
Farm or Lab? A Comparative Study of Oregano’s Leaf and Callus Volatile Isolates Chemistry and Cytotoxicity

 ,
,  , ,
, ,  ,
,  ,
, 
Abstract
1. Introduction
2. Results
2.1. Callus Biomass Propagation from Oregano’s Leaf Explants
2.1.1. Optimizing Conditions for Callus Induction
2.1.2. Callus Biomass Production from Whole Plant-Derived Leaf Explants
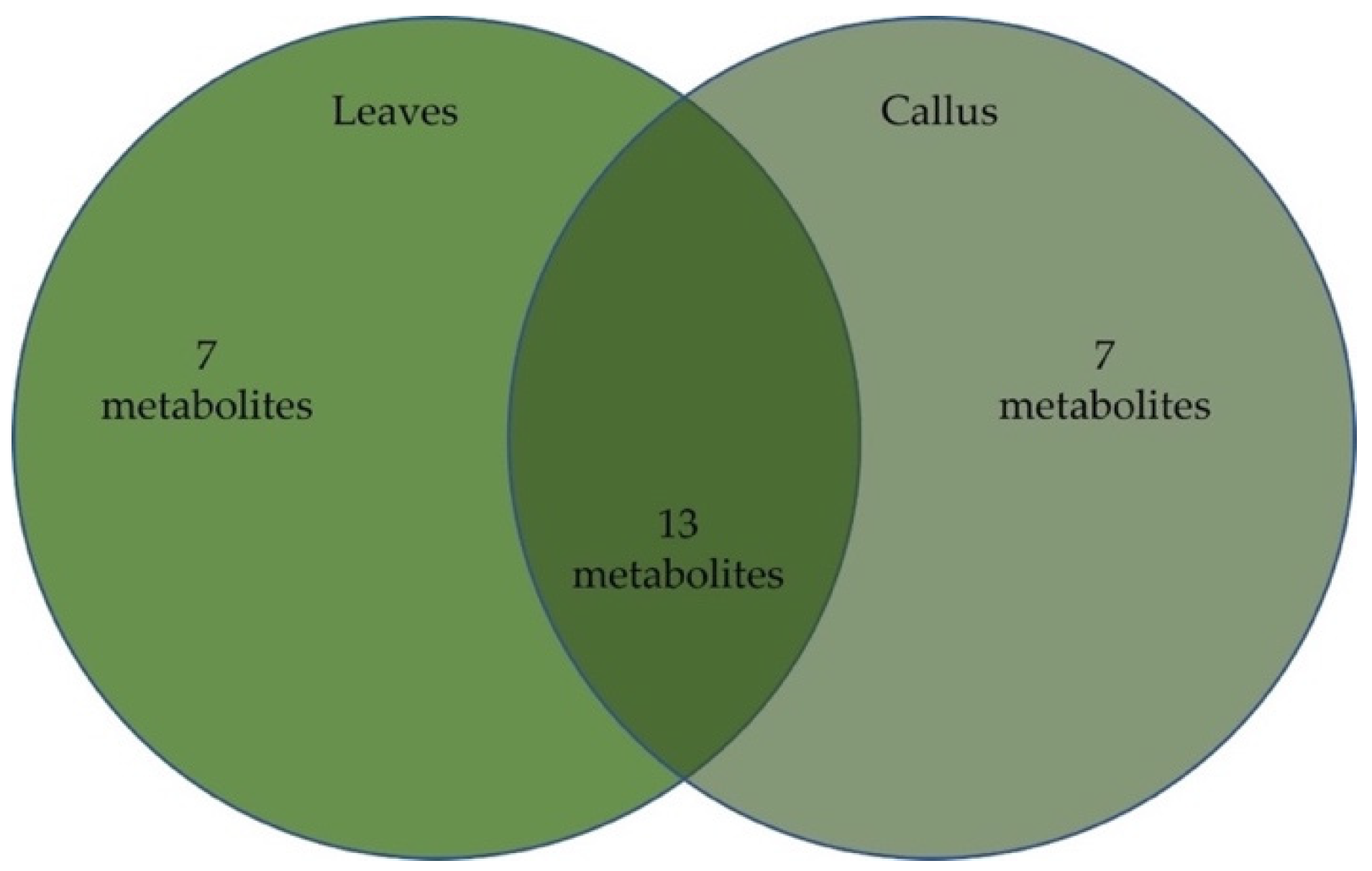
2.2. Chemical Composition of Oregano Leaves and Leaf-Derived Calluses
2.3. Determination of the Antioxidant Activity by the DPPH Radical Scavenging Assay
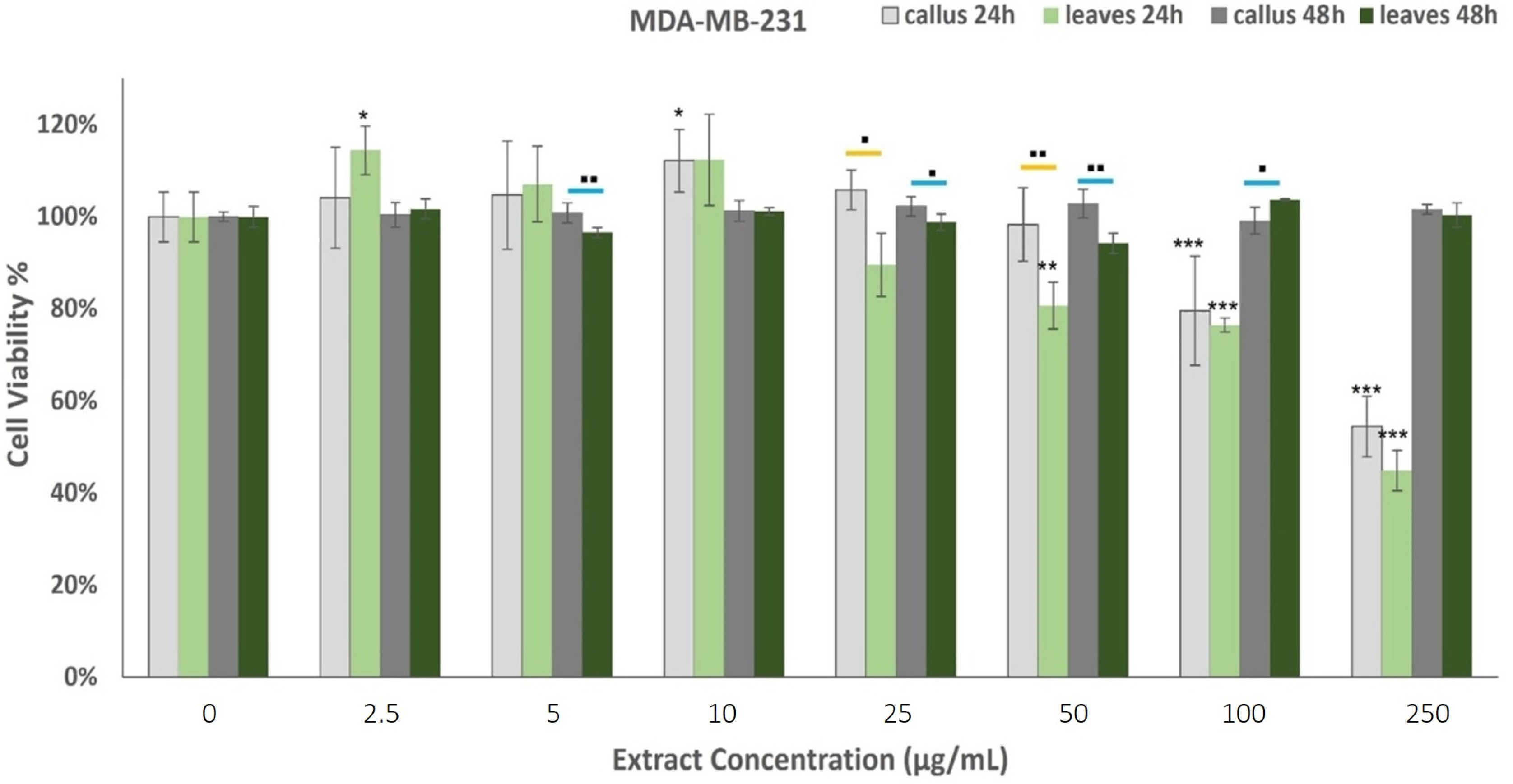
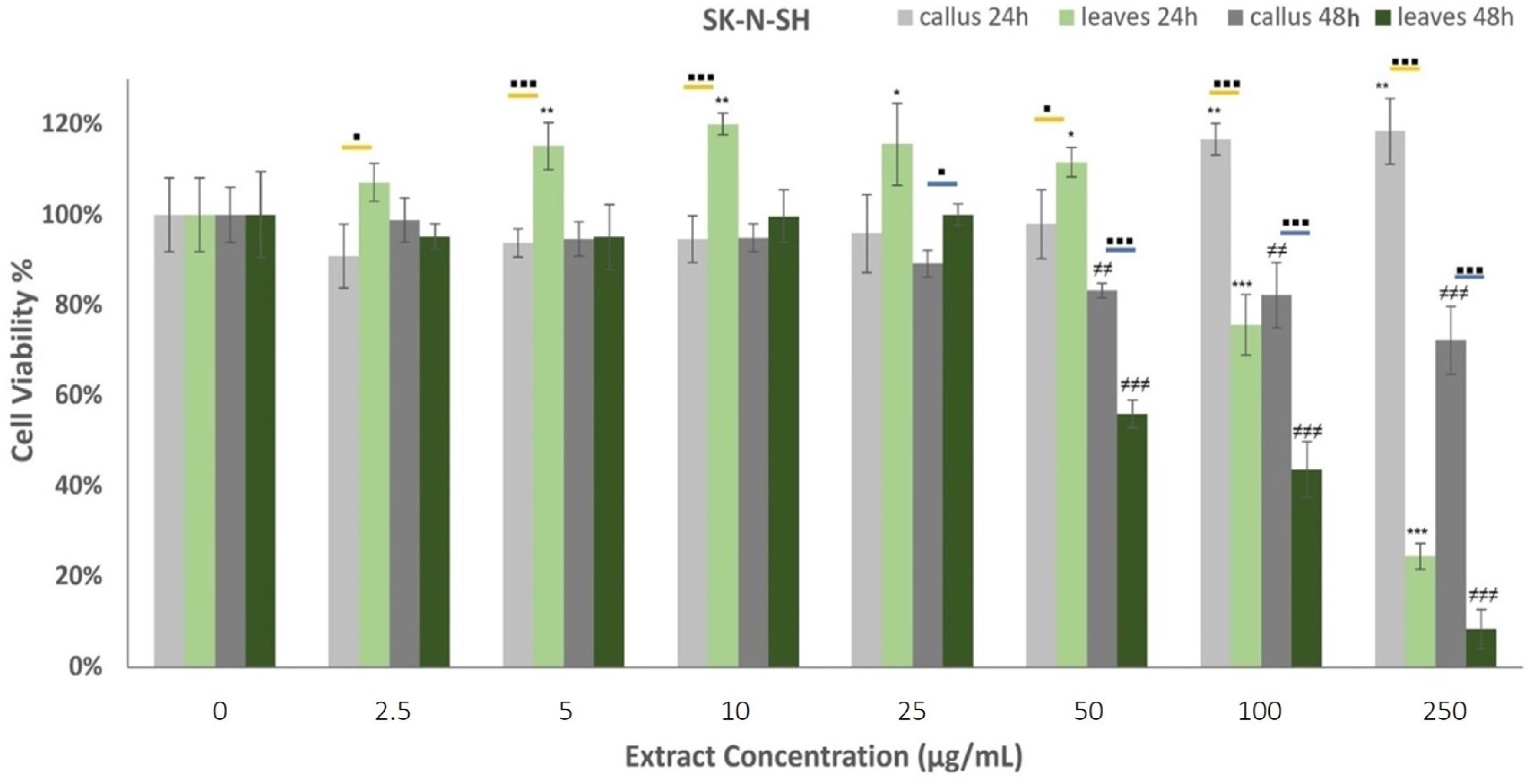
2.4. Anticancer Properties of Oregano Extracts
3. Discussion
4. Materials and Methods
4.1. In Vitro Callus Propagation
4.1.1. Plant Material and In Vitro Seedling Propagation
4.1.2. Callus Induction Optimization
4.1.3. Leaf-Derived Calluses from Commercial Plants
4.2. Herbal Tissue Extraction
4.3. Gas Chromatography–Mass Spectrometry (GC/MS)
4.4. Antioxidant Evaluation
4.5. Cytotoxicity Assays
4.5.1. Cell Culture
4.5.2. MTT Assay for Cell Viability
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, S.-L.; Yu, H.; Luo, H.-M.; Wu, Q.; Li, C.-F.; Steinmetz, A. Conservation and sustainable use of medicinal plants: Problems, progress, and prospects. Chin. Med. 2016, 11, 37. [Google Scholar] [CrossRef]
- Grigoriadou, K.; Krigas, N.; Sarropoulou, V.; Papanastasi, K.; Tsoktouridis, G.; Maloupa, E. In vitro propagation of medicinal and aromatic plants: The case of selected Greek species with conservation priority. Vitr. Cell. Dev. Biol.-Plant 2019, 55, 635–646. [Google Scholar] [CrossRef]
- Bhattacharjee, T.; Sen, S.; Chakraborty, R.; Maurya, P.K.; Chattopadhyay, A. Cultivation of Medicinal Plants: Special Reference to Important Medicinal Plants of India. In Herbal Medicine in India: Indigenous Knowledge, Practice, Innovation and Its Value; Sen, S., Chakraborty, R., Eds.; Springer Singapore: Singapore, 2020; pp. 101–115. [Google Scholar]
- Salamon, I. The Cultivation and Processing of Medicinal Plants in Slovakia. In Proceedings of the Second International Symposium on Agricultural Engineering, ISAE, Beograde, Serbia, 9–10 October 2015. [Google Scholar]
- Chandran, H.; Meena, M.; Barupal, T.; Sharma, K. Plant tissue culture as a perpetual source for production of industrially important bioactive compounds. Biotechnol. Rep. 2020, 26, e00450. [Google Scholar] [CrossRef]
- Bhatia, S.; Sharma, K.; Dahiya, R.; Bera, T. Modern Applications of Plant Biotechnology in Pharmaceutical Sciences; Academic Press: Cambridge, MA, USA, 2015; pp. 1–439. [Google Scholar]
- Thatoi, H.; Patra, J.K. Biotechnology and Pharmacological Evaluation of Medicinal Plants: An Overview. J. Herbs Spices Med. Plants 2011, 17, 214–248. [Google Scholar] [CrossRef]
- Ramani, S.; Jayabaskaran, C. Enhanced catharanthine and vindoline production in suspension cultures of Catharanthus roseus by ultraviolet-B light. J. Mol. Signal. 2008, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Misyri, V.; Tsekouras, V.; Iliopoulos, V.; Mavrikou, S.; Evergetis, E.; Moschopoulou, G.; Kintzios, S.; Haroutounian, S.A. Farm or lab? Chamazulene content of Artemisia arborescens (Vill.) L. essential oil and callus volatile metabolites isolate. Ind. Crops Prod. 2021, 160, 113114. [Google Scholar] [CrossRef]
- Deshpande, H.A.; Bhalsing, S.R. Isolation and characterization of diosgenin from in vitro cultured tissues of Helicteres isora L. Physiol. Mol. Biol. Plants 2014, 20, 89–94. [Google Scholar] [CrossRef]
- Qiang, B.; Miao, J.; Phillips, N.; Wei, K.; Gao, Y. Recent Advances in the Tissue Culture of American Ginseng (Panax quinquefolius). Chem. Biodivers. 2020, 17, e2000366. [Google Scholar] [CrossRef]
- Escrich, A.; Almagro, L.; Moyano, E.; Cusido, R.M.; Bonfill, M.; Hosseini, B.; Palazon, J. Improved biotechnological production of paclitaxel in Taxus media cell cultures by the combined action of coronatine and calix[8]arenes. Plant Physiol. Biochem. 2021, 163, 68–75. [Google Scholar] [CrossRef]
- Kintzios, S. Secondary Metabolite Production from Plant Cell Cultures: The Success Stories of Rosmarinic Acid and Taxol. In Bioactive Molecules and Medicinal Plants; Ramawat, K.G., Merillon, J.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 85–100. [Google Scholar]
- Bagheri, F.; Tahvilian, R.; Karimi, N.; Chalabi, M.; Azami, M. Shikonin Production by Callus Culture of Onosma bulbotrichom as Active Pharmaceutical Ingredient. Iran. J. Pharm. Res. 2018, 17, 495–504. [Google Scholar]
- Besher, S.; Al-Ammouri, Y.; Murshed, R. Production of tropan alkaloids in the in vitro and callus cultures of Hyoscyamus aureus and their genetic stability assessment using ISSR markers. Physiol. Mol. Biol. Plants 2014, 20, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Patel, V.; Modi, A.; Kumar, S.; Shukla, Y.M. Phyto-factories of anti-cancer compounds: A tissue culture perspective. Beni-Suef Univ. J. Basic Appl. Sci. 2022, 11, 43. [Google Scholar] [CrossRef]
- Niazian, M.; Sabbatini, P. Traditional in vitro strategies for sustainable production of bioactive compounds and manipulation of metabolomic profile in medicinal, aromatic and ornamental plants. Planta 2021, 254, 111. [Google Scholar] [CrossRef]
- Marchev, A.S.; Yordanova, Z.P.; Georgiev, M.I. Green (cell) factories for advanced production of plant secondary metabolites. Crit. Rev. Biotechnol. 2020, 40, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Lautié, E.; Russo, O.; Ducrot, P.; Boutin, J.A. Unraveling Plant Natural Chemical Diversity for Drug Discovery Purposes. Front. Pharmacol. 2020, 11, 397. [Google Scholar] [CrossRef]
- Joseph, R.C.; Silva da Fonseca Diniz, M.; Magno do Nascimento, V.; Barbosa Muribeca, A.d.J.; Costa Santiago, J.C.; da Cunha Borges, L.; da Costa Sá, P.R.; Portal Gomes, P.W.; da Silva Cardoso, J.C.; Rocha de Castro, M.N.; et al. Secure and Sustainable Sourcing of Plant Tissues for the Exhaustive Exploration of Their Chemodiversity. Molecules 2020, 25, 5992. [Google Scholar] [CrossRef]
- Köksal, Ö.; Gunes, E.; Özer, O.O.; Ozden, M. Analysis of effective factors on information sources at Turkish Oregano farms. Afr. J. Agric. Res. 2010, 5, 142–149. [Google Scholar]
- Morshedloo, M.R.; Salami, S.A.; Nazeri, V.; Maggi, F.; Craker, L. Essential oil profile of oregano (Origanum vulgare L.) populations grown under similar soil and climate conditions. Ind. Crops Prod. 2018, 119, 183–190. [Google Scholar] [CrossRef]
- Murillo-Amador, B.; Nieto-Garibay, A.; López-Aguilar, R.; Troyo-Diéguez, E.; Rueda-Puente, E.O.; Flores-Hernández, A.; Ruiz-Espinoza, F.H. Physiological, morphometric characteristics and yield of Origanum vulgare L. and Thymus vulgaris L. exposed to open-field and shade-enclosure. Ind. Crops Prod. 2013, 49, 659–667. [Google Scholar] [CrossRef]
- Arafeh, R.M.; Shibli, R.A.; Al-Mahmoud, M.; Shatnawi, M.A. Callusing, Cell Suspension Culture and Secondary Metabolites Production in Persian Oregano (Origanum vulgare L.) and Arabian Oregano (O. syriacum L.). Jordan J. Agric. Sci. 2006, 2, 274–282. [Google Scholar]
- Kumari, N.; Pardha-Saradhi, P. Regeneration of plants from callus cultures of Origanum vulgare L. Plant Cell Rep. 1992, 11, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.; Cardinali, A.; Ruta, C.; Fortunato, I.M.; Lattanzio, V.M.T.; Linsalata, V.; Cicco, N. Relationship of secondary metabolism to growth in oregano (Origanum vulgare L.) shoot cultures under nutritional stress. Environ. Exp. Bot. 2009, 65, 54–62. [Google Scholar] [CrossRef]
- Li, Y.-P.; Tang, D.-B.; Wang, X.-Q.; Wang, M.; Zhang, Q.-F.; Liu, Y.; Shen, B.-Y.; Chen, J.-G.; Yin, Z.-P. Development of Origanum vulgare Cell Suspension Culture to Produce Polyphenols and the Stimulation Effect of Salicylic Acid Elicitation and Phenylalanine Feeding. Biotechnol. Bioprocess Eng. 2021, 26, 456–467. [Google Scholar] [CrossRef]
- Veenstra, J.P.; Johnson, J.J. Oregano (Origanum vulgare) extract for food preservation and improvement in gastrointestinal health. Int. J. Nutr. 2019, 3, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Tang, H.; Yang, C.; Chen, J.; Wang, L.; Dong, Q.; Shi, W.; Li, L.; Liu, Y. Evaluation of the slow-release polylactic acid/polyhydroxyalkanoates active film containing oregano essential oil on the quality and flavor of chilled pufferfish (Takifugu obscurus) fillets. Food Chem. 2022, 385, 132693. [Google Scholar] [CrossRef]
- Mercati, F.; Dall’Aglio, C.; Acuti, G.; Faeti, V.; Tardella, F.M.; Pirino, C.; De Felice, E.; Scocco, P. Oregano Feed Supplementation Affects Glycoconjugates Production in Swine Gut. Animals 2020, 10, 149. [Google Scholar] [CrossRef]
- Alarcon-Rojo, A.; Janacua-Vidales, H.; Renteria, A. Oregano Essential Oil in Animal Production. InTech 2017, 13, 225–235. [Google Scholar]
- Oleynikov, V. Antioxidant and antimicrobial properties of oregano extract (Origani vulgaris herba L.). Foods Raw Mater. 2020, 8, 84–90. [Google Scholar] [CrossRef]
- Skoufogianni, E.; Solomou, A.D.; Danalatos, N.G. Ecology, Cultivation and Utilization of the Aromatic Greek Oregano (Origanum vulgare L.): A Review. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 545–552. [Google Scholar] [CrossRef]
- Gutiérrez-Grijalva, E.P.; Picos-Salas, M.A.; Leyva-López, N.; Criollo-Mendoza, M.S.; Vazquez-Olivo, G.; Heredia, J.B. Flavonoids and Phenolic Acids from Oregano: Occurrence, Biological Activity and Health Benefits. Plants 2017, 7, 2. [Google Scholar] [CrossRef]
- Jafari Khorsand, G.; Morshedloo, M.R.; Mumivand, H.; Emami Bistgani, Z.; Maggi, F.; Khademi, A. Natural diversity in phenolic components and antioxidant properties of oregano (Origanum vulgare L.) accessions, grown under the same conditions. Sci. Rep. 2022, 12, 5813. [Google Scholar] [CrossRef] [PubMed]
- de Torre, M.P.; Vizmanos, J.L.; Cavero, R.Y.; Calvo, M.I. Improvement of antioxidant activity of oregano (Origanum vulgare L.) with an oral pharmaceutical form. Biomed Pharm. 2020, 129, 110424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.L.; Guo, Y.S.; Wang, C.H.; Li, G.Q.; Xu, J.J.; Chung, H.Y.; Ye, W.C.; Li, Y.L.; Wang, G.C. Phenolic compounds from Origanum vulgare and their antioxidant and antiviral activities. Food Chem. 2014, 152, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Chuang, L.T.; Tsai, T.H.; Lien, T.J.; Huang, W.C.; Liu, J.J.; Chang, H.; Chang, M.L.; Tsai, P.J. Ethanolic Extract of Origanum vulgare Suppresses Propionibacterium acnes-Induced Inflammatory Responses in Human Monocyte and Mouse Ear Edema Models. Molecules 2018, 23, 1987. [Google Scholar] [CrossRef]
- Coccimiglio, J.; Alipour, M.; Jiang, Z.H.; Gottardo, C.; Suntres, Z. Antioxidant, Antibacterial, and Cytotoxic Activities of the Ethanolic Origanum vulgare Extract and Its Major Constituents. Oxid Med. Cell. Longev. 2016, 2016, 1404505. [Google Scholar] [CrossRef]
- Oaldje Pavlovic, M.; Lunić, T.; Mandic, M.; Alimpić Aradski, A.; Duletic, S.; Nedeljković, B. The Marvellous Oregano Spices; Multidisciplinary Digital Publishing Institute: Basel, Switzerland, 2021. [Google Scholar]
- Khaleel, C.; Tabanca, N.; Buchbauer, G. α-Terpineol, a natural monoterpene: A review of its biological properties. Open Chem. 2018, 16, 349–361. [Google Scholar] [CrossRef]
- Balahbib, A.; El Omari, N.; Hachlafi, N.E.L.; Lakhdar, F.; El Menyiy, N.; Salhi, N.; Mrabti, H.N.; Bakrim, S.; Zengin, G.; Bouyahya, A. Health beneficial and pharmacological properties of p-cymene. Food Chem. Toxicol. 2021, 153, 112259. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, L.A.; Pina, L.T.S.; Serafini, M.R.; Tavares, D.d.S.; Guimarães, A.G. Antitumor Effects of Carvacrol and Thymol: A Systematic Review. Front. Pharmacol. 2021, 12, 702487. [Google Scholar] [CrossRef]
- Han, X.; Parker, T.L. Anti-inflammatory, tissue remodeling, immunomodulatory, and anticancer activities of oregano (Origanum vulgare) essential oil in a human skin disease model. Biochim. Open 2017, 4, 73–77. [Google Scholar] [CrossRef]
- de Cássia da Silveira, E.S.R.; Lima, T.C.; da Nóbrega, F.R.; de Brito, A.E.M.; de Sousa, D.P. Analgesic-Like Activity of Essential Oil Constituents: An Update. Int. J. Mol. Sci. 2017, 18, 2392. [Google Scholar] [CrossRef]
- Global Burden of Disease 2019 Cancer Collaboration. Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life Years for 29 Cancer Groups From 2010 to 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. JAMA Oncol. 2022, 8, 420–444. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA A Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, I.D.L.; Soares, J.C.S. Conventional Cancer Treatment. In Advances in Cancer Treatment: From Systemic Chemotherapy to Targeted Therapy; Cavalcanti, I.D.L., Soares, J.C.S., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 29–56. [Google Scholar]
- Falzone, L.; Salomone, S.; Libra, M. Evolution of Cancer Pharmacological Treatments at the Turn of the Third Millennium. Front. Pharmacol. 2018, 9, 1300. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, A.J.; Jahan, S.; Singh, R.; Saxena, J.; Ashraf, S.A.; Khan, A.; Choudhary, R.K.; Balakrishnan, S.; Badraoui, R.; Bardakci, F.; et al. Plants in Anticancer Drug Discovery: From Molecular Mechanism to Chemoprevention. BioMed Res. Int. 2022, 2022, 5425485. [Google Scholar] [CrossRef]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in Cancer Treatment: From Preclinical Studies to Clinical Practice. Front. Pharmacol. 2020, 10, 1614. [Google Scholar] [CrossRef]
- Khan, A.W.; Farooq, M.; Haseeb, M.; Choi, S. Role of Plant-Derived Active Constituents in Cancer Treatment and Their Mechanisms of Action. Cells 2022, 11, 1326. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, A.; Ramachandran, S.; Gupta, N.; Kaushik, I.; Wright, S.; Srivastava, S.; Das, H.; Srivastava, S.; Prasad, S.; Srivastava, S.K. Role of Phytochemicals in Cancer Prevention. Int. J. Mol. Sci. 2019, 20, 4981. [Google Scholar] [CrossRef] [PubMed]
- Sagharyan, M.; Ganjeali, A.; Cheniany, M.; Mousavi Kouhi, S.M. Optimization of Callus Induction with Enhancing Production of Phenolic Compounds Production and Antioxidants Activity in Callus Cultures of Nepeta binaloudensis Jamzad (Lamiaceae). Iran. J. Biotechnol. 2020, 18, 47–55. [Google Scholar] [CrossRef]
- Suran, D.; Tumendalai, B.; Bayarmaa, G.-A. In vitro Seed Germination and Callus Induction of Ferula ferulaeoides (Steud.) Korov. (Apiaceae). Mong. J. Biol. Sci. 2016, 14, 53–58. [Google Scholar] [CrossRef]
- Braga, K.; Coimbra, M.; Castro, A. In vitro germination, callus induction and phenolic compounds contents from Pyrostegia venusta (Ker gawl.) miers. Acta Sci.-Biol. Sci. 2015, 37, 151–158. [Google Scholar] [CrossRef]
- Darwesh, H.; Nour Eldeen, A.; Fayad, E. In-vitro investigation for improving secondary metabolites in Origanum vulgare plants using tissue culture technique at Taif Governorate, KSA. Res. J. Pharm. Biol. Chem. Sci. 2015, 6, 1117–1122. [Google Scholar]
- Wang, Y.; Wang, Y.; Li, K.; Song, X.; Chen, J. Characterization and Comparative Expression Profiling of Browning Response in Medinilla formosana after Cutting. Front. Plant Sci. 2016, 7, 1897. [Google Scholar] [CrossRef] [PubMed]
- Yari Khosroushahi, A.; Naderi-Manesh, H.; Toft Simonsen, H. Effect of Antioxidants and Carbohydrates in Callus Cultures of Taxus brevifolia: Evaluation of Browning, Callus Growth, Total Phenolics and Paclitaxel Production. Bioimpacts 2011, 1, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Siddique, A.B.; Islam, S.M.S. Effect of light and dark on callus induction and regeneration in tobacco (Nicotiana tabacum L.). Bangladesh J. Bot. 2018, 44, 643–651. [Google Scholar] [CrossRef]
- Kintzios, S.; Papanastasiou, I.; Tourgelis, P.; Papastellatos, C.; Georgopoulos, V.; Drossopoulos, J. The Effects of Light on Callus Growth and Somatic Embryogenesis from Lavandula vera and Teucrium chamaedrys: A Preliminary Study. J. Herbs Spices Med. Plants 2002, 9, 223–227. [Google Scholar] [CrossRef]
- Jones, A.M.P.; Saxena, P.K. Inhibition of Phenylpropanoid Biosynthesis in Artemisia annua L.: A Novel Approach to Reduce Oxidative Browning in Plant Tissue Culture. PLoS ONE 2013, 8, e76802. [Google Scholar] [CrossRef]
- Pence, V.C. Evaluating costs for the in vitro propagation and preservation of endangered plants. Vitr. Cell. Dev. Biol.-Plant 2011, 47, 176–187. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Iwase, A.; Rymen, B.; Lambolez, A.; Kojima, M.; Takebayashi, Y.; Heyman, J.; Watanabe, S.; Seo, M.; De Veylder, L.; et al. Wounding Triggers Callus Formation via Dynamic Hormonal and Transcriptional Changes. Plant Physiol. 2017, 175, 1158–1174. [Google Scholar] [CrossRef]
- Efferth, T. Biotechnology Applications of Plant Callus Cultures. Engineering 2019, 5, 50–59. [Google Scholar] [CrossRef]
- Hosseini, B.; Bighamat, A. Effects of different concentrations of growth regulators and explants type on callus induction, embryogenesis and shoot regeneration of Origanum vulgare ssp. Gracile. Iran. J. Rangel. For. Plant Breed. Genet. Res. 2016, 24, 264–276. [Google Scholar] [CrossRef]
- Sökmen, M.; Serkedjieva, J.; Daferera, D.; Gulluce, M.; Polissiou, M.; Tepe, B.; Akpulat, H.A.; Sahin, F.; Sokmen, A. In vitro antioxidant, antimicrobial, and antiviral activities of the essential oil and various extracts from herbal parts and callus cultures of Origanum acutidens. J. Agric. Food Chem. 2004, 52, 3309–3312. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant Callus: Mechanisms of Induction and Repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef] [PubMed]
- Elgengaihi, S.; Taha, H.; Kamel, A.M. In vivo and in vitro comparative studies of Origanum species. J. Food Agric. Environ. 2006, 4, 127–134. [Google Scholar]
- Devasigamani, L.; Kuppan, N. Callus Induction and Regeneration of Multiple Shoots from In Vitro Apical Bud Explant of Origanum vulgare. An Important Medicinal Plant. Int. J. Res. Pharm. Chem. 2013, 3, 898–903. [Google Scholar]
- Adil, M.; Haider Abbasi, B.; ul Haq, I. Red light controlled callus morphogenetic patterns and secondary metabolites production in Withania somnifera L. Biotechnol. Rep. 2019, 24, e00380. [Google Scholar] [CrossRef]
- Moon, H.K.; Stomp, A.M. Effects of medium components and light on callus induction, growth, and frond regeneration in Lemna gibba (Duckweed). Vitr. Cell. Dev. Biol.-Plant 1997, 33, 20–25. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Khan, T.; Khurshid, R.; Nadeem, M.; Drouet, S.; Hano, C. UV-C mediated accumulation of pharmacologically significant phytochemicals under light regimes in in vitro culture of Fagonia indica (L.). Sci. Rep. 2021, 11, 679. [Google Scholar] [CrossRef] [PubMed]
- Kintzios, S.; Barberaki, M.; Drossopoulos, J.; Turgelis, P.; Konstas, J. Effect of Medium Composition and Explant Source on the Distribution Profiles Selected Micronutrients in Mistletoe Tissue Cultures. J. Plant Nutr. 2003, 26, 369–397. [Google Scholar] [CrossRef]
- Alhasnawi, A.N.; Zain, C.R.C.M.; Kadhimi, A.A.; Isahak, A.; Mohamad, A.; Ashraf, M.F.; Doni, F.; Yusoff, W.M.W. Impact of exogenous ascorbic acid on biochemical activities of rice callus treated with salt stress. AIP Conf. Proc. 2016, 1784, 020005. [Google Scholar] [CrossRef]
- Hönig, M.; Plíhalová, L.; Husičková, A.; Nisler, J.; Doležal, K. Role of Cytokinins in Senescence, Antioxidant Defence and Photosynthesis. Int. J. Mol. Sci. 2018, 19, 4045. [Google Scholar] [CrossRef]
- Pazurkiewicz-Kocot, K.; Kita, A.; Haduch, A. The Effect of Kinetin on the Chlorophyll Pigments Content in Leaves of Zea mays L. In Seedlings and Accumulation of Some Metal Ions; University of Silesia: Katowice, Poland, 2011. [Google Scholar]
- Du, J.; Ma, W.; Li, Y.; Lu, X.; Geng, Z.; Huang, H.; Yuan, Y.; Liu, Y.; Wang, X.; Wang, J. UPLC–MS-Based Non-targeted Analysis of Endogenous Metabolite Changes in the Leaves of Scabiosa tschiliensis Grüning Induced by 6-Benzylaminopurine and Kinetin. Front. Plant Sci. 2021, 12, 700623. [Google Scholar] [CrossRef] [PubMed]
- Acidri, R.; Sawai, Y.; Sugimoto, Y.; Handa, T.; Sasagawa, D.; Masunaga, T.; Yamamoto, S.; Nishihara, E. Exogenous Kinetin Promotes the Nonenzymatic Antioxidant System and Photosynthetic Activity of Coffee (Coffea arabica L.) Plants Under Cold Stress Conditions. Plants 2020, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Alamri, S.A.; Alam, P.; Ashraf, M.; Ahmad, P. Potential of exogenously sourced kinetin in protecting Solanum lycopersicum from NaCl-induced oxidative stress through up-regulation of the antioxidant system, ascorbate-glutathione cycle and glyoxalase system. PLoS ONE 2018, 13, e0202175. [Google Scholar] [CrossRef] [PubMed]
- Dhindsa, R.S.; Plumb-Dhindsa, P.L.; Reid, D.M. Leaf senescence and lipid peroxidation: Effects of some phytohormones, and scavengers of free radicals and singlet oxygen. Physiol. Plant. 1982, 56, 453–457. [Google Scholar] [CrossRef]
- de Moura, F.B.; Vieira, M.R.d.S.; SimÕEs, A.d.N.; Ferreira-Silva, S.L.; de Souza, C.A.V.; de Souza, E.S.; da Rocha, A.T.; da Silva, L.F.; JÚNior, M.A. Physiological Effect of Kinetin on the Photosynthetic Apparatus and Antioxidant Enzymes Activities During Production of Anthurium. Hortic. Plant J. 2018, 4, 182–192. [Google Scholar] [CrossRef]
- Tan, X.; Calderon-Villalobos, L.I.; Sharon, M.; Zheng, C.; Robinson, C.V.; Estelle, M.; Zheng, N. Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature 2007, 446, 640–645. [Google Scholar] [CrossRef]
- Ma, Q.; Ding, Y.; Chang, J.; Sun, X.; Zhang, L.; Wei, Q.; Cheng, Y.; Chen, L.; Xu, J.; Deng, X. Comprehensive insights on how 2,4-dichlorophenoxyacetic acid retards senescence in post-harvest citrus fruits using transcriptomic and proteomic approaches. J. Exp. Bot. 2013, 65, 61–74. [Google Scholar] [CrossRef]
- Campanoni, P.; Nick, P. Auxin-Dependent Cell Division and Cell Elongation. 1-Naphthaleneacetic Acid and 2,4-Dichlorophenoxyacetic Acid Activate Different Pathways. Plant Physiol. 2005, 137, 939–948. [Google Scholar] [CrossRef]
- Ali, A.; Mohammad, S.; Khan, M.A.; Raja, N.I.; Arif, M.; Kamil, A.; Mashwani, Z.-u.-R. Silver nanoparticles elicited in vitro callus cultures for accumulation of biomass and secondary metabolites in Caralluma tuberculata. Artif. Cells Nanomed. Biotechnol. 2019, 47, 715–724. [Google Scholar] [CrossRef]
- Ramabulana, A.-T.; Steenkamp, P.A.; Madala, N.E.; Dubery, I.A. Application of Plant Growth Regulators Modulates the Profile of Chlorogenic Acids in Cultured Bidens pilosa Cells. Plants 2021, 10, 437. [Google Scholar] [CrossRef]
- Ren, X.; Liu, Y.; Jeong, B.R. Callus induction and browning suppression in tree peony Paeonia ostii ‘Fengdan’. Hortic. Environ. Biotechnol. 2020, 61, 591–600. [Google Scholar] [CrossRef]
- Holderbaum, D.F.; Kon, T.; Kudo, T.; Guerra, M.P. Enzymatic Browning, Polyphenol Oxidase Activity, and Polyphenols in Four Apple Cultivars: Dynamics during Fruit Development. HortScience 2010, 45, 1150–1154. [Google Scholar] [CrossRef]
- Economou, G.; Panagopoulos, G.; Karamanos, A.; Tarantilis, P.; Kalivas, D.; Kotoulas, V. An assessment of the behavior of carvacrol–rich wild Lamiaceae species from the eastern Aegean under cultivation in two different environments. Ind. Crops Prod. 2014, 54, 62–69. [Google Scholar] [CrossRef]
- Evergetis, E.; Bellini, R.; Balatsos, G.; Michaelakis, A.; Carrieri, M.; Veronesi, R.; Papachristos, D.P.; Puggioli, A.; Kapsaski-Kanelli, V.-N.; Haroutounian, S.A. From Bio-Prospecting to Field Assessment: The Case of Carvacrol Rich Essential Oil as a Potent Mosquito Larvicidal and Repellent Agent. Front. Ecol. Evol. 2018, 6, 204. [Google Scholar] [CrossRef]
- Evergetis, E.; Haroutounian, S.A. Volatile systematics: A novel biochemical interpretation of essential oil compounds enhances their chemophenetic significance. Biochem. Syst. Ecol. 2020, 92, 104087. [Google Scholar] [CrossRef]
- Crozier, A.; Moritz, T. Chapter 2-Physico-chemical methods of plant hormone analysis. In New Comprehensive Biochemistry; Hooykaas, P.J.J., Hall, M.A., Libbenga, K.R., Eds.; Elsevier: Amsterdam, The Netherlands, 1999; Volume 33, pp. 23–60. [Google Scholar]
- Fan, K.; Li, X.; Cao, Y.; Qi, H.; Li, L.; Zhang, Q.; Sun, H. Carvacrol inhibits proliferation and induces apoptosis in human colon cancer cells. Anti-Cancer Drugs 2015, 26, 813–823. [Google Scholar] [CrossRef]
- Heidarian, E.; Keloushadi, M. Antiproliferative and Anti-invasion Effects of Carvacrol on PC3 Human Prostate Cancer Cells through Reducing pSTAT3, pAKT, and pERK1/2 Signaling Proteins. Int. J. Prev. Med. 2019, 10, 156. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, S.J.; Ahmad, A.; Irshad, M.; Rizvi, M. Cytotoxic effect of Carvacrol on human cervical cancer cells. Biol. Med. 2011, 2011, 307–312. [Google Scholar] [CrossRef]
- Al-Kalaldeh, J.Z.; Abu-Dahab, R.; Afifi, F.U. Volatile oil composition and antiproliferative activity of Laurus nobilis, Origanum syriacum, Origanum vulgare, and Salvia triloba against human breast adenocarcinoma cells. Nutr. Res. 2010, 30, 271–278. [Google Scholar] [CrossRef]
- Hu, H.; Zheng, X.; Hu, H. Chemical composition, antimicrobial, antioxidant and cytotoxic activity of the essential oil from the leaves of Acanthopanax leucorrhizus (Oliv.) Harms. Env. Toxicol. Pharm. 2012, 34, 618–623. [Google Scholar] [CrossRef]
- Horton, A.W.; Eshleman, D.N.; Schuff, A.R.; Perman, W.H. Correlation of cocarcinogenic activity among n-alkanes with their physical effects on phospholipid micelles. J. Natl. Cancer Inst. 1976, 56, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, C.; Matsuo, A.; Valenca Pereira, F.; Rabaça, A.; Farias, C.; Girola, N.; Massaoka, M.H.; Azevedo, R.A.; Scutti, J.; Arruda, D.; et al. Pyrostegia venusta heptane extract containing saturated aliphatic hydrocarbons induces apoptosis on B16F10-Nex2 melanoma cells and displays antitumor activity in vivo. Pharmacogn. Mag. 2014, 10, S363–S376. [Google Scholar] [CrossRef]
- Theodossiou, T.A.; Ali, M.; Grigalavicius, M.; Grallert, B.; Dillard, P.; Schink, K.O.; Olsen, C.E.; Wälchli, S.; Inderberg, E.M.; Kubin, A.; et al. Simultaneous defeat of MCF7 and MDA-MB-231 resistances by a hypericin PDT–tamoxifen hybrid therapy. NPJ Breast Cancer 2019, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Adams, R. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy; Allured Publ. Corp.: Carol Stream IL, USA, 1995; Volume 16. [Google Scholar]
- Masada, Y. Analysis of Essential Oils by Gas Chromatography and Mass Spectrometry; Halsted Press: New York, NY, USA, 1976; Volume 14, p. 17A. [Google Scholar]
- Nickavar, B.; Kamalinejad, M.; Haj-Yahya, M.; Shafaghi, B. Comparison of the Free Radical Scavenging Activity of Six Iranian Achillea. Species. Pharm. Biol. 2006, 44, 208–212. [Google Scholar] [CrossRef]
- Paivana, G.; Mavrikou, S.; Kaltsas, G.; Kintzios, S. Bioelectrical Analysis of Various Cancer Cell Types Immobilized in 3D Matrix and Cultured in 3D-Printed Well. Biosensors 2019, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Mavrikou, S.; Tsekouras, V.; Karageorgou, M.-A.; Moschopoulou, G.; Kintzios, S. Anticancer and biochemical effects of Viscum album L. protein extracts on HeLa cells. Plant Cell Tissue Organ Cult. 2020, 140, 369–378. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infected Explants | Callus Induction | Brown Callus | Healthy Callus |
|---|---|---|---|
| 8.49% | 56.70% | 10.91% | 46.23% |
| Components | RI | Leaves | Callus | Identification |
|---|---|---|---|---|
| β-Thujene | 910 | 0.2 | nd | a, b |
| α-phellandrene | 915 | nd | 0.2 | a, b |
| Sabinene | 962 | 0.4 | nd | a, b, c |
| β-Pinene | 964 | nd | 0.3 | a, b |
| β-Myrcene | 986 | 0.6 | 0.7 | a, b |
| α-Terpinene | 1012 | 0.4 | 0.2 | a, b |
| o-Cymene | 1020 | 1.2 | 0.7 | a, b |
| β-Phellandrene | 1024 | 0.5 | nd | a, b, c |
| γ-Terpinene | 1055 | 1.9 | 0.4 | a, b |
| cis-Sabinenehydrate | 1062 | 1.7 | 0.8 | a, b |
| trans-Sabinenehydrate | 1094 | 13.9 | 7.5 | a, b |
| Linalool | 1100 | 3.0 | nd | a, b, c |
| Borneol | 1161 | 0.3 | nd | a, b |
| Terpinen-4-ol | 1174 | 0.3 | 1.8 | a, b, c |
| Terpineol | 1187 | 1.9 | 1.3 | a, b, c |
| Linalyl acetate | 1256 | 4.4 | nd | a, b |
| Thymol | 1295 | 0.4 | 0.1 | a, b, c |
| Carvacrol | 1304 | 55.4 | 39.2 | a, b, c |
| Carvvacrol-1-TMS | 1334 | nd | 9.4 | a, b |
| Caryophyllene | 1415 | 1.3 | 1.5 | a, b, c |
| Bicyclogermacrene | 1495 | 0.7 | nd | a, b |
| 1-Hexadecanol-1-TMS | 1964 | nd | 0.9 | a, b |
| Hexadodecanoic acid ethylester | 1992 | nd | 0.7 | a, b |
| Hexacosane | 3088 | nd | 2.1 | a, b, c |
| Octacosane | 3279 | 1.6 | 6.5 | a, b, c |
| Nonacosane | 3485 | nd | 0.7 | a, b, c |
| Triacontane | 3543 | 2.1 | 6.2 | a, b, c |
| Total | 92.0 | 81.2 |
| Treatment Code | 2,4 D (mg/L) | Cytokine (mg/L) | Ascorbic Acid (mg/L) | Light (Hours) | |
|---|---|---|---|---|---|
| BAP | Kinetin | ||||
| BL | 0.5 | 3 | - | - | 16 |
| BD | 0.5 | 3 | - | - | - |
| BAL | 0.5 | 3 | - | 10 | 16 |
| BAD | 0.5 | 3 | - | 10 | - |
| KL | 2 | - | 2 | - | 16 |
| KD | 2 | - | 2 | - | - |
| KAL | 2 | - | 2 | 10 | 16 |
| KAD | 2 | - | 2 | 10 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kakalis, A.; Tsekouras, V.; Mavrikou, S.; Moschopoulou, G.; Kintzios, S.; Evergetis, E.; Iliopoulos, V.; Koulocheri, S.D.; Haroutounian, S.A. Farm or Lab? A Comparative Study of Oregano’s Leaf and Callus Volatile Isolates Chemistry and Cytotoxicity. Plants 2023, 12, 1472. https://doi.org/10.3390/plants12071472
Kakalis A, Tsekouras V, Mavrikou S, Moschopoulou G, Kintzios S, Evergetis E, Iliopoulos V, Koulocheri SD, Haroutounian SA. Farm or Lab? A Comparative Study of Oregano’s Leaf and Callus Volatile Isolates Chemistry and Cytotoxicity. Plants. 2023; 12(7):1472. https://doi.org/10.3390/plants12071472
Chicago/Turabian StyleKakalis, Antonis, Vasileios Tsekouras, Sofia Mavrikou, Georgia Moschopoulou, Spyridon Kintzios, Epameinondas Evergetis, Vasilios Iliopoulos, Sofia D. Koulocheri, and Serkos A. Haroutounian. 2023. "Farm or Lab? A Comparative Study of Oregano’s Leaf and Callus Volatile Isolates Chemistry and Cytotoxicity" Plants 12, no. 7: 1472. https://doi.org/10.3390/plants12071472
APA StyleKakalis, A., Tsekouras, V., Mavrikou, S., Moschopoulou, G., Kintzios, S., Evergetis, E., Iliopoulos, V., Koulocheri, S. D., & Haroutounian, S. A. (2023). Farm or Lab? A Comparative Study of Oregano’s Leaf and Callus Volatile Isolates Chemistry and Cytotoxicity. Plants, 12(7), 1472. https://doi.org/10.3390/plants12071472