
Structural Variations of Broccoli Polyphenolics and Their Antioxidant Capacity as a Function of Growing Temperature



Abstract
1. Introduction
2. Results and Discussion
2.1. The Effect of High and Low Growth Temperature on Free and Bound Total Phenolics in Broccoli
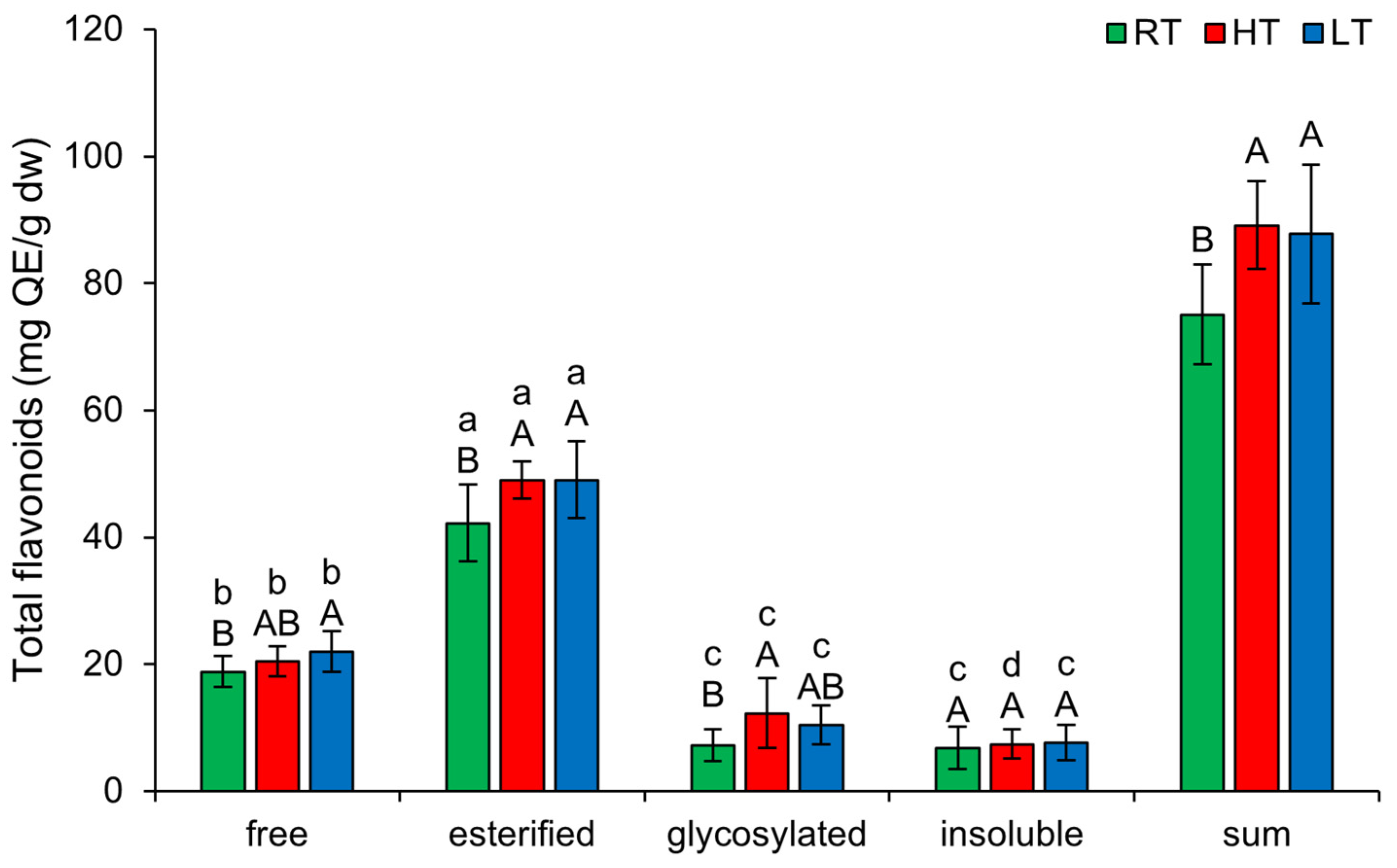
2.2. The Effect of High and Low Growth Temperature on Free and Bound Total Flavonoids in Broccoli
2.3. The Effect of High and Low Growth Temperature on Free and Bound Total Tannins in Broccoli
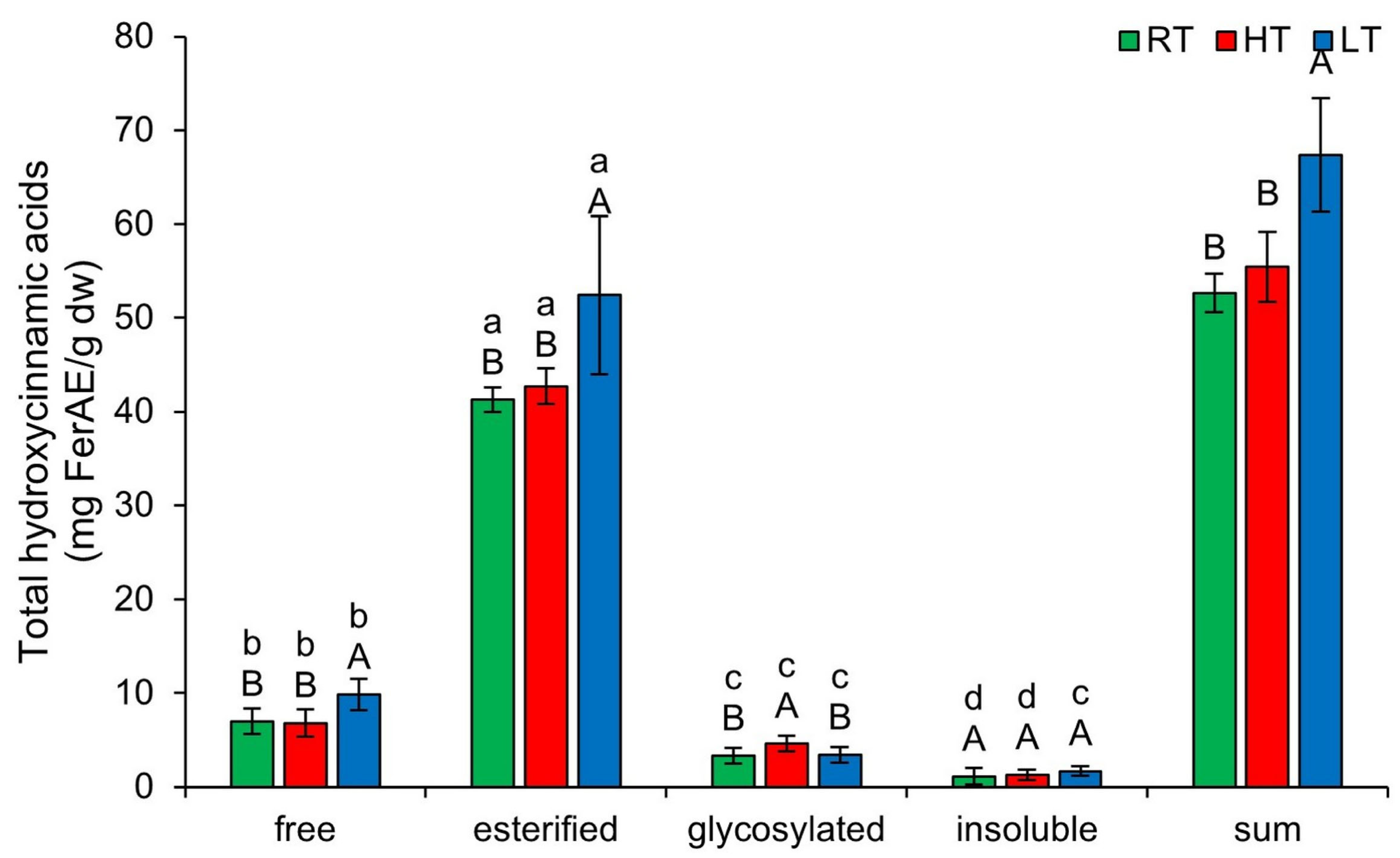
2.4. The Effect of High and Low Growth Temperature on Free and Bound Total Hydroxycinnamic Acids in Broccoli
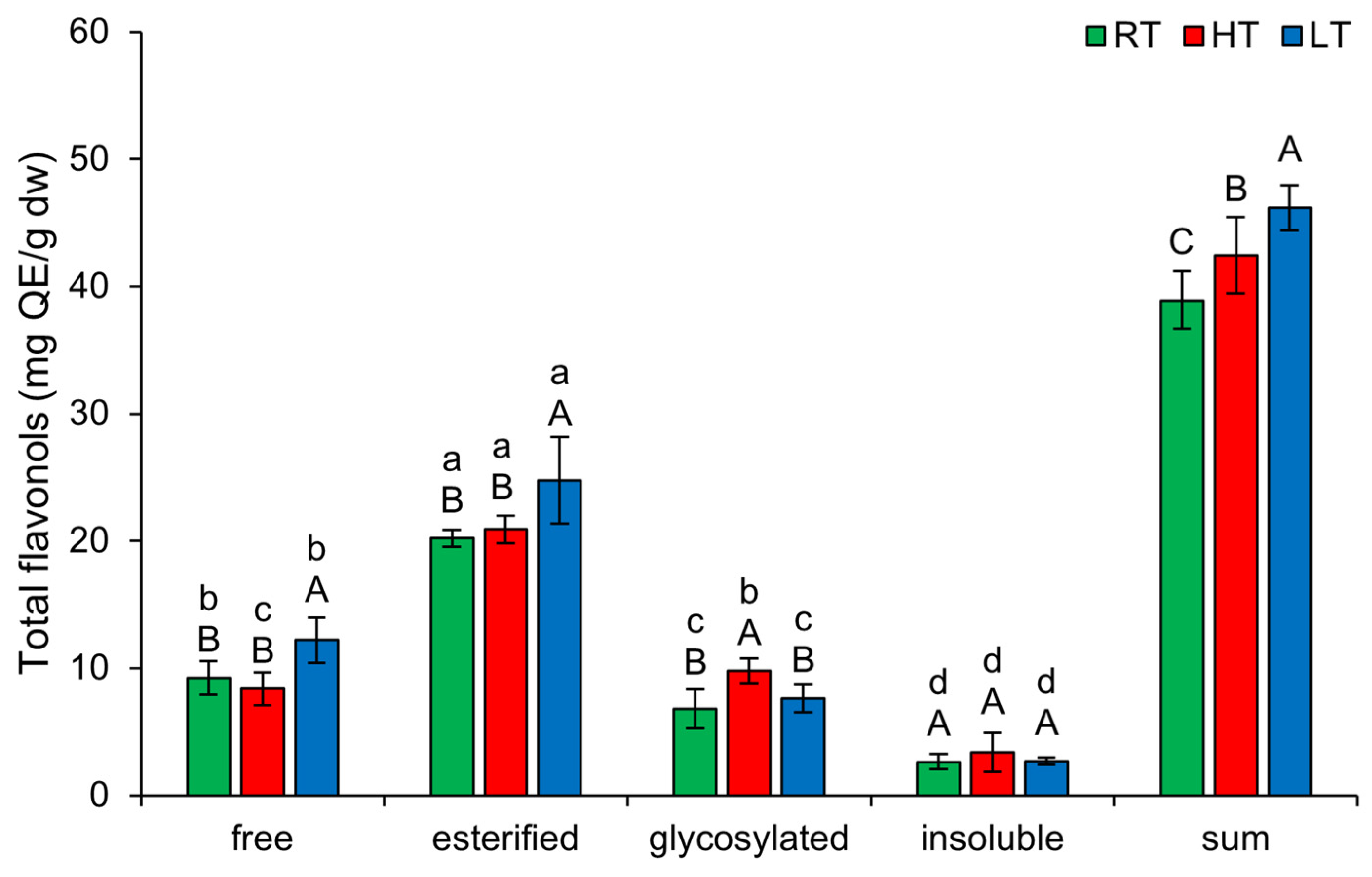
2.5. The Effect of High and Low Growth Temperature on Free and Bound Total Flavonols in Broccoli
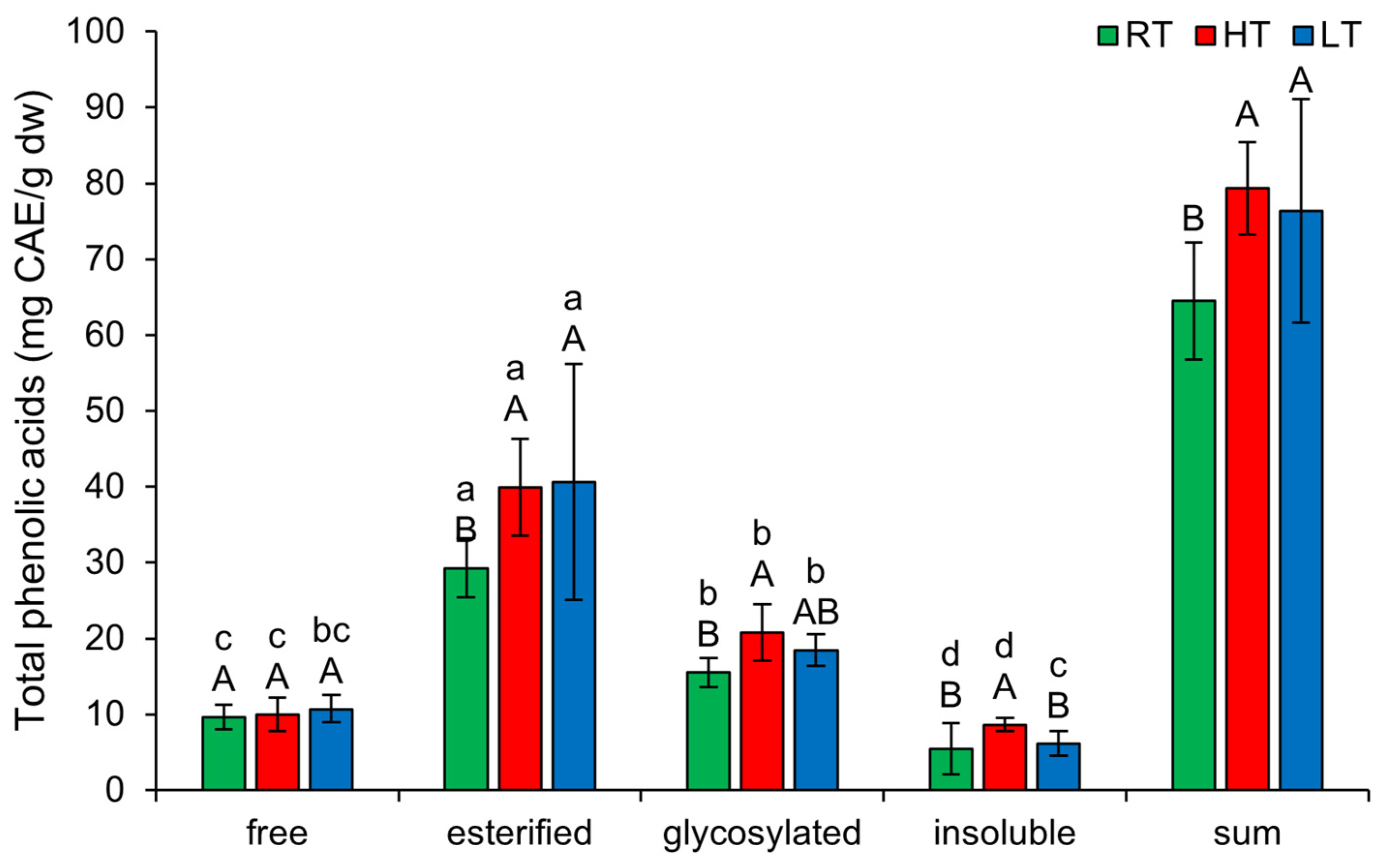
2.6. The Effect of High and Low Growth Temperature on Free and Bound Total Phenolic Acids in Broccoli
2.7. The Effect of High and Low Growth Temperature on Free and Bound Individual Polyphenolics in Broccoli
2.8. The Effect of High and Low Growth Temperature on Antioxidant Capacity of Free and Bound Polyphenolics in Broccoli
2.9. Statistical Analyses
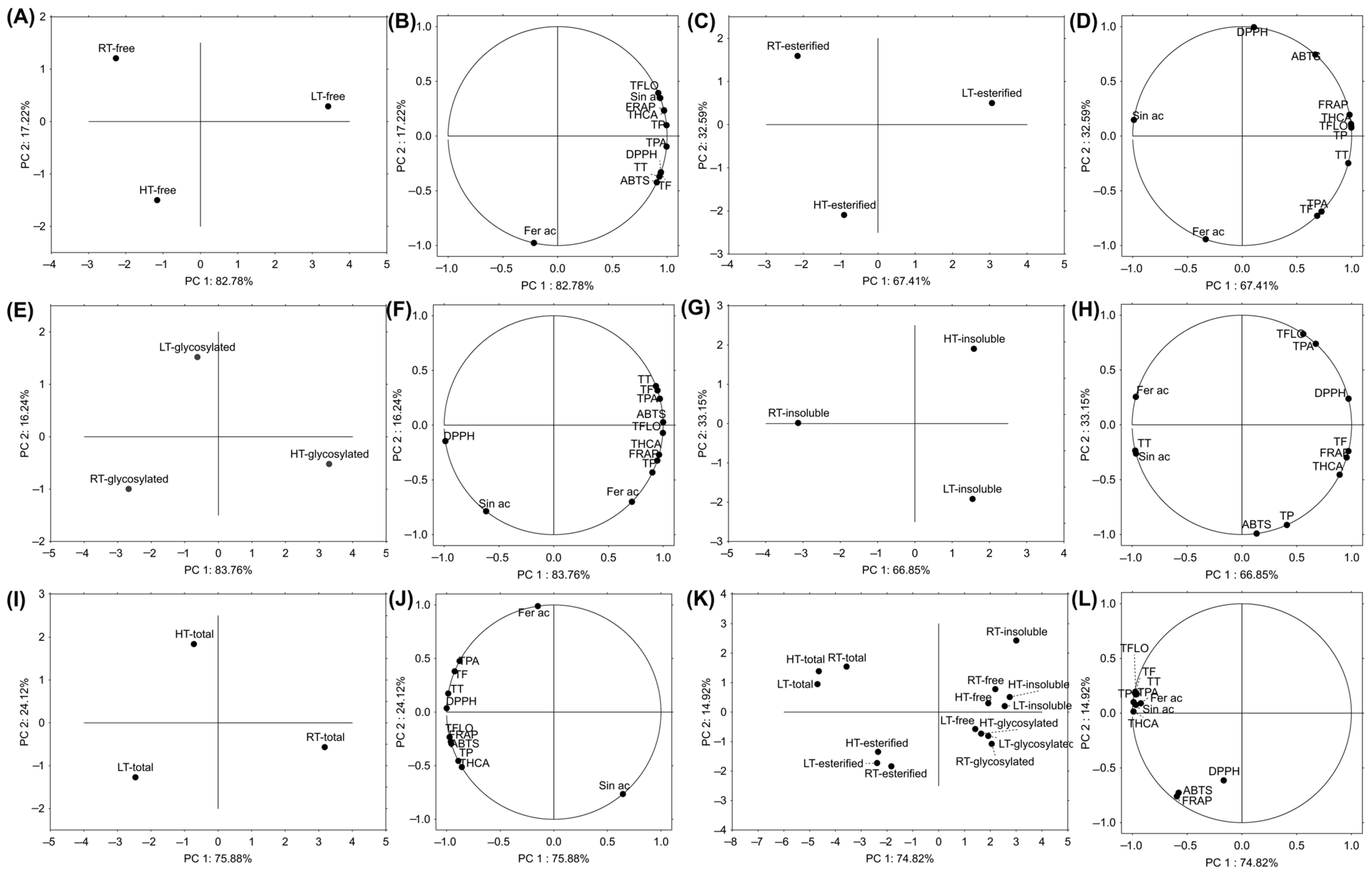
2.9.1. Pearson’s Correlations
2.9.2. Principal Component Analysis
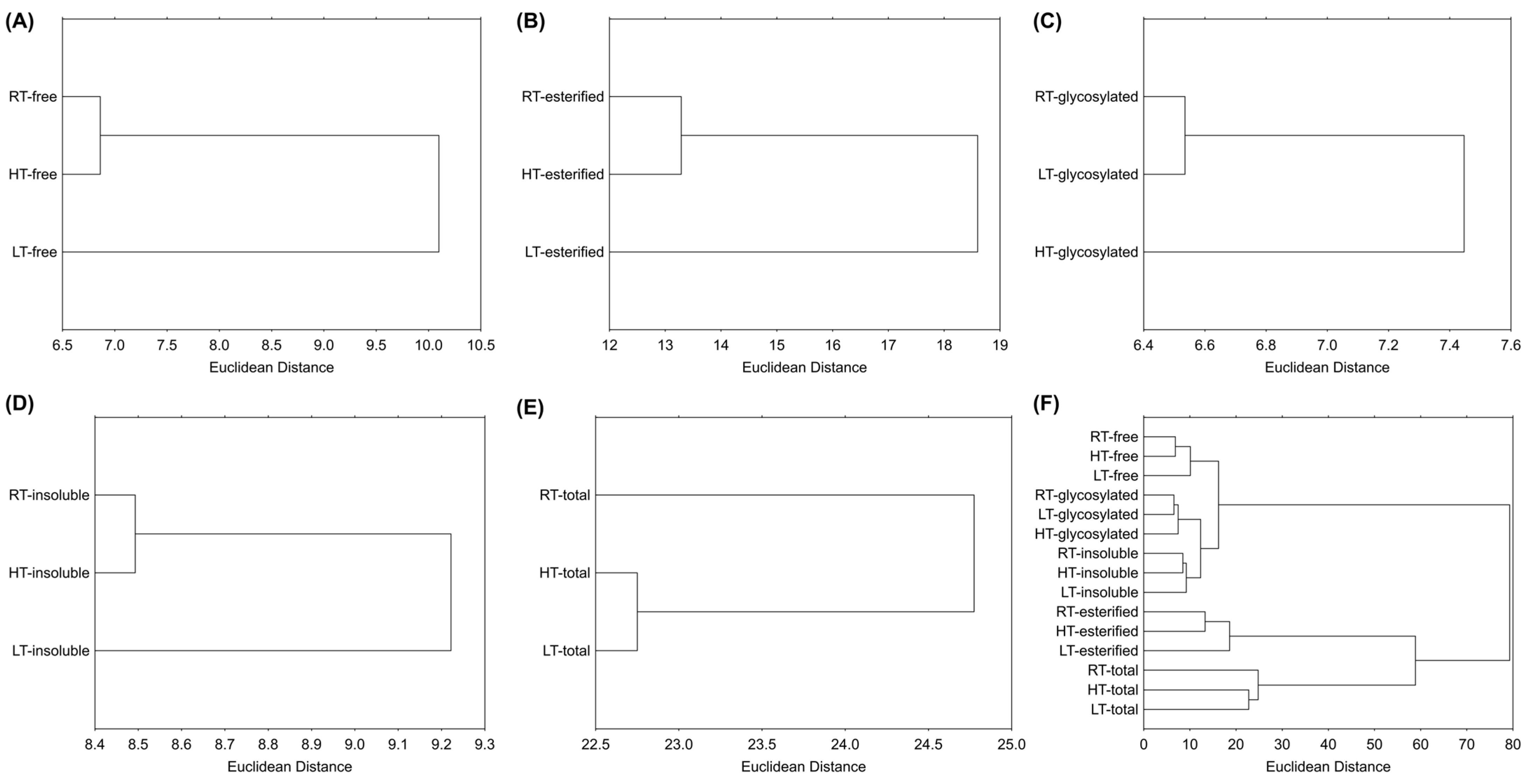
2.9.3. Hierarchical Clustering
3. Materials and Methods
3.1. Plant Material
3.2. Extraction of Different Polyphenolic Structural Forms
3.3. Spectrophotometric Analyses of Different Groups of Polyphenolics
3.4. High-Performance Liquid Chromatography Analysis of Individual Polyphenolic Compounds
3.5. Determination of Antioxidant Capacity of Different Polyphenolic Structural Forms
3.6. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABTS | 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) |
| CAE | caffeic acid equivalents |
| DPPH | 2,2-diphenyl-1-picrylhydrazyl |
| Fer ac | ferulic acid |
| FRAP | ferric ion reducing antioxidant power |
| GAE | gallic acid equivalents |
| HC | hierarchical clustering |
| HT | high temperature |
| LT | low temperature |
| QE | quercetin equivalents |
| PCA | principal component analysis |
| RT | room temperature |
| Sin ac | sinapic acid |
| TF | total flavonoids |
| TFLO | total flavonols |
| THCA | total hydroxycinnamic acids |
| TP | total phenolics |
| TPA | total phenolic acids |
| TT | total tannins |
References
- Nardini, M.; Ghiselli, A. Determination of free and bound phenolic acids in beer. Food Chem. 2004, 84, 137–143. [Google Scholar]
- Zhang, B.; Zhang, Y.; Li, H.; Deng, Z.; Tsao, R. A review on insoluble-bound phenolics in plant-based food matrix and their contribution to human health with future perspectives. Trends Food Sci. Technol. 2020, 105, 347–362. [Google Scholar]
- Lou, X.M.; Xu, H.D.; Hanna, M.; Yuan, L. Identification and quantification of free, esterified, glycosylated and insoluble-bound phenolic compounds in hawthorn berry fruit (Crataegus pinnatifida) and antioxidant activity evaluation. LWT-Food Sci. Technol. 2020, 130, 10. [Google Scholar]
- Shahidi, F.; Hossain, A. Importance of insoluble-bound phenolics to the antioxidant potential is dictated by source material. Antioxidants 2023, 12, 203. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Arruda, H.S.; Pereira, G.A.; de Morais, D.R.; Eberlin, M.N.; Pastore, G.M. Determination of free, esterified, glycosylated and insoluble-bound phenolics composition in the edible part of araticum fruit (Annona crassiflora Mart.) and its by-products by HPLC-ESI-MS/MS. Food Chem. 2018, 245, 738–749. [Google Scholar]
- Šola, I.; Stić, P.; Rusak, G. Effect of flooding and drought on the content of phenolics, sugars, photosynthetic pigments and vitamin C, and antioxidant potential of young Chinese cabbage. Eur. Food Res. Technol. 2021, 247, 1913–1920. [Google Scholar]
- Šola, I.; Poljuha, D.; Mikulic-Petkovsek, M.; Davosir, D.; Pinterić, M.; Bilić, J.; Veberic, R.; Hudina, M.; Rusak, G. Biopotential of underutilized Rosaceae inflorescences: LC-DAD-MS phytochemical profiles associated with antioxidant, antidiabetic, anti-inflammatory and antiproliferative activity in vitro. Plants 2022, 11, 271. [Google Scholar] [CrossRef]
- Rocchetti, G.; Gregorio, R.P.; Lorenzo, J.M.; Barba, F.J.; Oliveira, P.G.; Prieto, M.A.; Simal-Gandara, J.; Mosele, J.I.; Motilva, M.J.; Tomas, M.; et al. Functional implications of bound phenolic compounds and phenolics-food interaction: A review. Compr. Rev. Food Sci. Food Saf. 2022, 21, 811–842. [Google Scholar]
- Harbaum, B.M.; Hubbermann, E.M.; Zhu, Z.; Schwarz, K. Free and bound phenolic compounds in leaves of pak choi (Brassica campestris L. ssp. chinensis var. communis) and Chinese leaf mustard (Brassica juncea Coss). Food Chem. 2008, 110, 838–846. [Google Scholar]
- Kang, O.J. Distribution of free, esterified, and insoluble bound forms of phenolics in tea seeds and their antioxidant activity. Food Sci. Biotechnol. 2017, 26, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Shen, D.; Wang, R.; Li, Q.; Mo, R.; Zheng, Y.; Zhou, Y.; Liu, Y. Phenolic profiles and antioxidant activities of free, esterified and bound phenolic compounds in walnut kernel. Food Chem. 2021, 350, 129217. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, X.; Guo, Z.; Feng, X.; Huang, P.; Du, M.; Zalán, Z.; Kan, J. Distribution and natural variation of free, esterified, glycosylated, and insoluble-bound phenolic compounds in brocade orange (Citrus sinensis L. Osbeck) peel. Food Res. Int. 2022, 153, 110958. [Google Scholar] [CrossRef] [PubMed]
- Divya Priya, A.; Martin, A. UHPLC-MS/MS based comprehensive phenolic profiling, antimicrobial and antioxidant activities of Indian Rhodomyrtus tomentosa fruits. Sci. Rep. 2025, 15, 945. [Google Scholar]
- Acosta-Estrada, B.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Sun, J.; Chu, Y.-F.; Wu, X.; Liu, R.H. Antioxidant and antiproliferative activities of common fruits. J. Agric. Food Chem. 2002, 50, 7449–7454. [Google Scholar] [CrossRef]
- Adom, K.K.; Liu, R.H. Antioxidant activity of grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [Google Scholar] [CrossRef]
- Ross, K.A.; Beta, T.; Arntfield, S.D. A comparative study on the phenolic acids identified and quantified in dry beans using HPLC as affected by different extraction and hydrolysis methods. Food Chem. 2009, 113, 336–344. [Google Scholar] [CrossRef]
- Rocchetti, G.; Chiodelli, G.; Giuberti, G.; Lucini, L. Bioaccessibility of phenolic compounds following in vitro large intestine fermentation of nuts for human consumption. Food Chem. 2018, 245, 633–640. [Google Scholar] [CrossRef]
- Gong, E.S.; Luo, S.; Li, T.; Liu, C.; Zhang, G.; Chen, J.; Zeng, Z.; Liu, R.H. Phytochemical profiles and antioxidant activity of processed brown rice products. Food Chem. 2017, 232, 67–78. [Google Scholar] [CrossRef]
- Chen, P.X.; Dupuis, J.H.; Marcone, M.F.; Pauls, P.K.; Liu, R.; Liu, Q.; Tang, Y.; Zhang, B.; Tsao, R. Physicochemical properties and in vitro digestibility of cooked regular and nondarkening cranberry beans (Phaseolus vulgaris L.) and their effects on bioaccessibility, phenolic composition, and antioxidant activity. J. Agric. Food Chem. 2015, 63, 10448–10458. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, V.A. Effects of different cooking and drying methods on phenolic acids, carotenoids, and antioxidant activity of emmer (Triticum turgidum ssp. dicoccum) bulgur. Cereal Chem. 2019, 96, 1093–1102. [Google Scholar] [CrossRef]
- Martini, D.; Ciccoritti, R.; Nicoletti, I.; Nocente, F.; Corradini, D.; D’Egidio, M.G.; Taddei, F. From seed to cooked pasta: Influence of traditional and non-conventional transformation processes on total antioxidant capacity and phenolic acid content. Int. J. Food Sci. Nutr. 2018, 69, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, X.; Deng, J.; Ouyang, D.; Wang, D.; Liang, Y.; Chen, Y.; Sun, Y. Effect of thermal processing on free and bound phenolic compounds and antioxidant activities of hawthorn. Food Chem. 2020, 332, 127429. [Google Scholar] [CrossRef]
- Yeo, J.; Shahidi, F. Critical evaluation of changes in the ratio of insoluble bound to soluble phenolics on antioxidant activity of lentils during germination. J. Agric. Food Chem. 2015, 63, 379–381. [Google Scholar] [CrossRef]
- de Camargo, A.C.; Speisky, H.; Bridi, R.; Núñez Pizarro, P.; Larena, A.; Pinaffi-Langley, A.C.d.C.; Shahidi, F.; Schwember, A.R. Chickpeas from a Chilean region affected by a climate-related catastrophe: Effects of water stress on grain yield and flavonoid composition. Molecules 2022, 27, 691. [Google Scholar] [CrossRef]
- Martini, D.; Taddei, F.; Ciccoritti, R.; Pasquini, M.; Nicoletti, I.; Corradini, D.; D’Egidio, M.G. Variation of total antioxidant activity and of phenolic acid, total phenolics and yellow coloured pigments in durum wheat (Triticum turgidum L. var. durum) as a function of genotype, crop year and growing area. J. Cereal Sci. 2015, 65, 175–185. [Google Scholar]
- Antognoni, F.; Potente, G.; Biondi, S.; Mandrioli, R.; Marincich, L.; Ruiz, K.B. Free and conjugated phenolic profiles and antioxidant activity in quinoa seeds and their relationship with genotype and environment. Plants 2021, 10, 1046. [Google Scholar] [CrossRef]
- Gmižić, D.; Pinterić, M.; Lazarus, M.; Šola, I. High growing temperature changes nutritional value of broccoli (Brassica oleracea L. convar. botrytis (L.) Alef. var. cymosa Duch.) seedlings. Foods 2023, 12, 582. [Google Scholar]
- Šola, I.; Davosir, D.; Kokić, E.; Zekirovski, J. Effect of hot- and cold-water treatment on broccoli bioactive compounds, oxidative stress parameters and biological effects of their extracts. Plants 2023, 12, 1135. [Google Scholar] [CrossRef]
- Šola, I.; Gmižić, D.; Pinterić, M.; Tot, A.; Ludwig-Müller, J. Adjustments of the phytochemical profile of broccoli to low and high growing temperatures: Implications for the bioactivity of its extracts. Int. J. Mol. Sci. 2024, 25, 3677. [Google Scholar] [CrossRef] [PubMed]
- Šola, I.; Vujčić Bok, V.; Popović, M.; Gagić, S. Phytochemical composition and functional properties of Brassicaceae microgreens: Impact of in vitro digestion. Int. J. Mol. Sci. 2024, 25, 11831. [Google Scholar] [CrossRef] [PubMed]
- Šola, I.; Gmižić, D.; Miškec, K.; Ludwig-Müller, J. Impact of water stress on metabolic intermediates and regulators in broccoli sprouts, and cellular defense potential of their extracts. Int. J. Mol. Sci. 2025, 26, 632. [Google Scholar] [CrossRef] [PubMed]
- Šola, I.; Poljuha, D.; Pavičić, I.; Jurinjak Tušek, A.; Šamec, D. Climate change and plant foods: The influence of environmental stressors on plant metabolites and future food sources. Foods 2025, 14, 416. [Google Scholar] [CrossRef]
- Mishra, S.; Spaccarotella, K.; Gido, J.; Samanta, I.; Chowdhary, G. Effects of heat stress on plant-nutrient relations: An update on nutrient uptake, transport, and assimilation. Int. J. Mol. Sci. 2023, 24, 15670. [Google Scholar] [CrossRef]
- Ruelland, E.; Vaultier, M.N.; Zachowski, A.; Hurry, V. Cold signalling and cold acclimation in plants. In Advances in Botanical Research; Kader, J.C., Delseney, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2009; Volume 49, pp. 35–150. [Google Scholar]
- Dias, M.C.; Pinto, D.; Silva, A.M.S. Plant flavonoids: Chemical characteristics and biological activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Lund, M.N. Reactions of plant polyphenols in foods: Impact of molecular structure. Trends Food Sci. Technol. 2021, 112, 241–251. [Google Scholar] [CrossRef]
- Shahidi, F.; Yeo, J.D. Insoluble-bound phenolics in food. Molecules 2016, 21, 1216. [Google Scholar] [CrossRef]
- Chandrasekara, A.; Shahidi, F. Bioactivities and antiradical properties of millet grains and hulls. J. Agric. Food Chem. 2011, 59, 9563–9571. [Google Scholar] [CrossRef]
- Sahraeian, S.; Rashidinejad, A.; Golmakani, M.-T. Recent advances in the conjugation approaches for enhancing the bioavailability of polyphenols. Food Hydrocoll. 2024, 146, 109221. [Google Scholar] [CrossRef]
- Andreu, A.; Ćorović, M.; Garcia-Sanz, C.; Santos, A.S.; Milivojević, A.; Ortega-Nieto, C.; Mateo, C.; Bezbradica, D.; Palomo, J.M. Enzymatic glycosylation strategies in the production of bioactive compounds. Catalysts 2023, 13, 1359. [Google Scholar] [CrossRef]
- Kytidou, K.; Artola, M.; Overkleeft, H.S.; Aerts, J.M.F.G. Plant glycosides and glycosidases: A treasure-trove for therapeutics. Front. Plant Sci. 2020, 11, 357. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef] [PubMed]
- Chaaban, H.; Ioannou, I.; Chebil, L.; Slimane, M.; Gérardin, C.; Paris, C.; Charbonnel, C.; Chekir, L.; Ghoul, M. Effect of heat processing on thermal stability and antioxidant activity of six flavonoids. J. Food Process Preserv. 2017, 41, e13203. [Google Scholar] [CrossRef]
- Haslam, E. Natural polyphenols (vegetable tannins) as drugs: Possible modes of action. J. Nat. Prod. 1996, 59, 205–215. [Google Scholar] [CrossRef]
- Lees, G.L.; Hinks, C.F.; Suttill, N.H. Effect of high temperature on condensed tannin accumulation in leaf tissues of big trefoil (Lotus uliginosus Schkuhr). J. Sci. Food Agric. 1994, 65, 415–421. [Google Scholar] [CrossRef]
- Top, S.M.; Preston, C.M.; Dukes, J.S.; Tharayil, N. Climate Influences the content and chemical composition of foliar tannins in green and senesced tissues of Quercus rubra. Front. Plant Sci. 2017, 8, 423. [Google Scholar] [CrossRef]
- Poudel, P.R.; Koyama, K.; Goto-Yamamoto, N. Evaluating the influence of temperature on proanthocyanidin biosynthesis in developing grape berries (Vitis vinifera L.). Mol. Biol. Rep. 2020, 47, 3501–3510. [Google Scholar] [CrossRef]
- Terletskaya, N.V.; Shadenova, E.A.; Litvinenko, Y.A.; Ashimuly, K.; Erbay, M.; Mamirova, A.; Nazarova, I.; Meduntseva, N.D.; Kudrina, N.O.; Korbozova, N.K.; et al. Influence of cold stress on physiological and phytochemical characteristics and secondary metabolite accumulation in microclones of Juglans regia L. Int. J. Mol. Sci. 2024, 25, 4991. [Google Scholar] [CrossRef]
- Khawula, S.; Gokul, A.; Niekerk, L.A.; Basson, G.; Keyster, M.; Badiwe, M.; Klein, A.; Nkomo, M. Insights into the effects of hydroxycinnamic acid and its secondary metabolites as antioxidants for oxidative stress and plant growth under environmental stresses. Curr. Issues Mol. Biol. 2023, 46, 81–95. [Google Scholar] [CrossRef]
- Chandrakanth, N.N.; Zhang, C.; Freeman, J.; de Souza, W.R.; Bartley, L.E.; Mitchell, R.A.C. Modification of plant cell walls with hydroxycinnamic acids by BAHD acyltransferases. Front. Plant Sci. 2023, 13, 1088879. [Google Scholar]
- Ma, Y.; Zhao, S.; Ma, X.; Dong, G.; Liu, C.; Ding, Y.; Hou, B. A high temperature responsive UDP-glucosyltransferase gene OsUGT72F1 enhances heat tolerance in rice and Arabidopsis. Plant Cell Rep. 2025, 44, 48. [Google Scholar] [PubMed]
- Behr, M.; Neutelings, G.; El Jaziri, M.; Baucher, M. You want it sweeter: How glycosylation affects plant response to oxidative stress. Front. Plant Sci. 2020, 11, 1443. [Google Scholar]
- Attri, S.; Sharma, K.; Raigond, P.; Goel, G. Colonic fermentation of polyphenolics from sea buckthorn (Hippophae rhamnoides) berries: Assessment of effects on microbial diversity by principal component analysis. Food Res. Int. 2018, 105, 324–332. [Google Scholar]
- Beveridge, T.; Loubert, E.; Harrison, J.E. Simple measurement of phenolic esters in plant cell walls. Food Res. Int. 2000, 33, 775–783. [Google Scholar]
- Lafay, S.; Gil-Izquierdo, A. Bioavailability of phenolic acids. Phytochem. Rev. 2007, 7, 301. [Google Scholar]
- Jones, P.; Vogt, T. Glycosyltransferases in secondary plant metabolism: Tranquilizers and stimulant controllers. Planta 2001, 213, 164–174. [Google Scholar]
- Le Roy, J.; Huss, B.; Creach, A.; Hawkins, S.; Neutelings, G. Glycosylation is a major regulator of phenylpropanoid availability and biological activity in plants. Front. Plant Sci. 2016, 7, 735. [Google Scholar]
- Osete-Alcaraz, A.; Bautista-Ortín, A.B.; Gómez-Plaza, E. The role of soluble polysaccharides in tannin-cell wall interactions in model solutions and in wines. Biomolecules 2019, 10, 36. [Google Scholar] [CrossRef]
- Su, X.; Wang, W.; Xia, T.; Gao, L.; Shen, G.; Pang, Y. Characterization of a heat responsive UDP: Flavonoid glucosyltransferase gene in tea plant (Camellia sinensis). PLoS ONE 2018, 13, e0207212. [Google Scholar]
- Zagoskina, N.V.; Zubova, M.Y.; Nechaeva, T.L.; Kazantseva, V.V.; Goncharuk, E.A.; Katanskaya, V.M.; Baranova, E.N.; Aksenova, M.A. Polyphenols in plants: Structure, biosynthesis, abiotic stress regulation, and practical applications (review). Int. J. Mol. Sci. 2023, 24, 13874. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhao, Y.; Chen, Y.; Jiang, C.; Wang, X.; Zhuo, R.; Lu, M.-Z.; Zang, J. Lignin biosynthesis and accumulation in response to abiotic stresses in woody plants. For. Res. 2022, 2, 9. [Google Scholar]
- Cheng, Z.-Y.; Sun, L.; Wang, X.-J.; Sun, R.; An, Y.-Q.; An, B.-L.; Zhu, M.-X.; Zhao, C.-F.; Bai, J.-G. Ferulic acid pretreatment alleviates heat stress in blueberry seedlings by inducing antioxidant enzymes, proline, and soluble sugars. Biol. Plant 2018, 62, 534–542. [Google Scholar]
- Alhaithloul, H.A.S.; Galal, F.H.; Seufi, A.M. Effect of extreme temperature changes on phenolic, flavonoid contents and antioxidant activity of tomato seedlings (Solanum lycopersicum L.). PeerJ 2021, 9, e11193. [Google Scholar]
- Khan, K.A.; Saleem, M.H.; Afzal, S.; Hussain, I.; Ameen, F.; Fahad, S. Ferulic acid: Therapeutic potential due to its antioxidant properties, role in plant growth, and stress tolerance. Plant Growth Regul. 2024, 104, 1329–1353. [Google Scholar]
- Kaplan, F.; Kopka, J.; Haskell, D.W.; Zhao, W.; Schiller, K.C.; Gatzke, N.; Sung, D.Y.; Guy, C.L. Exploring the temperature-stress metabolome of Arabidopsis. Plant Physiol. 2004, 136, 4159–4168. [Google Scholar]
- Solecka, D.; Boudet, A.-M.; Kacperska, A. Phenylpropanoid and anthocyanin changes in low-temperature treated winter oilseed rape leaves. Plant Physiol. Biochem. 1999, 37, 491–496. [Google Scholar]
- Huilan, Z.; Liu, S.; Yao, L.; Wang, L.; Li, C. Free and bound phenolics of buckwheat varieties: HPLC characterization, antioxidant activity, and inhibitory potency towards α-glucosidase with molecular docking analysis. Antioxidants 2019, 8, 606. [Google Scholar] [CrossRef]
- Prakash, O.; Baskaran, R.; Kudachikar, V.B. Characterization, quantification of free, esterified and bound phenolics in Kainth (Pyrus pashia Buch.-Ham. Ex D.Don) fruit pulp by UPLC-ESI-HRMS/MS and evaluation of their antioxidant activity. Food Chem. 2019, 299, 114–125. [Google Scholar]
- Yu, M.; Yang, L.; Xue, Q.; Yin, P.; Sun, L.; Liu, Y. Comparison of free, esterified, and insoluble-bound phenolics and their Bioactivities in three organs of Lonicera japonica and L. macranthoides. Molecules 2019, 24, 970. [Google Scholar] [CrossRef]
- Akoglu, H. User’s guide to correlation coefficients. Turk. J. Emerg. Med. 2018, 18, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Meghanathan, N. Assortativity analysis of real-world network graphs based on centrality metrics. Comput. Inf. Sci. 2016, 9, 7. [Google Scholar] [CrossRef]
- Greenacre, M.; Groenen, P.J.F.; Hastie, T.; D’Enza, A.I.; Markos, A.; Tuzhilina, E. Principal component analysis. Nat. Rev. Methods Primers 2022, 2, 100. [Google Scholar] [CrossRef]
- Dewitte, G.; Walmagh, M.; Diricks, M.; Lepak, A.; Gutmann, A.; Nidetzky, B.; Desmet, T. Screening of recombinant glycosyltransferases reveals the broad acceptor specificity of stevia UGT-76G1. J. Biotechnol. 2016, 233, 49–55. [Google Scholar] [CrossRef]
- Murtagh, F.; Contreras, P. Algorithms for hierarchical clustering: An overview. WIREs Data Mining Knowl. Discov. 2012, 2, 86–97. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Galvão, M.A.M.; de Arruda, A.O.; Ferraz Bezerra, I.C.; Ferreira, M.R.A.; Soares, L.A.L. Evaluation of the Folin-Ciocalteu method and quantification of total tannins in stem barks and pods from Libidibia ferrea (Mart. ex Tul) L. P. Queiroz. Braz. Arch. Biol. Technol. 2018, 61, e18170586. [Google Scholar] [CrossRef]
- Howard, L.R.; Clear, J.R.; Brownmiller, C. Antioxidant capacity and phenolic content in blueberries as affected by genotype and growing season. J. Sci. Food Agric. 2003, 83, 1238–1247. [Google Scholar] [CrossRef]
- Jain, R.; Rao, B.; Tare, A.B. Comparative analysis of the spectrophotometry based total phenolic acid estimation methods. J. Anal. Chem. 2017, 72, 972–976. [Google Scholar] [CrossRef]
- Šola, I.; Vujčić Bok, V.; Dujmović, M.; Rusak, G. Developmentally-related changes in phenolic and L-ascorbic acid content, and antioxidant capacity of Chinese cabbage sprouts. J. Food Sci. Technol. 2020, 57, 702–712. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ferulic Acid | Sinapic Acid | |
|---|---|---|
| (mg/g dw) | ||
| free | ||
| RT | 0.28 ± 0.04 b | 2.30 ± 0.41 b |
| HT | 0.65 ± 0.06 b | 2.01 ± 0.55 b |
| LT | 0.33 ± 0.05 b | 3.62 ± 0.87 ab |
| ΔHT (%) | 135.47 * | −12.69 |
| ΔLT (%) | 17.84 | 57.50 * |
| esterified | ||
| RT | 1.36 ± 0.44 a | 9.70 ± 0.25 a |
| HT | 2.33 ± 0.48 a | 9.12 ± 0.47 a |
| LT | 1.31 ± 0.27 a | 8.14 ± 4.83 a |
| ΔHT (%) | 70.82 | −6.02 |
| ΔLT (%) | −4.03 | −16.06 * |
| glycosylated | ||
| RT | 0.07 ± 0.01 b | 0.13 ± 0.03 c |
| HT | 0.09 ± 0.03 c | 0.08 ± 0.01 c |
| LT | 0.05 ± 0.01 b | 0.07 ± 0.01 b |
| ΔHT (%) | 36.94 | −36.84 |
| ΔLT (%) | −26.68 | −46.72 |
| bound | ||
| RT | 0.17 ± 00 b | 0.99 ± 00 c |
| HT | 0.11 ± 0.11 c | 0.47 ± 0.21 c |
| LT | 0.09 ± 0.05 b | 0.62 ± 0.42 b |
| ΔHT (%) | −36.68 | −52.21 |
| ΔLT (%) | −49.49 | −37.72 |
| sum | ||
| RT | 1.87 ± 0.47 | 13.12 ± 0.51 |
| HT | 3.18 ± 0.57 | 11.68 ± 0.56 |
| LT | 1.77 ± 0.19 | 12.45 ± 3.86 |
| ΔHT (%) | 69.53 * | −10.99 |
| ΔLT (%) | −5.68 | −5.10 |
| ABTS (% of Inhibition) | DPPH (% of Inhibition) | FRAP (% of Reduction) | |
|---|---|---|---|
| Free | |||
| RT | 48.77 ± 6.79 b | 78.87 ± 2.14 a | 80.90 ± 0.83 c |
| HT | 54.93 ± 4.81 b | 79.36 ± 2.32 a | 80.61 ± 3.09 c |
| LT | 59.32 ± 6.73 b | 79.86 ± 1.36 a | 85.46 ± 2.40 b |
| ΔHT (%) | 12.64 * | 0.63 | −0.36 |
| ΔLT (%) | 21.62 * | 1.26 | 5.63 * |
| Esterified | |||
| RT | 74.45 ± 3.21 a | 79.59 ± 2.27 a | 95.93 ± 0.63 a |
| HT | 73.37 ± 4.75 a | 78.81 ± 2.68 a | 95.95 ± 0.47 a |
| LT | 75.25 ± 3.88 a | 79.43 ± 2.55 a | 96.43 ± 0.45 a |
| ΔHT (%) | −1.45 | −0.98 | 0.02 |
| ΔLT (%) | 1.08 | −0.19 | 0.52 * |
| Glycosylated | |||
| RT | 57.71 ± 5.27 b | 80.86 ± 0.97 a | 86.46 ± 2.25 b |
| HT | 64.90 ± 1.93 a | 78.37 ± 1.98 a | 87.75 ± 2.13 b |
| LT | 60.35 ± 2.43 b | 79.69 ± 1.27 a | 86.49 ± 2.69 b |
| ΔHT (%) | 12.45 * | −3.08 * | 1.49 |
| ΔLT (%) | 4.58 | −1.45 | 0.03 |
| Bound | |||
| RT | 48.92 ± 21.27 b | 74.18 ± 8.30 b | 78.16 ± 8.03 c |
| HT | 45.27 ± 24.89 c | 79.70 ± 3.71 a | 81.57 ± 5.78 c |
| LT | 54.96 ± 10.15 b | 78.29 ± 3.68 a | 83.02 ± 2.62 c |
| ΔHT (%) | −7.47 | 7.44 | 4.37 |
| ΔLT (%) | 12.33 | 5.55 | 6.22 |
| Sum | |||
| RT | 57.46 ± 4.97 | 78.37 ± 2.79 | 85.36 ± 1.92 |
| HT | 59.62 ± 5.58 | 79.06 ± 1.80 | 86.47 ± 2.17 |
| LT | 62.47 ± 3.68 | 79.32 ± 1.65 | 87.85 ± 1.13 |
| ΔHT (%) | 3.75 | 0.88 | 1.30 |
| ΔLT (%) | 8.71 * | 1.21 | 2.91 * |
| Free | |||||||||||
| TP | TT | TPA | THCA | TF | TFLO | ABTS | DPPH | FRAP | Fer ac | Sin ac | |
| TP | 1.00 | ||||||||||
| TT | 0.89 | 1.00 | |||||||||
| TPA | 0.98 | 0.96 | 1.00 | ||||||||
| THCA | 0.99 | 0.82 | 0.95 | 1.00 | |||||||
| TF | 0.90 | 1.00 * | 0.97 | 0.84 | 1.00 | ||||||
| TFLO | 0.95 | 0.71 | 0.88 | 0.99 | 0.73 | 1.00 | |||||
| ABTS | 0.86 | 1.00 * | 0.94 | 0.78 | 1.00 * | 0.67 | 1.00 | ||||
| DPPH | 0.91 | 1.00 * | 0.97 | 0.84 | 1.00 | 0.74 | 1.00 | 1.00 | |||
| FRAP | 0.99 | 0.82 | 0.94 | 1.00 * | 0.84 | 0.99 | 0.78 | 0.84 | 1.00 | ||
| Fer ac | −0.31 | 0.16 | −0.12 | −0.44 | 0.12 | −0.58 | 0.22 | 0.12 | −0.44 | 1.00 | |
| Sin ac | 0.97 | 0.74 | 0.90 | 0.99 | 0.77 | 1.00 * | 0.70 | 0.77 | 0.99 | −0.54 | 1.00 |
| Esterified | |||||||||||
| TP | TT | TPA | THCA | TF | TFLO | ABTS | DPPH | FRAP | Fer ac | Sin ac | |
| TP | 1.00 | ||||||||||
| TT | 0.95 | 1.00 | |||||||||
| TPA | 0.67 | 0.87 | 1.00 | ||||||||
| THCA | 1.00 * | 0.94 | 0.64 | 1.00 | |||||||
| TF | 0.63 | 0.84 | 1.00 * | 0.60 | 1.00 | ||||||
| TFLO | 1.00 * | 0.94 | 0.66 | 1.00 * | 0.62 | 1.00 | |||||
| ABTS | 0.72 | 0.46 | −0.03 | 0.75 | −0.08 | 0.73 | 1.00 | ||||
| DPPH | 0.18 | −0.14 | −0.61 | 0.22 | −0.65 | 0.20 | 0.81 | 1.00 | |||
| FRAP | 0.99 | 0.90 | 0.58 | 1.00 | 0.53 | 0.99 | 0.80 | 0.30 | 1.00 | ||
| Fer ac | −0.41 | −0.09 | 0.41 | −0.44 | 0.46 | −0.42 | −0.92 | −0.97 | −0.51 | 1.00 | |
| Sin ac | −0.97 | −0.99 | −0.82 | −0.97 | −0.79 | −0.97 | −0.55 | 0.04 | −0.94 | 0.19 | 1.00 |
| Glycosylated | |||||||||||
| TP | TT | TPA | THCA | TF | TFLO | ABTS | DPPH | FRAP | Fer ac | Sin ac | |
| TP | 1.00 | ||||||||||
| TT | 0.69 | 1.00 * | |||||||||
| TPA | 0.77 | 0.99 | 1.00 | ||||||||
| THCA | 0.99 | 0.80 | 0.87 | 1.00 | |||||||
| TF | 0.72 | 1.00 | 1.00 | 0.83 | 1.00 | ||||||
| TFLO | 0.93 | 0.91 | 0.95 | 0.98 | 0.92 | 1.00 | |||||
| ABTS | 0.89 | 0.94 | 0.98 | 0.95 | 0.96 | 1.00 | 1.00 | ||||
| DPPH | −0.83 | −0.98 | −1.00 | −0.91 | −0.98 | −0.98 | −0.99 | 1.00 | |||
| FRAP | 0.99 | 0.77 | 0.84 | 1.00 * | 0.79 | 0.97 | 0.94 | −0.89 | 1.00 | ||
| Fer ac | 0.95 | 0.42 | 0.52 | 0.88 | 0.46 | 0.76 | 0.69 | −0.61 | 0.90 | 1.00 | |
| Sin ac | −0.22 | −0.86 | −0.79 | −0.38 | −0.84 | −0.56 | −0.64 | 0.73 | −0.33 | 0.11 | 1.00 |
| Insoluble | |||||||||||
| TP | TT | TPA | THCA | TF | TFLO | ABTS | DPPH | FRAP | Fer ac | Sin ac | |
| TP | 1.00 | ||||||||||
| TT | −0.18 | 1.00 | |||||||||
| TPA | −0.40 | −0.83 | 1.00 | ||||||||
| THCA | 0.78 | −0.76 | 0.26 | 1.00 | |||||||
| TF | 0.62 | −0.89 | 0.48 | 0.97 | 1.00 | ||||||
| TFLO | −0.53 | −0.74 | 0.99 | 0.12 | 0.34 | 1.00 | |||||
| ABTS | 0.96 | 0.10 | −0.64 | 0.57 | 0.37 | −0.75 | 1.00 | ||||
| DPPH | 0.18 | −1.00 * | 0.83 | 0.76 | 0.89 | 0.74 | −0.11 | 1.00 | |||
| FRAP | 0.66 | −0.86 | 0.42 | 0.99 | 1.00 * | 0.29 | 0.42 | 0.86 | 1.00 | ||
| Fer ac | −0.63 | 0.88 | −0.46 | −0.98 | −1.00 * | −0.33 | −0.38 | −0.88 | −1.00 * | 1.00 | |
| Sin ac | −0.16 | 1.00 * | −0.84 | −0.74 | −0.87 | −0.76 | 0.13 | −1.00 | −0.84 | 0.87 | 1.00 |
| Sum | |||||||||||
| TP | TT | TPA | THCA | TF | TFLO | ABTS | DPPH | FRAP | Fer ac | Sin ac | |
| TP | 1.00 | ||||||||||
| TT | 0.80 | 1.00 | |||||||||
| TPA | 0.57 | 0.95 | 1.00 | ||||||||
| THCA | 1.00 * | 0.76 | 0.51 | 1.00 | |||||||
| TF | 0.65 | 0.98 | 0.99 | 0.60 | 1.00 | ||||||
| TFLO | 0.97 | 0.92 | 0.74 | 0.95 | 0.81 | 1.00 | |||||
| ABTS | 0.98 | 0.89 | 0.70 | 0.97 | 0.77 | 1.00 * | 1.00 | ||||
| DPPH | 0.87 | 0.99 | 0.90 | 0.84 | 0.94 | 0.96 | 0.94 | 1.00 | |||
| FRAP | 0.98 | 0.90 | 0.71 | 0.97 | 0.78 | 1.00 * | 1.00 * | 0.95 | 1.00 | ||
| Fer ac | −0.32 | 0.32 | 0.60 | −0.38 | 0.51 | −0.09 | −0.15 | 0.19 | −0.13 | 1.00 | |
| Sin ac | −0.23 | −0.77 | −0.93 | −0.16 | −0.89 | −0.45 | −0.39 | −0.67 | −0.41 | −0.85 | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šola, I.; Gmižić, D. Structural Variations of Broccoli Polyphenolics and Their Antioxidant Capacity as a Function of Growing Temperature. Plants 2025, 14, 1186. https://doi.org/10.3390/plants14081186
Šola I, Gmižić D. Structural Variations of Broccoli Polyphenolics and Their Antioxidant Capacity as a Function of Growing Temperature. Plants. 2025; 14(8):1186. https://doi.org/10.3390/plants14081186
Chicago/Turabian StyleŠola, Ivana, and Daria Gmižić. 2025. "Structural Variations of Broccoli Polyphenolics and Their Antioxidant Capacity as a Function of Growing Temperature" Plants 14, no. 8: 1186. https://doi.org/10.3390/plants14081186
APA StyleŠola, I., & Gmižić, D. (2025). Structural Variations of Broccoli Polyphenolics and Their Antioxidant Capacity as a Function of Growing Temperature. Plants, 14(8), 1186. https://doi.org/10.3390/plants14081186