
Beneficial Effects of ACC Deaminase-Producing Rhizobacteria on the Drought Stress Resistance of Coffea arabica L.

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Identification and Characterization of ACCd-Producing Rhizobacteria Strains
2.2. Effect of ACC Deaminase-Producing Strains on the Growth and Physiological Status of Coffea arabica L.
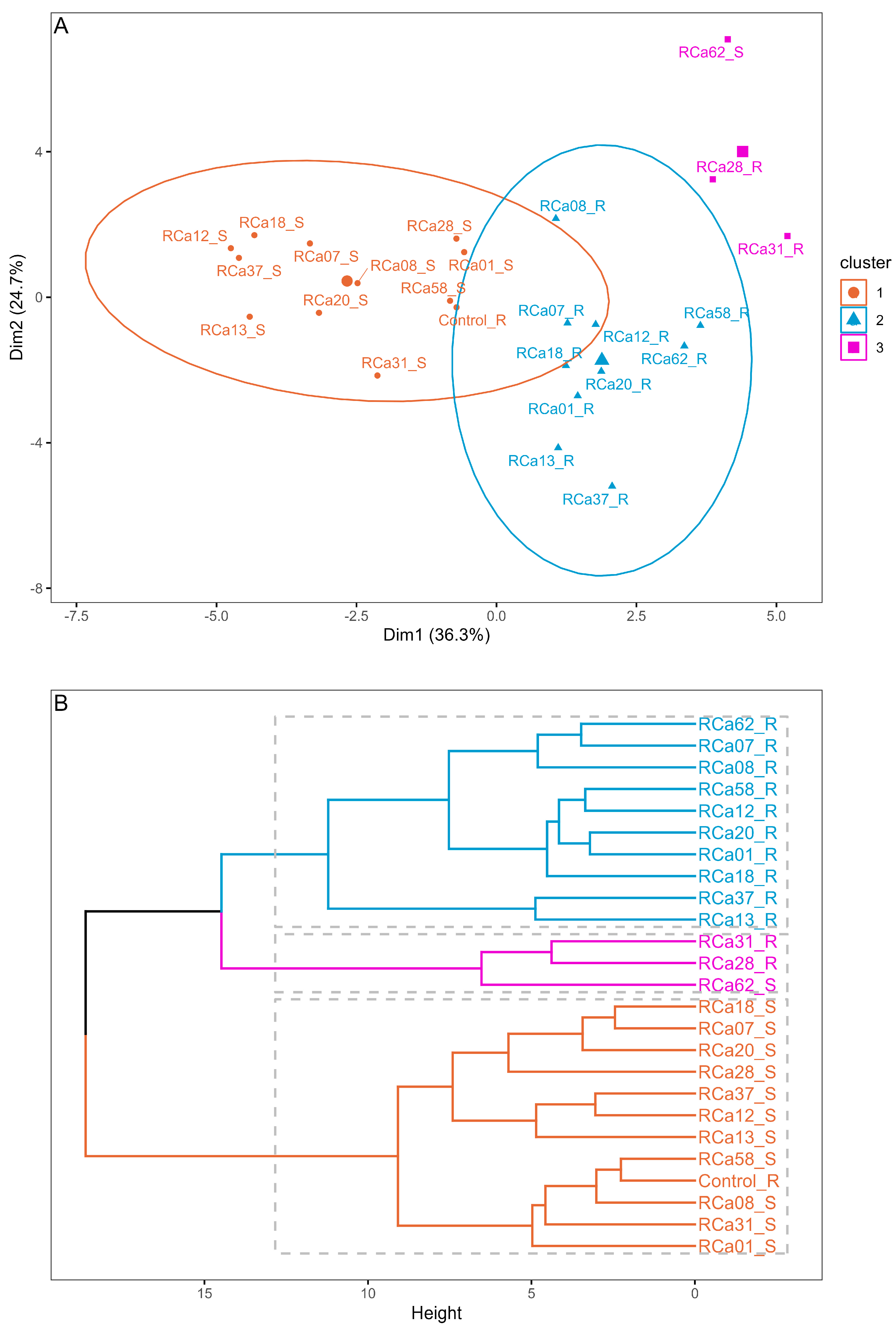
2.2.1. Differentiation Between Treatments
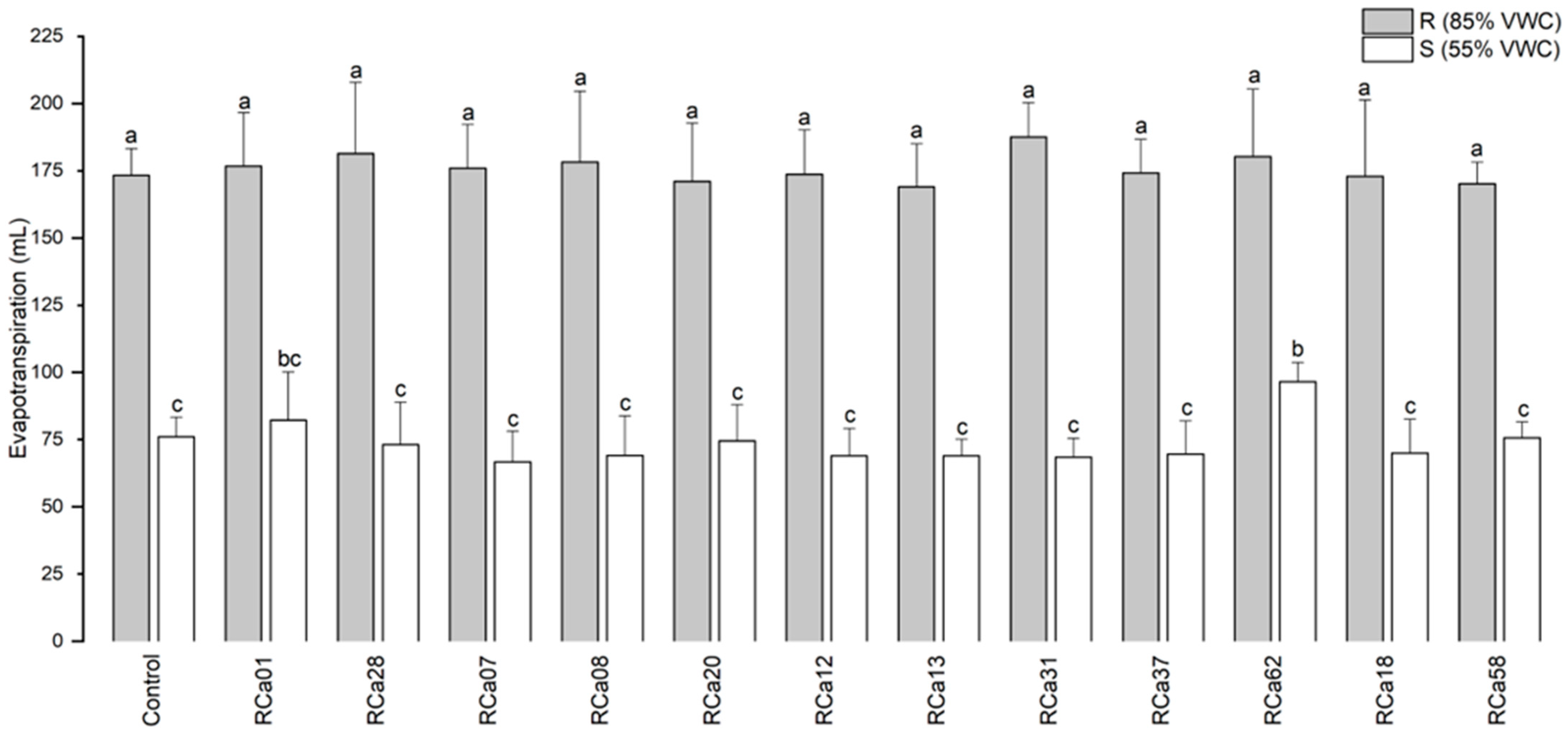
2.2.2. Effects on Evapotranspiration
2.2.3. Effects on Leaf Functional Traits
2.2.4. Effect on Plant Growth
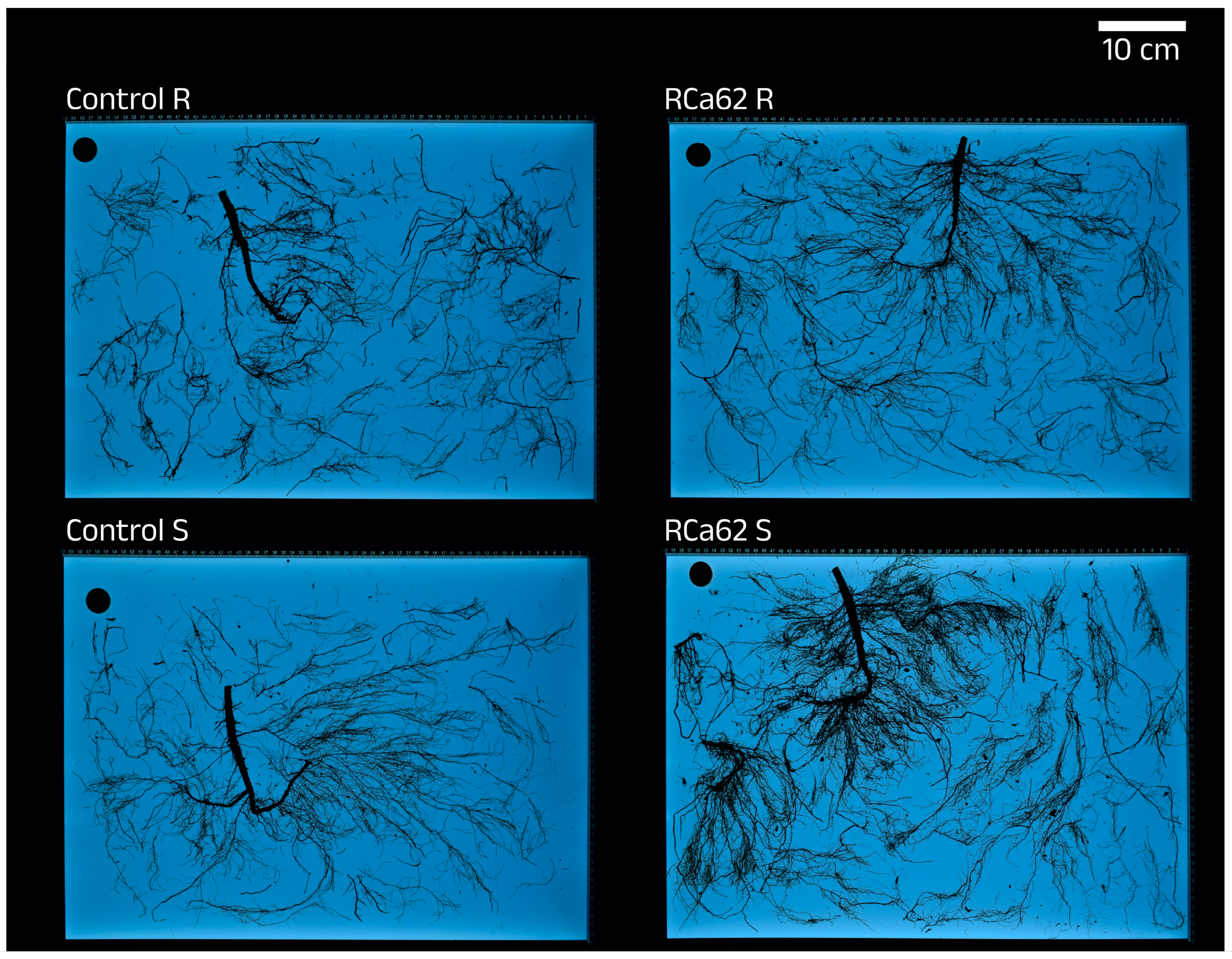
2.2.5. Effects on Root Development
2.2.6. Effect on Pigment Concentrations
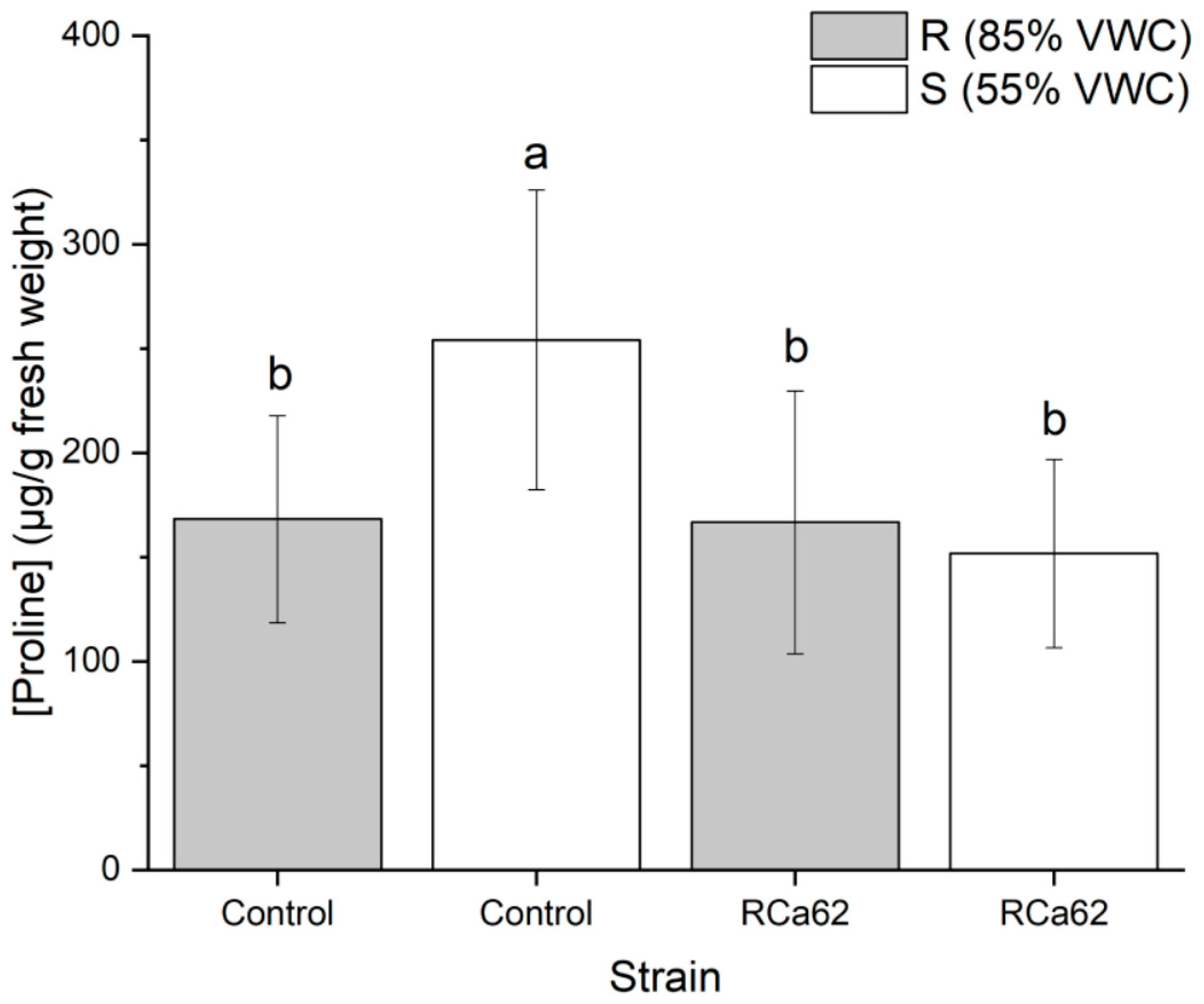
2.2.7. Effect on Proline Accumulation
3. Discussion
3.1. ACCd-Producing Rhizobacteria Strains
3.2. Bacterial Effect on Evapotranspiration
3.3. Bacterial Effect on Leaf Functional Traits
3.4. Bacterial Effect on Plant Growth
3.5. Bacterial Effect on Root Development
3.6. Bacterial Effect on Pigment Concentrations
3.7. Bacterial Effect on Proline Accumulation
3.8. Further Remarks
4. Materials and Methods
4.1. Sampling
4.2. Isolation and Selection of ACC Deaminase-Producing Rhizobacteria
4.3. Characterization of Plant Growth-Promoting Traits on ACC Deaminase-Producing Strains
4.4. Pot Experiment Design
4.5. Effects of ACC Deaminase-Producing Strains on the Growth and Physiological Status of Coffea arabica
4.6. Proline Accumulation
4.7. Data Analysis
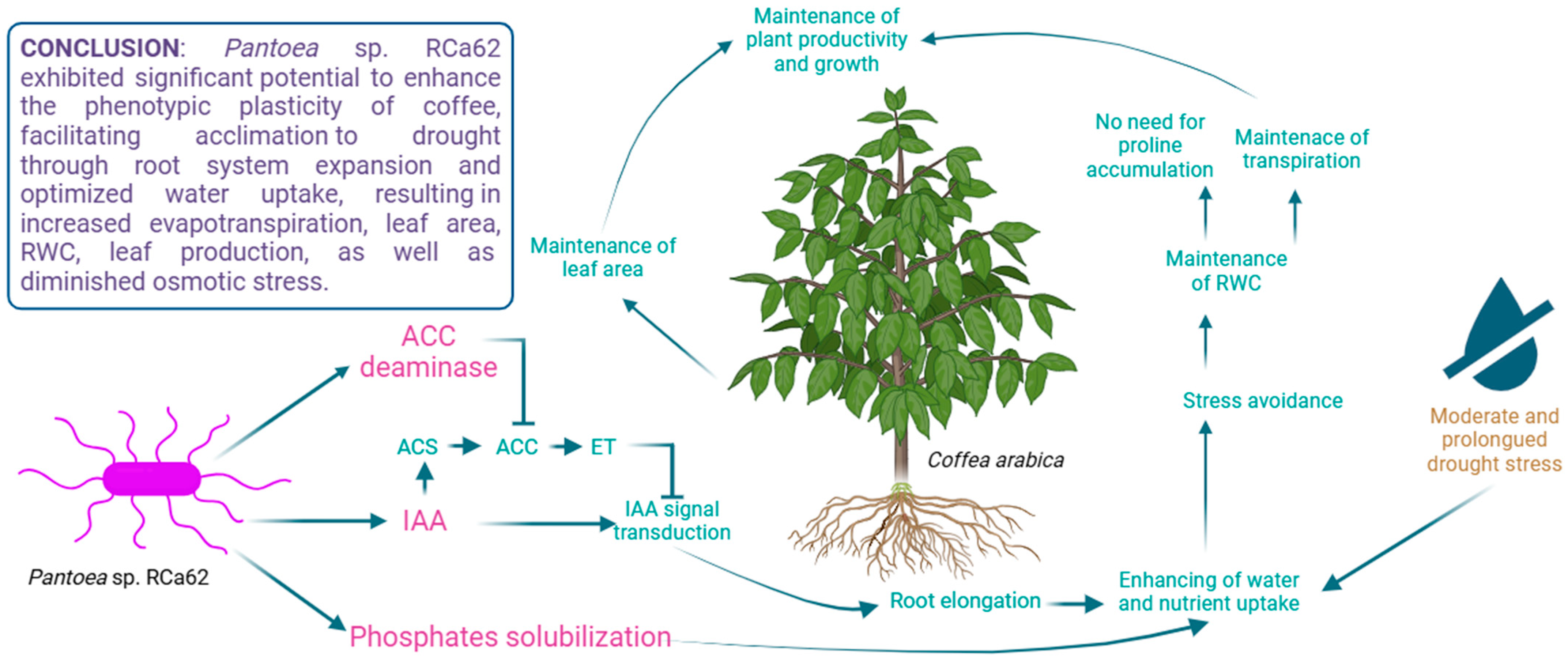
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACC | 1-aminocyclopropane-1-carboxylic acid |
| ACCd | ACC deaminase |
| PGPR | Plant growth promoting rhizobacteria |
| R | Full-irrigation treatment |
| RWC | Relative water content |
| S | Drought stress treatment |
| VWC | Volumetric water content |
References
- Beining, A.M. Ecophysiological Diversity of Wild Coffea arabica Populations in Ethiopia: Drought Adaptation Mechanisms. Ph.D. Thesis, Universität Bonn, Bonn, Germany, 2007. [Google Scholar]
- Amrouk, M.; Palmeri, F.; Magrini, E. Global Coffee Market and Recent Price Developments; FAO: Rome, Italy, 2025. [Google Scholar]
- Tucker, C.M. Coffee culture. In Local Experiences, Global Connections, 2nd ed.; Anthropology of Stuff series; Routledge: New York, NY, USA, 2018; ISBN 9781315678795. [Google Scholar]
- Wright, D.R.; Gordon, A.; Bennett, R.E.; Selinske, M.J.; Lentini, P.E.; Garrard, G.E.; Rodewald, A.D.; Bekessy, S.A. Biodiverse coffee plantations provide co-benefits without compromising yield. J. Sustain. Agric. Environ. 2024, 3, 1–12. [Google Scholar] [CrossRef]
- Lugo-Pérez, J.; Hajian-Forooshani, Z.; Perfecto, I.; Vandermeer, J. The importance of shade trees in promoting carbon storage in the coffee agroforest systems. Agric. Ecosyst. Environ. 2023, 355, 108594. [Google Scholar] [CrossRef]
- Wildtruth, F.; Perfecto, I. Effects of canopy connectivity on the arboreal ant community in coffee shade trees. Biotropica 2023, 55, 1106–1113. [Google Scholar] [CrossRef]
- Villers, L.; Arizpe, N.; Orellana, R.; Conde, C.; Hernandez, J. Impactos del cambio climático en la floración y desarrollo del fruto del café en Veracruz, Mexico. Interciencia 2009, 34, 322–329. [Google Scholar]
- Melke, A.; Fetene, M. Eco-physiological basis of drought stress in coffee (Coffea arabica, L.) in Ethiopia. Theor. Exp. Plant Physiol. 2014, 26, 225–239. [Google Scholar] [CrossRef]
- Menezes-Silva, P.E.; Sanglard, L.M.V.P.; Ávila, R.T.; Morais, L.E.; Martins, S.C.V.; Nobres, P.; Patreze, C.M.; Ferreira, M.A.; Araújo, W.L.; Fernie, A.R.; et al. Photosynthetic and metabolic acclimation to repeated drought events play key roles in drought tolerance in coffee. J. Exp. Bot. 2017, 68, 4309–4322. [Google Scholar] [CrossRef]
- Da Matta, F.M.; Ramalho, C. Impacts of drought and temperature stress on coffee physiology and production: A review. Brazilian J. Plant Physiol. 2006, 18, 55–81. [Google Scholar] [CrossRef]
- Ahmed, S.; Brinkley, S.; Smith, E.; Sela, A.; Theisen, M.; Thibodeau, C.; Warne, T.; Anderson, E.; Van Dusen, N.; Giuliano, P.; et al. Climate change and coffee quality: Systematic review on the effects of environmental and management variation on secondary metabolites and sensory attributes of Coffea arabica and Coffea canephora. Front. Plant Sci. 2021, 12, 708013. [Google Scholar] [CrossRef]
- da Silva Tavares, P.; Giarolla, A.; Chou, S.C.; de Paula Silva, A.J.; de Arruda Lyra, A. Climate change impact on the potential yield of Arabica coffee in southeast Brazil. Reg. Environ. Change 2018, 18, 873–883. [Google Scholar] [CrossRef]
- Laderach, P.; Lundy, M.; Jarvis, A.; Ramirez, J.; Portilla, E.P.; Schepp, K.; Eitzinger, A. Predicted impact of climate change on coffee supply chains. In The Economic, Social and Political Elements of Climate Change; Filho, W.L., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 703–723. ISBN 978-3-642-14775-3. [Google Scholar]
- Pham, Y.; Reardon-Smith, K.; Mushtaq, S.; Cockfield, G. The impact of climate change and variability on coffee production: A systematic review. Clim. Change 2019, 156, 609–630. [Google Scholar] [CrossRef]
- Ovalle-Rivera, O.; Läderach, P.; Bunn, C.; Obersteiner, M.; Schroth, G. Projected shifts in Coffea arabica suitability among major global producing regions due to climate change. PLoS ONE 2015, 10, e0124155. [Google Scholar] [CrossRef]
- Jawo, T.O.; Kyereh, D.; Lojka, B. The impact of climate change on coffee production of small farmers and their adaptation strategies: A review. Clim. Dev. 2023, 15, 93–109. [Google Scholar] [CrossRef]
- Kim, T.-W.; Jehanzaib, M. Drought risk analysis, forecasting and assessment under climate change. Water 2020, 12, 1862. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Deepak; Singh, S.; Bakshi, M.; Bansal; Sangeeta; Bansal, S.; Thakur, A.; Singh, S.; Bakshi, M.; Bansal, S. Changes in crop physiology under drought stress: A review. J. Pharmacogn. Phytochem. 2019, 8, 1251–1253. [Google Scholar]
- Dietz, K.J.; Zörb, C.; Geilfus, C.M. Drought and crop yield. Plant Biol. 2021, 23, 881–893. [Google Scholar] [CrossRef]
- Grover, M.; Bodhankar, S.; Sharma, A.; Sharma, P.; Singh, J.; Nain, L. PGPR mediated alterations in root traits: Way toward sustainable crop production. Front. Sustain. Food Syst. 2021, 4, 287. [Google Scholar] [CrossRef]
- Barnawal, D.; Singh, R.; Singh, R.P. Role of plant growth promoting rhizobacteria in drought tolerance. In PGPR Amelioration in Sustainable Agriculture; Singh, A.K., Kumar, A., Singh, P.K., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 107–128. ISBN 9780128158791. [Google Scholar]
- Shaffique, S.; Shah, A.A.; Odongkara, P.; Elansary, H.O.; Khan, A.L.; Adhikari, A.; Kang, S.M.; Lee, I.J. Deciphering the ABA and GA biosynthesis approach of Bacillus pumilus, mechanistic approach, explaining the role of metabolic region as an aid in improving the stress tolerance. Sci. Rep. 2024, 14, 28923. [Google Scholar] [CrossRef]
- Ali, S.; Khan, N. Delineation of mechanistic approaches employed by plant growth promoting microorganisms for improving drought stress tolerance in plants. Microbiol. Res. 2021, 249, 126771. [Google Scholar] [CrossRef]
- Fernandes, I.; Marques, I.; Paulo, O.S.; Batista, D.; Partelli, F.L.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. Understanding the impact of drought in Coffea genotypes: Transcriptomic analysis supports a common high resilience to moderate water deficit but a genotype dependent sensitivity to severe water deficit. Agronomy 2021, 11, 2255. [Google Scholar] [CrossRef]
- Sati, D.; Pande, V.; Pandey, S.C.; Samant, M. Recent advances in PGPR and molecular mechanisms involved in drought stress resistance. J. Soil Sci. Plant Nutr. 2023, 23, 106–124. [Google Scholar] [CrossRef]
- Gowtham, H.G.; Singh, S.B.; Shilpa, N.; Aiyaz, M.; Nataraj, K.; Udayashankar, A.C.; Amruthesh, K.N.; Murali, M.; Poczai, P.; Gafur, A.; et al. Insight into recent progress and perspectives in improvement of antioxidant machinery upon PGPR augmentation in plants under drought stress: A review. Antioxidants 2022, 11, 1763. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Stress control and ACC deaminase. In Principles of Plant-Microbe Interactions; Lugtenberg, B., Ed.; Springer: Cham, Switzerland, 2015; pp. 257–264. ISBN 9783319085753. [Google Scholar]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.d.C.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt stress in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, L.; Hao, R.; Bai, X.; Wang, Y.; Yu, X. Drought-tolerant plant growth-promoting rhizobacteria isolated from jujube (Ziziphus jujuba) and their potential to enhance drought tolerance. Plant Soil 2020, 452, 423–440. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-Ul-Hye, M.; Hussain, S.; Riaz, M.; Qayyum, M.F. Mitigation of drought stress in maize through inoculation with drought tolerant ACC deaminase containing PGPR under axenic conditions. Pakistan J. Bot. 2020, 52, 49–60. [Google Scholar] [CrossRef]
- Buqori, D.M.A.I.; Sugiharto, B.; Suherman; Siswoyo, T.A.; Hariyono, K. Mitigating drought stress by application of drought-tolerant Bacillus spp. enhanced root architecture, growth, antioxidant and photosynthetic genes expression in sugarcane. Sci. Rep. 2025, 15, 5259. [Google Scholar] [CrossRef]
- Gupta, S.; Pandey, S. ACC deaminase producing bacteria with multifarious plant growth promoting traits alleviates salinity stress in french bean (Phaseolus vulgaris) plants. Front. Microbiol. 2019, 10, 1506. [Google Scholar] [CrossRef]
- Gupta, A.; Rai, S.; Bano, A.; Sharma, S.; Kumar, M.; Binsuwaidan, R.; Suhail Khan, M.; Upadhyay, T.K.; Alshammari, N.; Saeed, M.; et al. ACC Deaminase produced by PGPR mitigates the adverse effect of osmotic and salinity stresses in Pisum sativum through modulating the antioxidants activities. Plants 2022, 11, 3419. [Google Scholar] [CrossRef]
- Yuan, Y.; Shi, Y.; Liu, Z.; Fan, Y.; Liu, M.; Ningjing, M.; Li, Y. Promotional properties of ACC deaminase-producing bacterial strain DY1-3 and its enhancement of maize resistance to salt and drought stresses. Microorganisms 2023, 11, 2654. [Google Scholar] [CrossRef]
- Urgiles-Gómez, N.; Avila-Salem, M.E.; Loján, P.; Encalada, M.; Hurtado, L.; Araujo, S.; Collahuazo, Y.; Guachanamá, J.; Poma, N.; Granda, K.; et al. Plant growth-promoting microorganisms in coffee production: From isolation to field application. Agronomy 2021, 11, 1531. [Google Scholar] [CrossRef]
- Arshad, M.; Shaharoona, B.; Mahmood, T. Inoculation with Pseudomonas spp. containing ACC-deaminase partially eliminates the effects of drought stress on growth, yield, and ripening of pea (Pisum sativum L.). Pedosphere 2008, 18, 611–620. [Google Scholar] [CrossRef]
- Bhise, K.K.; Bhagwat, P.K.; Dandge, P.B. Synergistic effect of Chryseobacterium gleum sp. SUK with ACC deaminase activity in alleviation of salt stress and plant growth promotion in Triticum aestivum L. 3 Biotech 2017, 7, 105. [Google Scholar] [CrossRef]
- Panwar, M.; Tewari, R.; Gulati, A.; Nayyar, H. Indigenous salt-tolerant rhizobacterium Pantoea dispersa (PSB3) reduces sodium uptake and mitigates the effects of salt stress on growth and yield of chickpea. Acta Physiol. Plant. 2016, 38, 278. [Google Scholar] [CrossRef]
- Shaharoona, B.; Arshad, M.; Zahir, Z.A.; Khalid, A. Performance of Pseudomonas spp. containing ACC-deaminase for improving growth and yield of maize (Zea mays L.) in the presence of nitrogenous fertilizer. Soil Biol. Biochem. 2006, 38, 2971–2975. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. The PGPR Stenotrophomonas maltophilia SBP-9 augments resistance against biotic and abiotic stress in wheat plants. Front. Microbiol. 2017, 8, 1945. [Google Scholar] [CrossRef]
- Wu, Z.; Yue, H.; Lu, J.; Li, C. Characterization of rhizobacterial strain Rs-2 with ACC deaminase activity and its performance in promoting cotton growth under salinity stress. World J. Microbiol. Biotechnol. 2012, 28, 2383–2393. [Google Scholar] [CrossRef]
- Cardinale, M.; Ratering, S.; Suarez, C.; Zapata Montoya, A.M.; Geissler-Plaum, R.; Schnell, S. Paradox of plant growth promotion potential of rhizobacteria and their actual promotion effect on growth of barley (Hordeum vulgare L.) under salt stress. Microbiol. Res. 2015, 181, 22–32. [Google Scholar] [CrossRef]
- Gupta, G.; Jha, P. Screening of potential PGPR candidates as future biofertilizers-A strategic approach from lab to field. Res. J. Biotechnol. 2015, 10, 48–62. [Google Scholar]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef]
- Dunn, M.F.; Becerra-Rivera, V.A. The biosynthesis and functions of polyamines in the interaction of plant growth-promoting rhizobacteria with plants. Plants 2023, 12, 2671. [Google Scholar] [CrossRef] [PubMed]
- Knights, H.E.; Jorrin, B.; Haskett, T.L.; Poole, P.S. Deciphering bacterial mechanisms of root colonization. Environ. Microbiol. Rep. 2021, 13, 428–444. [Google Scholar] [CrossRef] [PubMed]
- Muleta, D.; Assefa, F.; Hjort, K.; Roos, S.; Granhall, U. Characterization of rhizobacteria isolated from wild Coffea arabica L. Eng. Life Sci. 2009, 9, 100–108. [Google Scholar] [CrossRef]
- Vega, F.E.; Pava-Ripoll, M.; Posada, F.; Buyer, J.S. Endophytic bacteria in Coffea arabica L. J. Basic Microbiol. 2005, 45, 371–380. [Google Scholar] [CrossRef]
- Chen, M.; Li, N.; Zhang, X.-F.; Zhou, X.-K.; Shi, R.; Su, Y.-X.; Liu, J.-J.; Cao, Y.; Mo, M.H.; Ma, L. Sphingobacterium faecale sp. nov., a 1-aminocyclopropane-1-carboxylate deaminase producing bacterium isolated from camel faeces. Int. J. Syst. Evol. Microbiol. 2022, 72, 005215. [Google Scholar] [CrossRef]
- Simarmata, R.; Ngadiman, N.; Rohman, M.S.; Simanjuntak, P. Identification of 1-aminocyclopropane-1-carboxilid acid (ACC)-deaminase producing endophytic bacteria from local agricultural plantation based on 16S ribosomal RNA gene as genetic marker. Biotropic J. Trop. Biol. 2019, 3, 13–23. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. The multifarious PGPR Serratia marcescens CDP-13 augments induced systemic resistance and enhanced salinity tolerance of wheat (Triticum aestivum L.). PLoS ONE 2016, 11, e0155026. [Google Scholar] [CrossRef]
- Zahir, Z.A.; Ghani, U.; Naveed, M.; Nadeem, S.M.; Asghar, H.N. Comparative effectiveness of Pseudomonas and Serratia sp. containing ACC-deaminase for improving growth and yield of wheat (Triticum aestivum L.) under salt-stressed conditions. Arch. Microbiol. 2009, 191, 415–424. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef]
- Palmer, M.; Steenkamp, E.T.; Coetzee, M.P.A.; Chan, W.Y.; van Zyl, E.; De Maayer, P.; Coutinho, T.A.; Blom, J.; Smits, T.H.M.; Duffy, B.; et al. Phylogenomic resolution of the bacterial genus Pantoea and its relationship with Erwinia and Tatumella. Antonie Van Leeuwenhoek 2017, 110, 1287–1309. [Google Scholar] [CrossRef]
- Mergaert, J.; Verdonck, L.; Kersters, K. Transfer of Erwinia ananas (synonym, Erwinia uredovora) and Erwinia stewartii to the genus Pantoea emend. as Pantoea ananas (Serrano 1928) comb. nov. and Pantoea stewartii (Smith 1898) comb. nov., respectively, an. Int. J. Syst. Bacteriol. 1993, 43, 162–173. [Google Scholar] [CrossRef]
- Muleta, D.; Assefa, F.; Börjesson, E.; Granhall, U. Phosphate-solubilising rhizobacteria associated with Coffea arabica L. in natural coffee forests of southwestern Ethiopia. J. Saudi Soc. Agric. Sci. 2013, 12, 73–84. [Google Scholar] [CrossRef]
- Mekete, T.; Hallmann, J.; Kiewnick, S.; Sikora, R. Endophytic bacteria from ethiopian coffee plants and their potential to antagonise meloidogyne incognita. Nematology 2009, 11, 117–127. [Google Scholar] [CrossRef]
- Ferreira Pires, J.; de Souza Cardoso, L.; Freitas Schwan, R.; Ferreira Silva, C. Diversity of microbiota found in coffee processing wastewater treatment plant. World J. Microbiol. Biotechnol. 2017, 33, 211. [Google Scholar] [CrossRef]
- Solis Pino, A.F.; Delgado Espinosa, Z.Y.; Ramos Cabrera, E.V. Characterization of the rhizosphere bacterial microbiome and coffee bean fermentation in the Castillo-Tambo and Bourbon varieties in the Popayán-Colombia plateau. BMC Plant Biol. 2023, 23, 217. [Google Scholar] [CrossRef]
- Seyedsayamdost, M.R.; Cleto, S.; Carr, G.; Vlamakis, H.; João Vieira, M.; Kolter, R.; Clardy, J. Mixing and matching siderophore clusters: Structure and biosynthesis of serratiochelins from Serratia sp. V4. J. Am. Chem. Soc. 2012, 134, 13550–13553. [Google Scholar] [CrossRef]
- Weakland, D.R.; Smith, S.N.; Bell, B.; Tripathi, A.; Mobley, H.L.T. The Serratia marcescens siderophore serratiochelin is necessary for full virulence during bloodstream infection. Infect. Immun. 2020, 88, e00117-20. [Google Scholar] [CrossRef]
- Appel, T.M.; Quijano-Martínez, N.; De La Cadena, E.; Mojica, M.F.; Villegas, M.V. Microbiological and clinical aspects of Raoultella spp. Front. Public Health 2021, 9, 686789. [Google Scholar] [CrossRef]
- Santoyo, G.; Valencia-Cantero, E.; Orozco-Mosqueda, M.d.C.; Peña-Cabriales, J.J.; Farías-Rodríguez, R. Role of siderophores in antagonic activity of Pseudomonas fluorescens ZUM80 against plant fungi. Terra Latinoam. 2010, 28, 53–60. [Google Scholar]
- Sivasakthi, S.; Usharani, G.; Saranraj, P. Biocontrol potentiality of plant growth promoting bacteria (PGPR)—Pseudomonas fluorescens and Bacillus subtilis: A review. Afr. J. Agric. Res. 2014, 9, 1265–1277. [Google Scholar]
- Ciríaco da Silva, E.; Custódio Nogueira, R.J.; Almeida da Silva, M.; Bandeira de Albuquerque, M. Drought stress and plant nutrition. Plant Stress 2011, 5, 32–41. [Google Scholar]
- Chen, Q.; Liu, S. Identification and characterization of the phosphate-solubilizing bacterium Pantoea sp. S32 in reclamation soil in Shanxi, China. Front. Microbiol. 2019, 10, 2171. [Google Scholar] [CrossRef]
- Li, L.; Chen, R.; Zuo, Z.; Lv, Z.; Yang, Z.; Mao, W.; Liu, Y.; Zhou, Y.; Huang, J.; Song, Z. Evaluation and improvement of phosphate solubilization by an isolated bacterium Pantoea agglomerans ZB. World J. Microbiol. Biotechnol. 2020, 36, 27. [Google Scholar] [CrossRef]
- Son, H.J.; Park, G.T.; Cha, M.S.; Heo, M.S. Solubilization of insoluble inorganic phosphates by a novel salt- and pH-tolerant Pantoea agglomerans R-42 isolated from soybean rhizosphere. Bioresour. Technol. 2006, 97, 204–210. [Google Scholar] [CrossRef]
- Apine, O.A.; Jadhav, J.P. Optimization of medium for indole-3-acetic acid production using Pantoea agglomerans strain PVM. J. Appl. Microbiol. 2011, 110, 1235–1244. [Google Scholar] [CrossRef]
- Novák, V. Evapotranspiration in the Soil-Plant-Atmosphere System; Springer Netherlands: Dordrecht, The Netherlands, 2012; ISBN 978-94-007-3839-3. [Google Scholar]
- Martins, S.C.V.; Sanglard, M.L.; Morais, L.E.; Menezes-Silva, P.E.; Mauri, R.; Avila, R.T.; Vital, C.E.; Cardoso, A.A.; DaMatta, F.M. How do coffee trees deal with severe natural droughts? An analysis of hydraulic, diffusive and biochemical components at the leaf level. Trees Struct. Funct. 2019, 33, 1679–1693. [Google Scholar] [CrossRef]
- Carvalho da Silva, P.; Ribeiro Junior, W.Q.; Gerosa Ramos, M.L.; Cruz Rocha, O.; Delly Veiga, A.; Henriques Silva, N.; de Oliveira Brasileiro, L.; Cardoso Santana, C.; Filgueiras Soares, G.; Vitória Malaquias, J.; et al. Physiological changes of arabica coffee under different intensities and durations of water stress in the Brazilian Cerrado. Plants 2022, 11, 2198. [Google Scholar] [CrossRef]
- León-Burgos, A.F.; Unigarro, C.; Balaguera-López, H.E. Can prolonged conditions of water deficit alter photosynthetic performance and water relations of coffee plants in central-west Colombian? S. Afr. J. Bot. 2022, 149, 366–375. [Google Scholar] [CrossRef]
- Da Matta, F.M.; Chaves, A.R.M.; Pinheiro, H.A.; Ducatti, C.; Loureiro, M.E. Drought tolerance of two field-grown clones of Coffea canephora. Plant Sci. 2003, 164, 111–117. [Google Scholar] [CrossRef]
- Avila, R.T.; Cardoso, A.A.; de Almeida, W.L.; Costa, L.C.; Machado, K.L.G.; Barbosa, M.L.; de Souza, R.P.B.; Oliveira, L.A.; Batista, D.S.; Martins, S.C.V.; et al. Coffee plants respond to drought and elevated [CO2] through changes in stomatal function, plant hydraulic conductance, and aquaporin expression. Environ. Exp. Bot. 2020, 177, 104148. [Google Scholar] [CrossRef]
- Naves, V.L. Coffee Trees Under Rainfall Exclusion: Evidences for Canopy Acclimation to Water Shortage. Ph.D. Thesis, Universidade Federal de Lavras, Lavras, Brazil, 2018. [Google Scholar]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought stress in plants: Causes, consequences, and tolerance. In Drought Stress Tolerance in Plants, Vol 1: Physiology and Biochemistry; Hossain, M., Wani, S., Bhattacharjee, S., Burritt, D., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2016; pp. 1–16. ISBN 9783319288994. [Google Scholar]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An overview. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–33. [Google Scholar]
- Hu, W.; Lu, Z.; Meng, F.; Li, X.; Cong, R.; Ren, T.; Sharkey, T.D.; Lu, J. The reduction in leaf area precedes that in photosynthesis under potassium deficiency: The importance of leaf anatomy. New Phytol. 2020, 227, 1749–1763. [Google Scholar] [CrossRef] [PubMed]
- Kathpalia, R.; Bhatla, S.C. Plant water relations. In Plant Physiology, Development and Metabolism; Bhatla, S., Lal, M., Eds.; Springer: Singapore, 2018; pp. 3–36. [Google Scholar]
- Sharma-Natu, P.; Ghildiyal, M.C. Potential targets for improving photosynthesis and crop yield. Curr. Sci. 2005, 88, 1918–1928. [Google Scholar]
- Da Matta, F.M. Exploring drought tolerance in coffee: A physiological approach with some insights for plant breeding. Brazilian J. Plant Physiol. 2004, 16, 1–6. [Google Scholar] [CrossRef]
- Cai, Z.Q.; Chen, Y.J.; Guo, Y.H.; Cao, K.F. Responses of two field-grown coffee species to drought and re-hydration. Photosynthetica 2005, 43, 187–193. [Google Scholar] [CrossRef]
- Worku, M.; Astatkie, T. Dry matter partitioning and physiological responses of Coffea arabica varieties to soil moisture deficit stress at the seedling stage in Southwest Ethiopia. African J. Agric. Res. 2010, 5, 2066–2072. [Google Scholar]
- Khan, A.; Singh, A.V. Multifarious effect of ACC deaminase and EPS producing Pseudomonas sp. and Serratia marcescens to augment drought stress tolerance and nutrient status of wheat. World J. Microbiol. Biotechnol. 2021, 37, 198. [Google Scholar] [CrossRef]
- Gowtham, H.G.; Singh, B.; Murail, M.; Shilpa, N.; Prasad, M.; Aiyaz, M.; Amruthesh, K.N.; Niranjana, S.R. Induction of drought tolerance in tomato upon the application of ACC deaminase producing plant growth promoting rhizobacterium Bacillus subtilis Rhizo SF 48. Microbiol. Res. 2020, 234, 126422. [Google Scholar] [CrossRef]
- Parkash, V.; Singh, S. A review on potential plant-basedwater stress indicators for vegetable crops. Sustainability 2020, 12, 3945. [Google Scholar] [CrossRef]
- Cisneros, C.A.; Franco, J.M.; Realpe Fernández, M.; Fuenmayor, J.C. Influencia de microorganismos en la disponibilidad de fósforo en plántulas de café (Coffea arabica). Biotecnoloía En El Sect. Agropecu. Y Agroindustrial 2017, 15, 19. [Google Scholar] [CrossRef]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Rivas-Ubach, A.; Oravec, M.; Vecerova, K.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; et al. Opposite metabolic responses of shoots and roots to drought. Sci. Rep. 2014, 4, 6829. [Google Scholar] [CrossRef]
- Lynch, J.P.; Chimungu, J.G.; Brown, K.M. Root anatomical phenes associated with water acquisition from drying soil: Targets for crop improvement. J. Exp. Bot. 2014, 65, 6155–6166. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Souchère, V., Alberola, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. ISBN 9789048126668. [Google Scholar]
- Fonta, J.E.; Giri, J.; Vejchasarn, P.; Lynch, J.P.; Brown, K.M. Spatiotemporal responses of rice root architecture and anatomy to drought. Plant Soil 2022, 479, 443–464. [Google Scholar] [CrossRef]
- Fry, E.L.; Evans, A.L.; Sturrock, C.J.; Bullock, J.M.; Bardgett, R.D. Root architecture governs plasticity in response to drought. Plant Soil 2018, 433, 189–200. [Google Scholar] [CrossRef]
- Aloni, R.; Aloni, E.; Langhans, M.; Ullrich, C.I. Role of cytokinin and auxin in shaping root architecture: Regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. Ann. Bot. 2006, 97, 883–893. [Google Scholar] [CrossRef]
- Fu, X.; Harberd, N.P. Auxin promotes Arabidopsis root growth by modulating gibberellin response. Nature 2003, 421, 740–743. [Google Scholar] [CrossRef]
- Shi, H.; Chen, L.; Ye, T.; Liu, X.; Ding, K.; Chan, Z. Modulation of auxin content in Arabidopsis confers improved drought stress resistance. Plant Physiol. Biochem. 2014, 82, 209–217. [Google Scholar] [CrossRef]
- Uzma, M.; Iqbal, A.; Hasnain, S. Drought tolerance induction and growth promotion by indole acetic acid producing Pseudomonas aeruginosa in Vigna radiata. PLoS ONE 2022, 17, e0262932. [Google Scholar] [CrossRef]
- Etesami, H.; Glick, B.R. Bacterial indole-3-acetic acid: A key regulator for plant growth, plant-microbe interactions, and agricultural adaptive resilience. Microbiol. Res. 2024, 281, 127602. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Pandey, S.S.; Pandey, A.; Chanotiya, C.S.; Kalra, A. Plant growth-promoting rhizobacteria enhance wheat salt and drought stress tolerance by altering endogenous phytohormone levels and TaCTR1/TaDREB2 expression. Physiol. Plant. 2017, 161, 502–514. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Hassan, M.J.; Li, Z.; Peng, Y. Indole-3-acetic acid improves drought tolerance of white clover via activating auxin, abscisic acid and jasmonic acid related genes and inhibiting senescence genes. BMC Plant Biol. 2020, 20, 150. [Google Scholar] [CrossRef]
- Firdaus, N.K.; Oktafiyanto, M.F.; Sasmita, K.D.; Pranowo, D. Efficacy of phytohormones producing isolates on the growth of two stem-cutting robusta coffee (Coffea canephora). IOP Conf. Ser. Earth Environ. Sci. 2023, 1172, 012002. [Google Scholar] [CrossRef]
- Zemlyanskaya, E.V.; Omelyanchuk, N.A.; Ubogoeva, E.V.; Mironova, V.V. Deciphering auxin-ethylene crosstalk at a systems level. Int. J. Mol. Sci. 2018, 19, 4060. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, J.; Zhu, M.; Wan, H.; Chen, Z.; Yang, N.; Duan, J.; Wei, Z.; Hu, T.; Liu, F. Effects of plant growth promoting hizobacteria (PGPR) strain Bacillus licheniformis with biochar amendment on potato growth and water use efficiency under reduced irrigation regime. Agronomy 2022, 12, 1031. [Google Scholar] [CrossRef]
- Le, A.T.; Pék, Z.; Takács, S.; Neményi, A.; Helyes, L. The effect of plant growth-promoting rhizobacteria on yield, water use efficiency and Brix degree of processing tomato. Plant, Soil Environ. 2018, 64, 523–529. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Amby, D.B.; Hegelund, J.N.; Fimognari, L.; Großkinsky, D.K.; Westergaard, J.C.; Müller, R.; Moelbak, L.; Liu, F.; Roitsch, T. Bacillus licheniformis FMCH001 increases water use efficiency via growth stimulation in both normal and drought conditions. Front. Plant Sci. 2020, 11, 297. [Google Scholar] [CrossRef]
- Gholamin, R.; Khayatnezhad, M. Assessment of the correlation between chlorophyll content and drought resistance in corn cultivars (Zea Mays). Helix 2020, 10, 93–97. [Google Scholar] [CrossRef]
- Nikolaeva, M.K.; Maevskaya, S.N.; Shugaev, A.G.; Bukhov, N.G. Effect of drought on chlorophyll content and antioxidant enzyme activities in leaves of three wheat cultivars varying in productivity. Russ. J. Plant Physiol. 2010, 57, 87–95. [Google Scholar] [CrossRef]
- Chekol, H.; Bezuayehu, Y.; Warkineh, B.; Shimber, T.; Mierek-Adamska, A.; Dąbrowska, G.B.; Degu, A. Unraveling drought tolerance and sensitivity in coffee genotypes: Insights from seed traits, germination, and growth-physiological responses. Agriculture 2023, 13, 1754. [Google Scholar] [CrossRef]
- Pompelli, M.F.; Barata-Luís, R.; Vitorino, H.S.; Gonçalves, E.R.; Rolim, E.V.; Santos, M.G.; Almeida-Cortez, J.S.; Ferreira, V.M.; Lemos, E.E.; Endres, L. Photosynthesis, photoprotection and antioxidant activity of purging nut under drought deficit and recovery. Biomass Bioenergy 2010, 34, 1207–1215. [Google Scholar] [CrossRef]
- Tamirat, W. Review on role of proline on coffee under drought conditions. J. Environ. Earth Sci. 2019, 9, 1–6. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Basu, S.; Ramegowda, V.; Pereira, A. Mechanisms of drought tolerance in rice. In Achieving Sustainable Cultivation of Rice; Sasaki, T., Ed.; Burleigh Dodds Science Publishing Limited: Sawston, UK, 2016; pp. 131–163. [Google Scholar]
- Panda, D.; Mishra, S.S.; Behera, P.K. Drought tolerance in rice: Focus on recent mechanisms and approaches. Rice Sci. 2021, 28, 119–132. [Google Scholar] [CrossRef]
- Bhise, K.K.; Dandge, P.B. Alleviation of salinity stress in rice plant by encapsulated salt tolerant plant growth promoting bacteria Pantoea agglomerans strain KL and its root colonization ability. Arch. Agron. Soil Sci. 2019, 65, 1955–1968. [Google Scholar] [CrossRef]
- Claussen, W. Proline as a measure of stress in tomato plants. Plant Sci. 2005, 168, 241–248. [Google Scholar] [CrossRef]
- Cavatte, P.C.; Oliveira, Á.A.G.; Morais, L.E.; Martins, S.C.V.; Sanglard, L.M.V.P.; Damatta, F.M. Could shading reduce the negative impacts of drought on coffee? A morphophysiological analysis. Physiol. Plant. 2012, 144, 111–122. [Google Scholar] [CrossRef]
- Barea, J.M.; Pozo, M.J.; Azcón, R.; Azcón-Aguilar, C. Microbial co-operation in the rhizosphere. J. Exp. Bot. 2005, 56, 1761–1778. [Google Scholar] [CrossRef]
- Alemneh, A.A.; Zhou, Y.; Ryder, M.H.; Denton, M.D. Soil environment influences plant growth-promotion traits of isolated rhizobacteria. Pedobiologia 2022, 90, 150785. [Google Scholar] [CrossRef]
- Pantoja-Guerra, M.; Valero-Valero, N.; Ramírez, C.A. Total auxin level in the soil–plant system as a modulating factor for the effectiveness of PGPR inocula: A review. Chem. Biol. Technol. Agric. 2023, 10, 6. [Google Scholar] [CrossRef]
- Martínez, O.A.; Jorquera, M.A.; Crowley, D.E.; de la Mora, M.L. Influence of nitrogen fertilisation on pasture culturable rhizobacteria occurrence and the role of environmental factors on their potential PGPR activities. Biol. Fertil. Soils 2011, 47, 875–885. [Google Scholar] [CrossRef]
- Descroix, F.; Snoeck, J. Environmental factors suitable for coffee cultivation. In Coffee: Growing, Processing, Sustainable Production: A Guidebook for Growers, Processors, Traders, and Researchers; Wintgens, J.N., Ed.; WILEY-VCH Verlag GMBH & Co. KGaA: Weinheim, Germany, 2004; pp. 164–177. ISBN 3527307311. [Google Scholar]
- Muñoz-Villers, L.E.; Geris, J.; Alvarado-Barrientos, M.S.; Holwerda, F.; Dawson, T. Coffee and shade trees show complementary use of soil water in a traditional agroforestry ecosystem. Hydrol. Earth Syst. Sci. 2020, 24, 1649–1668. [Google Scholar] [CrossRef]
- Padovan, M.P.; Cortez, V.J.; Navarrete, L.F.; Navarrete, E.D.; Deffner, A.C.; Centeno, L.G.; Munguía, R.; Barrios, M.; Vílchez-Mendoza, J.S.; Vega-Jarquín, C.; et al. Root distribution and water use in coffee shaded with Tabebuia rosea Bertol. and Simarouba glauca DC. compared to full sun coffee in sub-optimal environmental conditions. Agrofor. Syst. 2015, 89, 857–868. [Google Scholar] [CrossRef]
- Montoani Silva, B.; César de Oliveira, G.; Evaldo Serafim, M.; Eloize Carducci, C.; Andressa da Silva, É.; Martins Barbosa, S.; Beatriz Batista de Melo, L.; Junior Reis dos Santos, W.; Henrique Pereira Reis, T.; Henrique Caputo de Oliveira, C.; et al. Soil management and water-use efficiency in Brazilian coffee crops. In Coffee—Production and Research; IntechOpen: London, UK, 2020. [Google Scholar]
- Clairmont, L.K.; Stevens, K.J.; Slawson, R.M. Site-specific differences in microbial community structure and function within the rhizosphere and rhizoplane of wetland plants is plant species dependent. Rhizosphere 2019, 9, 56–68. [Google Scholar] [CrossRef]
- York, L.M.; Carminati, A.; Mooney, S.J.; Ritz, K.; Bennett, M.J. The holistic rhizosphere: Integrating zones, processes, and semantics in the soil influenced by roots. J. Exp. Bot. 2016, 67, 3629–3643. [Google Scholar] [CrossRef]
- Dworkin, M.; Foster, J.W. Experiments with some microorganisms which utilize ethane and hydrogen. J. Bacteriol. 1958, 75, 592–603. [Google Scholar] [CrossRef]
- Zafar-ul-Hye, M.; Danish, S.; Abbas, M.; Ahmad, M.; Munir, T.M. ACC Deaminase producing PGPR Bacillus amyloliquefaciens and Agrobacterium fabrum along with biochar improve wheat productivity under drought stress. Agronomy 2019, 9, 343. [Google Scholar] [CrossRef]
- Yang, A.; Yen, C. PCR optimization of BOX-A1R PCR for microbial source tracking of Escherichia coli in waterways. J. Exp. Microbiol. Immunol. 2012, 16, 85–89. [Google Scholar]
- Madeira, F.; Madhusoodanan, N.; Lee, J.; Eusebi, A.; Niewielska, A.; Tivey, A.R.N.; Lopez, R.; Butcher, S. The EMBL-EBI Job Dispatcher sequence analysis tools framework in 2024. Nucleic Acids Res. 2024, 52, W521–W525. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of blue agar CAS assay for siderophore detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Jain, D.K.; Patriquin, D.G. Characterization of a substance produced by Azospirillum which causes branching of wheat root hairs. Can. J. Microbiol. 1985, 31, 206–210. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the Salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [PubMed]
- Hara, S.; Morikawa, T.; Wasai, S.; Kasahara, Y.; Koshiba, T.; Yamazaki, K.; Fujiwara, T.; Tokunaga, T.; Minamisawa, K. Identification of nitrogen-fixing Bradyrhizobium associated with roots of field-grown sorghum by metagenome and proteome analyses. Front. Microbiol. 2019, 10, 407. [Google Scholar] [CrossRef]
- Schöllhorn, R.; Burris, R.H. Acetylene as a competitive inhibitor of N-2 fixation. Proc. Natl. Acad. Sci. USA 1967, 58, 213–216. [Google Scholar] [CrossRef]
- Estrada-de los Santos, P.; Bustillos-Cristales, R.; Caballero-Mellado, J. Burkholderia, a genus rich in plant-associated nitrogen fixers with wide environmental and geographic distribution. Appl. Environ. Microbiol. 2001, 67, 2790–2798. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Barrs, H.; Weatherley, P. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413. [Google Scholar] [CrossRef]
- Narayanasamy, S.; Thangappan, S.; Uthandi, S. Plant growth-promoting Bacillus sp. cahoots moisture stress alleviation in rice genotypes by triggering antioxidant defense system. Microbiol. Res. 2020, 239, 126518. [Google Scholar] [CrossRef]
- Seethepalli, A.; Dhakal, K.; Griffiths, M.; Guo, H.; Freschet, G.T.; York, L.M. RhizoVision Explorer: Open-source software for root image analysis and measurement standardization. AoB Plants 2021, 13, plab056. [Google Scholar] [CrossRef]
- Hiscox, J.D.; Israelstam, G.F. A method for the extraction of chlorophyll from leaf tissue without maceration. Can. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Shabnam, N.; Tripathi, I.; Sharmila, P.; Pardha-Saradhi, P. A rapid, ideal, and eco-friendlier protocol for quantifying proline. Protoplasma 2016, 253, 1577–1582. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | ACCd (μmol α-Ketobutyrate/mg Protein/h) | Nitrogen fixation | Iron Solubilization by Siderophores (Halo Diameter in mm) | PSI | IAA Production at 72 h (μg/mL) |
|---|---|---|---|---|---|
| Serratia sp. RCa01 | 1.13 ± 0.31 ef | – | 7.0 ± 0.8 a | 1.6 ± 0.1 e | 1.36 ± 0.25 c |
| Serratia sp. RCa28 | 0.65 ± 0.14 fg | – | 6.8 ± 1.0 ab | 1.4 ± 0.1 e | – |
| Stenotrophomonas sp. RCa07 | 1.83 ± 0.09 de | – | – | – | Not determined |
| Sphingobacterium sp. RCa08 | 0.38 ± 0.14 g | – | – | – | – |
| Sphingobacterium sp. RCa20 | 0.37 ± 0.03 g | – | – | – | – |
| Raoultella sp. RCa12 | 1.22 ± 0.26 ef | – | 6.0 ± 0.0 ab | 1.7 ± 0.2 de | – |
| Chryseobacterium sp. RCa13 | 2.93 ± 0.08 cd | – | – | – | Not determined |
| Pantoea sp. RCa31 | 9.32 ± 0.42 a | – | – | 2.7 ± 0.3 ab | 27.73 ± 2.32 ab |
| Pantoea sp. RCa37 | 2.76 ± 0.27 cd | – | – | 3.2 ± 0.2 a | 25.26 ± 1.30 b |
| Pantoea sp. RCa62 | 5.77 ± 0.15 ab | – | – | 2.3 ± 0.3 bc | 30.23 ± 1.19 a |
| Pseudomonas sp. RCa18 | 9.90 ± 1.07 a | – | 6.5 ± 0.6 ab | 1.6 ± 0.2 e | – |
| Pseudomonas sp. RCa58 | 4.66 ± 0.55 bc | – | 5.5 ± 0.6 b | 2.1 ± 0.1 cd | – |
| Strain | Primary Growth (cm) | Relative Primary Growth (%) | Secondary Growth (cm) | Relative Secondary Growth (%) | ||||
|---|---|---|---|---|---|---|---|---|
| R | S | R | S | R | S | R | S | |
| Control | 2.4 ± 0.3 abcde | 0.9 ± 0.3 fgh | 6.16 ± 1.38 abcd | 2.26 ± 0.62 fg | 0.05 ± 0.03 ab | 0.01 ± 0.04 abcdefg | 7.08 ± 4.32 ab | 1.55 ± 6.39 abcdef |
| Serratia sp. RCa01 | 2.5 ± 0.9 abcd | 1.0 ± 1.2 efgh | 6.04 ± 2.10 abcde | 2.59 ± 3.05 efg | 0.03 ± 0.02 abcde | 0.05 ± 0.08 a | 4.38 ± 3.16 abcde | 8.61 ± 13.85 a |
| Serratia sp. RCa28 | 3.1 ± 0.8 ab | 1.7 ± 1.1 bcdefg | 7.26 ± 2.27 ab | 4.11 ± 2.53 bcdefg | 0.03 ± 0.0 abc | 0.01 ± 0.03 abcdefg | 4.83 ± 4.23 abcd | 1.73 ± 4.33 abcdef |
| Stenotrophomonas sp. RCa07 | 2.4 ± 1.8 abcd | 1.2 ± 0.8 defgh | 6.09 ± 4.37 abcd | 3.20 ± 2.10 defg | 0.04 ± 0.03 abc | −0.01 ± 0.05 cdefg | 5.94 ± 4.59 abc | −1.06 ± 7.17 cdef |
| Sphingobacterium sp. RCa08 | 2.2 ± 0.9 abcdef | 1.3 ± 0.5 defgh | 5.56 ± 1.97 abcdef | 3.50 ± 1.93 cdefg | 0.02 ± 0.03 abcdefg | 0.01 ± 0.03 abcdefg | 3.35 ± 4.4 abcdef | 1.62 ± 4.61 abcdef |
| Sphingobacterium sp. RCa20 | 1.8 ± 1.1 bcdefg | 1.6 ± 1.4 cdefgh | 4.54 ± 3.05 bcdefg | 3.99 ± 3.26 bcdefg | 0.03 ± 0.02 abcd | −0.02 ± 0.03 defg | 4.52 ± 3.22 abcde | −3.18 ± 6.21 ef |
| Raoultella sp. RCa12 | 2.2 ± 1.7 abcdef | 0.7 ± 1.4 gh | 5.66 ± 4.35 abcdef | 1.64 ± 3.30 g | 0.03 ± 0.03 abc | −0.02 ± 0.05 efg | 5.82 ± 4.51 abc | −2.51 ± 8.15 def |
| Chryseobacterium sp. RCa13 | 1.4 ± 0.8 cdefgh | 0.7 ± 0.6 gh | 3.50 ± 1.84 cdefg | 2.03 ± 1.73 g | 0.04 ± 0.04 ab | −0.02 ± 0.03 fg | 6.75 ± 5.9 abc | −3 ± 4.76 def |
| Pantoea sp. RCa31 | 2.7 ± 0.5 abc | 0.5 ± 0.6 gh | 6.24 ± 1.21 abcd | 1.59 ± 2.10 g | 0.02 ± 0.01 abcdefg | 0.02 ± 0.02 abcdef | 3 ± 2.31 abcdef | 3.72 ± 2.45 abcdef |
| Pantoea sp. RCa37 | 2.2 ± 0.3 abcdef | 0.4 ± 1.4 h | 5.79 ± 1.09 abcde | 1.26 ± 3.62 g | 0.02 ± 0.03 abcdefg | 0 ± 0.03 bcdefg | 4.28 ± 7.44 abcdef | 0.19 ± 6.04 bcdef |
| Pantoea sp. RCa62 | 2.6 ± 0.6 abc | 3.1 ± 0.8 ab ** | 6.84 ± 2.14 abc | 7.19 ± 1.12 ab ** | 0.05 ± 0.03 ab | 0.01 ± 0.01 abcdefg | 7.69 ± 5.8 ab | 1.24 ± 2.16 abcdef |
| Pseudomonas sp. RCa18 | 2.7 ± 0.7 abc | 1.4 ± 0.8 cdefgh | 7.20 ± 1.66 ab | 3.39 ± 2.05 cdefg | 0.03 ± 0.03 abcde | −0.03 ± 0.03 g | 4.15 ± 4.62 abcdef | −3.57 ± 4.75 f |
| Pseudomonas sp. RCa58 | 3.3 ± 1.0 a | 2.1 ± 0.8 abcdef | 8.74 ± 2.84 a | 5.70 ± 2.98 abcdef | 0.03 ± 0.02 abcdef | 0.03 ± 0.03 abcdef | 4.22 ± 2.71 abcdef | 4.05 ± 4.58 abcdef |
| Total mean | 2.4 ± 0.23 A | 1.3 ± 0.25 B | 6.12 ± 0.59 A | 3.26 ± 0.63 B | 0.03 ± 0.01 A | 0.004 ± 0.01 B | 5.08 ± 1.01 A | 0.72 ± 1.53 B |
| Strain | Dry Weight of the Shoot (g) | Dry Weight of the Root (g) | Total Dry Weight (g) | |||
|---|---|---|---|---|---|---|
| R | S | R | S | R | S | |
| Control | 10.4 ± 2.84 bcd | 8.93 ± 1.95 d | 5.29 ± 1.93 b | 6.89 ± 2.29 b | 15.68 ± 4.65 | 15.83 ± 4.04 |
| Serratia sp. RCa01 | 9.68 ± 2.57 bcd | 10.76 ± 3.37 abcd | 5.01 ± 1.65 b | 6.77 ± 3.34 b | 14.69 ± 4.08 | 17.54 ± 6.61 |
| Serratia sp. RCa28 | 13.18 ± 3.26 abc | 11.98 ± 6.3 abcd | 7.65 ± 2.82 ab | 7.36 ± 3.87 b | 20.83 ± 6.02 | 19.33 ± 10.16 |
| Stenotrophomonas sp. RCa07 | 10.93 ± 3.3 abcd | 11.11 ± 1.73 abcd | 5.75 ± 2.5 b | 6.83 ± 0.58 b | 16.67 ± 5.57 | 17.94 ± 1.86 |
| Sphingobacterium sp. RCa08 | 11.17 ± 2.33 abcd | 9.02 ± 1.39 cd | 6.52 ± 1.93 b | 6.65 ± 1.77 b | 17.69 ± 4.15 | 15.67 ± 3.12 |
| Sphingobacterium sp. RCa20 | 10.9 ± 3.33 abcd | 10.75 ± 2.56 abcd | 5.58 ± 2.39 b | 6.27 ± 2.23 b | 16.47 ± 5.44 | 17.02 ± 4.57 |
| Raoultella sp. RCa12 | 9.9 ± 1.87 bcd | 8.71 ± 1.65 d | 6 ± 2.17 b | 6.2 ± 1.57 b | 15.91 ± 3.98 | 14.91 ± 3.01 |
| Chryseobacterium sp. RCa13 | 10.51 ± 2.73 abcd | 8.97 ± 2.13 d | 5.47 ± 1.47 b | 5.2 ± 1.89 b | 15.98 ± 3.54 | 14.17 ± 3.46 |
| Pantoea sp. RCa31 | 13.75 ± 3.95 ab | 10.19 ± 2.39 bcd | 6.8 ± 2.56 b | 5.76 ± 1.56 b | 20.55 ± 5.95 | 15.95 ± 3.71 |
| Pantoea sp. RCa37 | 9.95 ± 3.06 bcd | 8.45 ± 1.68 d | 4.68 ± 2.13 b | 6.32 ± 2.13 b | 14.63 ± 5.06 | 14.77 ± 3.63 |
| Pantoea sp. RCa62 | 11.14 ± 2.47 abcd | 14.9 ± 3.92 a ** | 5.6 ± 1.56 b | 10.57 ± 3.25 a * | 16.75 ± 3.32 | 25.48 ± 7.02 |
| Pseudomonas sp. RCa18 | 10.02 ± 4.01 bcd | 11.15 ± 2.47 abcd | 6.05 ± 4.01 b | 7.04 ± 1.05 b | 16.07 ± 7.93 | 18.19 ± 3.21 |
| Pseudomonas sp. RCa58 | 11.04 ± 1.88 abcd | 10.66 ± 3.03 abcd | 5.49 ± 1.77 b | 5.95 ± 1.65 b | 16.52 ± 3.13 | 16.62 ± 4.56 |
| Total mean | 10.97 ± 0.67 | 10.43 ± 0.71 | 5.84 ± 0.51 B | 6.76 ± 0.54 A | 16.80 ± 1.12 | 17.19 ± 1.20 |
| Strain | Number of Leaf Buds | Fresh Weight of the Leaf Buds (g) | Dry Weight of the Leaf Buds (g) | |||
|---|---|---|---|---|---|---|
| R | S | R | S | R | S | |
| Control | 3.9 ± 1.6 abcd | 2.3 ± 2.4 bcd | 1.54 ± 1.32 abcd | 0.25 ± 0.38 g | 0.40 ± 0.33 abcd | 0.08 ± 0.11 f |
| Serratia sp. RCa01 | 3.7 ± 1.8 abcd | 3.0 ± 2.1 bcd | 1.41 ± 0.59 abcde | 0.90 ± 0.66 bcdefg | 0.39 ± 0.17 abcd | 0.27 ± 0.21 bcdef |
| Serratia sp. RCa28 | 4.1 ± 1.9 abcd | 4.6 ± 3.6 abc | 1.57 ± 0.57 abc | 0.85 ± 0.58 bcdefg | 0.35 ± 0.16 abcde | 0.29 ± 0.21 bcdef |
| Stenotrophomonas sp. RCa07 | 2.7 ± 1.1 bcd | 2.1 ± 2.0 cd | 1.05 ± 0.57 bcdefg | 0.45 ± 0.52 fg | 0.27 ± 0.16 bcdef | 0.15 ± 0.18 def |
| Sphingobacterium sp. RCa08 | 3.0 ± 1.2 bcd | 2.4 ± 1.7 abcd | 0.95 ± 0.24 bcdefg | 0.55 ± 0.54 efg | 0.26 ± 0.08 bcdef | 0.18 ± 0.16 def |
| Sphingobacterium sp. RCa20 | 3.0 ± 1.3 bcd | 2.4 ± 1.5 bcd | 1.41 ± 1.04 abcde | 0.34 ± 0.36 g | 0.37 ± 0.29 abcd | 0.10 ± 0.11 ef |
| Raoultella sp. RCa12 | 4.0 ± 1.8 abcd | 2.9 ± 0.9 bcd | 1.40 ± 0.89 abcde | 0.51 ± 0.36 efg | 0.35 ± 0.22 abcde | 0.17 ± 0.13 def |
| Chryseobacterium sp. RCa13 | 3.9 ± 1.3 abcd | 3.1 ± 2.2 bcd | 1.15 ± 0.55 bcdefg | 0.64 ± 0.49 cdefg | 0.30 ± 0.14 bcdef | 0.19 ± 0.14 cdef |
| Pantoea sp. RCa31 | 5.9 ± 2.0 a | 2.4 ± 1.9 bcd | 2.17 ± 1.51 a | 0.64 ± 0.56 cdefg | 0.57 ± 0.38 a | 0.19 ± 0.16 cdef |
| Pantoea sp. RCa37 | 4.0 ± 1.6 abcd | 2.0 ± 1.2 d | 1.36 ± 0.86 abcdef | 0.28 ± 0.20 g | 0.36 ± 0.18 abcde | 0.09 ± 0.07 f |
| Pantoea sp. RCa62 | 3.7 ± 1.3 abcd | 4.7 ± 3.3 ab | 1.72 ± 0.87 ab | 1.57 ± 1.15 abc *** | 0.45 ± 0.19 ab | 0.45 ± 0.33 ab ** |
| Pseudomonas sp. RCa18 | 3.0 ± 1.4 bcd | 2.3 ± 1.8 bcd | 0.90 ± 0.57 bcdefg | 0.36 ± 0.37 g | 0.24 ± 0.16 bcdef | 0.11 ± 0.11 ef |
| Pseudomonas sp. RCa58 | 3.9 ± 0.7 abcd | 2.6 ± 2.2 bcd | 1.68 ± 0.78 ab | 0.63 ± 0.70 defg | 0.44 ± 0.22 abc | 0.18 ± 0.19 def |
| Total mean | 3.8 ± 0.3 A | 2.8 ± 0.5 B | 1.41 ± 0.18 A | 0.61 ± 0.13 B | 0.37 ± 0.05 A | 0.19 ± 0.04 B |
| Strain | Total Root Length (m) | Mean Root Diameter (mm) | Total Root Surface (cm2) | Total Root Volume (cm3) | Dry Shoot-to-Root Ratio | Total Root Length (m) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| R | S | R | S | R | S | R | S | R | S | |
| Control | 66.68 ± 37.68 b | 100.65 ± 43.84 ab | 0.66 ± 0.06 abc | 0.64 ± 0.05 bc | 1315 ± 629 b | 1936 ± 775 ab | 73 ± 34 | 86 ± 17 | 2.05 ± 0.33 ab | 1.39 ± 0.38 f |
| Serratia sp. RCa01 | 55.32 ± 33.87 b | 101.05 ± 45.29 ab | 0.68 ± 0.07 abc | 0.63 ± 0.08 c | 1119 ± 588 b | 1924 ± 826 ab | 55 ± 20 | 81 ± 30 | 2.02 ± 0.32 ab | 1.73 ± 0.35 abcdef |
| Serratia sp. RCa28 | 114.81 ± 70.23 ab | 81.19 ± 33.65 ab | 0.66 ± 0.08 abc | 0.69 ± 0.04 abc | 2164 ± 1152 ab | 1706 ± 737 ab | 100 ±37 | 78 ± 26 | 1.81 ± 0.27 abcde | 1.63 ± 0.19 abcdef |
| Stenotrophomonas sp. RCa07 | 70.45 ± 40.07 b | 84.18 ± 23.76 ab | 0.68 ± 0.07 abc | 0.68 ± 0.05 abc | 1414 ± 743 b | 1748 ± 383 ab | 69 ± 33 | 81 ± 8 | 2.16 ± 0.68 ab | 1.64 ± 0.28 abcdef |
| Sphingobacterium sp. RCa08 | 106.61 ± 51.81 ab | 90.68 ± 28.29 ab | 0.65 ± 0.11 bc | 0.65 ± 0.05 abc | 1972 ± 892 ab | 1792 ± 503 ab | 87 ± 35 | 75 ± 10 | 1.86 ± 0.5 abcde | 1.44 ± 0.32 def |
| Sphingobacterium sp. RCa20 | 69.04 ± 51.97 b | 75.76 ± 36.21 ab | 0.71 ± 0.09 abc | 0.69 ± 0.08 abc | 1375 ± 895 b | 1531 ± 588 ab | 72 ± 38 | 71 ± 14 | 2.09 ± 0.4 ab | 1.82 ± 0.44 abcde |
| Raoultella sp. RCa12 | 82.67 ± 41.45 ab | 80.77 ± 17.37 ab | 0.65 ± 0.05 abc | 0.64 ± 0.03 bc | 1624 ± 736 ab | 1633 ± 334 ab | 72 ± 34 | 74 ± 10 | 1.78 ± 0.36 abcdef | 1.45 ± 0.33 cdef |
| Chryseobacterium sp. RCa13 | 53.34 ± 32.82 b | 65.23 ± 28.73 b | 0.77 ± 0.12 a | 0.68 ± 0.04 abc | 1116 ± 538 b | 1342 ± 569 b | 66 ± 17 | 62 ± 21 | 2.00 ± 0.47 abc | 1.98 ± 0.87 abcdef |
| Pantoea sp. RCa31 | 98.77 ± 61.18 ab | 70.1 ± 25.61 ab | 0.63 ± 0.06 c | 0.66 ± 0.02 abc | 1869 ± 1020 ab | 1423 ± 505 b | 90 ± 44 | 65 ± 20 | 2.19 ± 0.75 ab | 1.82 ± 0.33 abcde |
| Pantoea sp. RCa37 | 56.36 ± 42.95 b | 92.92 ± 30.04 ab | 0.74 ± 0.12 abc | 0.63 ± 0.08 c | 1162 ± 805 b | 1774 ± 580 ab | 58 ± 25 | 71 ± 27 | 2.37 ± 0.59 a | 1.41 ± 0.3 ef |
| Pantoea sp. RCa62 | 74.42 ± 41.2 ab | 138.89 ± 59.93 a | 0.66 ± 0.07 abc | 0.65 ± 0.06 bc | 1412 ± 659 b | 2684 ± 971 a | 64 ± 23 | 109 ± 33 | 2.15 ± 0.65 ab | 1.45 ± 0.24 cdef |
| Pseudomonas sp. RCa18 | 77.97 ± 83.26 b | 81.04 ± 12.67 ab | 0.75 ± 0.12 ab | 0.63 ± 0.05 c | 1477 ± 1416 b | 1589 ± 177 ab | 76 ± 59 | 68 ± 8 | 1.94 ± 0.5 abcd | 1.59 ± 0.29 bcdef |
| Pseudomonas sp. RCa58 | 85.48 ± 59.29 ab | 83.88 ± 31.86 ab | 0.64 ± 0.06 bc | 0.64 ± 0.06 bc | 1600 ± 937 ab | 1615 ± 462 ab | 76 ± 37 | 75 ± 19 | 2.15 ± 0.52 ab | 1.81 ± 0.26 abcde |
| Total mean | 77.84 ± 11.56 B | 88.18 ± 8.09 A | 0.68 ± 0.02 A | 0.65 ± 0.01 B | 1509.22± 195.86 B | 1745.98 ± 143.63 A | 73.63 ± 7.79 | 76.40 ± 4.88 | 2.04 ± 0.11 A | 1.63 ± 0.09 B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasso-Arreola, Y.; Ibarra, J.A.; Rosas-Cárdenas, F.d.F.; Estrada-de los Santos, P. Beneficial Effects of ACC Deaminase-Producing Rhizobacteria on the Drought Stress Resistance of Coffea arabica L. Plants 2025, 14, 1084. https://doi.org/10.3390/plants14071084
Jasso-Arreola Y, Ibarra JA, Rosas-Cárdenas FdF, Estrada-de los Santos P. Beneficial Effects of ACC Deaminase-Producing Rhizobacteria on the Drought Stress Resistance of Coffea arabica L. Plants. 2025; 14(7):1084. https://doi.org/10.3390/plants14071084
Chicago/Turabian StyleJasso-Arreola, Yesenia, J. Antonio Ibarra, Flor de Fátima Rosas-Cárdenas, and Paulina Estrada-de los Santos. 2025. "Beneficial Effects of ACC Deaminase-Producing Rhizobacteria on the Drought Stress Resistance of Coffea arabica L." Plants 14, no. 7: 1084. https://doi.org/10.3390/plants14071084
APA StyleJasso-Arreola, Y., Ibarra, J. A., Rosas-Cárdenas, F. d. F., & Estrada-de los Santos, P. (2025). Beneficial Effects of ACC Deaminase-Producing Rhizobacteria on the Drought Stress Resistance of Coffea arabica L. Plants, 14(7), 1084. https://doi.org/10.3390/plants14071084