
Elaeocarpus sylvestris var. ellipticus Extract and Its Major Component, Geraniin, Inhibit Herpes Simplex Virus-1 Replication



Abstract
1. Introduction
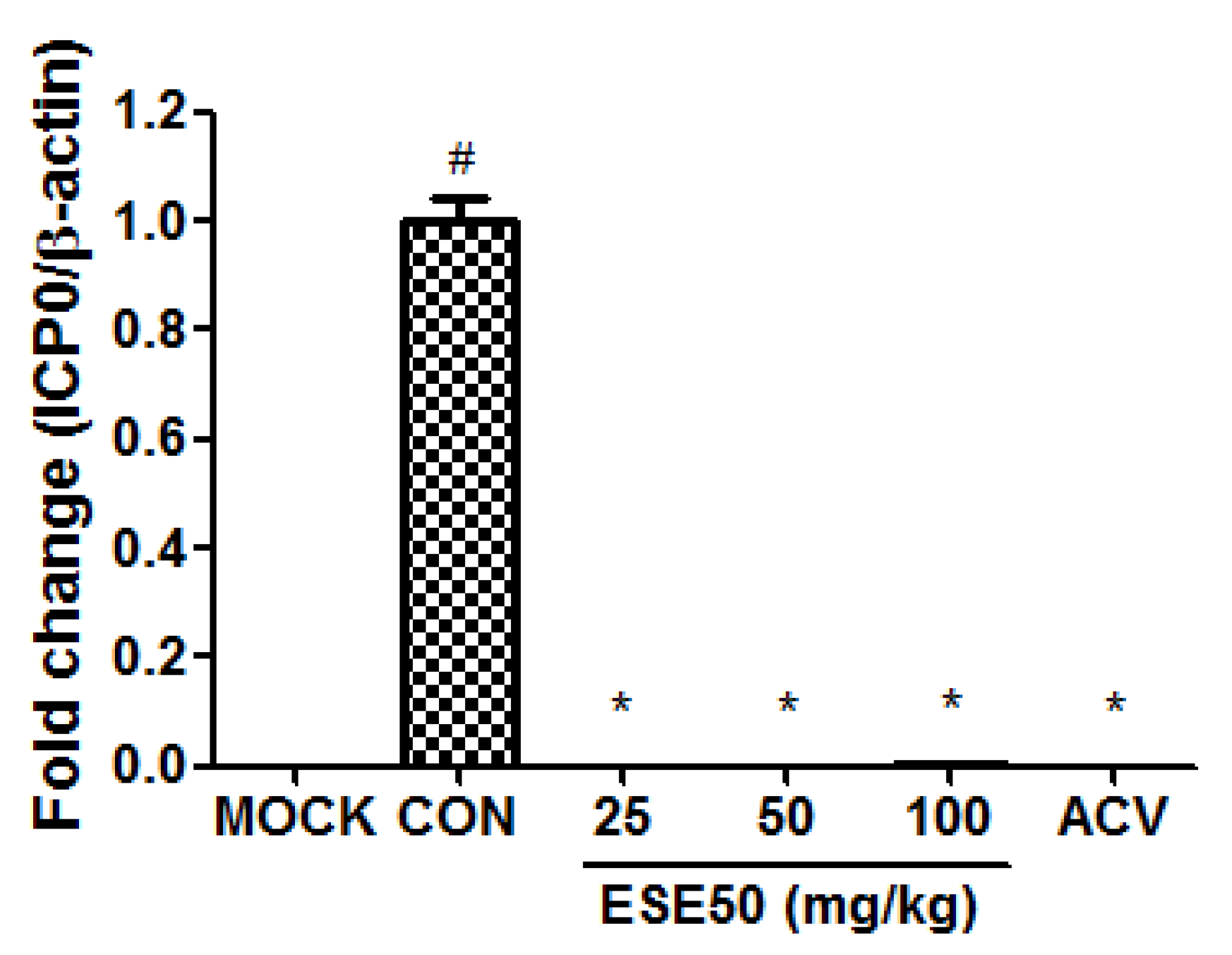
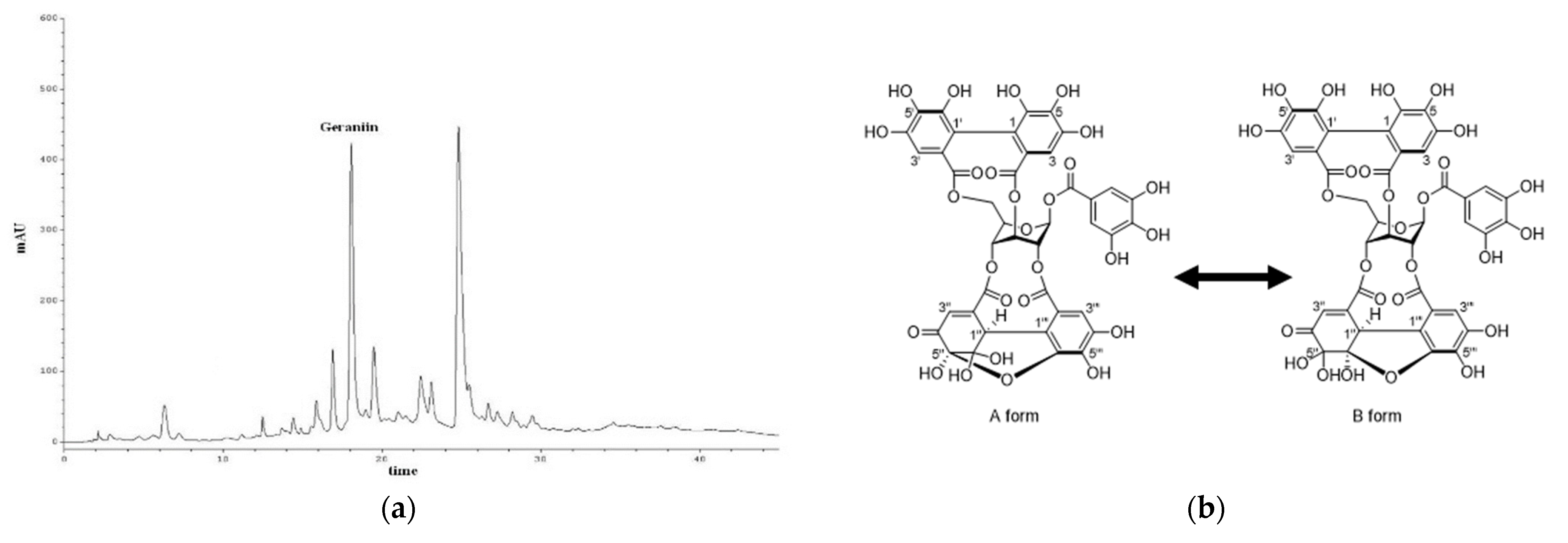
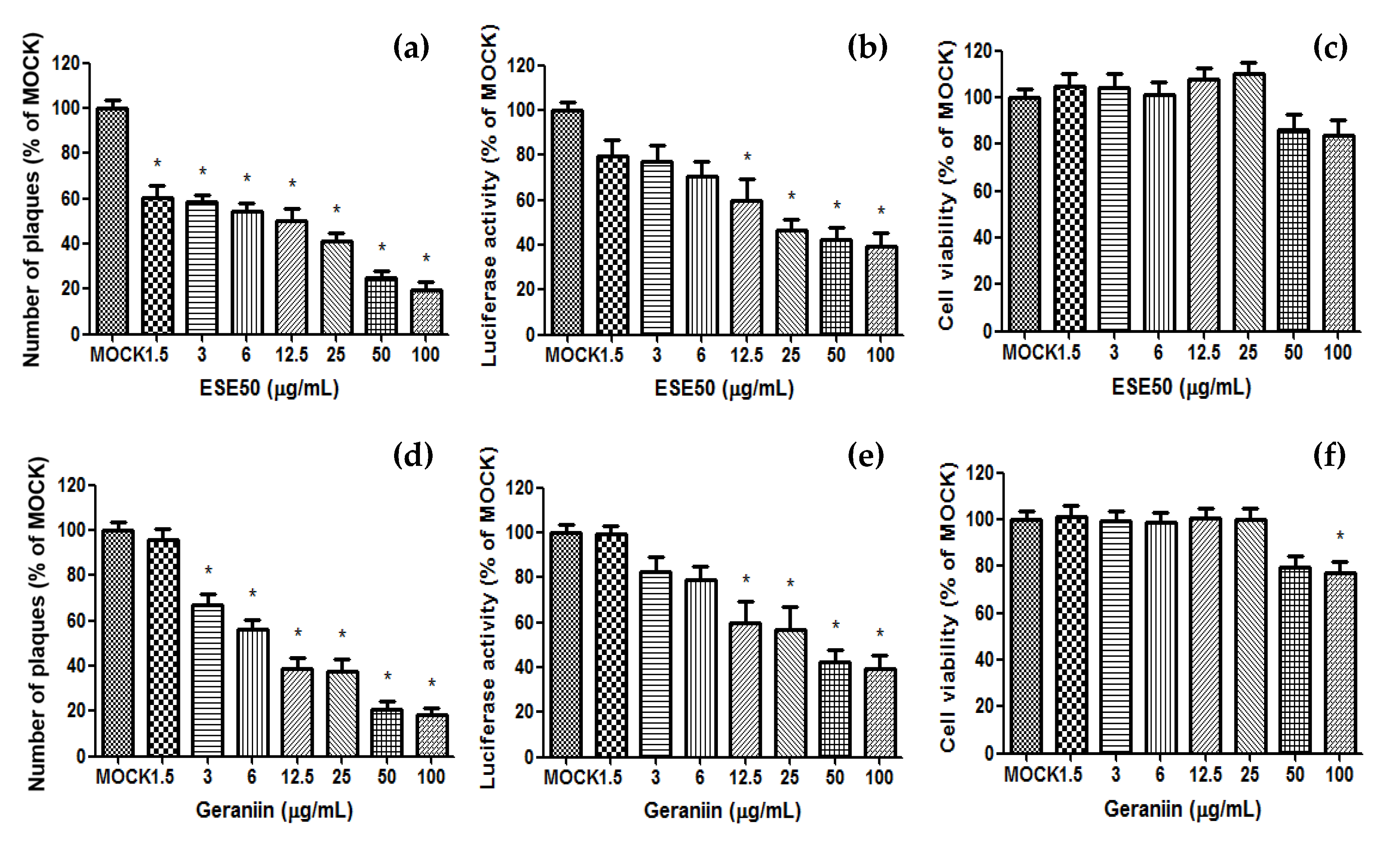
2. Results and Discussion
3. Materials and Methods
3.1. Analytical Procedures for Phytochemical Analysis
3.2. Sample Preparation
3.3. High-Performance Liquid Chromatography Analysis
3.4. Cytotoxicity Assay
3.5. Antiviral Assays
3.6. Animals
3.7. In Vivo Conditions
3.8. In Vivo Protocols
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shahin, V.H.; Oberleithner, H.; Ludwig, Y.; Windoffer, B.; Schillers, H.; Kuhn, J.E. The Genome of HSV-1 Translocates through the Nuclear Pore as a Condensed Rod-Like Structure. J. Cell Sci. 2006, 119, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Zhuo, C.; He, Z.; Wang, H.; Liu, W.; Zhang, R.; Wang, Y. Anti-Herpes Simplex Virus Activity of Polysaccharides from Eucheuma gelatinae. World J. Microb. Biot. 2015, 31, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Pujol, C.A.; Sepúlveda, S.C.; Richmond, V.; Maier, M.S.; Damonte, E.B. Polyhydroxylated Sulfated Steroids Derived from 5alpha-Cholestanes as Antiviral Agents against Herpes Simplex Virus. Arch. Virol. 2016, 161, 1993–1999. [Google Scholar] [CrossRef] [PubMed]
- Arduino, P.G.; Porter, P.S. Herpes Simplex Virus Type 1 Infection: Overview on Relevant Clinico-Pathological Features. J. Oral Pathol. Med. 2008, 37, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Whitley, R.J. Herpes Simplex Encephalitis: Adolescents and Adults. Antiviral Res. 2006, 71, 141–148. [Google Scholar] [CrossRef]
- Shivkumar, M.; Milho, R.; May, J.S.; Nicoll, M.P.; Efstathiou, S.; Stevenson, P.G. Herpes Simplex Virus 1 Targets the Murine Olfactory Neuroepithelium for Host Entry. J. Virol. 2013, 87, 10477–10488. [Google Scholar] [CrossRef] [PubMed]
- Roizman, B.; Knipe, D.M. Herpes Simplex Viruses and Their Replication. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2001; pp. 2399–2459. [Google Scholar]
- Rong, B.L.; Libermann, T.A.; Kogawa, K.; Ghosh, S.; Cao, L.X.; Langston, D.P.; Dunkel, E.C. HSV-1-Inducible Proteins Bind to NF-κB-Like Sites in the HSV-1 Genome. Virology 1992, 189, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Halford, W.P.; Schaffer, P.A. ICP0 Is Required for Efficient Reactivation of Herpes Simplex Virus Type 1 from Neuronal Latency. J. Virol. 2001, 75, 3240–3249. [Google Scholar] [CrossRef] [PubMed]
- Leib, D.A.; Coen, D.M.; Bogard, C.L.; Hicks, K.A.; Yager, D.R.; Knipe, D.M.; Tyler, K.L.; Schaffer, P.A. Immediate-Early Regulatory Gene Mutants Define Different Stages in the Establishment and Reactivation of Herpes Simplex Virus Latency. J. Virol. 1986, 63, 759–768. [Google Scholar] [CrossRef]
- Cai, W.; Schaffer, P.A. A Cellular Function Can Enhance Gene Expression and Plating Efficiency of a Mutant Defective in the Gene for ICP0, a Transactivating Protein of Herpes Simplex Virus Type 1. J. Virol. 1991, 65, 4078–4090. [Google Scholar] [CrossRef]
- Everett, R.D. Construction and Characterization of Herpes Simplex Virus Type 1 Mutants with Defined Lesions in Immediate-Early Gene 1. J. Gen. Virol. 1989, 70, 1185–1202. [Google Scholar] [CrossRef] [PubMed]
- Sacks, W.R.; Schaffer, P.A. Deletion Mutants in the Gene Encoding the Herpes Simplex Virus Type 1 Immediate-Early Protein ICP0 Exhibit Impaired Growth in Cell Culture. J. Virol. 1987, 61, 829–839. [Google Scholar] [CrossRef] [PubMed]
- Kukhanova, M.K.; Korovina, A.N.; Kochetkov, S.N. Human Herpes Simplex Virus: Life Cycle and Development of Inhibitors. Biochemistry 2014, 79, 1635–1652. [Google Scholar] [CrossRef]
- Field, H.J. Pathogenicity of Herpes Simplex Virus Mutants Containing Drug Resistance Mutations in the Viral DNA Polymerase Gene. J. Virol. 1986, 60, 286–289. [Google Scholar] [CrossRef] [PubMed]
- Field, H.J. Persistent Herpes Simplex Virus Infection and Mechanisms of Virus Drug Resistance. Eur. J. Clin. Microbiol. Infect. Dis. 1989, 8, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.P.; Sasse, F.; Brönstrup, M.; Diez, J.; Meyerhans, A. Antiviral Drug Discovery: Broad-Spectrum Drugs from Nature. Nat. Prod. Rep. 2015, 32, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Lee, I.S.; Mar, W. Inhibition of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Activity by 1,2,3,4,6-Penta-O-galloyl-beta-D-glucose in Murine Macrophage Cells. Arch. Pharm. Res. 2003, 26, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.T.; Kang, S.C.; Song, Y.J. The Extract of Elaeocarpus sylvestris Inhibits Human Cytomegalovirus Immediate Early Gene Expression and Replication In Vitro. Mol. Med. Rep. 2014, 9, 744–748. [Google Scholar]
- Okuda, T. Geraniin, a New Ellagitannin from Geranium thunbergii. Tetrahedron Lett. 1976, 17, 3721–3722. [Google Scholar] [CrossRef]
- Perera, A.; Ton, S.H.; Palanisamy, U.D. Perspectives on Geraniin, a Multifunctional Natural Bioactive Compound. Trends Food Sci. Technol. 2015, 44, 243–257. [Google Scholar] [CrossRef]
- Taguri, T.; Tanaka, T.; Kouno, I. Antimicrobial Activity of 10 Different Plant Polyphenols against Bacteria Causing Food-Borne Disease. Biol. Pharm. Bull. 2004, 27, 1965–1969. [Google Scholar] [CrossRef] [PubMed]
- Ito, H. Metabolites of the Ellagitannin Geraniin and Their Antioxidant Activities. Planta Med. 2011, 77, 1110–1115. [Google Scholar] [CrossRef]
- Kang, K.A.; Lee, I.K.; Zhang, R.; Piao, M.J.; Kim, K.C.; Kim, S.Y.; Shin, T.; Kim, B.J.; Lee, N.H.; Hyun, J.W. Radioprotective Effect of Geraniin via the Inhibition of Apoptosis Triggered by γ-Radiation-Induced Oxidative Stress. Cell Biol. Toxicol. 2011, 27, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.T.; Chang, S.S.; Hsu, F.L. Antihypertensive Action of Geraniin in Rats. J. Pharm. Pharmacol. 1994, 46, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Kashiwada, Y.; Nonaka, G.I.; Nishioka, I.; Chang, J.-J.; Lee, K.-H. Antitumor Agents, 129. Tannins and Related Compounds as Selective Cytotoxic Agents. J. Nat. Prod. 1992, 55, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Miguel, O.G.; Calixto, J.B.; Santos, A.R.; Messana, I.; Ferrari, F.; Cechinel Filho, V.; Pizzolatti, M.G.; Yunes, R.A. Chemical and Preliminary Analgesic Evaluation of Geraniin and Furosin Isolated from Phyllanthus sellowianus. Planta Med. 1996, 62, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Notka, F.; Meier, G.R.; Wagner, R. Inhibition of Wild-Type Human Immunodeficiency Virus and Reverse Transcriptase Inhibitor-Resistant Variants by Phyllanthus amarus. Antiviral Res. 2003, 58, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, H.; Zhou, W.; Feng, M.; Zhou, P. Anti-Hepatitis B Virus Activities of Geranium caroliniaum L. Extracts and Identification of the Active Components. Biol. Pharm. Bull. 2008, 31, 743–747. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Cheng, H.Y.; Lin, T.C.; Chiang, L.C.; Lin, C.C. The In Vitro Activity of Geraniin and 1,3,4,6-Tetra-O-galloyl-beta-D-glucose Isolated from Phyllanthus urinaria against Herpes Simplex Virus Type 1 and Type 2 Infection. J. Ethnopharmacol. 2007, 110, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Hsiung, C.D. Virus Assay and Neutralization Test. Diagnostic Virology; Yale University Press: New Haven, CT, USA, 1982; pp. 25–35. [Google Scholar]
- Hierholzer, J.C.; Killington, R.A. Virus Isolation and Quantitation. In Virology Methods Manual; Academic Press: Cambridge, MA, USA, 1996; pp. 25–46. [Google Scholar]
- Joo, Y.H.; Lee, Y.G.; Lim, Y.; Jeon, H.; Kim, E.H.; Choi, J.; Hong, W.; Jeon, H.; Ahrweiler, M.; Kim, H.; et al. Potent antiviral activity of the extract of Elaeocarpus sylvestris against influenza A virus in vitro and in vivo. Phytomedicine 2022, 97, 153892. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| EtOH (%) | 0 | 30 | 50 | 70 | 100 |
|---|---|---|---|---|---|
| EC50 1 | 143.8 ± 2.51 | 42.6 ± 0.85 | 23.2 ± 0.22 | 28.5 ± 0.37 | 63.8 ± 1.82 |
| IC50 2 | >500 | 358.5 ± 2.37 | 342.8 ± 2.59 | 209.4 ± 3.61 | 184.3 ± 1.68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-G.; Park, D.W.; Kwon, J.E.; Kim, H.; Kang, S.C. Elaeocarpus sylvestris var. ellipticus Extract and Its Major Component, Geraniin, Inhibit Herpes Simplex Virus-1 Replication. Plants 2024, 13, 1437. https://doi.org/10.3390/plants13111437
Lee Y-G, Park DW, Kwon JE, Kim H, Kang SC. Elaeocarpus sylvestris var. ellipticus Extract and Its Major Component, Geraniin, Inhibit Herpes Simplex Virus-1 Replication. Plants. 2024; 13(11):1437. https://doi.org/10.3390/plants13111437
Chicago/Turabian StyleLee, Yeong-Geun, Dae Won Park, Jeong Eun Kwon, Hyunggun Kim, and Se Chan Kang. 2024. "Elaeocarpus sylvestris var. ellipticus Extract and Its Major Component, Geraniin, Inhibit Herpes Simplex Virus-1 Replication" Plants 13, no. 11: 1437. https://doi.org/10.3390/plants13111437
APA StyleLee, Y.-G., Park, D. W., Kwon, J. E., Kim, H., & Kang, S. C. (2024). Elaeocarpus sylvestris var. ellipticus Extract and Its Major Component, Geraniin, Inhibit Herpes Simplex Virus-1 Replication. Plants, 13(11), 1437. https://doi.org/10.3390/plants13111437