The Challenge for Rapid Detection of High-Structured Circular RNA: Assay of Potato Spindle Tuber Viroid Based on Recombinase Polymerase Amplification and Lateral Flow Tests


 ,
,  and
and
Abstract
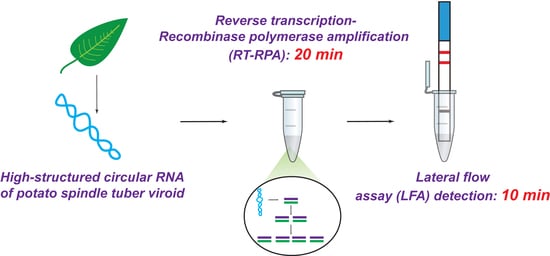
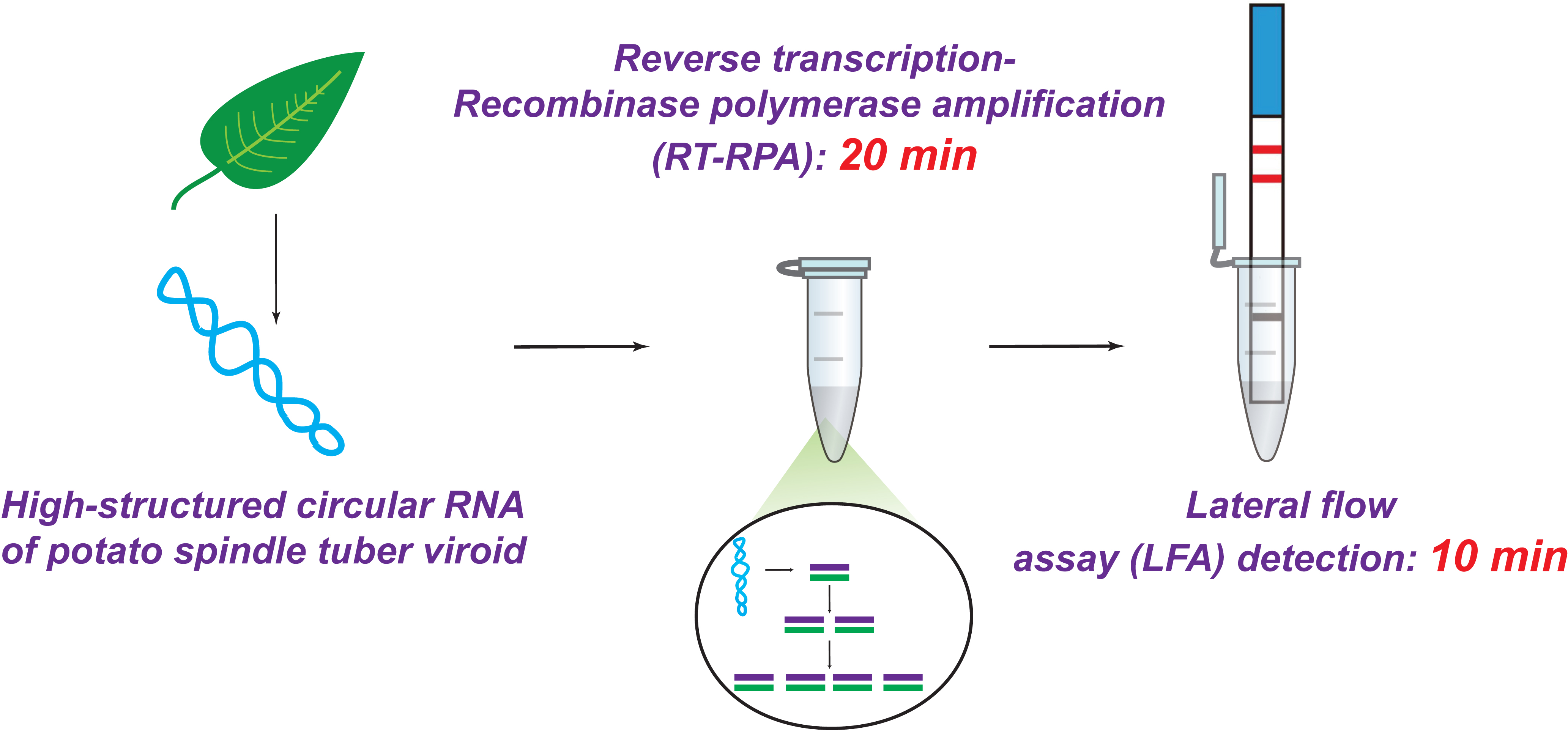
1. Introduction
2. Results and Discussion
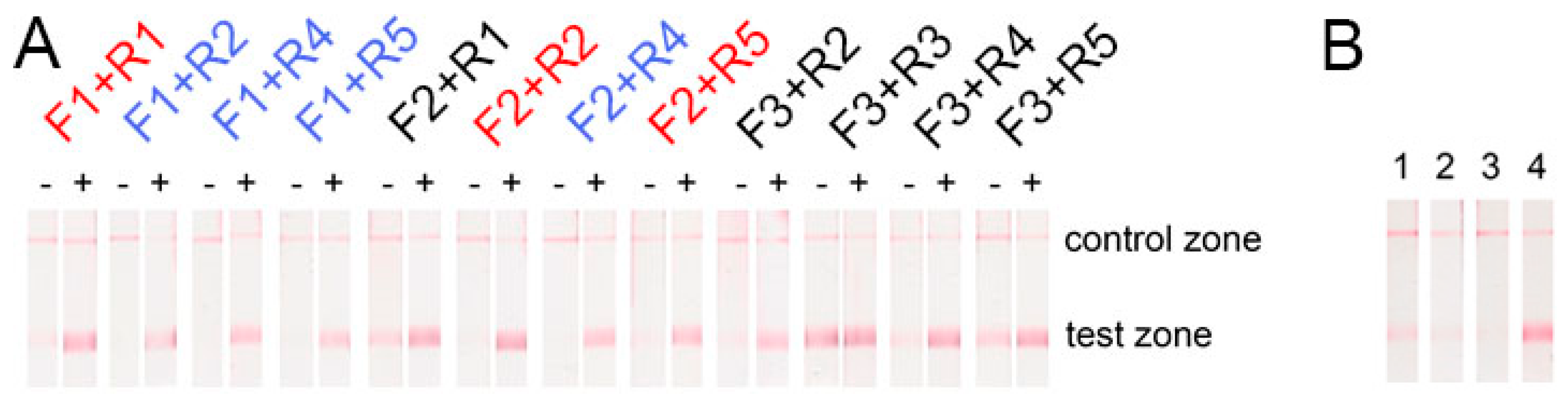
2.1. Selection of Primers for Recombinase Polymerase Amplification Lateral Flow Assay (RPA-LFA)
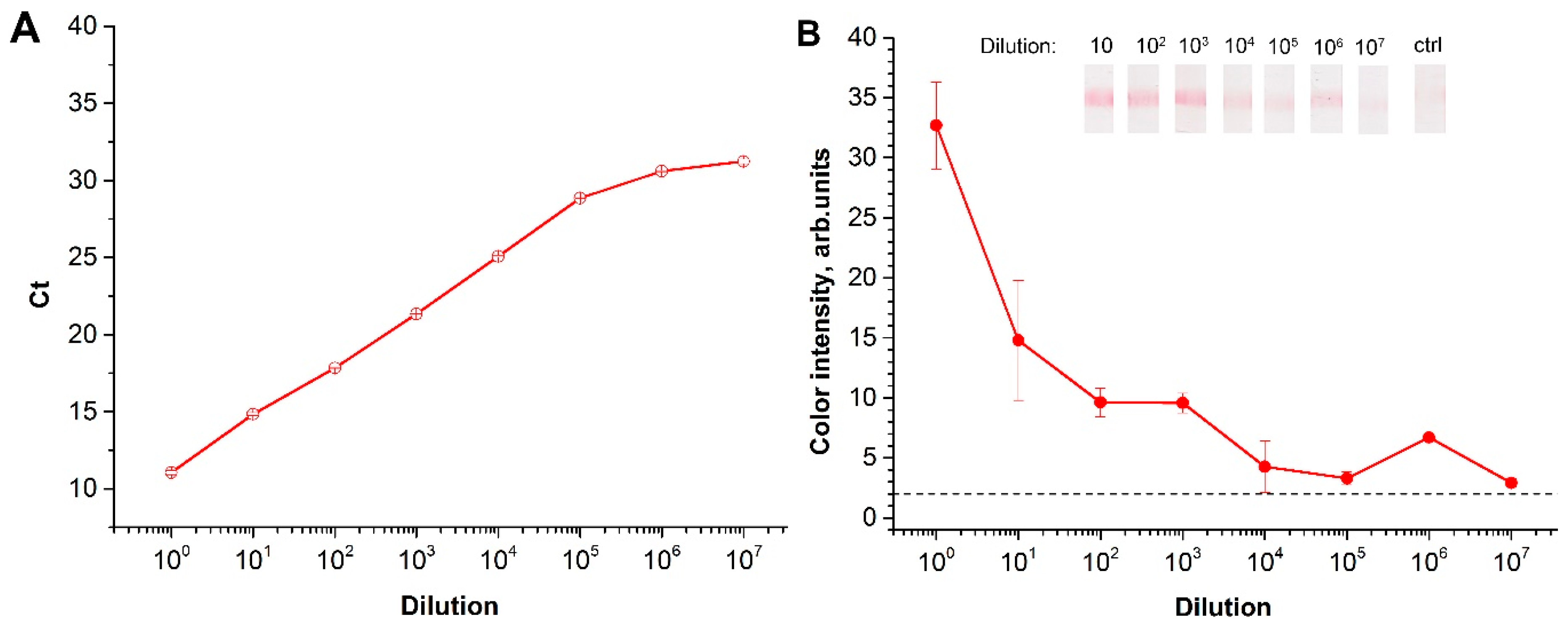
2.2. Characterization of Reverse Transcription (RT)-RPA-LFA
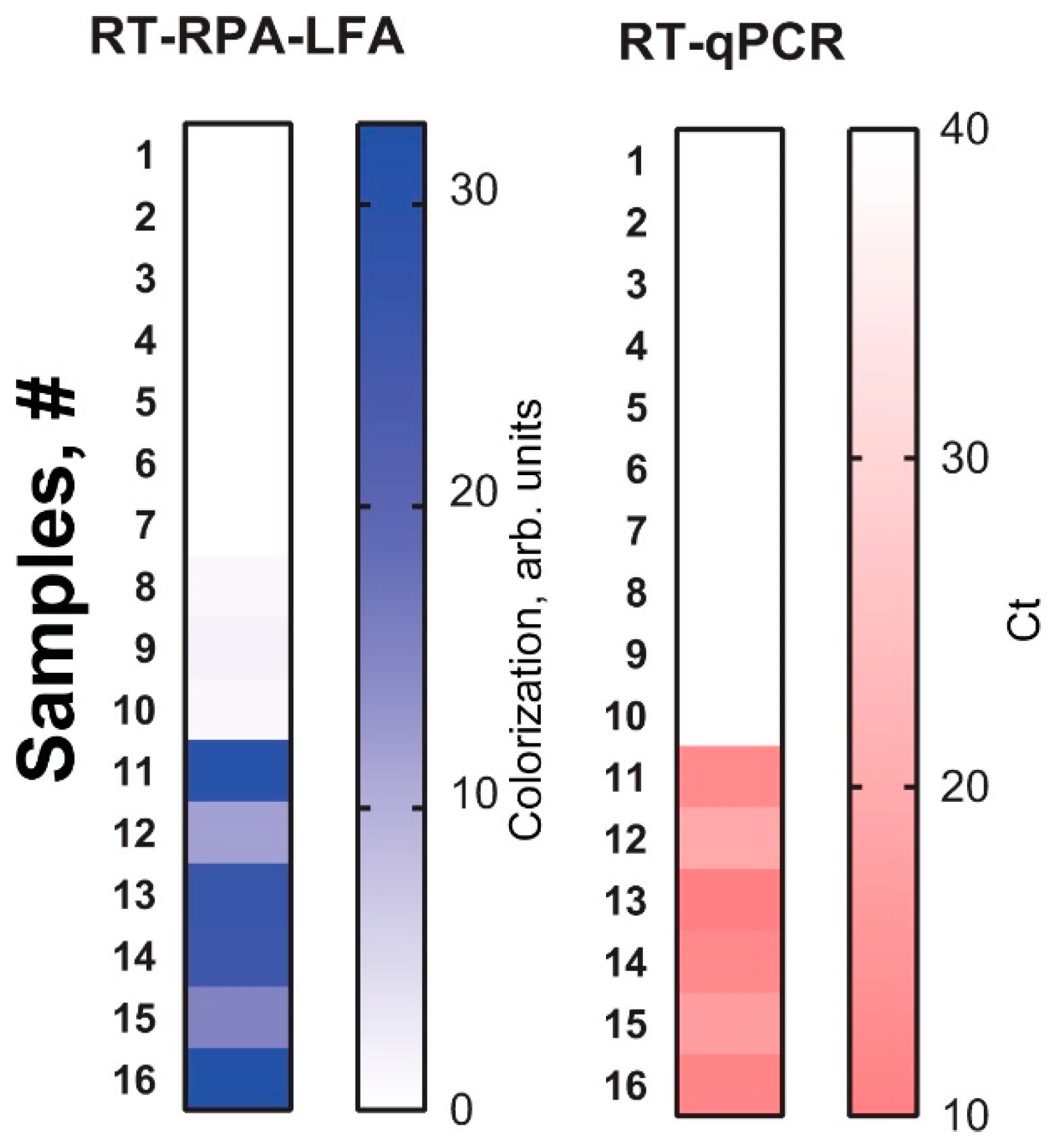
2.3. RT-RPA-LFA of Healthy and Potato Spindle Tuber Viroid (PSTVd)-Infected Potatoes
3. Materials and Methods
3.1. In Vitro Transcription of PSTVd RNA
3.2. Purification of PSTVd RNA
3.3. Preparation of Conjugate of Gold Nanoparticles with Antibodies
3.4. Preparation of LFA Test Strips
3.5. Sample Collection and Characterization
3.6. Primer Design for RPA
3.7. RPA with the LFA test
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hammond, R.W. Economic significance of viroids in vegetable and filed crops. In Viroids and Satellites; Hadidi, A., Flores, R., Palukaitis, P., Randles, J., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 5–14. [Google Scholar]
- Steger, G.; Perreault, J.P. Structure and Associated Biological Functions of Viroids. Adv. Virus Res. 2016, 94, 141–172. [Google Scholar] [CrossRef]
- Keese, P.; Symons, R.H. Domains in viroids: Evidence of intermolecular RNA rearrangements and their contribution to viroid evolution. Proc. Natl. Acad. Sci. USA 1985, 82, 4582–4586. [Google Scholar] [CrossRef]
- Gast, F.U.; Kempe, D.; Spieker, R.L.; Sanger, H.L. Secondary structure probing of potato spindle tuber viroid (PSTVd) and sequence comparison with other small pathogenic RNA replicons provides evidence for central non-canonical base-pairs, large A-rich loops, and a terminal branch. J. Mol. Biol. 1996, 262, 652–670. [Google Scholar] [CrossRef]
- Zhong, X.; Leontis, N.; Qian, S.; Itaya, A.; Qi, Y.; Boris-Lawrie, K.; Ding, B. Tertiary structural and functional analyses of a viroid RNA motif by isostericity matrix and mutagenesis reveal its essential role in replication. J. Virol. 2006, 80, 8566–8581. [Google Scholar] [CrossRef]
- Qi, Y.; Pelissier, T.; Itaya, A.; Hunt, E.; Wassenegger, M.; Ding, B. Direct role of a viroid RNA motif in mediating directional RNA trafficking across a specific cellular boundary. Plant Cell 2004, 16, 1741–1752. [Google Scholar] [CrossRef]
- Takeda, R.; Petrov, A.I.; Leontis, N.B.; Ding, B. A three-dimensional RNA motif in Potato spindle tuber viroid mediates trafficking from palisade mesophyll to spongy mesophyll in Nicotiana benthamiana. Plant Cell 2011, 23, 258–272. [Google Scholar] [CrossRef]
- Zhong, X.; Tao, X.; Stombaugh, J.; Leontis, N.; Ding, B. Tertiary structure and function of an RNA motif required for plant vascular entry to initiate systemic trafficking. EMBO J. 2007, 26, 3836–3846. [Google Scholar] [CrossRef]
- Adkar-Purushothama, C.R.; Perreault, J.P. Current overview on viroid-host interactions. Wiley Interdiscip. Rev. RNA 2020, 11, e1570. [Google Scholar] [CrossRef]
- Adkar-Purushothama, C.R.; Iyer, P.S.; Perreault, J.P. Potato spindle tuber viroid infection triggers degradation of chloride channel protein CLC-b-like and Ribosomal protein S3a-like mRNAs in tomato plants. Sci. Rep. 2017, 7, 8341. [Google Scholar] [CrossRef]
- Pelissier, T.; Thalmeir, S.; Kempe, D.; Sanger, H.L.; Wassenegger, M. Heavy de novo methylation at symmetrical and non-symmetrical sites is a hallmark of RNA-directed DNA methylation. Nucleic Acids Res. 1999, 27, 1625–1634. [Google Scholar] [CrossRef]
- Matsushita, Y.; Yanagisawa, H.; Sano, T. Vertical and Horizontal Transmission of Pospiviroids. Viruses 2018, 10, 706. [Google Scholar] [CrossRef] [PubMed]
- Mackie, A.E.; Barbetti, M.J.; Rodoni, B.; McKirdy, S.J.; Jones, R.A.C. Effects of a Potato Spindle Tuber Viroid Tomato Strain on the Symptoms, Biomass, and Yields of Classical Indicator and Currently Grown Potato and Tomato Cultivars. Plant Dis. 2019, 103, 3009–3017. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Jin, B.; Liu, W.; Chen, L.; Chen, J. Highly sensitive fluorescent-labeled probes and glass slide hybridization for the detection of plant RNA viruses and a viroid. Acta Biochim. Biophys. Sin. 2007, 39, 326–334. [Google Scholar] [CrossRef]
- Nakahara, K.; Hataya, T.; Hayashi, Y.; Sugimoto, T.; Kimura, I.; Shikata, E. A mixture of synthetic oligonucleotide probes labeled with biotin for the sensitive detection of potato spindle tuber viroid. J. Virol. Methods 1998, 71, 219–227. [Google Scholar] [CrossRef]
- Singh, R.P.; Boucher, A.; Lakshman, D.K.; Tavantzis, S.M. Multimeric non-radioactive cRNA probes improve detection of potato spindle tuber viroid (PSTVd). J. Virol. Methods 1994, 49, 221–233. [Google Scholar] [CrossRef]
- Vachev, T.; Ivanova, D.; Yahubyan, G.; Naimov, S.; Minkov, I.; Gozmanova, M. Detection of Potato spindle tuber viroid sequence variants derived from PSTVd-infected Phelipanche ramosa in flower organs of tomato plants. Biotechnol. Biotechnol. Equip. 2014, 28, 402–407. [Google Scholar] [CrossRef]
- Torchetti, E.M.; Navarro, B.; Di Serio, F. A single polyprobe for detecting simultaneously eight pospiviroids infecting ornamentals and vegetables. J. Virol. Methods 2012, 186, 141–146. [Google Scholar] [CrossRef]
- Peiman, M.; Xie, C. Development and evaluation of a multiplex RT-PCR for detecting main viruses and a viroid of potato. Acta Virol. 2006, 50, 129–133. [Google Scholar]
- Bostan, H.; Nie, X.; Singh, R.P. An RT-PCR primer pair for the detection of Pospiviroid and its application in surveying ornamental plants for viroids. J. Virol. Methods 2004, 116, 189–193. [Google Scholar] [CrossRef]
- Shamloul, A.M.; Faggioli, F.; Keith, J.M.; Hadidi, A. A novel multiplex RT-PCR probe capture hybridization (RT-PCR-ELISA) for simultaneous detection of six viroids in four genera: Apscaviroid, Hostuviroid, Pelamoviroid, and Pospiviroid. J. Virol. Methods 2002, 105, 115–121. [Google Scholar] [CrossRef]
- Nie, X.; Singh, R.P. A novel usage of random primers for multiplex RT-PCR detection of virus and viroid in aphids, leaves, and tubers. J. Virol. Methods 2001, 91, 37–49. [Google Scholar] [CrossRef]
- Boonham, N.; Perez, L.G.; Mendez, M.S.; Peralta, E.L.; Blockley, A.; Walsh, K.; Barker, I.; Mumford, R.A. Development of a real-time RT-PCR assay for the detection of potato spindle tuber viroid. J. Virol. Methods 2004, 116, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Botermans, M.; van de Vossenberg, B.T.; Verhoeven, J.T.; Roenhorst, J.W.; Hooftman, M.; Dekter, R.; Meekes, E.T. Development and validation of a real-time RT-PCR assay for generic detection of pospiviroids. J. Virol. Methods 2013, 187, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Monger, W.; Tomlinson, J.; Booonham, N.; Marn, M.V.; Plesko, I.M.; Molinero-Demilly, V.; Tassus, X.; Meekes, E.; Toonen, M.; Papayiannis, L.; et al. Development and inter-laboratory evaluation of real-time PCR assays for the detection of pospiviroids. J. Virol. Methods 2010, 169, 207–210. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. ISPM 27 Diagnostic Protocols 7: Potato Spindle Tuber Viroid; International Plant Protection Convention: Rome, Italy, 2015. [Google Scholar]
- European and Mediterranean Plant Protection Organization. Summary Sheet of Validation Data for a Diagnostic Test; International Plant Protection Convention: Rome, Italy, 2016; Volume 11-17239. [Google Scholar]
- Jeffries, C.; James, C. Development of an EU protocol for the detection and diagnosis of Potato spindle tuber pospiviroid. EPPO Bull. 2005, 35, 125–132. [Google Scholar] [CrossRef]
- Dobnik, D.; Morisset, D.; Lenarcic, R.; Ravnikar, M. Simultaneous detection of RNA and DNA targets based on multiplex isothermal amplification. J. Agric. Food Chem. 2014, 62, 2989–2996. [Google Scholar] [CrossRef]
- Tsutsumi, N.; Yanagisawa, H.; Fujiwara, Y.; Ohara, T. Detection of Potato Spindle Tuber Viroid by Reverse Transcription Loop-mediated Isothermal Amplification. Res. Bull. Plant Prot. Serv. Jpn. 2010, 46, 7. [Google Scholar]
- Lenarcic, R.; Morisset, D.; Mehle, N.; Ravnikar, M. Fast real-time detection of Potato spindle tuber viroid by RT-LAMP. Plant Pathol. 2013, 62, 1147–1156. [Google Scholar] [CrossRef]
- Fischbach, J.; Xander, N.C.; Frohme, M.; Glokler, J.F. Shining a light on LAMP assays--a comparison of LAMP visualization methods including the novel use of berberine. Biotechniques 2015, 58, 189–194. [Google Scholar] [CrossRef]
- Owens, R.A.; Sano, T.; Duran-Vila, N. Plant viroids: Isolation, characterization/detection, and analysis. Methods Mol. Biol. 2012, 894, 253–271. [Google Scholar] [CrossRef]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA detection using recombination proteins. PLoS Biol. 2006, 4, e204. [Google Scholar] [CrossRef] [PubMed]
- Hammond, R.W.; Zhang, S. Development of a rapid diagnostic assay for the detection of tomato chlorotic dwarf viroid based on isothermal reverse-transcription-recombinase polymerase amplification. J. Virol. Methods 2016, 236, 62–67. [Google Scholar] [CrossRef]
- Safenkova, I.V.; Ivanov, A.V.; Slutskaya, E.S.; Samokhvalov, A.V.; Zherdev, A.V.; Dzantiev, B.B. Key significance of DNA-target size in lateral flow assay coupled with recombinase polymerase amplification. Anal. Chim. Acta 2020, 1102, 109–118. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W.; Sambrook, J. The Condensed Protocols from Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2006; 800p. [Google Scholar]
- Byzova, N.A.; Safenkova, I.V.; Chirkov, S.N.; Zherdev, A.V.; Blintsov, A.N.; Dzantiev, B.B.; Atabekov, I.G. Development of immunochromatographic test systems for express detection of plant viruses. Appl. Biochem. Microbiol. 2009, 45, 204–209. [Google Scholar] [CrossRef]
- Kibbe, W.A. OligoCalc: An online oligonucleotide properties calculator. Nucleic Acids Res. 2007, 35, W43–W46. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Safenkova, I.V.; Zherdev, A.V.; Dzantiev, B.B. Nucleic acid lateral flow assay with recombinase polymerase amplification: Solutions for highly sensitive detection of RNA virus. Talanta 2020, 210, 120616. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | 5′-3′ Sequence | Position at RNA of Potato Spindle Tuber Viroid (PSTVd) | Modification |
|---|---|---|---|
| PSTV F1 1 | ggttcacacctgacctcctgagcagaaaag | 25–54 | FAM |
| PSTV F2 | cggggaaacctggagcgaactggcaat | 94–120 | FAM |
| PSTV F3 | gggagtgcccagcggccgacaggagtaatt | 131–160 | FAM |
| PSTV R1 | accctcgccccgaagcaagtaagatag | 301–327 | Biotin |
| PSTV R2 | accgggtagtagccgaagcgacagcgc | 239–265 | Biotin |
| PSTV R3 | caccctcgccccgaagcaagtaagatagag | 299–328 | Biotin |
| PSTV R4 | aaaaagcggttctcgggagcttcagttgtt | 269–298 | Biotin |
| PSTV R5 | ggagcttcagttgtttccaccgggtagtag | 254–283 | Biotin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanov, A.V.; Shmyglya, I.V.; Zherdev, A.V.; Dzantiev, B.B.; Safenkova, I.V. The Challenge for Rapid Detection of High-Structured Circular RNA: Assay of Potato Spindle Tuber Viroid Based on Recombinase Polymerase Amplification and Lateral Flow Tests. Plants 2020, 9, 1369. https://doi.org/10.3390/plants9101369
Ivanov AV, Shmyglya IV, Zherdev AV, Dzantiev BB, Safenkova IV. The Challenge for Rapid Detection of High-Structured Circular RNA: Assay of Potato Spindle Tuber Viroid Based on Recombinase Polymerase Amplification and Lateral Flow Tests. Plants. 2020; 9(10):1369. https://doi.org/10.3390/plants9101369
Chicago/Turabian StyleIvanov, Aleksandr V., Irina V. Shmyglya, Anatoly V. Zherdev, Boris B. Dzantiev, and Irina V. Safenkova. 2020. "The Challenge for Rapid Detection of High-Structured Circular RNA: Assay of Potato Spindle Tuber Viroid Based on Recombinase Polymerase Amplification and Lateral Flow Tests" Plants 9, no. 10: 1369. https://doi.org/10.3390/plants9101369
APA StyleIvanov, A. V., Shmyglya, I. V., Zherdev, A. V., Dzantiev, B. B., & Safenkova, I. V. (2020). The Challenge for Rapid Detection of High-Structured Circular RNA: Assay of Potato Spindle Tuber Viroid Based on Recombinase Polymerase Amplification and Lateral Flow Tests. Plants, 9(10), 1369. https://doi.org/10.3390/plants9101369