
Genotypic Variability in Root Morphology in a Diverse Wheat Genotypes Under Drought and Low Phosphorus Stress

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
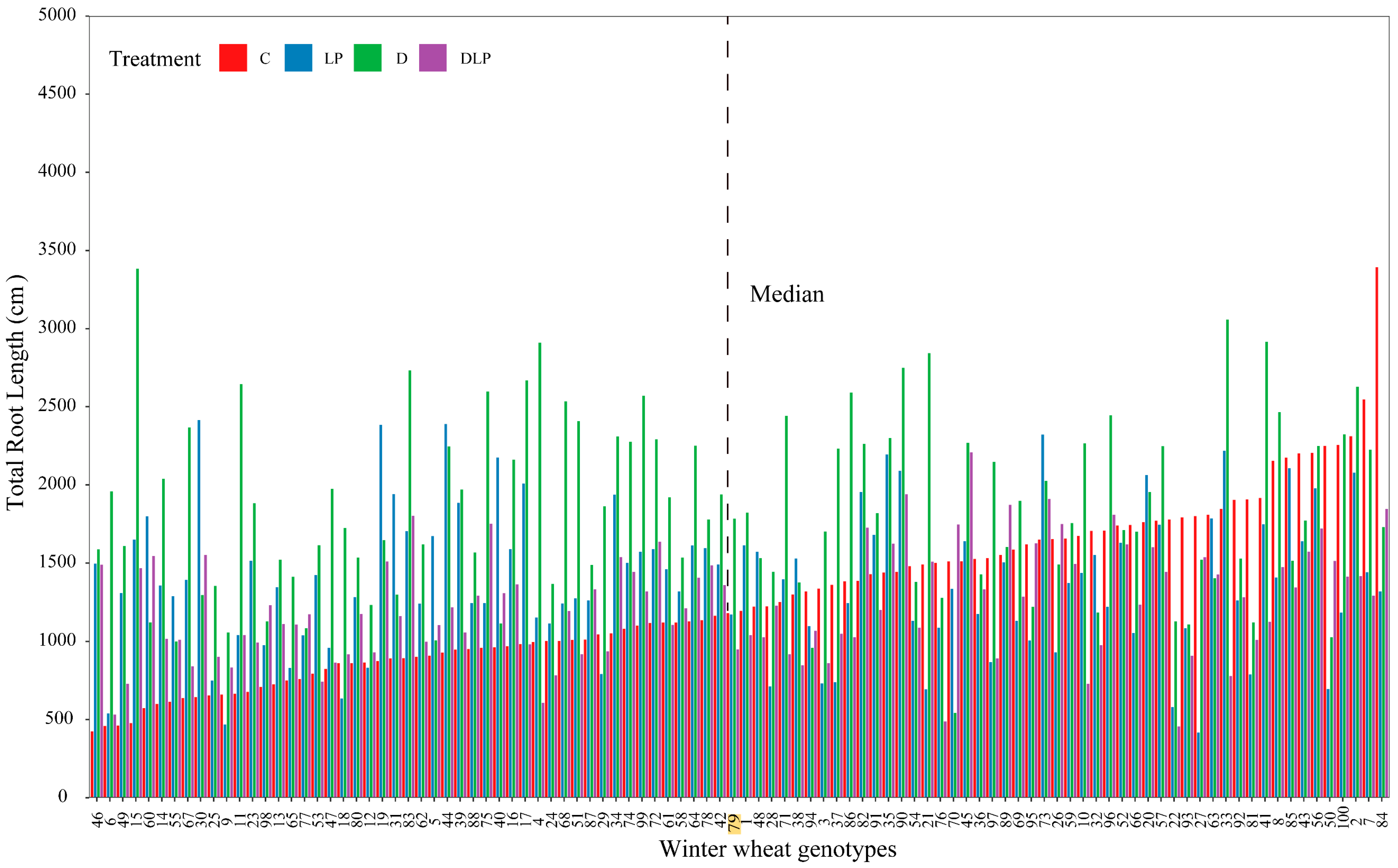
2.1. Phenotypic Variation of Winter Wheat Genotypes
2.2. Correlation Among Traits
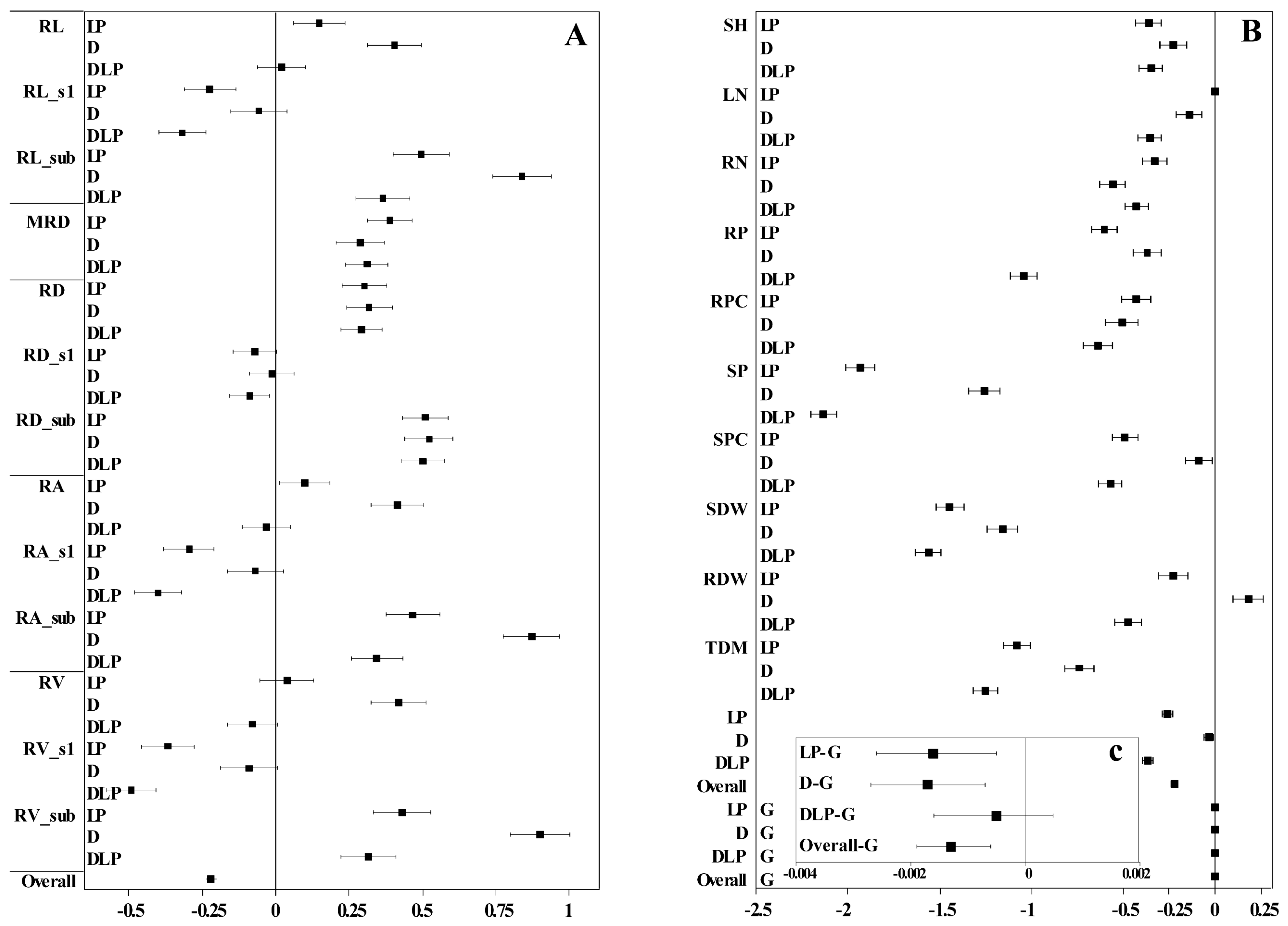
2.3. Determination of Root Trait Variability
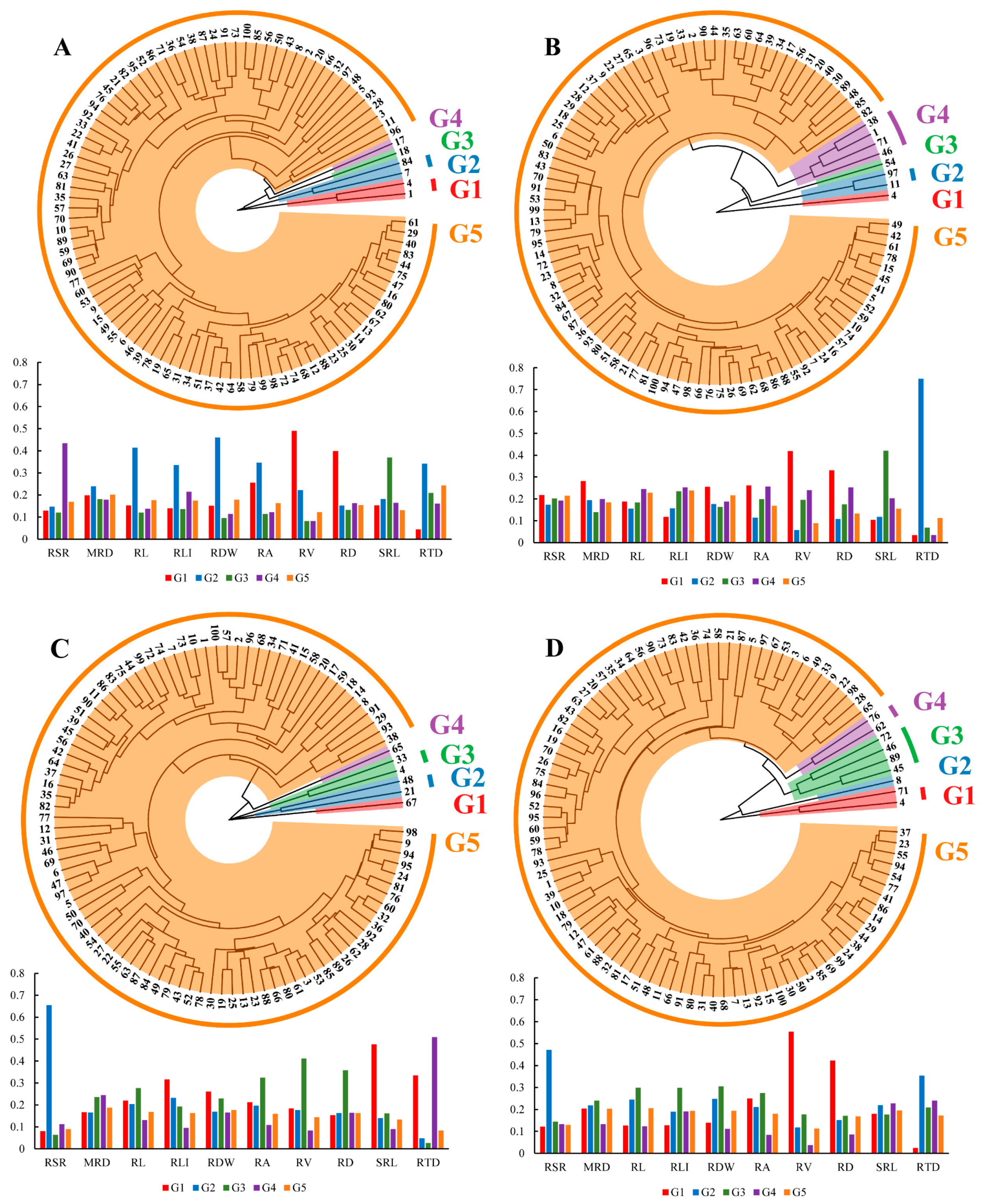
2.4. Genotype Homogeneous Grouping Based on Root Trait Variation
3. Discussion
3.1. Genetic Variation of Winter Wheat Root Traits
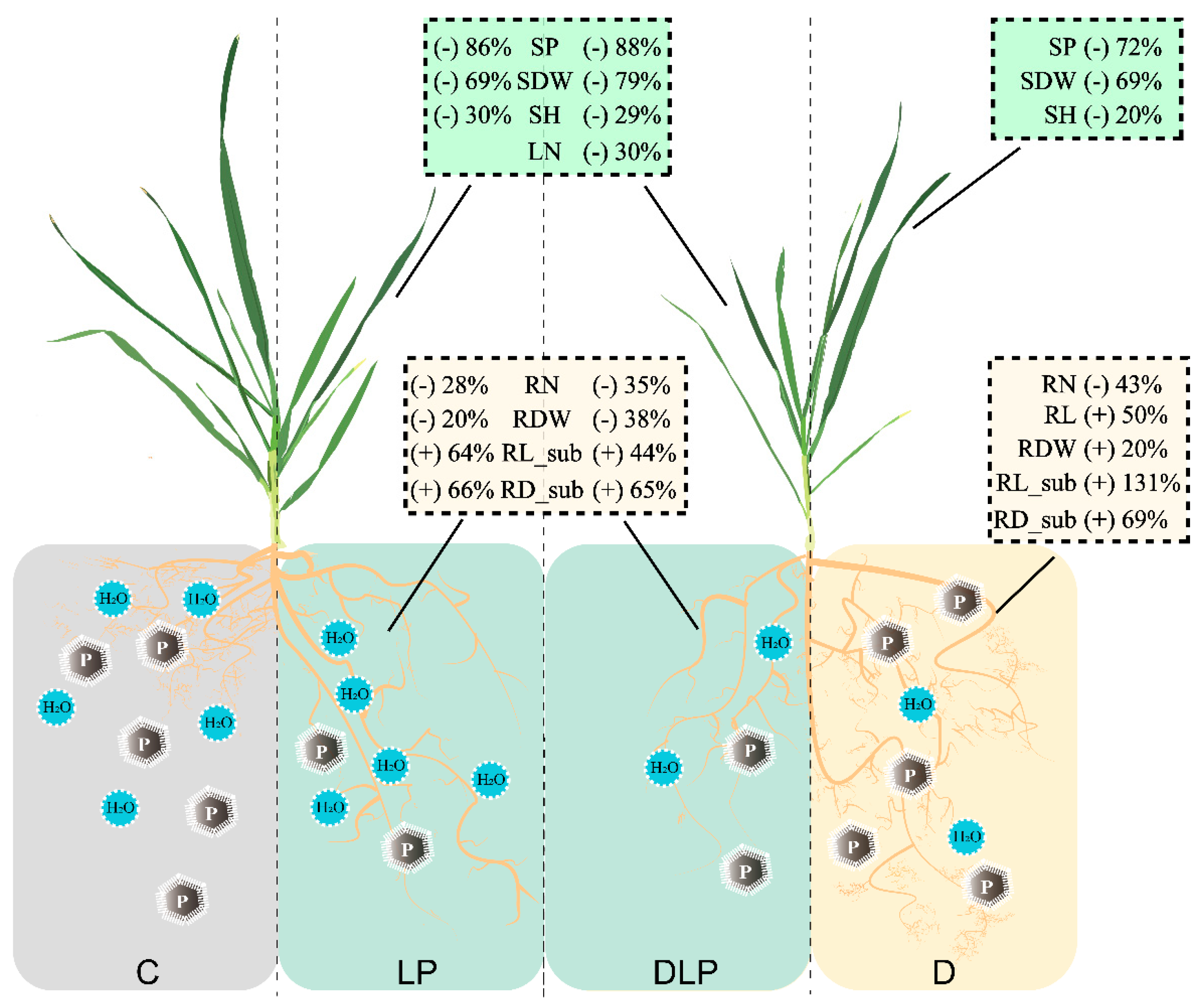
3.2. Variation of Winter Wheat Shoot and Root Traits to Drought and Phosphorus Stress
3.3. Genotype Selection Based on Root Trait Properties for Breeding Programs
4. Materials and Methods
4.1. Plant Materials and Experimental Design
4.2. Sampling
- Stress sensitivity index ),
- Tolerance index ,
- Stress index ,
- Harmonized mean index ,
- Mean productivity index ,
- Geometric mean productivity index ,
- Stress tolerance index .
4.3. Statistics and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Thorup-Kristensen, K.; Salmerón Cortasa, M.; Loges, R. Winter wheat roots grow twice as deep as spring wheat roots, is this important for N uptake and N leaching losses? Plant Soil 2009, 322, 101–114. [Google Scholar] [CrossRef]
- Wu, M.; Xu, Y.; Zheng, J.; Hao, Z. North Expansion of Winter Wheat Planting Area in China under Different Emissions Scenarios. Agriculture 2022, 12, 763. [Google Scholar] [CrossRef]
- Niu, X.; Hu, T.; Zhang, F.; Feng, P. Severity and duration of osmotic stress on partial root system: Effects on root hydraulic conductance and root growth. Plant Growth Regul. 2015, 79, 177–186. [Google Scholar] [CrossRef]
- Ludwiczak, A.; Osiak, M.; Cárdenas-Pérez, S.; Lubińska-Mielińska, S.; Piernik, A. Osmotic Stress or Ionic Composition: Which Affects the Early Growth of Crop Species More? Agronomy 2021, 11, 15. [Google Scholar] [CrossRef]
- Raghothama, K.G. PHOSPHATE ACQUISITION. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 665–693. [Google Scholar] [CrossRef]
- Mat Hassan, H.; Marschner, P.; McNeill, A.; Tang, C. Growth, P uptake in grain legumes and changes in rhizosphere soil P pools. Biol. Fertil. Soils 2012, 48, 151–159. [Google Scholar] [CrossRef]
- Cong, W.F.; Suriyagoda, L.D.B.; Lambers, H. Tightening the Phosphorus Cycle through Phosphorus-Efficient Crop Genotypes. Trends Plant Sci. 2020, 25, 967–975. [Google Scholar] [CrossRef]
- Li, S.-X.; Wang, Z.-H.; Malhi, S.S.; Li, S.-Q.; Gao, Y.-J.; Tian, X.-H. Chapter 7 Nutrient and Water Management Effects on Crop Production, and Nutrient and Water Use Efficiency in Dryland Areas of China. In Advances in Agronomy; Sparks, D.L., Ed.; Elsevier: Amsterdam, The Netherlands, 2009; Volume 102, pp. 223–265. [Google Scholar]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Sun, X.; Ren, W.; Wang, P.; Chen, F.; Yuan, L.; Pan, Q.; Mi, G. Evaluation of maize root growth and genome-wide association studies of root traits in response to low nitrogen supply at seedling emergence. Crop J. 2021, 9, 794–804. [Google Scholar] [CrossRef]
- Li, Y.; Wei, R.; Zhang, R.; Sun, S.; Xu, G. Facilitated legume nodulation, phosphate uptake and nitrogen transfer by arbuscular inoculation in an upland rice and mung bean intercropping system. Plant Soil 2009, 315, 285–296. [Google Scholar] [CrossRef]
- Lynch, J.P.; Brown, K.M. Topsoil foraging—An architectural adaptation of plants to low phosphorus availability. Plant Soil 2001, 237, 225–237. [Google Scholar] [CrossRef]
- Lambers, H.; Raven, J.A.; Shaver, G.R.; Smith, S.E. Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef]
- Lindh, M.; Zhang, L.; Falster, D.; Franklin, O.; Brännström, Å. Plant diversity and drought: The role of deep roots. Ecol. Modell. 2014, 290, 85–93. [Google Scholar] [CrossRef]
- Colombi, T.; Walter, A. Genetic Diversity under Soil Compaction in Wheat: Root Number as a Promising Trait for Early Plant Vigor. Front. Plant Sci. 2017, 8, 420. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Peng, Z.; Mao, X.; Wang, J.; Chang, X.; Reynolds, M.; Jing, R. Genome-wide association study reveals genomic regions controlling root and shoot traits at late growth stages in wheat. Ann. Bot. 2019, 124, 993–1006. [Google Scholar] [CrossRef]
- Song, P.; Wang, J.; Guo, X.; Yang, W.; Zhao, C. High-throughput phenotyping: Breaking through the bottleneck in future crop breeding. Crop J. 2021, 9, 633–645. [Google Scholar] [CrossRef]
- Chen, Y.L.; Dunbabin, V.M.; Diggle, A.J.; Siddique, K.H.M.; Rengel, Z. Development of a novel semi-hydroponic phenotyping system for studying root architecture. Funct. Plant Biol. 2011, 38, 355–363. [Google Scholar] [CrossRef]
- Qiao, S.; Fang, Y.; Wu, A.; Xu, B.; Zhang, S.; Deng, X.; Djalovic, I.; Siddique, K.H.M.; Chen, Y. Dissecting root trait variability in maize genotypes using the semi-hydroponic phenotyping platform. Plant Soil 2018, 439, 75–90. [Google Scholar] [CrossRef]
- Liu, S.; Begum, N.; An, T.; Zhao, T.; Xu, B.; Zhang, S.; Deng, X.; Lam, H.M.; Nguyen, H.T.; Siddique, K.H.M.; et al. Characterization of Root System Architecture Traits in Diverse Soybean Genotypes Using a Semi-Hydroponic System. Plants 2021, 10, 2781. [Google Scholar] [CrossRef]
- Chen, Y.; Palta, J.; Prasad, P.V.V.; Siddique, K.H.M. Phenotypic variability in bread wheat root systems at the early vegetative stage. BMC Plant Biol. 2020, 20, 185. [Google Scholar] [CrossRef] [PubMed]
- Mollier, A.; Pellerin, S. Maize root system growth and development as influenced by phosphorus deficiency. J. Exp. Bot. 1999, 50, 487–497. [Google Scholar] [CrossRef]
- Zhao, J.; Bodner, G.; Rewald, B.; Leitner, D.; Nagel, K.A.; Nakhforoosh, A. Root architecture simulation improves the inference from seedling root phenotyping towards mature root systems. J. Exp. Bot. 2017, 68, 965–982. [Google Scholar] [CrossRef] [PubMed]
- Violle, C.; Navas, M.L.; Vile, D.; Kazakou, E.; Fortunel, C.; Hummel, I.; Garnier, E. Let the concept of trait be functional! Oikos 2007, 116, 882–892. [Google Scholar] [CrossRef]
- ZarzyŃSka, K.; Boguszewska-MaŃKowska, D.; Nosalewicz, A. Differences in size and architecture of the potato cultivars root system and their tolerance to drought stress. Plant, Soil Environ. 2017, 63, 159–164. [Google Scholar] [CrossRef]
- Fradgley, N.; Evans, G.; Biernaskie, J.M.; Cockram, J.; Marr, E.C.; Oliver, A.G.; Ober, E.; Jones, H. Effects of breeding history and crop management on the root architecture of wheat. Plant Soil 2020, 452, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Paz-Vidal, P.; Castillo-Rosales, D.; López, M.D.; Tejos, I.M.; Noriega, F.; Schoebitz, M. Root Morphological and Physiological Traits, Rhizosphere Biological Activity, and Mycorrhizal Colonization of 14 Winter Wheat Varieties Released in Chile Between 1965 and 2020. J. Soil Sci. Plant Nutr. 2023, 23, 5389–5403. [Google Scholar] [CrossRef]
- Li, C.; Li, L.; Reynolds, M.P.; Wang, J.; Chang, X.; Mao, X.; Jing, R. Recognizing the hidden half in wheat: Root system attributes associated with drought tolerance. J. Exp. Bot. 2021, 72, 5117–5133. [Google Scholar] [CrossRef]
- Ferrieri, A.P.; Machado, R.A.R.; Arce, C.C.M.; Kessler, D.; Baldwin, I.T.; Erb, M. Localized micronutrient patches induce lateral root foraging and chemotropism in Nicotiana attenuata. J. Integr. Plant. Biol. 2017, 59, 759–771. [Google Scholar] [CrossRef]
- San-Oh, Y.; Kondo, M.; Ookawa, T.; Hirasawa, T. Ecophysiological Analysis on Effect of Planting Pattern on Biomass Production and Grain Yield in Rice. Jpn. Agric. Res. Q. 2008, 42, 79–89. [Google Scholar] [CrossRef]
- Liu, Z.; Qin, T.; Atienza, M.; Zhao, Y.; Nguyen, H.; Sheng, H.; Olukayode, T.; Song, H.; Panjvani, K.; Magalhaes, J.; et al. Constitutive basis of root system architecture: Uncovering a promising trait for breeding nutrient- and drought-resilient crops. aBIOTECH 2023, 4, 315–331. [Google Scholar] [CrossRef]
- Borden, K.A.; Thomas, S.C.; Isaac, M.E. Variation in fine root traits reveals nutrient-specific acquisition strategies in agroforestry systems. Plant Soil 2019, 453, 139–151. [Google Scholar] [CrossRef]
- Lynch, J.P. Root phenes for enhanced soil exploration and phosphorus acquisition: Tools for future crops. Plant Physiol. 2011, 156, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.A.; Postma, J.A.; Lynch, J.P. Phene synergism between root hair length and basal root growth angle for phosphorus acquisition. Plant Physiol. 2015, 167, 1430–1439. [Google Scholar] [CrossRef] [PubMed]
- Palta, J.A.; Chen, X.; Milroy, S.P.; Rebetzke, G.J.; Dreccer, M.F.; Watt, M. Large root systems: Are they useful in adapting wheat to dry environments? Funct. Plant Biol. 2011, 38, 347–354. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Gowda, M.; Cairns, J.E.; Mwafulirwa, L.; Daniell, T.J.; Thierfelder, C.; Paterson, E.; Baggs, E.M. Evidence of a plant genetic basis for maize roots impacting soil organic matter mineralization. Soil Biol. Biochem. 2021, 161, 108402. [Google Scholar] [CrossRef]
- Zhu, J.; Kaeppler, S.M.; Lynch, J.P. Topsoil foraging and phosphorus acquisition efficiency in maize (Zea mays). Funct. Plant Biol. 2005, 32, 749–762. [Google Scholar] [CrossRef]
- Odone, A.; Popovic, O.; Thorup-Kristensen, K. Deep roots: Implications for nitrogen uptake and drought tolerance among winter wheat cultivars. Plant Soil 2023, 500, 13–32. [Google Scholar] [CrossRef]
- Homulle, Z.; George, T.S.; Karley, A.J. Root traits with team benefits: Understanding belowground interactions in intercropping systems. Plant Soil 2021, 471, 1–26. [Google Scholar] [CrossRef]
- Lalli, D.A.; Decroocq, V.; Blenda, A.V.; Schurdi-Levraud, V.; Garay, L.; Le Gall, O.; Damsteegt, V.; Reighard, G.L.; Abbott, A.G. Identification and mapping of resistance gene analogs (RGAs) in Prunus: A resistance map for Prunus. Theor. Appl. Genet. 2005, 111, 1504–1513. [Google Scholar] [CrossRef]
- Camilo, S.; Odindo, A.O.; Kondwakwenda, A.; Sibiya, J. Root Traits Related with Drought and Phosphorus Tolerance in Common Bean (Phaseolus vulgaris L.). Agronomy 2021, 11, 552. [Google Scholar] [CrossRef]
- Jun, W.; Ping, L.; Zhiyong, L.; Zhansheng, W.; Yongshen, L.; Xinyuan, G. Dry matter accumulation and phosphorus efficiency response of cotton cultivars to phosphorus and drought. J. Plant Nutr. 2017, 40, 2349–2357. [Google Scholar] [CrossRef]
- Jain, N.K.; Dahama, A.K. Effect of phosphorus and zinc on yield, nutrient uptake and quality of wheat (Triticum aestivum). Indian J. Agric. Sci. 2007, 77, 310–313. [Google Scholar]
- Awad, W.; Byrne, P.F.; Reid, S.D.; Comas, L.H.; Haley, S.D. Great Plains Winter Wheat Varies for Root Length and Diameter under Drought Stress. Agronomy 2018, 110, 226–235. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Prasad, P.V.V.; Kumari, J.; Rengel, Z. Root length and root lipid composition contribute to drought tolerance of winter and spring wheat. Plant Soil 2018, 439, 57–73. [Google Scholar] [CrossRef]
- Lynch, J.P.; Brown, K.M. Root strategies for phosphorus acquisition. In The Ecophysiology of Plant-Phosphorus Interactions; White, P.J., Hammond, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 83–116. [Google Scholar]
- Chen, Y.L.; Dunbabin, V.M.; Postma, J.A.; Diggle, A.J.; Palta, J.A.; Lynch, J.P.; Siddique, K.H.M.; Rengel, Z. Phenotypic variability and modelling of root structure of wild Lupinus angustifolius genotypes. Plant Soil 2011, 348, 345–364. [Google Scholar] [CrossRef]
- Strock, C.F.; Morrow de la Riva, L.; Lynch, J.P. Reduction in Root Secondary Growth as a Strategy for Phosphorus Acquisition. Plant Physiol. 2018, 176, 691–703. [Google Scholar] [CrossRef]
- Ma, Z.; Bielenberg, D.G.; Brown, K.M.; Lynch, J.P. Regulation of root hair density by phosphorus availability in Arabidopsis thaliana. Plant Cell Environ. 2001, 24, 459–467. [Google Scholar] [CrossRef]
- Beroueg, A.; Lecompte, F.; Mollier, A.; Pages, L. Genetic Variation in Root Architectural Traits in Lactuca and Their Roles in Increasing Phosphorus-Use-Efficiency in Response to Low Phosphorus Availability. Front. Plant Sci. 2021, 12, 658321. [Google Scholar] [CrossRef] [PubMed]
- Robin, A.H.K.; Ghosh, S.; Shahed, M.A. PEG-Induced Osmotic Stress Alters Root Morphology and Root Hair Traits in Wheat Genotypes. Plants 2021, 10, 20. [Google Scholar] [CrossRef]
- Zhang, C.; Simpson, R.J.; Kim, C.M.; Warthmann, N.; Delhaize, E.; Dolan, L.; Byrne, M.E.; Wu, Y.; Ryan, P.R. Do longer root hairs improve phosphorus uptake? Testing the hypothesis with transgenic Brachypodium distachyon lines over-expressing endogenous RSL genes. New Phytol. 2018, 217, 1654–1666. [Google Scholar] [CrossRef] [PubMed]
- Suriyagoda, L.D.; Ryan, M.H.; Renton, M.; Lambers, H. Adaptive shoot and root responses collectively enhance growth at optimum temperature and limited phosphorus supply of three herbaceous legume species. Ann. Bot. 2012, 110, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Milly, P.C.; Dunne, K.A.; Vecchia, A.V. Global pattern of trends in streamflow and water availability in a changing climate. Nature 2005, 438, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Jha, U.C.; Nayyar, H.; Parida, S.K.; Beena, R.; Pang, J.; Siddique, K.H.M. Breeding and genomics approaches for improving phosphorus-use efficiency in grain legumes. Environ. Exp. Bot. 2023, 205, 105120. [Google Scholar] [CrossRef]
- Schneider, H.M.; Lynch, J.P. Should Root Plasticity Be a Crop Breeding Target? Front. Plant Sci. 2020, 11, 546. [Google Scholar] [CrossRef]
- Lynch, J.P.; Galindo-Castañeda, T.; Schneider, H.M.; Sidhu, J.S.; Rangarajan, H.; York, L.M. Root phenotypes for improved nitrogen capture. Plant Soil 2023, 502, 31–85. [Google Scholar] [CrossRef]
- Richardson, A.E.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; Smith, S.E.; Harvey, P.R.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; et al. Plant and microbial strategies to improve the phosphorus efficiency of agriculture. Plant Soil 2011, 349, 121–156. [Google Scholar] [CrossRef]
- Zhan, A.; Schneider, H.; Lynch, J.P. Reduced Lateral Root Branching Density Improves Drought Tolerance in Maize. Plant Physiol. 2015, 168, 1603–1615. [Google Scholar] [CrossRef]
- Fan, S.; Blake, T.J. Comparison of polyethylene glycol 3350 induced osmotic stress and soil drying for drought simulation in three woody species. Trees 1997, 11, 342–348. [Google Scholar] [CrossRef]
- Cui, G.; Zhao, Y.; Zhang, J.; Chao, M.; Xie, K.; Zhang, C.; Sun, F.; Liu, S.; Xi, Y. Proteomic analysis of the similarities and differences of soil drought and polyethylene glycol stress responses in wheat (Triticum aestivum L.). Plant Mol. Biol. 2019, 100, 391–410. [Google Scholar] [CrossRef]
- Salehi Eskandari, B.; Ghaderian, S.M.; Schat, H. The role of nickel (Ni) and drought in serpentine adaptation: Contrasting effects of Ni on osmoprotectants and oxidative stress markers in the serpentine endemic, Cleome heratensis, and the related non-serpentinophyte, Cleome foliolosa. Plant Soil 2017, 417, 183–195. [Google Scholar] [CrossRef]
- Wu, S.; Hu, C.; Tan, Q.; Nie, Z.; Sun, X. Effects of molybdenum on water utilization, antioxidative defense system and osmotic-adjustment ability in winter wheat (Triticum aestivum) under drought stress. Plant Physiol. Biochem. 2014, 83, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ramírez, D.A.; Qin, J.; Dormatey, R.; Bi, Z.; Sun, C.; Wang, H.; Bai, J. Water restriction scenarios and their effects on traits in potato with different degrees of drought tolerance. Sci. Hortic 2019, 256, 108525. [Google Scholar] [CrossRef]
- Bao, S. Soil and Agricultural Chemistry Analysis. China Agriculture Press: Beijing, China, 2007. [Google Scholar]
- Irfan, M.; Abbas, M.; Shah, J.; Akram, M.; Depar, N.; Memon, M. Categorization and Identification of Brassica Genotypes for Phosphorus Utilization Efficiency. Int. J. Agric. Biol. 2020, 23, 227–234. [Google Scholar]
- Grzesiak, S.; Hordyńska, N.; Szczyrek, P.; Grzesiak, M.T.; Noga, A.; Szechyńska-Hebda, M. Variation among wheat (Triticum easativum L.) genotypes in response to the drought stress: I—Selection approaches. J. Plant Interact. 2018, 14, 30–44. [Google Scholar] [CrossRef]
- Kothari, D.; Pargaien, N.; Tewari, L.M.; Dikshit, H.K.; Mishra, G.P.; Aski, M.S.; Bansal, R.; Gupta, S.; Kumar, S.; Nair, R.M. Genetic Variation for Traits Related to Phosphorus Use Efficiency in Vigna Species. Agronomy 2023, 13, 305. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Taiyun, W.; Viliam, S. R Package ‘corrplot’: Visualization of a Correlation Matrix (Version 0.92); R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef]
- Wang, J.; Li, H.; Zhang, L.; Meng, L. Users’ Manual of QTL IciMapping; The Quantitative Genetics Group, Institute of Crop Science, Chinese Academy of Agricultural Sciences (CAAS): Beijing, China, 2019. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: AnRPackage for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. factoextra: Extract and Visualize the Results of Multivariate Data Analyses_, R package Version 1.0.7. 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 22 November 2024).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Walach, J.; Filzmoser, P.; Hron, K. Chapter Seven—Data Normalization and Scaling: Consequences for the Analysis in Omics Sciences. In Comprehensive Analytical Chemistry; Jaumot, J., Bedia, C., Tauler, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 82, pp. 165–196. [Google Scholar]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The Meta-Analysis of Response Ratios in Experimental Ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Abbreviation | Description | Unit |
|---|---|---|---|
| Maximum root depth | MRD | The longest seminal root length | cm |
| Root number | RN | Seminal and primary root number (from seeds) | Number per plant |
| Root length | RL | Total root length per plant | cm |
| Root diameter | RD | Average root diameter | mm |
| Root area | RA | Total root surface area | cm2 |
| Root volume | RV | Total root volume | cm3 |
| Root length density | RLD | Root length per unit area (0–110 cm depth, 1430 cm2) | cm·cm−2 |
| Specific root length | SRL | Total root length per unit root dry mass | mg−1 dry mass |
| Root length Intensity | RLI | Total root length per unit root depth | cm·cm−1 |
| Root tissue density | RTD | Root dry mass per unit root volume | mg·cm−3 |
| Root growth rate | RGR | Average daily root growth (based on the longest seminal or primary root growth at 65 days after transplanting) | cm·d−1 |
| Root dry weight | RDW | Root dry weight per plant | g |
| Root phosphorus concentration | RPC | Root phosphorus concentration per gram dry weight | mg·g−1 |
| Root phosphorus content | RP | Root phosphorus content per plant | mg |
| Shoot dry weight | SDW | Shoot dry weight per plant | g |
| Root–shoot ratio | RSR | Root-to-shoot dry mass ratio | |
| Total dry mass | TDM | Total dry mass (sum of root and shoot dry weight) | mg |
| Shoot height | SH | Shoot height measured to the tallest leaf | cm |
| Leaf number | LN | Number of leaves per plant | |
| Tiller number | TN | Number of tillers per plant | |
| Shoot growth rate | SGR | Average daily shoot growth | cm·d−1 |
| Shoot phosphorus concentration | SPC | Shoot phosphorus concentration per gram of dry weight | mg·g−1 |
| Shoot phosphorus content | SP | Shoot phosphorus content per plant | mg |
| Total phosphorus content | TP | Total phosphorus content per plant | mg |
| Trait | CV | Mean | ||||||
|---|---|---|---|---|---|---|---|---|
| C | LP | D | DLP | C | LP | D | DLP | |
| MRD | 0.31 | 0.28 | 0.36 | 0.27 | 55.10 c | 79.87 a | 73.29 b | 74.81 b |
| RL | 0.61 | 0.52 | 0.51 | 0.45 | 1290.21 b | 1390.51 b | 1846.70 a | 1242.26 b |
| RD | 0.50 | 0.50 | 0.40 | 0.40 | 0.18 b | 0.24 a | 0.24 a | 0.23 a |
| RA | 0.57 | 0.50 | 0.52 | 0.45 | 97.24 b | 103.15 b | 143.20 a | 91.33 b |
| RV | 0.79 | 0.85 | 0.66 | 0.84 | 0.66 b | 0.69 b | 0.96 a | 0.62 b |
| SRL | 0.73 | 0.64 | 0.95 | 0.48 | 12954.18 b | 16974.74 a | 17172.59 a | 18690.97 a |
| RLI | 0.65 | 0.54 | 0.68 | 0.57 | 24.88 b | 18.37 c | 28.46 a | 17.83 c |
| RTD | 1.12 | 1.59 | 1.62 | 1.09 | 0.27 a | 0.22 ab | 0.26 a | 0.18 b |
| RDW | 0.51 | 0.32 | 0.49 | 0.28 | 0.12 b | 0.09 c | 0.14 a | 0.07 d |
| SDW | 0.48 | 0.37 | 0.53 | 0.38 | 0.52 a | 0.12 c | 0.16 b | 0.11 c |
| TDM | 0.45 | 0.27 | 0.42 | 0.29 | 0.63 a | 0.20 c | 0.30 b | 0.18 c |
| RSR | 0.54 | 0.52 | 1.47 | 0.56 | 0.25 c | 0.85 b | 1.19 a | 0.77 b |
| Items | LP | D | DLP | |
|---|---|---|---|---|
| STS | PAE | −0.730 ** | −0.668 ** | −0.617 ** |
| △RL | −0.456 ** | −0.532 ** | −0.425 ** | |
| △RL_s1 | −0.285 ** | −0.452 ** | −0.293 ** | |
| △RL_sub | −0.468 ** | −0.455 ** | −0.426 ** | |
| △RD | −0.217 * | −0.122 ns | −0.071 ns | |
| △RD_s1 | 0.088 ns | −0.044 ns | 0.026 ns | |
| △RD_sub | −0.246 * | −0.142 ns | −0.104 ns | |
| PAE | △RL | 0.510 ** | 0.588 ** | 0.515 ** |
| △RL_s1 | 0.373 ** | 0.495 ** | 0.431 ** | |
| △RL_sub | 0.497 ** | 0.480 ** | 0.493 ** | |
| △RD | 0.277 * | 0.190 ns | 0.165 ns | |
| △RD_s1 | −0.083 ns | 0.045 ns | 0.129 ns | |
| △RD_sub | 0.296 ** | 0.210 * | 0.186 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Chen, Y.; Xu, Y.; Sun, H.; Gao, Y.; Yan, P.; Song, Q.; Li, S.; Zhan, A. Genotypic Variability in Root Morphology in a Diverse Wheat Genotypes Under Drought and Low Phosphorus Stress. Plants 2024, 13, 3361. https://doi.org/10.3390/plants13233361
Li X, Chen Y, Xu Y, Sun H, Gao Y, Yan P, Song Q, Li S, Zhan A. Genotypic Variability in Root Morphology in a Diverse Wheat Genotypes Under Drought and Low Phosphorus Stress. Plants. 2024; 13(23):3361. https://doi.org/10.3390/plants13233361
Chicago/Turabian StyleLi, Xin, Yinglong Chen, Yuzhou Xu, Haoyang Sun, Yamin Gao, Peng Yan, Qilong Song, Shiqing Li, and Ai Zhan. 2024. "Genotypic Variability in Root Morphology in a Diverse Wheat Genotypes Under Drought and Low Phosphorus Stress" Plants 13, no. 23: 3361. https://doi.org/10.3390/plants13233361
APA StyleLi, X., Chen, Y., Xu, Y., Sun, H., Gao, Y., Yan, P., Song, Q., Li, S., & Zhan, A. (2024). Genotypic Variability in Root Morphology in a Diverse Wheat Genotypes Under Drought and Low Phosphorus Stress. Plants, 13(23), 3361. https://doi.org/10.3390/plants13233361