
MaxEnt-Based Distribution Modeling of the Invasive Species Phragmites australis Under Climate Change Conditions in Iraq


Abstract
1. Introduction
2. Results
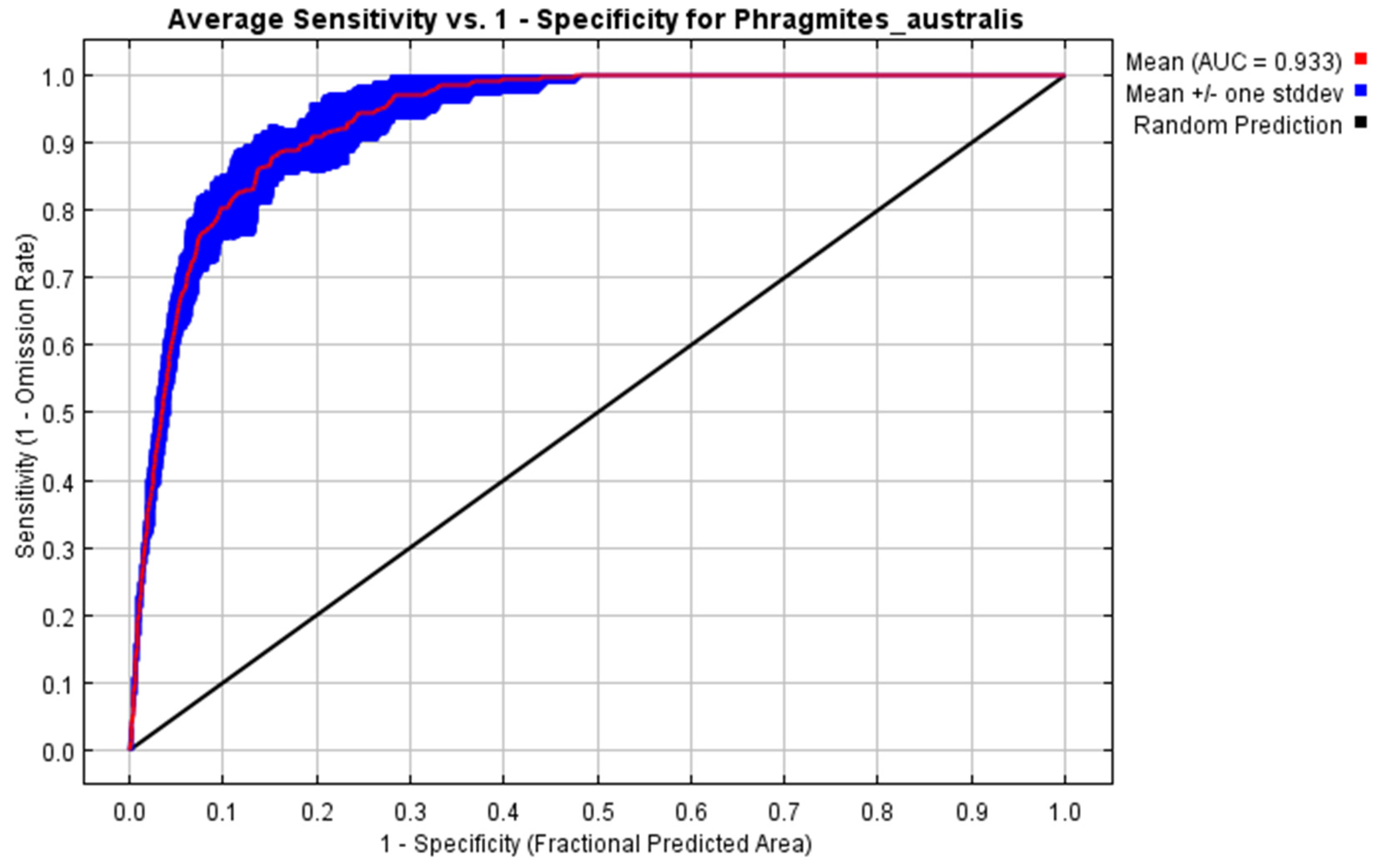
2.1. Model Performance
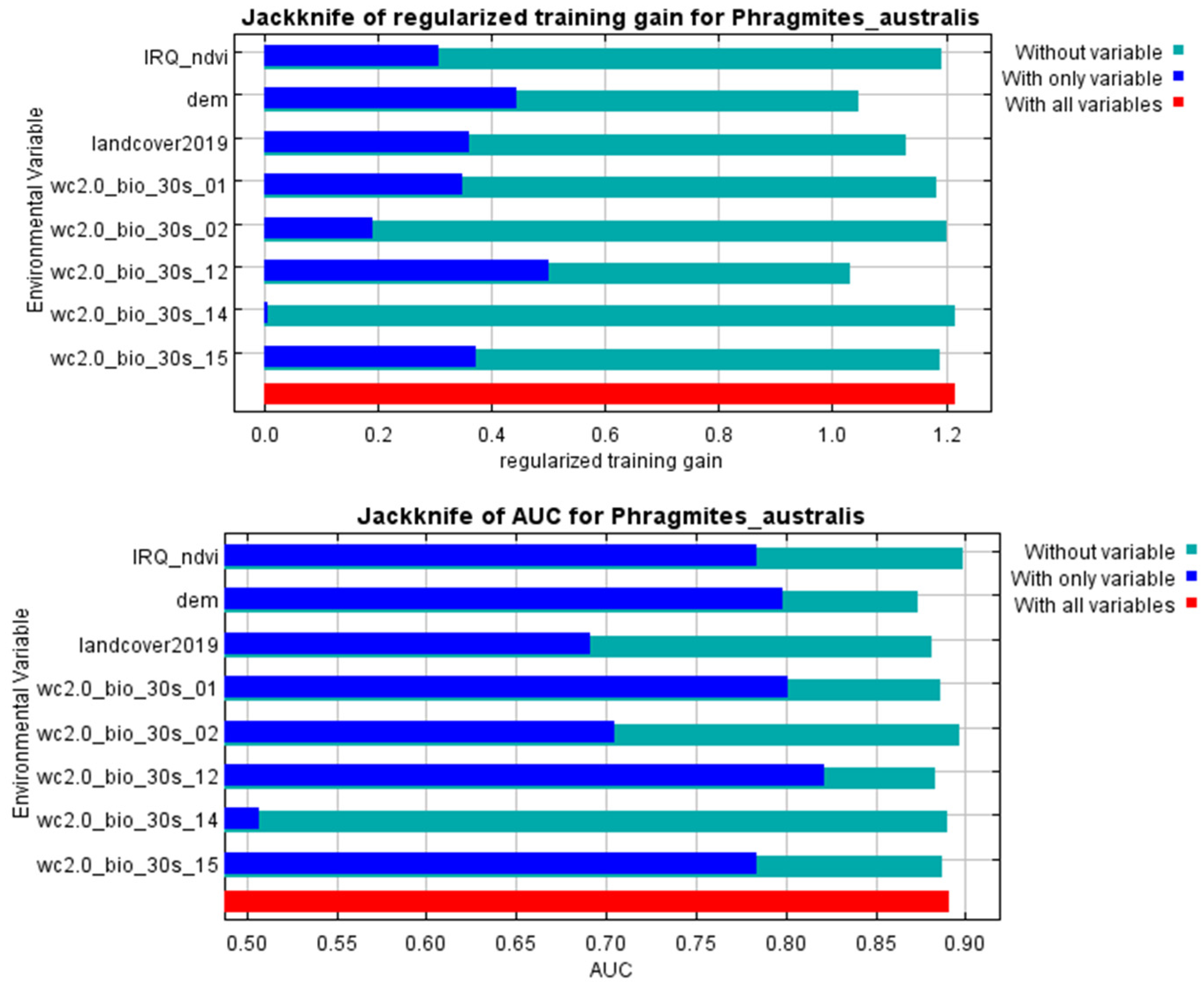
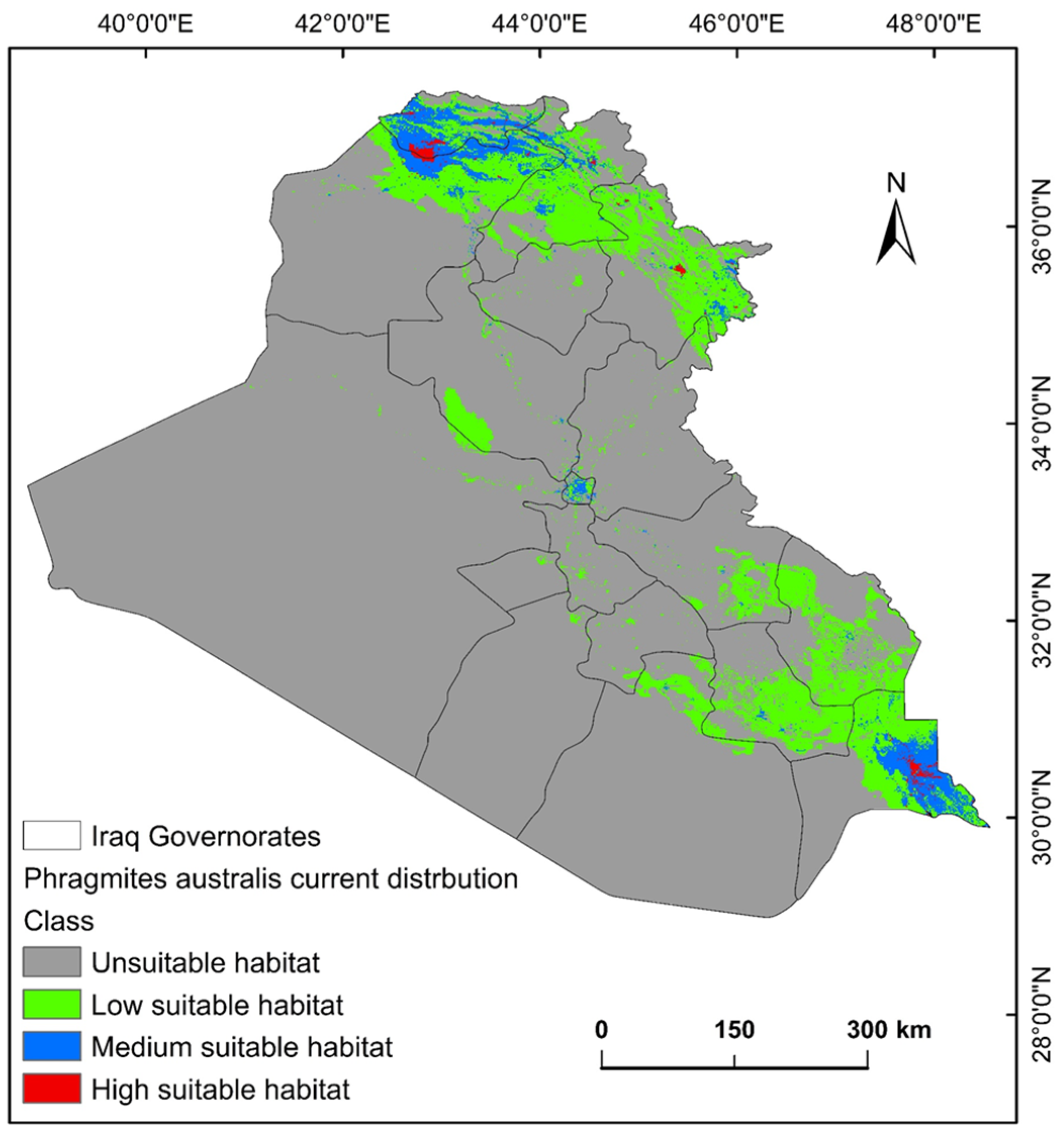
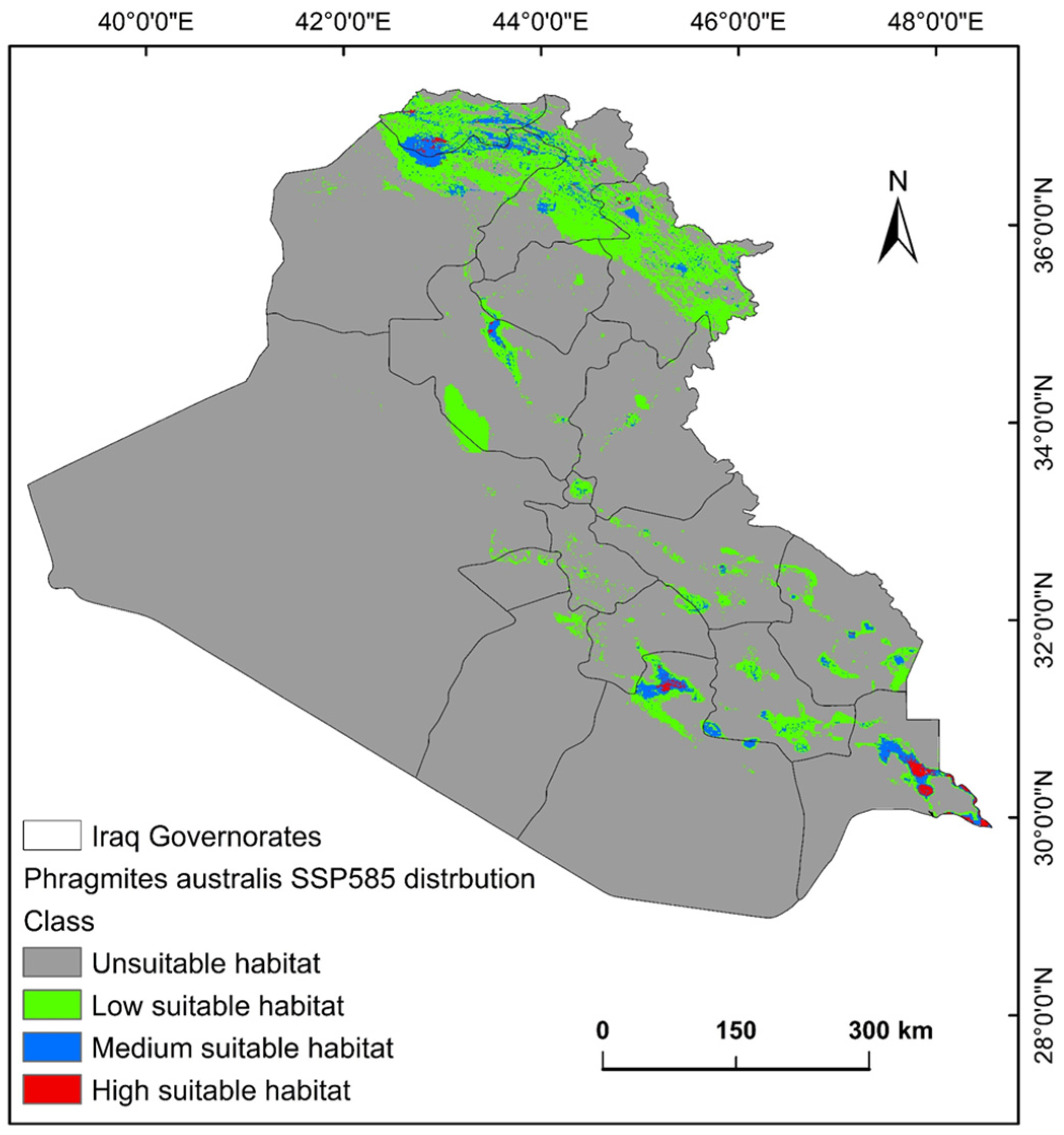
2.2. Suitability Distribution of P. australis and Conditioning Factors
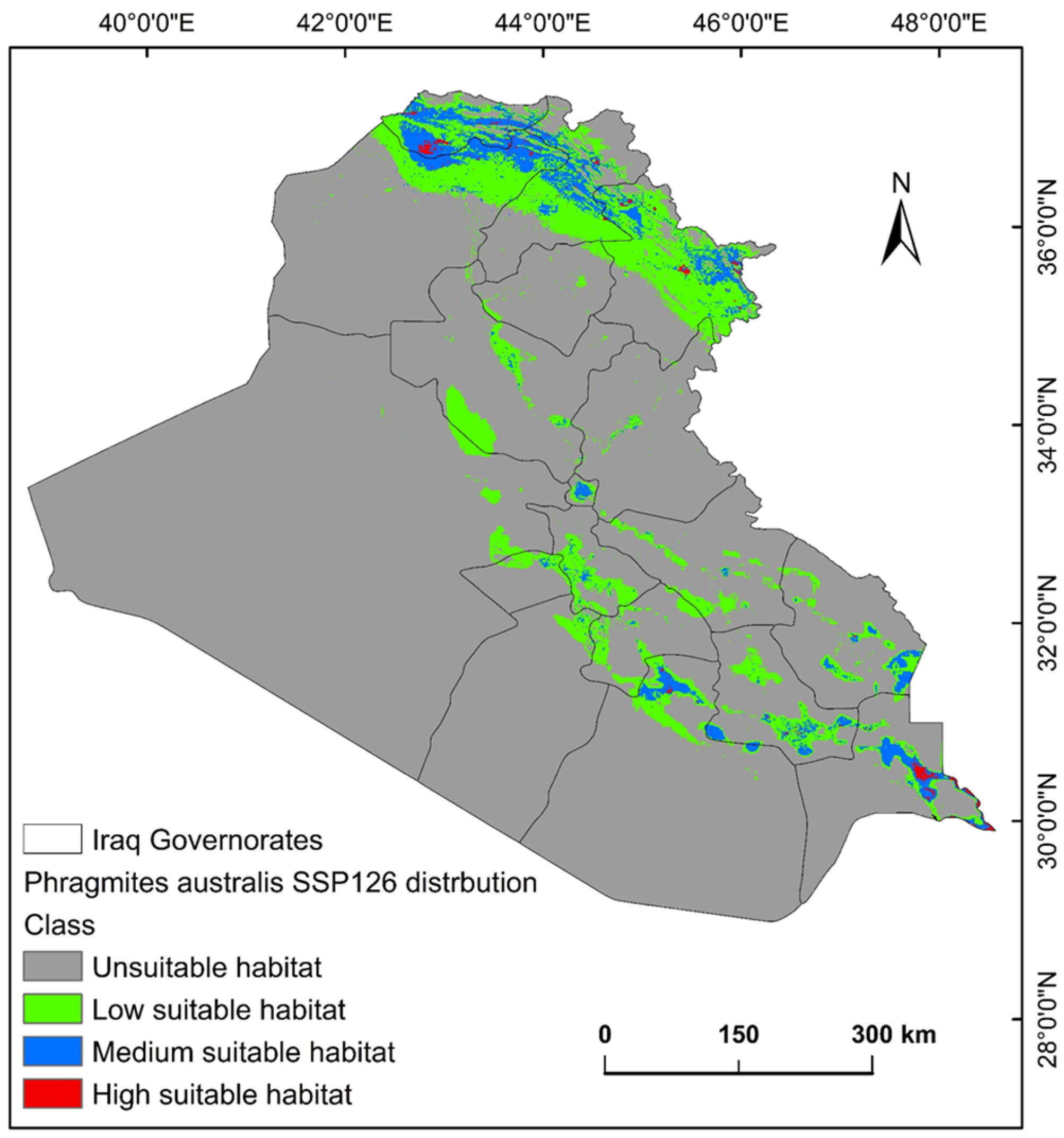
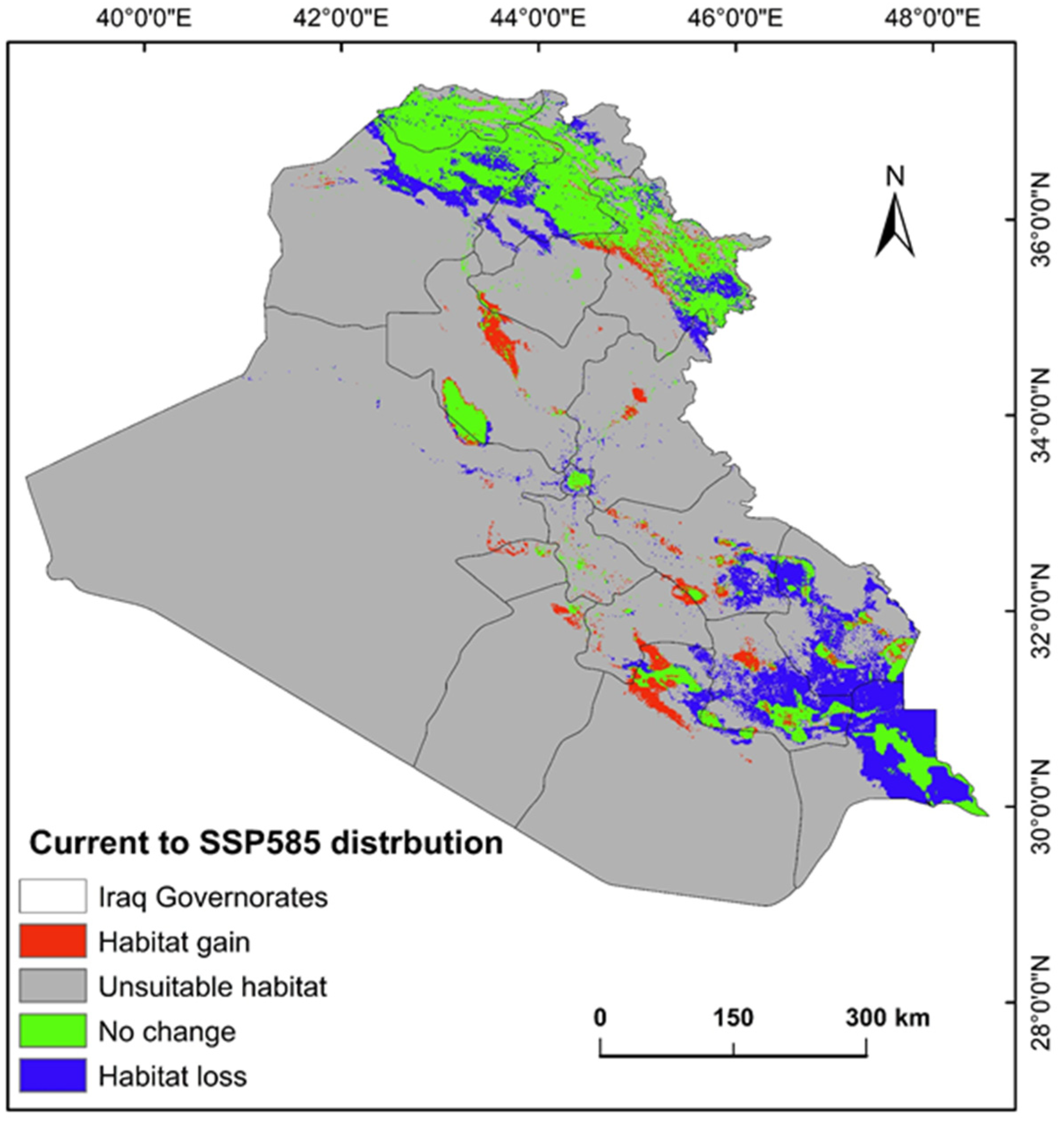
2.3. Shifts in Habitat Distribution of P. australis over Time in Iraq
3. Discussion
3.1. Distribution of P. australis in Iraq
3.2. Habitat Loss and Gain and Climate-Driven Redistribution
3.3. Implications for Biodiversity and Limitations
4. Materials and Methods
4.1. Study Area
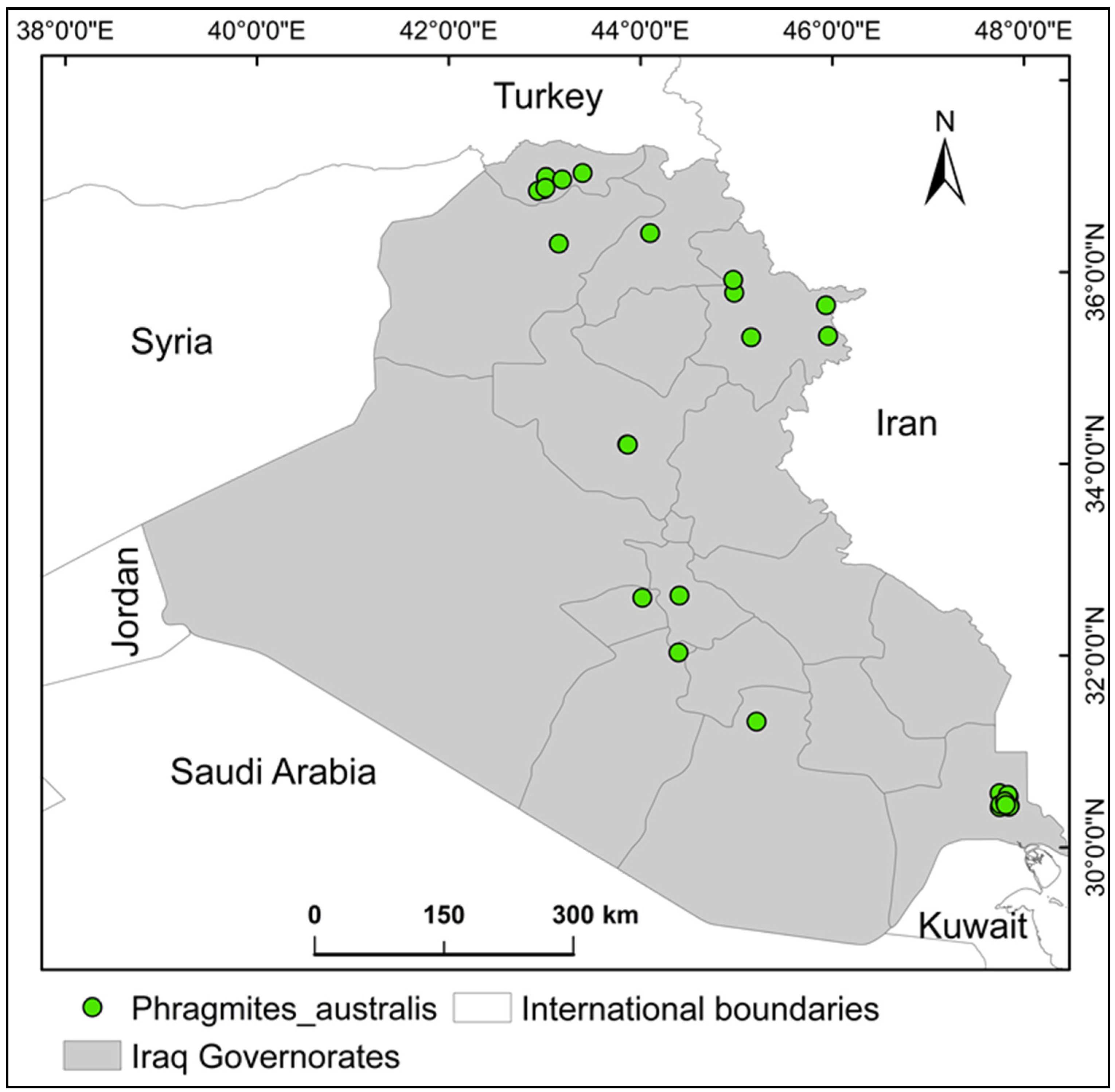
4.2. Phragmites Australis Occurrence Records
4.3. Conditioning Factors
4.4. MaxEnt Model
4.5. Model Evaluation
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pyšek, P.; Richardson, D.M. The biogeography of naturalization in alien plants. J. Biogeogr. 2006, 33, 2040–2050. [Google Scholar] [CrossRef]
- Chen, B.; Sun, Y.; Han, Z.; Huang, H.; Zhang, H.; Li, Y.; Zhang, G.; Liu, W. Challenges in preventing and controlling invasive alien species in China. J. Biosaf 2020, 29, 157–163. [Google Scholar]
- Richardson, D.M.; Pyšek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- O’Connor, B.; Bojinski, S.; Röösli, C.; Schaepman, M. Monitoring global changes in biodiversity and climate is more important than ever as ecological crisis intensifies. Ecol. Inform. 2019, 55, 101033. [Google Scholar] [CrossRef]
- Lee, H.; Calvin, K.; Dasgupta, D.; Krinner, G.; Mukherji, A.; Thorne, P.; Trisos, C.; Romero, J.; Aldunce, P.; Barrett, K. Climate Change 2023: Synthesis Report. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Lee, H., Romero, J., Eds.; IPCC: Geneva, Switzerland, 2023. [Google Scholar]
- Hassan, W.H.; Nile, B.K. Climate change and predicting future temperature in Iraq using CanESM2 and HadCM3 modeling. Model. Earth Syst. Environ. 2021, 7, 737–748. [Google Scholar] [CrossRef]
- Salman, S.A.; Shahid, S.; Ismail, T.; Ahmed, K.; Wang, X.-J. Selection of climate models for projection of spatiotemporal changes in temperature of Iraq with uncertainties. Atmos. Res. 2018, 213, 509–522. [Google Scholar] [CrossRef]
- Mansouri Daneshvar, M.R.; Ebrahimi, M.; Nejadsoleymani, H. An overview of climate change in Iran: Facts and statistics. Environ. Syst. Res. 2019, 8, 1–10. [Google Scholar] [CrossRef]
- Demircan, M.; Gürkan, H.; Eskioğlu, O.; Arabaci, H.; Coşkun, M. Climate change projections for Turkey: Three models and two scenarios. Turk. J. Water Sci. Manag. 2017, 1, 22–43. [Google Scholar] [CrossRef]
- Lu, X.; Siemann, E.; Shao, X.; Wei, H.; Ding, J. Climate warming affects biological invasions by shifting interactions of plants and herbivores. Glob. Chang. Biol. 2013, 19, 2339–2347. [Google Scholar] [CrossRef]
- Dukes, J.S.; Chiariello, N.R.; Loarie, S.R.; Field, C.B. Strong response of an invasive plant species (Centaurea solstitialis L.) to global environmental changes. Ecol. Appl. 2011, 21, 1887–1894. [Google Scholar] [CrossRef]
- Peterson, A.T.; Stewart, A.; Mohamed, K.I.; Araújo, M.B. Shifting global invasive potential of European plants with climate change. PLoS ONE 2008, 3, e2441. [Google Scholar] [CrossRef] [PubMed]
- Awchi, T.A.; Kalyana, M.M. Meteorological drought analysis in northern Iraq using SPI and GIS. Sustain. Water Resour. Manag. 2017, 3, 451–463. [Google Scholar] [CrossRef]
- Khwarahm, N.R. Modeling forest-shrubland fire susceptibility based on machine learning and geospatial approaches in mountains of Kurdistan Region, Iraq. Arab. J. Geosci. 2022, 15, 1184. [Google Scholar] [CrossRef]
- Khwarahm, N.R.; Najmaddin, P.M.; Ararat, K.; Qader, S. Past and future prediction of land cover land use change based on earth observation data by the CA–Markov model: A case study from Duhok governorate, Iraq. Arab. J. Geosci. 2021, 14, 1–14. [Google Scholar] [CrossRef]
- Khwarahm, N.R.; Qader, S.; Ararat, K.; Fadhil Al-Quraishi, A.M. Predicting and mapping land cover/land use changes in Erbil/Iraq using CA-Markov synergy model. Earth Sci. Inform. 2021, 14, 393–406. [Google Scholar] [CrossRef]
- Khwarahm, N.R. Mapping current and potential future distributions of the oak tree (Quercus aegilops) in the Kurdistan Region, Iraq. Ecol. Process. 2020, 9, 1–16. [Google Scholar] [CrossRef]
- Hama, A.A.; Khwarahm, N.R. Predictive mapping of two endemic oak tree species under climate change scenarios in a semiarid region: Range overlap and implications for conservation. Ecol. Inform. 2023, 73, 101930. [Google Scholar] [CrossRef]
- Khwarahm, N.R. Predicting the spatial distribution of Hyalomma ssp., vector ticks of Crimean–Congo Haemorrhagic fever in Iraq. Sustainability 2023, 15, 13669. [Google Scholar] [CrossRef]
- Brix, H.; Cížková, H. Introduction: Phragmites-dominated wetlands, their functions and sustainable use. Aquat. Bot. 2001, 69, 87–88. [Google Scholar] [CrossRef]
- Ahmad, S.A.; Ahmad, N.S.; Salih, S. Invasive alien species in Iraq. In Invasive Alien Species: Observations and Issues from Around the World; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2021; Volume 2, pp. 126–150. [Google Scholar]
- Al-Mayah, A.-R.A.; Al-Abbawy, D.A.; Al-Assadi, W.M.; Al-Saadi, S.A.; Al-Edany, T.Y. Status of aquatic macrophytes in Saffia nature reserve, South of Huwaiza Marsh, Iraq. Marsh Bull. 2012, 7, 1–16. [Google Scholar]
- Bedair, H.; Al-Saad, H.; Salman, N. Iraq’s southern marshes something special to be conserved; A Case Study. Marsh Bull. 2006, 2, 99–126. [Google Scholar]
- Rohal, C.B.; Cranney, C.; Hazelton, E.L.; Kettenring, K.M. Invasive Phragmites australis management outcomes and native plant recovery are context dependent. Ecol. Evol. 2019, 9, 13835–13849. [Google Scholar] [CrossRef] [PubMed]
- Kiviat, E. Ecosystem services of Phragmites in North America with emphasis on habitat functions. AoB Plants 2013, 5, plt008. [Google Scholar] [CrossRef]
- Kandoh, A.A.; Abed, S.A. Measurement the Heavy Elements of Phragmites australis and Syllibium marianum in Hor Al-Dalmaj, Southern Iraq. IOP Conf. Ser. Earth Environ. Sci. 2021, 790, 012034. [Google Scholar] [CrossRef]
- salman obaid AL-khazraji, S.; Al-Fatlawy, H.J.J. The Concentration of Some Heavy Metals in Phragmites Australis and Ceratophyllum. Demersum Plants in the Euphrates River, Iraq. Pak. J. Life Soc. Sci. 2024, 22, 2881–2903. [Google Scholar]
- Al-Iraqi, A.R.; Gandhi, B.P.; Folkard, A.M.; Barker, P.A.; Semple, K.T. Determine the optimal parameters for biogas production from common reed (Phragmites australis). BioEnergy Res. 2024, 17, 1302–1314. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Eller, F.; Lambertini, C.; Nguyen, L.X.; Brix, H. Increased invasive potential of non-native Phragmites australis: Elevated CO2 and temperature alleviate salinity effects on photosynthesis and growth. Glob. Chang. Biol. 2014, 20, 531–543. [Google Scholar] [CrossRef]
- Eller, F.; Skálová, H.; Caplan, J.S.; Bhattarai, G.P.; Burger, M.K.; Cronin, J.T.; Guo, W.-Y.; Guo, X.; Hazelton, E.L.; Kettenring, K.M. Cosmopolitan species as models for ecophysiological responses to global change: The common reed Phragmites australis. Front. Plant Sci. 2017, 8, 1833. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Lee, S. Principal bioclimatic variables of ten dominant plant species in Korea wetland using the Maxent model. Ecol. Eng. 2022, 183, 106729. [Google Scholar] [CrossRef]
- Guo, W.Y.; Lambertini, C.; Li, X.Z.; Meyerson, L.A.; Brix, H. Invasion of Old World Phragmites australis in the New World: Precipitation and temperature patterns combined with human influences redesign the invasive niche. Glob. Chang. Biol. 2013, 19, 3406–3422. [Google Scholar] [CrossRef]
- McCormick, M.K.; Whigham, D.F.; Stapp, J.R.; Hazelton, E.L.; McFarland, E.K.; Kettenring, K.M. Shoreline modification affects recruitment of invasive Phragmites australis. Wetl. Ecol. Manag. 2020, 28, 909–919. [Google Scholar] [CrossRef]
- Ward, M.J.M.; Degazio, T.; Bowman, J. Great Lakes coastal wetland plant biodiversity increases following the manual removal of invasive Phragmites australis. Wetl. Ecol. Manag. 2025, 33, 2. [Google Scholar] [CrossRef]
- Wang, C.; Liu, L.; Yin, M.; Liu, B.; Wu, Y.; Eller, F.; Gao, Y.; Brix, H.; Wang, T.; Guo, W. Chromosome-level genome assemblies reveal genome evolution of an invasive plant Phragmites australis. Commun. Biol. 2024, 7, 1007. [Google Scholar] [CrossRef]
- Blizzard, C. Reexamining Ecological Roles and Management Methods of Phragmites australis. Master’s Thesis, Johns Hopkins University, Baltimore, MD, USA, 2023. [Google Scholar]
- Nasser, M. Forests and forestry in Iraq: Prospects and limitations. Commonw. For. Rev. 1984, 63, 299–304. [Google Scholar]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef]
- HamadAmin, B.A.; Khwarahm, N.R. Mapping impacts of climate change on the distributions of two endemic tree species under socioeconomic pathway scenarios (SSP). Sustainability 2023, 15, 5469. [Google Scholar] [CrossRef]
- Nan, Q.; Li, C.; Li, X.; Zheng, D.; Li, Z.; Zhao, L. Modeling the Potential Distribution Patterns of the Invasive Plant Species Phytolacca americana in China in Response to Climate Change. Plants 2024, 13, 1082. [Google Scholar] [CrossRef]
- Wu, K.; Wang, Y.; Liu, Z.; Huo, W.; Cao, J.; Zhao, G.; Zhang, F.-G. Prediction of potential invasion of two weeds of the genus Avena in Asia under climate change based on Maxent. Sci. Total Environ. 2024, 950, 175192. [Google Scholar] [CrossRef]
- Yukimoto, S.; Koshiro, T.; Kawai, H.; Oshima, N.; Yoshida, K.; Urakawa, S.; Tsujino, H.; Deushi, M.; Tanaka, T.; Hosaka, M. MRI MRI-ESM2. 0 Model Output Prepared for CMIP6 CMIP Historical, Version 20190222; Earth System Grid Federation: Washington, DC, USA, 2019. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. A J. R. Meteorol. Soc. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; ACM Press: New York, NY, USA, 2004; pp. 655–662. [Google Scholar]
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.J.; Hijmans, R.; Huettmann, F.R.; Leathwick, J.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.; Li, J.; Peterson, A.T.; Graham, C.; Guisan, A.; Group, N.P.S.D.W. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Radha, K.O.; Khwarahm, N.R. An integrated approach to map the impact of climate change on the distributions of Crataegus azarolus and Crataegus monogyna in Kurdistan Region, Iraq. Sustainability 2022, 14, 14621. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable Used in Modeling | Abbreviation and Unit | Percent Contribution | Permutation Importance |
|---|---|---|---|
| Elevation | DEM (m) | 33.2 | 35.5 |
| wc_bio12 | bio12 (annual precipitation) (mm) | 31.7 | 50 |
| landcover2019 | LC 2019 | 19.1 | 4 |
| wc_bio15 | bio15 (precipitation seasonality (coefficient of variation) (mm) | 6.1 | 2.2 |
| wc_bio01 | bio1 (annual mean temperature) (°C) | 5.1 | 3.7 |
| wc_bio02 | bio2 (mean diurnal range) (°C) | 2.1 | 2.3 |
| IRAQ_NDVI | NDVI | 2 | 2 |
| wc_bio14 | bio14 (precipitation of driest month) (mm) | 0.7 | 0.3 |
| Current Distribution | SSP126_2041–2060 Distribution | SSP585_2041–2060 Distribution | ||||
|---|---|---|---|---|---|---|
| Class | Area (Km2) | Area % (Km2) | Area (Km2) | Area % (Km2) | Area (Km2) | Area % (Km2) |
| Unsuitable habitat | 366,566.50 | 85.12 | 374,098.03 | 86.87 | 387,760.80 | 90.04 |
| Low suitable habitat | 51,504.07 | 11.96 | 41,818.88 | 9.71 | 35,186.10 | 8.17 |
| Medium suitable habitat | 11,540.42 | 2.68 | 13,706.59 | 3.18 | 6762.17 | 1.57 |
| High suitable habitat | 1021.18 | 0.24 | 1008.66 | 0.23 | 923.09 | 0.21 |
| Total area | 430,632.17 | 100 | 430,632.17 | 100 | 430,632.17 | 100 |
| Current to SSP126_2041−2060 Change | Current to SSP585_2041−2060 | |||
|---|---|---|---|---|
| Class | Area (Km2) | Area % (Km2) | Area (Km2) | Area % (Km2) |
| Habitat gain | 15,403.22 | 3.58 | 7823.69 | 1.82 |
| Unsuitable | 351,163.28 | 81.55 | 358,742.81 | 83.31 |
| No change | 41,130.91 | 9.55 | 35,052.54 | 8.14 |
| Habitat loss | 22,934.75 | 5.33 | 29,013.12 | 6.74 |
| Total area | 430,632.17 | 100 | 430,632.17 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khwarahm, N.R. MaxEnt-Based Distribution Modeling of the Invasive Species Phragmites australis Under Climate Change Conditions in Iraq. Plants 2025, 14, 768. https://doi.org/10.3390/plants14050768
Khwarahm NR. MaxEnt-Based Distribution Modeling of the Invasive Species Phragmites australis Under Climate Change Conditions in Iraq. Plants. 2025; 14(5):768. https://doi.org/10.3390/plants14050768
Chicago/Turabian StyleKhwarahm, Nabaz R. 2025. "MaxEnt-Based Distribution Modeling of the Invasive Species Phragmites australis Under Climate Change Conditions in Iraq" Plants 14, no. 5: 768. https://doi.org/10.3390/plants14050768
APA StyleKhwarahm, N. R. (2025). MaxEnt-Based Distribution Modeling of the Invasive Species Phragmites australis Under Climate Change Conditions in Iraq. Plants, 14(5), 768. https://doi.org/10.3390/plants14050768