
Effect of Arbuscular Mycorrhizal Fungi on Nitrogen and Phosphorus Uptake Efficiency and Crop Productivity of Two-Rowed Barley under Different Crop Production Systems
 , ,
, ,  ,
,  ,
,
Abstract
1. Introduction
2. Results
2.1. Growth and Productivity of Two-Rowed Barley
2.2. Nitrogen (N) and Phosphorus (P) Content, Uptake and Use Efficiency of Two-Rowed Barley
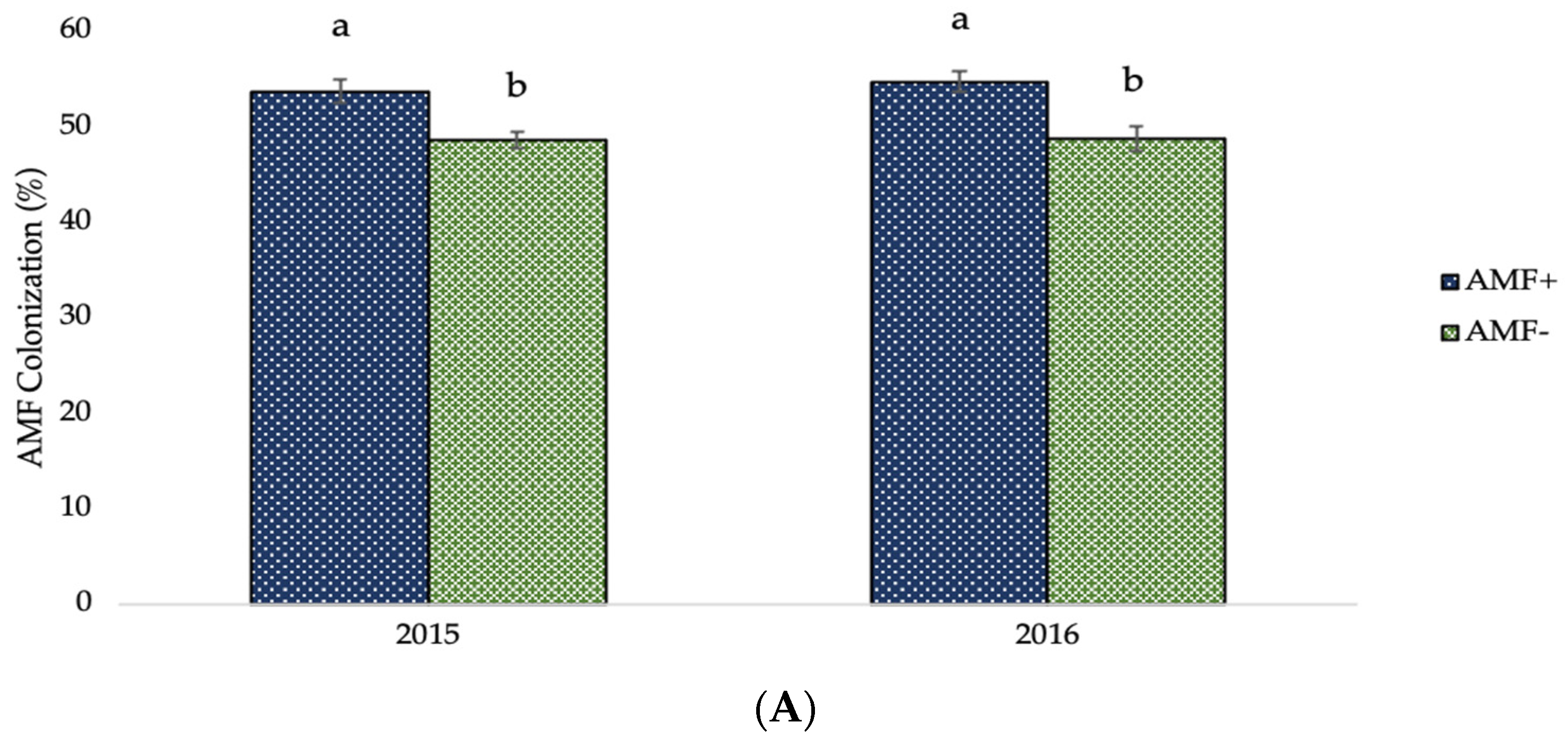
2.3. Arbuscular Mycorrhizal Fungi (AMF) Colonization and Weighted Mycorrhizal Dependency (WMD) of Two-Rowed Barley
3. Discussion
4. Materials and Methods
4.1. Site Description and Experimental Design
4.2. Sampling, Measurements and Methods
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miransari, M. Interactions between arbuscular mycorrhizal fungi and soil bacteria. Appl. Microbiol. Biotechnol. 2011, 89, 917–930. [Google Scholar] [CrossRef]
- Sbrana, C.; Avio, L.; Giovannetti, M. Beneficial mycorrhizal symbionts affecting the production of health-promoting phytochemicals. Electrophoresis 2014, 35, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R.; Rillig, M.C. Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol. 2018, 220, 1059–1075. [Google Scholar] [CrossRef]
- Feddermann, N.; Finlay, R.; Boller, T.; Elfstrand, M. Functional diversity in arbuscular mycorrhiza—The role of gene expression, phosphorous nutrition and symbiotic efficiency. Fungal Ecol. 2010, 3, 1–8. [Google Scholar] [CrossRef]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed]
- Menéndez, A.B.; Scervino, J.M.; Godeas, A.M. Arbuscular mycorrhizal populations associated with natural and cultivated vegetation on a site of Buenos Aires province, Argentina. Biol. Fertil. Soils 2001, 33, 373–381. [Google Scholar] [CrossRef]
- Manoharan, L.; Rosenstock, N.P.; Williams, A.; Hedlund, K. Agricultural management practices influence AMF diversity and community composition with cascading effects on plant productivity. Appl. Soil Ecol. 2017, 115, 53–59. [Google Scholar] [CrossRef]
- Kogelmann, W.J.; Lin, H.S.; Bryant, R.B.; Beege, D.B.; Wolf, A.M.; Petersen, G.W. A statewide assessment of the impacts of phosphorus index implementation in Pennsylvania. J. Soil Water Conserv. 2004, 59, 9–18. [Google Scholar]
- Bach, I.-M.; Essich, L.; Bauerle, A.; Müller, T. Efficiency of phosphorus fertilizers derived from recycled biogas digestate as applied to maize and ryegrass in soils with different pH. Agriculture 2022, 12, 325. [Google Scholar] [CrossRef]
- Zhu, C.; Ling, N.; Guo, J.; Wang, M.; Guo, S.; Shen, Q. Impacts of fertilization regimes on arbuscular mycorrhizal fungal (AMF) community composition were correlated with organic matter composition in maize rhizosphere soil. Front. Microbiol. 2016, 7, 1840. [Google Scholar] [CrossRef]
- Galvez, L.; Douds, D.D.; Drinkwater, L.E.; Wagoner, P. Effect of tillage and farming system upon VAM fungus populations and mycorrhizas and nutrient uptake of maize. Plant Soil 2001, 118, 299–308. [Google Scholar] [CrossRef]
- Yang, H.S.; Zhang, Q.; Dai, Y.J.; Liu, Q.; Tang, J.J.; Bian, X.M.; Chen, X. Effects of arbuscular mycorrhizal fungi on plant growth depend on root system: A meta-analysis. Plant Soil 2015, 389, 361–374. [Google Scholar] [CrossRef]
- Thirkell, T.J.; Charters, M.D.; Elliott, A.J.; Sait, S.M.; Field, K.J. Are mycorrhizal fungi our sustainable saviours? Considerations for achieving food security. J. Ecol. 2017, 105, 921–929. [Google Scholar] [CrossRef]
- Watts-Williams, S.J.; Gilbert, S.E. Arbuscular mycorrhizal fungi affect the concentration and distribution of nutrients in the grain differently in barley compared with wheat. Plants People Planet 2021, 3, 567–577. [Google Scholar] [CrossRef]
- Vosatka, M.; Dodd, J.C. Ecological considerations for successful application of arbuscular mycorrhizal fungi inoculum. In Mycorrhizal Technology in Agriculture; Gianinazzi, S., Schüepp, H., Barea, J.M., Haselwandter, K., Eds.; Birkhäuser Verlag: Basel, Switzerland, 2002. [Google Scholar]
- Jin, L.; Wang, Q.; Wang, Q.; Wang, X.J.; Gange, A.C. Mycorrhizal-induced growth depression in plants. Symbiosis 2017, 72, 81–88. [Google Scholar] [CrossRef]
- Sun, R.-T.; Zhang, Z.-Z.; Zhou, N.; Srivastava, A.; Kuča, K.; Abd-Allah, E.F.; Hashem, A.; Wu, Q.-S. A review of the interaction of medicinal plants and arbuscular mycorrhizal fungi in the rhizosphere. Not. Bot. Horti Agrobot. Cluj Napoca 2021, 49, 12454. [Google Scholar] [CrossRef]
- Kahiluoto, H.; Ketoja, E.; Vestberg, M. Contribution of arbuscular mycorrhiza to soil quality in contrasting cropping systems. Agric. Ecosyst. Environ. 2009, 134, 36–45. [Google Scholar] [CrossRef]
- Gosling, P.; Hodge, A.; Goodlass, G.; Bending, G.D. Arbuscular mycorrhizal fungi and organic farming. Agric. Ecosyst. Environ. 2006, 113, 17–35. [Google Scholar] [CrossRef]
- Koide, R.T.; Mosse, B. A history of research on arbuscular mycorrhiza. Mycorrhiza 2004, 14, 145–163. [Google Scholar] [CrossRef]
- Ho-Plágaro, T.; García-Garrido, J.M. Molecular regulation of arbuscular mycorrhizal symbiosis. Int. J. Mol. Sci. 2022, 23, 5960. [Google Scholar] [CrossRef]
- Ma, J.; Wang, W.; Yang, J.; Qin, S.; Yang, Y.; Sun, C.; Pei, G.; Zeeshan, M.; Liao, H.; Liu, L.; et al. Mycorrhizal symbiosis promotes the nutrient content accumulation and affects the root exudates in maize. BMC Plant Biol. 2022, 22, 64. [Google Scholar] [CrossRef] [PubMed]
- Sabra, M.; Aboulnasr, A.; Franken, P.; Perreca, E.; Wright, L.P.; Camehl, I. Beneficial root endophytic fungi increase growth and quality parameters of sweet basil in heavy metal contaminated soil. Front. Plant Sci. 2018, 9, 1726. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Zhang, M.; Wang, M.; Li, Y.; Liu, J.; Chen, Y. Inoculation with arbuscular mycorrhizal fungus modulates defense-related genes expression in banana seedlings susceptible to wilt disease. Plant Signal. Behav. 2021, 16, 1884782. [Google Scholar] [CrossRef] [PubMed]
- Jumrani, K.; Bhatia, V.S.; Kataria, S.; Alamri, S.A.; Siddiqui, M.H.; Rastogi, A. Inoculation with arbuscular mycorrhizal fungi alleviates the adverse effects of high temperature in soybean. Plants 2022, 11, 2210. [Google Scholar] [CrossRef]
- Gao, Y.; Huang, S.; Wang, Y.; Lin, H.; Pan, Z.; Zhang, S.; Zhang, J.; Wang, W.; Cheng, S.; Chen, Y. Analysis of the molecular and biochemical mechanisms involved in the symbiotic relationship between Arbuscular mycorrhiza fungi and Manihot esculenta Crantz. Front. Plant Sci. 2023, 14, 1130924. [Google Scholar] [CrossRef]
- Abdallah, C.; Valot, B.; Guillier, C.; Mounier, A.; Balliau, T.; Zivy, M.; van Tuinen, D.; Renaut, J.; Wipf, D.; Dumas-Gaudot, E.; et al. The membrane proteome of Medicago truncatula roots displays qualitative and quantitative changes in response to arbuscular mycorrhizal symbiosis. J. Proteomics 2014, 108, 354–368. [Google Scholar] [CrossRef]
- Mitra, D.; Valizadeh, K.; Chaudhary, P.; Ruparelia, J.; Smruthi, M.; Boutaj, H.; Das Mohapatra, P.K.; Panneerselvam, P. Involvement of strigolactone hormone in root development, influence and interaction with mycorrhizal fungi in plant: Mini-review. Curr. Res. Microb. Sci. 2021, 2, 100026. [Google Scholar] [CrossRef]
- Mishra, S.; Priyanka; Sharma, S. Metabolomic insights into endophyte-derived bioactive compounds. Front. Microbiol. 2022, 13, 835931. [Google Scholar] [CrossRef]
- Tsiokanos, E.; Cartabia, A.; Tsafantakis, N.; Lalaymia, I.; Termentzi, A.; Miguel, M.; Declerck, S.; Fokialakis, N. The metabolic profile of Anchusa officinalis L. differs according to its associated arbuscular mycorrhizal fungi. Metabolites 2022, 12, 573. [Google Scholar] [CrossRef]
- Preiti, G.; Calvi, A.; Romeo, M.; Badagliacca, G.; Bacchi, M. Seeding density and nitrogen fertilization effects on agronomic responses of some hybrid barley lines in a Mediterranean environment. Agronomy 2021, 11, 1942. [Google Scholar] [CrossRef]
- Bardehji, S.; Eshghizadeh, H.; Zahedi, M.; Sabzalian, M.; Gheisari, M. The combined effect of nitrogen fertilizer and sowing season on response to water-limited stress in barley (Hordeum vulgare L.). J. Agric. Sci. 2021, 159, 31–49. [Google Scholar] [CrossRef]
- Beslemes, D.; Tigka, E.; Roussis, I.; Kakabouki, I.; Mavroeidis, A.; Vlachostergios, D. Contribution of arbuscular mycorrhizal fungi to nitrogen and phosphorus uptake efficiency and productivity of faba bean crop on contrasting cropping systems. Not. Bot. Horti Agrobot. Cluj Napoca 2022, 50, 12806. [Google Scholar] [CrossRef]
- Feng, G.; Li, X.; Zhang, F.; Li, S. Effect of AM fungi on water and nutrition status of corn plants under salt stress. Chin. J. Appl. Ecol. 2000, 11, 595–598. [Google Scholar]
- Felföldi, Z.; Vidican, R.; Stoian, V.; Roman, I.A.; Sestras, A.F.; Rusu, T.; Sestras, R.E. Arbuscular mycorrhizal fungi and fertilization influence yield, growth and root colonization of different tomato genotype. Plants 2022, 11, 1743. [Google Scholar] [CrossRef]
- Gao, X.; Guo, H.; Zhang, Q.; Guo, H.; Zhang, L.; Zhang, C.; Gou, Z.; Liu, Y.; Wei, J.; Chen, A.; et al. Arbuscular mycorrhizal fungi (AMF) enhanced the growth, yield, fiber quality and phosphorus regulation in upland cotton (Gossypium hirsutum L.). Sci. Rep. 2020, 10, 2084. [Google Scholar] [CrossRef]
- Trisilawati, O.; Hartoyo, B.; Bermawie, N.; Pribadi, E.R. Application of AMF (Arbuscular Mycorrhizal Fungi) and organic fertilizer to increase the growth, biomass and bioactive content of Centella. IOP Conf. Ser. Earth Environ. Sci. 2019, 292, 012067. [Google Scholar] [CrossRef]
- Jadhav, D.N.; Patil, M.S. Effect of AM inoculation on growth and NPK uptake of groundnut. Acta Agron. Hung. 1996, 44, 153–159. [Google Scholar]
- Bi, Y.; Zhou, H. Changes in peanut canopy structure and photosynthetic characteristics induced by an arbuscular mycorrhizal fungus in a nutrient-poor environment. Sci. Rep. 2021, 11, 14832. [Google Scholar] [CrossRef]
- Evans, J. Nitrogen and photosynthesis in the flag leaf of wheat (Triticum aestivum L.). Plant Physiol. 1983, 72, 297–302. [Google Scholar] [CrossRef]
- Kakabouki, I.P.; Roussis, I.; Hela, D.; Papastylianou, P.; Folina, A.; Bilalis, D. Root growth dynamics and productivity of quinoa (Chenopodium quinoa Willd.) in response to fertilization and soil tillage. Folia Hortic. 2019, 31, 277–291. [Google Scholar] [CrossRef]
- Jerbi, M.; Labidi, S.; Laruelle, F.; Tisserant, B.; Jeddi, F.B.; Sahraoui, A.L.H. Mycorrhizal biofertilization improves grain yield and quality of hulless Barley (Hordeum vulgare ssp. nudum L.) under water stress conditions. J. Cereal Sci. 2022, 104, 103436. [Google Scholar] [CrossRef]
- Masrahi, A.S.; Alasmari, A.; Shahin, M.G.; Qumsani, A.T.; Oraby, H.F.; Awad-Allah, M.M.A. Role of arbuscular mycorrhizal fungi and phosphate solubilizing bacteria in improving yield, yield components, and nutrients uptake of barley under salinity soil. Agriculture 2023, 13, 537. [Google Scholar] [CrossRef]
- Lekberg, Y.; Koide, R.T. Is plant performance limited by abundance of arbuscular mycorrhizal fungi? a meta-analysis of studies published between 1988 and 2003. New Phytol. 2005, 168, 189–204. [Google Scholar] [CrossRef]
- Pellegrino, E.; Öpik, M.; Bonari, E.; Ercoli, L. Responses of wheat to arbuscular mycorrhizal fungi: A meta-analysis of field studies from 1975 to 2013. Soil Biol. Biochem. 2015, 84, 210–217. [Google Scholar] [CrossRef]
- Habiyaremye, C.; Schroeder, K.L.; Reganold, J.P.; White, D.; Packer, D.; Murphy, K.M. Effect of nitrogen and seeding rate on β-glucan, protein, and grain yield of naked food barley in no-till cropping systems in the Palouse region of the Pacific Northwest. Front. Sustain. Food Syst. 2021, 5, 663445. [Google Scholar] [CrossRef]
- Bilalis, D.; Karkanis, A.; Sidiras, N.; Travlos, I.; Efthimiadou, A.; Thomopoulos, P.; Kakabouki, I. Maize and legumes root growth and yield as influenced by organic fertilization, under Mediterranean environmental conditions. Rom. Agric. Res. 2012, 29, 211–217. [Google Scholar]
- Shepherd, K.; Cooper, P.; Allan, A.; Drennan, D.; Keatinge, J. Growth, water use and yield of barley in Mediterranean-type environments. J. Agric. Sci. 1987, 108, 365–378. [Google Scholar] [CrossRef]
- Plaza-Bonilla, D.; Lampurlanés, J.; Fernández, F.G.; Cantero-Martínez, C. Nitrogen fertilization strategies for improved Mediterranean rainfed wheat and barley performance and water and nitrogen use efficiency. Eur. J. Agron. 2021, 124, 126238. [Google Scholar] [CrossRef]
- Gallais, A.; Hirel, B. An approach to the genetics of nitrogen use efficiency in maize. J. Exp. Bot. 2004, 55, 295–306. [Google Scholar] [CrossRef]
- Vos, J.; Putten, P.E.L.; Birch, C.J. Effect of nitrogen supply on leaf appearance, leaf nitrogen economy and photosynthetic maize (Zea mays L.). Field Crop. Res. 2005, 93, 64–73. [Google Scholar] [CrossRef]
- Liu, Y.J.; Shi, G.X.; Mao, L.; Cheng, G.; Jiang, S.J.; Ma, X.J.; An, L.; Du, G.; Johnson, N.C.; Feng, H.Y. Direct and indirect influences of 8 yr of nitrogen and phosphorus fertilization on Glomeromycota in an alpine meadow ecosystem. New Phytol. 2012, 194, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Matson, P.A.; Parton, W.J.; Power, A.G.; Swift, M.J. Agricultural intensification and ecosystem properties. Science 1997, 277, 504–509. [Google Scholar] [CrossRef] [PubMed]
- McSwiney, C.P.; Robertson, G.P. Nonlinear response of N2O flux to incremental fertilizer addition in a continuous maize (Zea mays L.) cropping system. Glob. Chang. Biol. 2005, 11, 1712–1719. [Google Scholar] [CrossRef]
- Huber, D.M.; Watson, R.D. Nitrogen form and plant disease. Annu. Rev. Phytopathol. 1974, 12, 139–165. [Google Scholar] [CrossRef] [PubMed]
- Paustian, K.; Robertson, G.P.; Elliott, E.T. Management impacts on carbon storage and gas fluxes (CO2, CH4) in mid-latitude cropland ecosystems. In Soil Management and Greenhouse Effect; Lal, R., Kimble, J., Levine, E., Stewart, B.A., Eds.; CRC Press: Boca Raton, FL, USA, 1995; pp. 69–84. [Google Scholar]
- Dordas, C. Dry matter, nitrogen and phosphorus accumulation, partitioning and remobilization as affected by N and P fertilization and source–sink relation. Eur. J. Agron. 2009, 30, 129–139. [Google Scholar] [CrossRef]
- Ayadi, S.; Karmous, C.; Chamekh, Z.; Hammami, Z.; Baraket, M.; Esposito, S.; Rezgui, S.; Trifa, Y. Effects of nitrogen rates on grain yield and nitrogen agronomic efficiency of durum wheat genotypes under different environments. Ann. Appl. Biol. 2016, 168, 264–273. [Google Scholar] [CrossRef]
- Hodge, A.; Campbell, C.D.; Fitter, A.H. An arbuscular mycorrhizal fungus accelerates decomposition and acquires nitrogen directly from organic material. Nature 2001, 413, 297–299. [Google Scholar] [CrossRef]
- Barrett, G.; Campbell, C.D.; Fitter, A.H.; Hodge, A. The arbuscular mycorrhizal fungus Glomus hoi can capture and transfer nitrogen from organic patches to its associated host plant at low temperature. Appl. Soil Ecol. 2011, 48, 102–105. [Google Scholar] [CrossRef]
- Smith, F.A.; Smith, S.E. Mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef]
- Govindarajulu, M.; Pfeffer, P.E.; Jin, H.; Abubaker, J.; Douds, D.D.; Allen, J.W.; Bücking, H.; Lammers, P.J.; Shachar-Hill, Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 2005, 435, 819–823. [Google Scholar] [CrossRef]
- Leigh, J.; Hodge, A.; Fitter, A.H. Arbuscular mycorrhizal fungi can transfer substantial amounts of nitrogen to their host plant from organic material. New Phytol. 2009, 181, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Barrett, G.; Campbell, C.D.; Hodge, A. The direct response of the external mycelium of arbuscular mycorrhizal fungi to temperature and the implications for nutrient transfer. Soil Biol. Biochem. 2014, 78, 109–117. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R.; Glick, B.R. Contribution of arbuscular mycorrhizal fungi, phosphate–solubilizing bacteria, and silicon to P uptake by plant. Front. Plant Sci. 2021, 12, 699618. [Google Scholar] [CrossRef] [PubMed]
- Bulman, P.; Smith, D.L. Post-heading nitrogen uptake, retranslocation, and partitioning in spring barley. Crop Sci. 1994, 34, 977–984. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C. Enhancing nitrogen use efficiency in crop plants. Adv. Agron. 2005, 88, 97–185. [Google Scholar]
- Najafi, A.; Ardakani, M.R.; Rejali, F.; Sajedi, N. Response of winter barley to co-inoculation with Azotobacter and Mycorrhiza fungi influenced by plant growth promoting rhizobacteria. Ann. Biol. Res. 2012, 3, 4002–4006. [Google Scholar]
- Raklami, A.; Bechtaoui, N.; Tahiri, A.; Anli, M.; Meddich, A.; Oufdou, K. Use of rhizobacteria and mycorrhizae consortium in the open field as a strategy for improving crop nutrition, productivity and soil fertility. Front. Microbiol. 2019, 10, 1106. [Google Scholar] [CrossRef]
- Berhanu, G.; Kismányoky, T.; Sárdi, K. Effect of nitrogen fertilization and residue management on the productivity of winter barley (Hordeum vulgare L.). Acta Agron. Hung. 2013, 61, 101–111. [Google Scholar] [CrossRef]
- Golubkina, N.A.; Amagova, Z.; Matsadze, V.; Zamana, S.; Tallarita, A.; Caruso, G. Effects of arbuscular mycorrhizal fungi on yield, biochemical characteristics, and elemental composition of garlic and onion under selenium supply. Plants 2020, 9, 84. [Google Scholar] [CrossRef]
- Fernández Bidondo, L.; Bompadre, J.; Pergola, M.; Silvani, V.; Colombo, R.; Bracamonte, F.; Godeas, A. Differential interaction between two Glomus intraradices strains and a phosphate solubilizing bacterium in maize rhizosphere. Pedobiologia 2012, 55, 227–232. [Google Scholar] [CrossRef]
- Heydari, M.M.; Maleki, A. Effect of phosphorus sources and mycorrhizal inoculation on root colonization and phosphorus uptake of barley (Hordeum vulgare L.). Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 235–248. [Google Scholar]
- Nell, M.; Wawrosch, C.; Steinkellner, S.; Vierheilig, H.; Kopp, B.; Lössl, A. Root colonization by symbiotic arbuscular mycorrhizal fungi increases sesquiterpenic acid concentrations in Valeriana officinalis L. Planta Med. 2010, 76, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Joner, E.J.; Jakobsen, I. Growth and extracellular phosphatase activity of arbuscular mycorrhizal hyphae as influenced by soil organic matter. Soil Biol. Biochem. 1995, 27, 1153–1159. [Google Scholar] [CrossRef]
- Schweiger, P.F.; Jakobsen, I. The role of mycorrhizas in plant P nutrition: Fungal uptake kinetics and genotype variation. In Plant Nutrition—Molecular Biology and Genetics; Gissel-Nielsen, G., Jensen, A., Eds.; Kluwer: Dordrecht, The Netherlands, 1999; pp. 277–289. [Google Scholar]
- Galleguillos, C.; Aguirre, C.; Barea, J.M.; Azcon, R. Growth promoting effect of two Sinorhizobium meliloti strains (a wild type and its genetically modified derivative) on a non-legume plant species in specific interaction with two arbuscular mycorrhizal fungi. Plant Sci. 2000, 159, 57–63. [Google Scholar] [CrossRef]
- Lopez-Bellido, R.J.; Lopez-Bellido, L.; Benitez-Vega, J.; Lopez-Bellido, F.J. Tillage system, preceding crop and nitrogen fertilizer in wheat crop: Water utilization. Agron. J. 2007, 99, 66–72. [Google Scholar] [CrossRef]
- Muchow, R.C.; Davis, R. Effect of nitrogen supply on the comparative productivity of maize and sorghum in a semi-arid tropical environment II. Radiation interception and biomass accumulation. Field Crops Res. 1988, 18, 17–30. [Google Scholar] [CrossRef]
- Ryan, M.H.; Graham, J.H. Is there a role for arbuscular mycorrhizal fungi in production agriculture? Plant Soil 2002, 244, 263–271. [Google Scholar] [CrossRef]
- Sorensen, N.; Larsen, J.; Jakobsen, I. Mycorrhiza formation and nutrient concentration in leeks (Allium porrum) in relation to previous crop and cover crop management on high P soils. Plant Soil 2005, 273, 101–114. [Google Scholar] [CrossRef]
- Kahiluoto, H.; Ketoja, E.; Vestberg, M.; Saarela, I. Promotion of AM utilization through reduced P fertilization. 2. Field studies. Plant Soil 2001, 231, 65–79. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; Dezonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Ye, Q.; Zhang, H.; Wei, H.; Zhang, Y.; Wang, B.; Xia, K.; Huo, Z.; Dai, Q.; Xu, K. Effects of nitrogen fertilizer on nitrogen use efficiency and yield of rice under different soil conditions. Front. Agric. China 2007, 1, 30–36. [Google Scholar] [CrossRef]
- Kormanik, P.P.; McGraw, A.C. Quantification of vesicular-arbuscular mycorrhizae in plant roots. In Methods and Principles of Mycorrhizal Research; Schenck, N.C., Ed.; American Phytopathological Society: St. Paul, MN, USA, 1982; pp. 37–45. [Google Scholar]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular-arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Plenchette, C.; Fortin, J.A.; Furlan, V. Growth response of several plant species to mycorrhizae in a soil of moderate P fertility. I. Mycorrhizal dependency under field conditions. Plant Soil 1983, 70, 199–209. [Google Scholar] [CrossRef]
- Karagiannidis, N.; Thomidis, T.; Lazari, D.; Panou-Filotheou, E.; Karagiannidou, C. Effect of three Greek arbuscular mycorrhizal fungi in improving the growth, nutrient concentration, and production of essential oils of oregano and mint plants. Sci. Hort. 2011, 129, 329–334. [Google Scholar] [CrossRef]
- Chatzistathis, T.; Orfanoudakis, M.; Alifragis, D.; Therios, I. Colonization of Greek olive cultivars’ root system by arbuscular mycorrhiza fungus: Root morphology, growth, and mineral nutrition of olive plants. Sci. Agric. 2013, 70, 185–194. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variance | Df | Plant Height | Leaf Area Index (LAI) | Biomass Yield | Seed Yield | Harvest Index (HI) | Biomass Nitrogen (N) Uptake |
| Year (Y) | 1 | 90.039 *** | 1.371 ns | 7.757 ** | 20.061 *** | 0.006 ns | 0.088 ns |
| Inoculation (I) | 1 | 117.121 *** | 12.974 *** | 35.021 *** | 52.184 *** | 2.888 ns | 6.079 * |
| Fertilization (F) | 4 | 157.668 *** | 14.406 *** | 59.011 *** | 114.229 *** | 1.606 ns | 9.976 *** |
| Y × I | 1 | 0.024 ns | 0.027 ns | 0.009 ns | 0.010 ns | 0.001 ns | 0.007 ns |
| Y × F | 4 | 0.052 ns | 0.004 ns | 0.017 ns | 0.033 ns | 0.001 ns | 0.443 ns |
| I × F | 4 | 47.888 *** | 19.433 *** | 1.735 ns | 5.113 ** | 1.016 ns | 2.459 ns |
| Y × I × F | 4 | 0.015 ns | 0.004 ns | 0.002 ns | 0.020 ns | 0.002 ns | 0.313 ns |
| Source of Variance | Df | Seed N Uptake | Total Plant N Uptake | Nitrogen Harvest Index (NHI) | Biomass Phosphorus (P) Uptake | Seed P Uptake | Total Plant P Uptake |
| Year (Y) | 1 | 3.654 ns | 2.619 ns | 0.151 ns | 1.630 ns | 15.651 *** | 9.813 ** |
| Inoculation (I) | 1 | 4.893 * | 9.108 ** | 2.395 ns | 71.370 *** | 19.116 *** | 68.393 *** |
| Fertilization (F) | 4 | 31.069 *** | 33.613 *** | 2.906 * | 19.580 *** | 112.141 *** | 74.012 *** |
| Y × I | 1 | 0.001 ns | 0.001 ns | 0.024 ns | 0.271 ns | 1.121 ns | 0.918 ns |
| Y × F | 4 | 0.009 ns | 0.181 ns | 0.498 ns | 0.039 ns | 0.209 ns | 0.156 ns |
| I × F | 4 | 1.893 ns | 2.451 ns | 2.006 ns | 1.259 ns | 7.279 *** | 2.671 * |
| Y × I × F | 4 | 0.004 ns | 0.106 ns | 0.356 ns | 0.053 ns | 0.180 ns | 0.022 ns |
| Source of Variance | Df | Phosphorus Harvest Index (PHI) | Nitrogen Utilization Efficiency (NUtE) | Phosphorus Utilization Efficiency (PUtE) | AMF Colonization | Weighted Mycorrhizal Dependency (WMD) | |
| Year (Y) | 1 | 0.010 ns | 0.048 ns | 0.042 ns | 0.658 ns | 0.322 ns | |
| Inoculation (I) | 1 | 52.509 *** | 0.008 ns | 14.564 *** | 63.434 *** | - | |
| Fertilization (F) | 4 | 7.628 *** | 3.662 * | 6.479 *** | 21.626 *** | 7.229 *** | |
| Y × I | 1 | 0.006 ns | 0.009 ns | 0.848 ns | 0.434 ns | - | |
| Y × F | 4 | 0.014 ns | 0.172 ns | 0.099 ns | 0.510 ns | 0.472 ns | |
| I × F | 4 | 1.783 ns | 3.486 * | 8.657 *** | 4.426 ** | - | |
| Y × I × F | 4 | 0.174 ns | 0.117 ns | 0.011 ns | 0.584 ns | - | |
| Inoculation | Fertilization | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2015 | 2016 | ||||||||||||
| 100% Org. | 60% Org. | 100% Inorg. | 60% Inorg. | Control | Mean | 100% Org. | 60% Org. | 100% Inorg. | 60% Inorg. | Control | Mean | ||
| Plant Height (cm) | AMF+ | 84.2 a | 79.1 b | 84.8 a | 79.0 b | 77.7 b | 80.9 A | 87.2 a | 81.9 b | 87.8 a | 81.9 b | 80.5 b | 83.9 A |
| AMF− | 77.5 c | 80.1 b | 84.1 a | 78.8 bc | 68.3 d | 77.8 B | 80.3 c | 82.9 b | 87.1 a | 81.7 bc | 70.8 d | 80.6 B | |
| LSD† | *** | * | ns | ns | *** | *** | ns | ns | ns | *** | |||
| LAI (m2 m−2) | AMF+ | 4.39 a | 3.35 b | 3.54 ab | 3.22 b | 3.17 b | 3.53 A | 4.52 a | 3.45 b | 3.63 ab | 3.31 b | 3.25 b | 3.63 A |
| AMF− | 2.95 b | 3.15 b | 4.04 a | 3.33 b | 2.91 b | 3.28 B | 3.02 b | 3.22 b | 4.14 a | 3.40 b | 2.98 b | 3.35 B | |
| LSD† | ** | ns | ns | * | ns | ** | ns | ns | * | ns | |||
| Biomass Yield (tn ha−1) | AMF+ | 7.66 a | 7.35 a | 7.65 a | 6.92 ab | 6.12 b | 7.14 A | 7.95 a | 7.61 a | 7.92 a | 7.20 a | 6.34 b | 7.40 A |
| AMF− | 7.38 a | 6.86 ab | 7.34 a | 6.25 bc | 5.20 c | 6.61 B | 7.66 a | 7.11 ab | 7.61 a | 6.48 b | 5.41 c | 6.85 B | |
| LSD† | ns | ** | ns | ns | ** | ns | ** | ns | ns | * | |||
| Seed Yield (tn ha−1) | AMF+ | 4.35 a | 4.12 a | 4.33 a | 4.01 a | 3.54 b | 4.07 A | 4.52 a | 4.26 ab | 4.48 ab | 4.18 b | 3.67 c | 4.22 A |
| AMF− | 4.25 a | 4.05 a | 4.12 a | 3.62 b | 3.11 c | 3.83 B | 4.41 a | 4.21 a | 4.27 a | 3.75 b | 3.24 c | 3.98 B | |
| LSD† | * | ns | * | * | ** | * | ns | * | * | ** | |||
| Harvest Index (%) | AMF+ | 56.82 a | 56.03 a | 56.62 a | 58.04 a | 57.89 a | 57.08 A | 56.91 a | 56.00 a | 56.63 a | 58.19 a | 57.88 a | 57.12 A |
| AMF− | 57.69 a | 59.10 a | 56.20 a | 58.09 a | 59.72 a | 58.16 A | 57.75 a | 59.15 a | 56.21 a | 58.10 a | 59.89 a | 58.22 A | |
| LSD† | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | |||
| Inoculation | Fertilization | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2015 | 2016 | ||||||||||||
| 100% Org. | 60% Org. | 100% Inorg. | 60% Inorg. | Control | Mean | 100% Org. | 60% Org. | 100% Inorg. | 60% Inorg. | Control | Mean | ||
| Biomass N Uptake (kg N ha−1) | AMF+ | 23.79 a | 15.84 ab | 18.69 ab | 12.02 b | 15.25 ab | 17.12 A | 25.16 a | 16.75 bc | 19.52 ab | 14.19 bc | 10.99 c | 17.32 A |
| AMF− | 15.81 a | 15.86 a | 16.77 a | 13.42 a | 11.64 a | 14.70 B | 17.08 a | 16.17 ab | 17.14 a | 13.39 ab | 11.54 b | 15.06 B | |
| LSD† | * | ns | * | ns | ns | * | ns | * | ns | ns | |||
| Seed N Uptake (kg N ha−1) | AMF+ | 76.85 a | 68.52 ab | 74.15 a | 67.36 ab | 58.60 b | 69.09 A | 79.91 a | 70.87 ab | 76.82 a | 70.23 ab | 60.68 b | 71.70 A |
| AMF− | 71.28 a | 69.05 a | 75.56 a | 64.61 a | 50.33 b | 66.17 B | 74.00 a | 71.69 a | 78.34 a | 66.92 a | 52.51 b | 68.69 B | |
| LSD† | ns | ns | ns | ns | * | ns | ns | ns | ns | * | |||
| Total Plant N Uptake (kg N ha−1) | AMF+ | 100.64 a | 84.37 abc | 92.85 ab | 79.39 bc | 73.85 c | 86.22 A | 105.07 a | 87.62 b | 96.34 ab | 84.43 bc | 71.67 c | 89.02 A |
| AMF− | 87.09 a | 84.92 a | 92.33 a | 78.03 ab | 61.97 b | 80.87 B | 91.08 a | 87.87 a | 95.48 a | 80.31 ab | 64.05 b | 83.76 B | |
| LSD† | * | ns | ns | ns | * | * | ns | ns | ns | * | |||
| NHI (%) | AMF+ | 76.37 b | 81.17 ab | 79.97 ab | 84.80 a | 79.93 ab | 80.45 A | 76.00 b | 80.83 ab | 79.87 ab | 83.20 ab | 84.67 a | 80.91 A |
| AMF− | 82.10 a | 81.30 a | 82.10 a | 82.75 a | 81.27 a | 81.91 A | 81.57 a | 81.60 a | 82.10 a | 83.30 a | 81.97 a | 82.11 A | |
| LSD† | ns | ns | ns | ns | ns | ns | ns | ns | ns | ** | |||
| Biomass P Uptake (kg P ha−1) | AMF+ | 4.20 ab | 4.67 a | 3.94 ab | 3.58 bc | 2.94 c | 3.87 A | 4.46 ab | 4.83 a | 4.15 abc | 3.70 bc | 3.16 c | 4.06 A |
| AMF− | 3.19 ab | 3.41 a | 3.11 ab | 2.96 ab | 2.40 b | 3.01 B | 3.29 ab | 3.55 a | 3.26 ab | 3.05 ab | 2.33 b | 3.09 B | |
| LSD† | * | ** | * | ns | ns | * | ** | * | ns | * | |||
| Seed P Uptake (kg P ha−1) | AMF+ | 9.63 a | 8.16 b | 9.50 a | 7.85 b | 6.87 c | 8.40 A | 10.16 a | 8.66 b | 10.07 a | 8.33 b | 7.05 c | 8.85 A |
| AMF− | 8.62 a | 8.53 ab | 9.04 a | 7.59 bc | 6.75 c | 8.11 B | 8.90 a | 8.88 a | 9.32 a | 7.71 b | 7.01 b | 8.37 B | |
| LSD† | *** | ns | ns | ns | ns | *** | ns | * | ns | ns | |||
| Total Plant P Uptake (kg P ha−1) | AMF+ | 13.84 a | 12.83 a | 13.43 a | 11.43 b | 9.81 c | 12.27 A | 14.62 a | 13.49 a | 14.21 a | 12.03 b | 10.21 c | 12.91 A |
| AMF− | 11.80 a | 11.94 a | 12.15 a | 10.55 ab | 9.15 b | 11.12 B | 12.20 a | 12.43 a | 12.58 a | 10.76 ab | 9.34 b | 11.46 B | |
| LSD† | ** | * | * | ns | ns | ** | * | * | ns | * | |||
| PHI (%) | AMF+ | 69.67 a | 63.60 b | 70.77 ab | 68.77 ab | 70.03 a | 68.57 B | 69.50 a | 64.10 b | 70.86 a | 69.26 a | 69.15 a | 68.58 B |
| AMF− | 73.13 a | 71.47 a | 73.87 a | 72.27 a | 74.43 a | 73.03 A | 73.07 a | 71.46 a | 74.13 a | 71.97 a | 75.10 a | 73.15 A | |
| LSD† | ns | * | * | ns | ns | ns | * | * | ns | * | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beslemes, D.; Tigka, E.; Roussis, I.; Kakabouki, I.; Mavroeidis, A.; Vlachostergios, D. Effect of Arbuscular Mycorrhizal Fungi on Nitrogen and Phosphorus Uptake Efficiency and Crop Productivity of Two-Rowed Barley under Different Crop Production Systems. Plants 2023, 12, 1908. https://doi.org/10.3390/plants12091908
Beslemes D, Tigka E, Roussis I, Kakabouki I, Mavroeidis A, Vlachostergios D. Effect of Arbuscular Mycorrhizal Fungi on Nitrogen and Phosphorus Uptake Efficiency and Crop Productivity of Two-Rowed Barley under Different Crop Production Systems. Plants. 2023; 12(9):1908. https://doi.org/10.3390/plants12091908
Chicago/Turabian StyleBeslemes, Dimitrios, Evangelia Tigka, Ioannis Roussis, Ioanna Kakabouki, Antonios Mavroeidis, and Dimitrios Vlachostergios. 2023. "Effect of Arbuscular Mycorrhizal Fungi on Nitrogen and Phosphorus Uptake Efficiency and Crop Productivity of Two-Rowed Barley under Different Crop Production Systems" Plants 12, no. 9: 1908. https://doi.org/10.3390/plants12091908
APA StyleBeslemes, D., Tigka, E., Roussis, I., Kakabouki, I., Mavroeidis, A., & Vlachostergios, D. (2023). Effect of Arbuscular Mycorrhizal Fungi on Nitrogen and Phosphorus Uptake Efficiency and Crop Productivity of Two-Rowed Barley under Different Crop Production Systems. Plants, 12(9), 1908. https://doi.org/10.3390/plants12091908