
Cloning of the Soybean GmNHL1 Gene and Functional Analysis under Salt Stress


Abstract
:1. Introduction
2. Results
2.1. Bioinformatics Analysis of GmNHL1 Gene
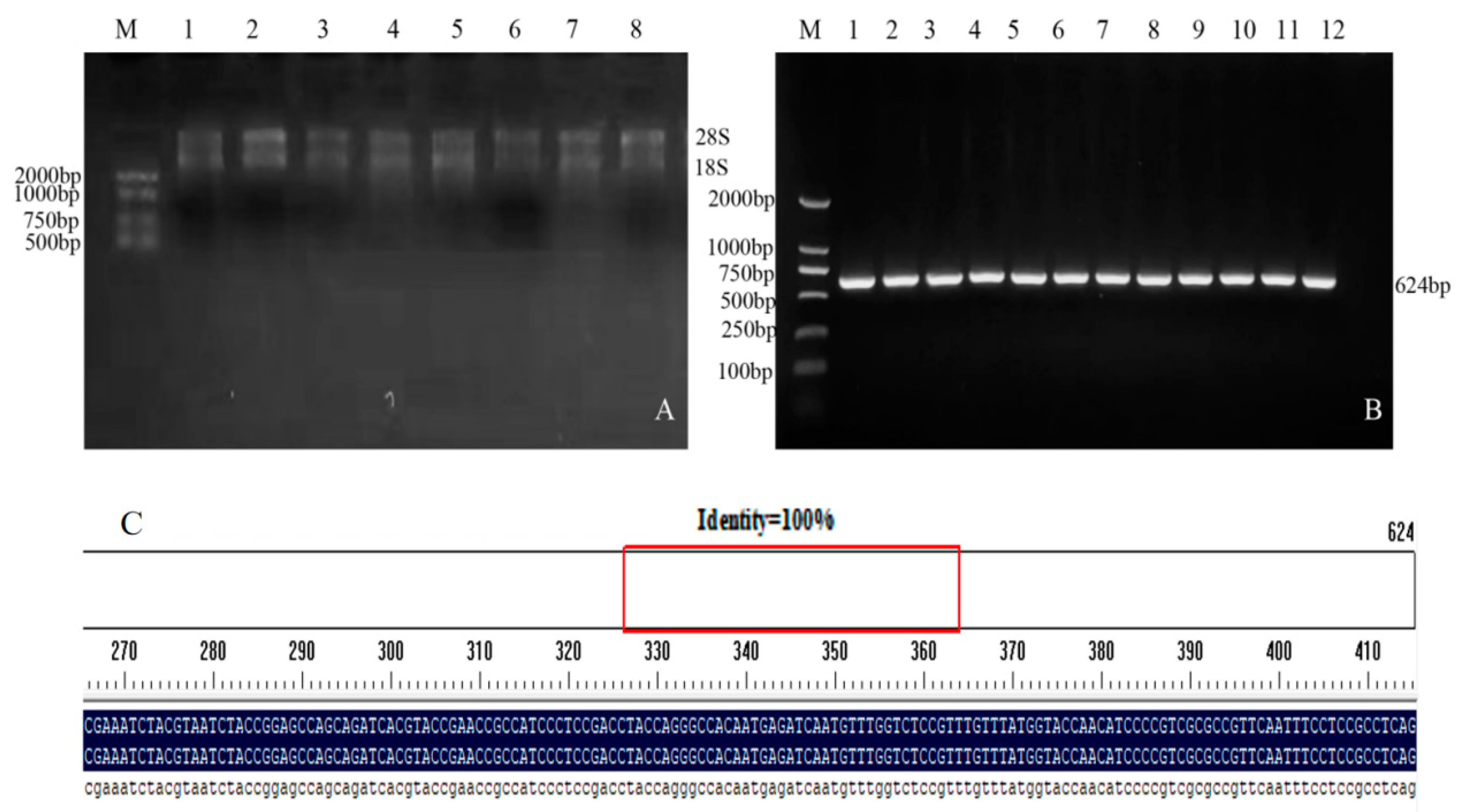
2.2. Cloning of Target Genes
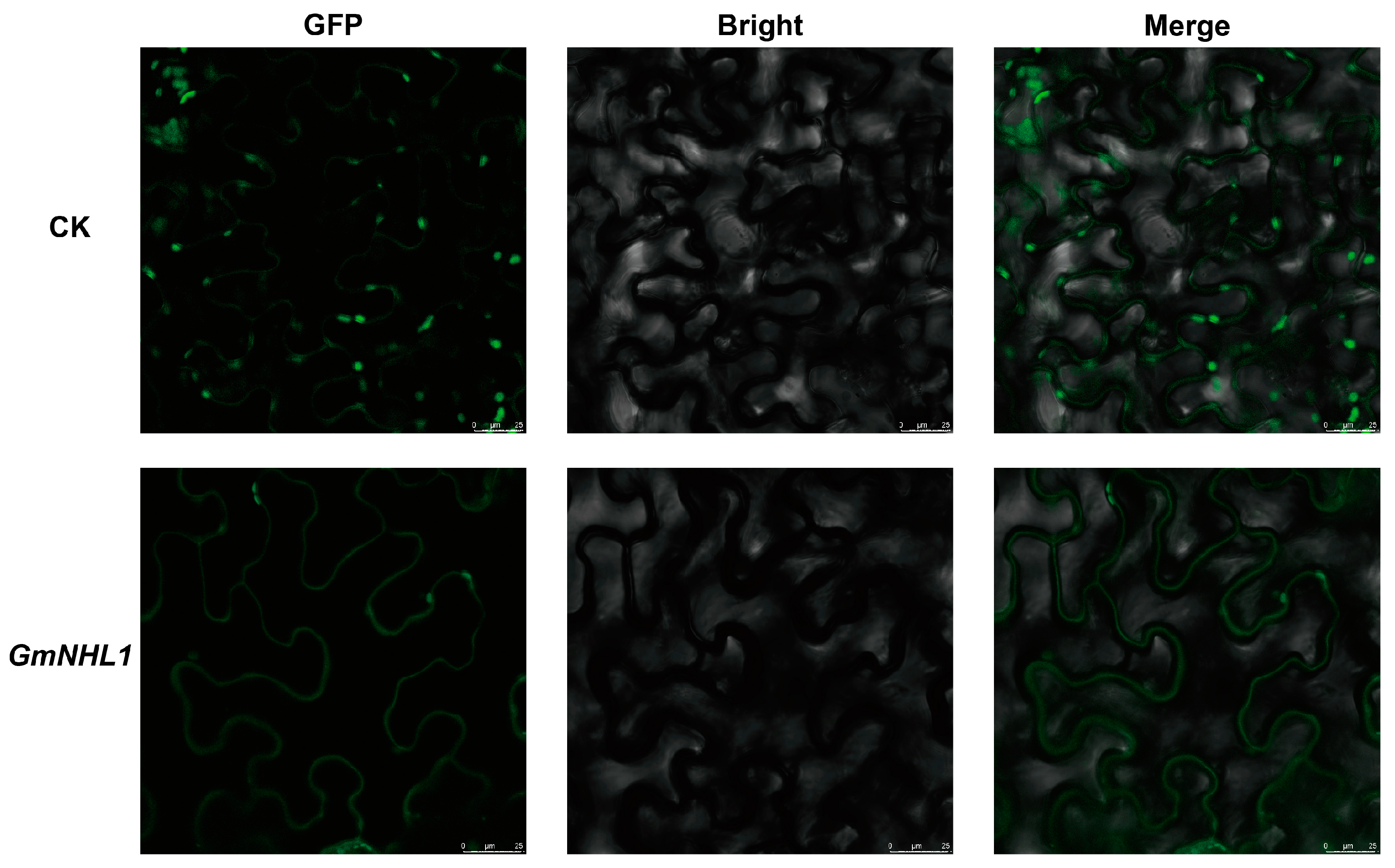
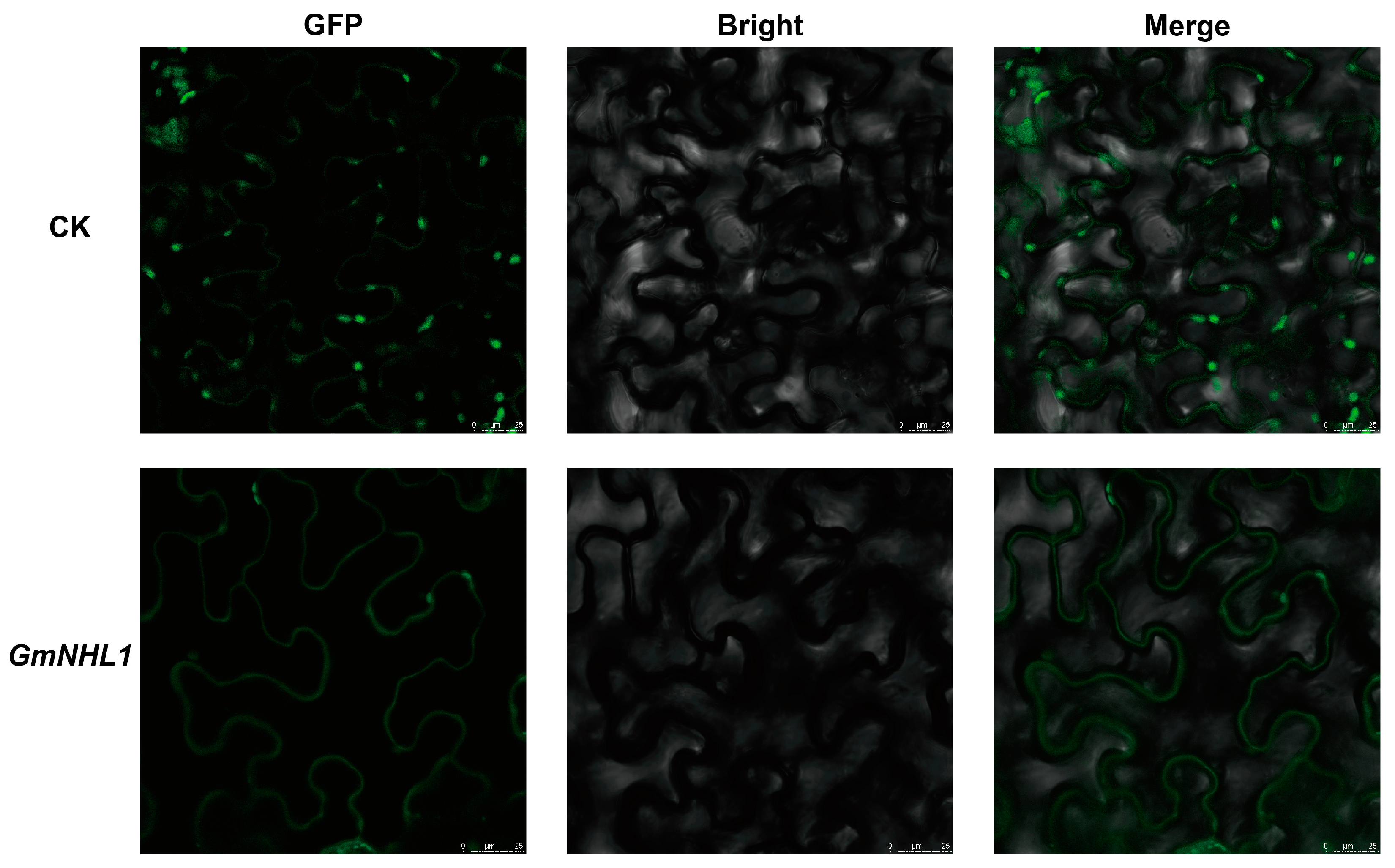
2.3. Sub-Cellular Localization
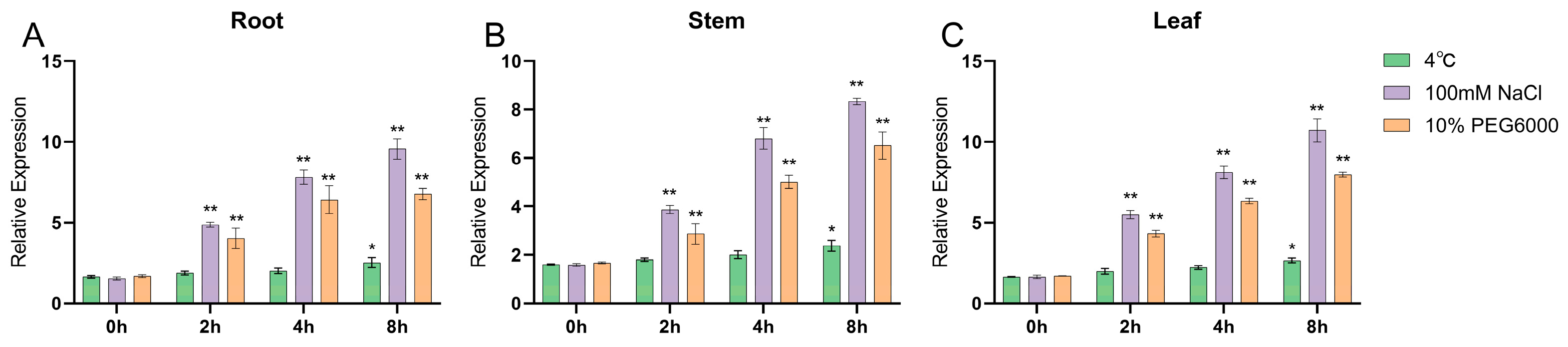
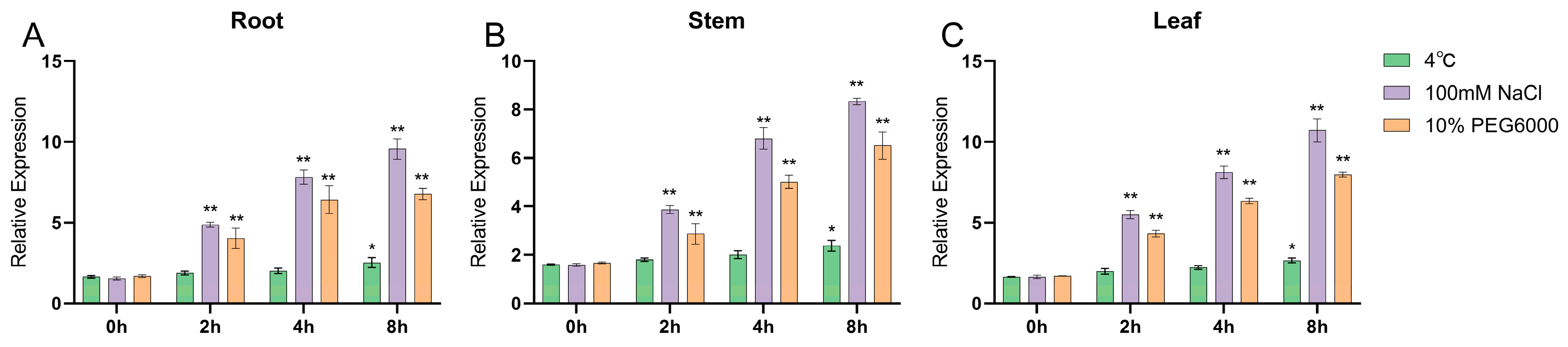
2.4. Analysis of Gene Expression Patterns under Different Stress Conditions
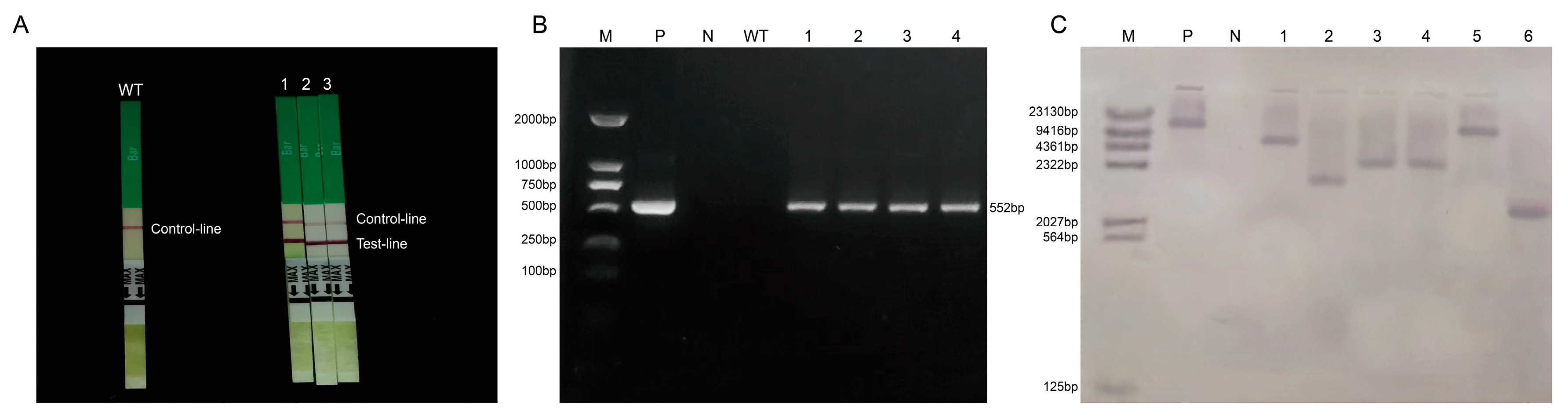
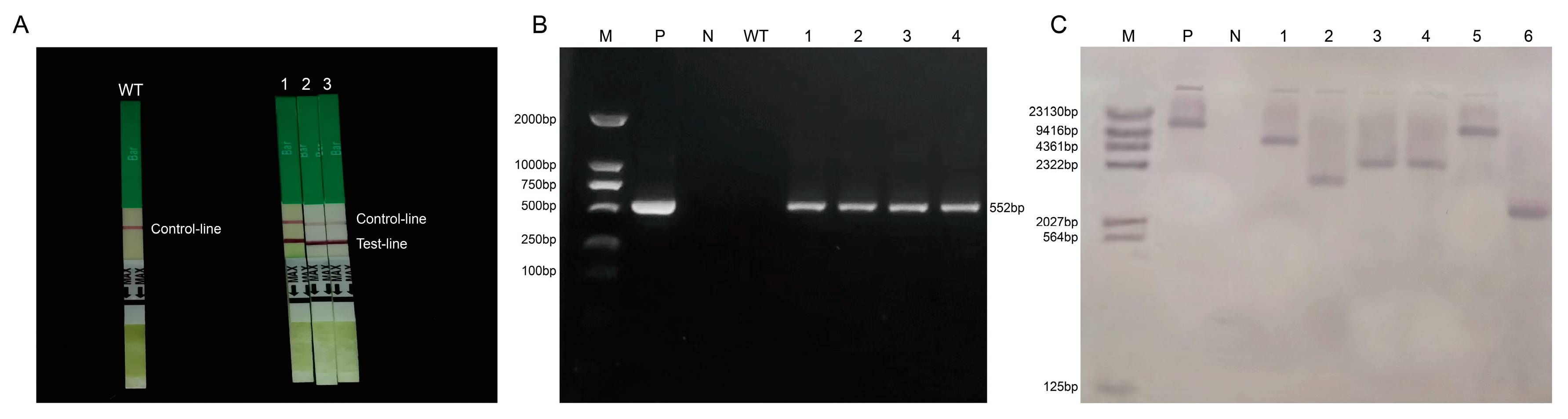
2.5. Genetic Transformation and Identification of Positive Plants
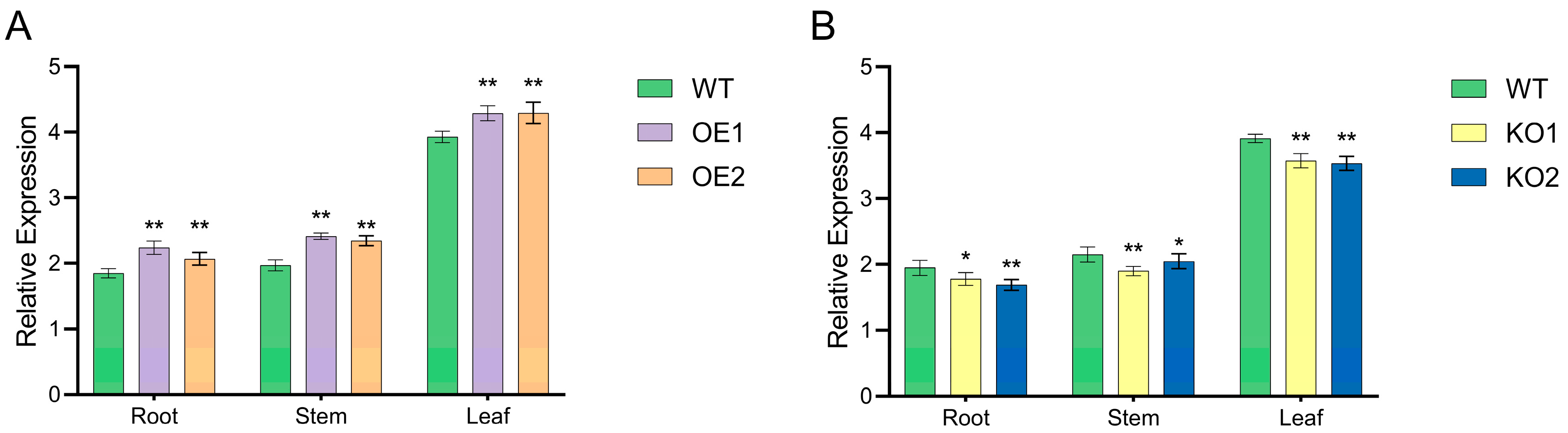
2.6. Fluorescence Quantitative PCR Detection of T2 Generation Positive Plants
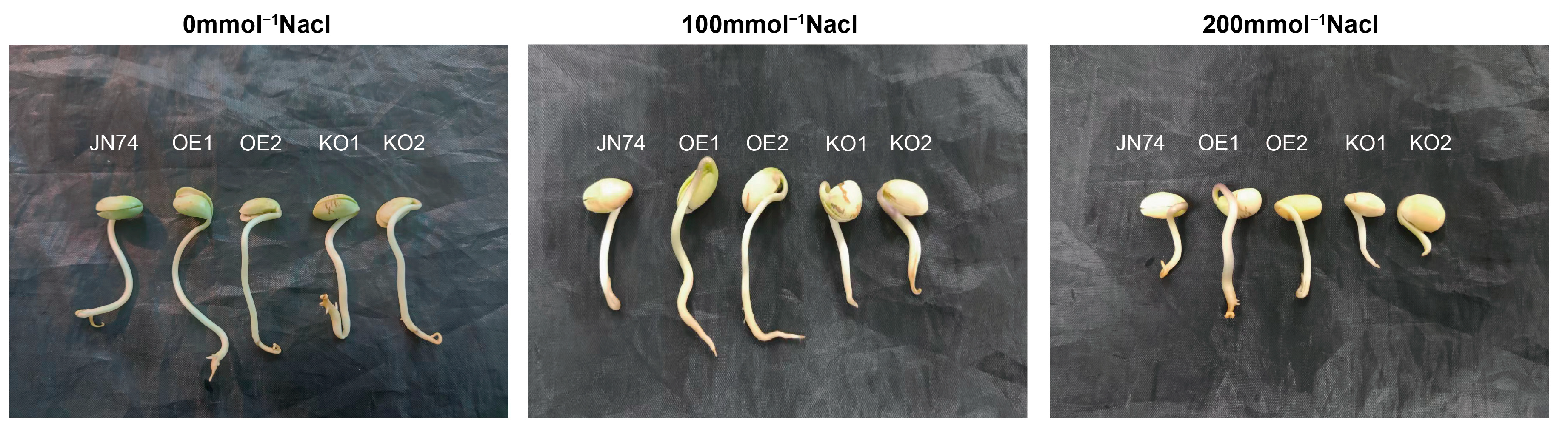
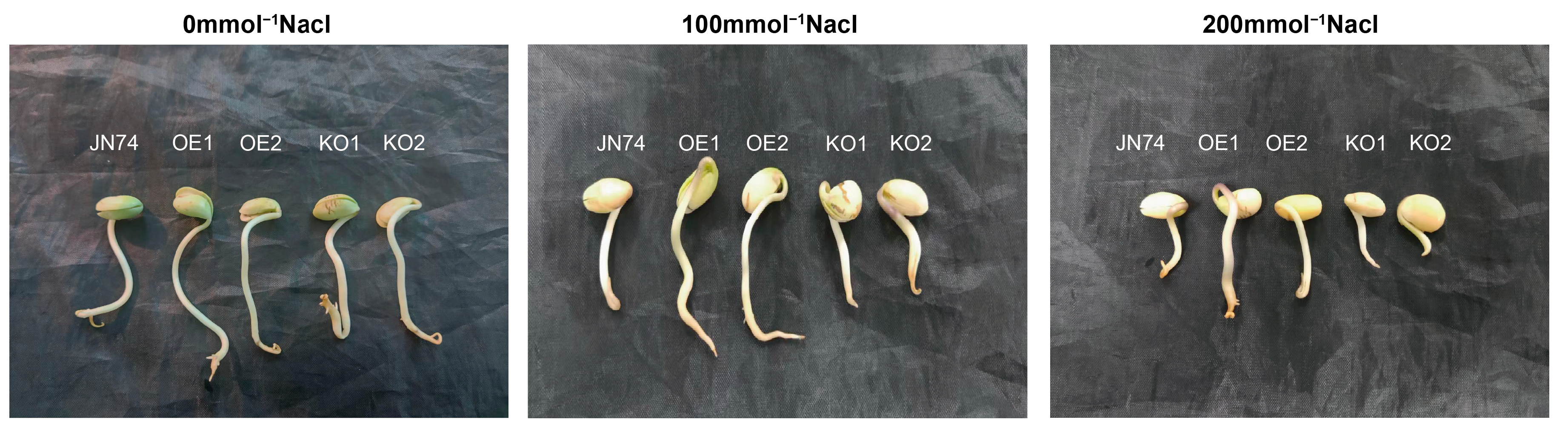
2.7. Identification of Salt Tolerance in Transgenic GmNHL1 Soybean during Germination
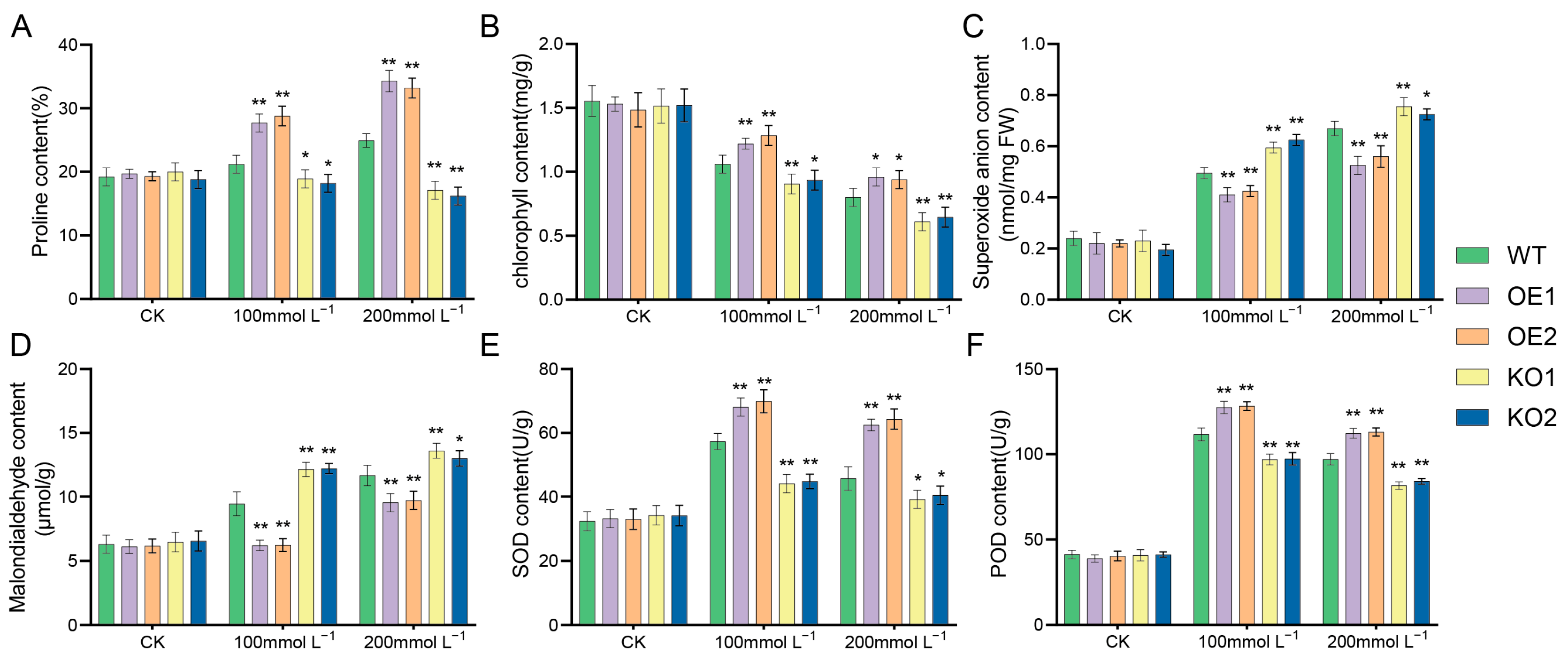
2.8. Comparative Examination of Various Soybean Strains’ Responses to Distinct Salt Stress Conditions
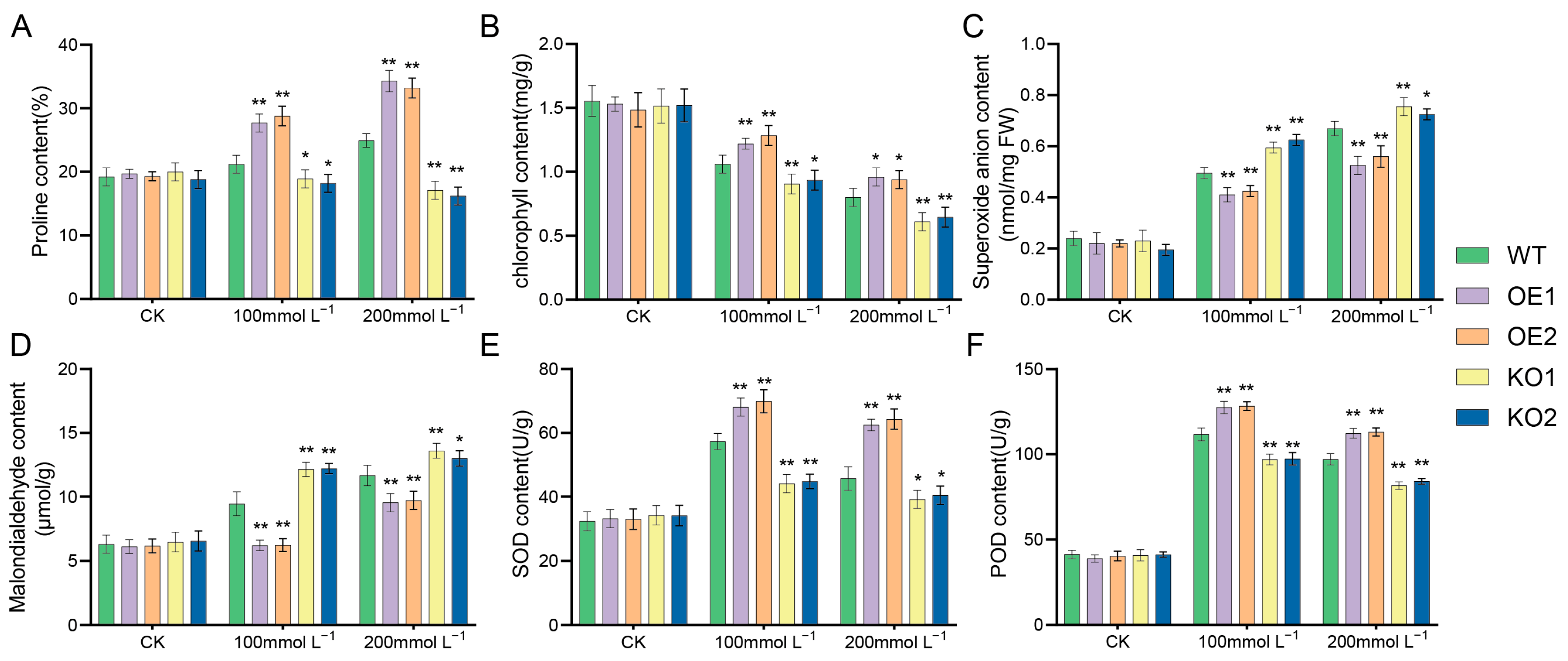
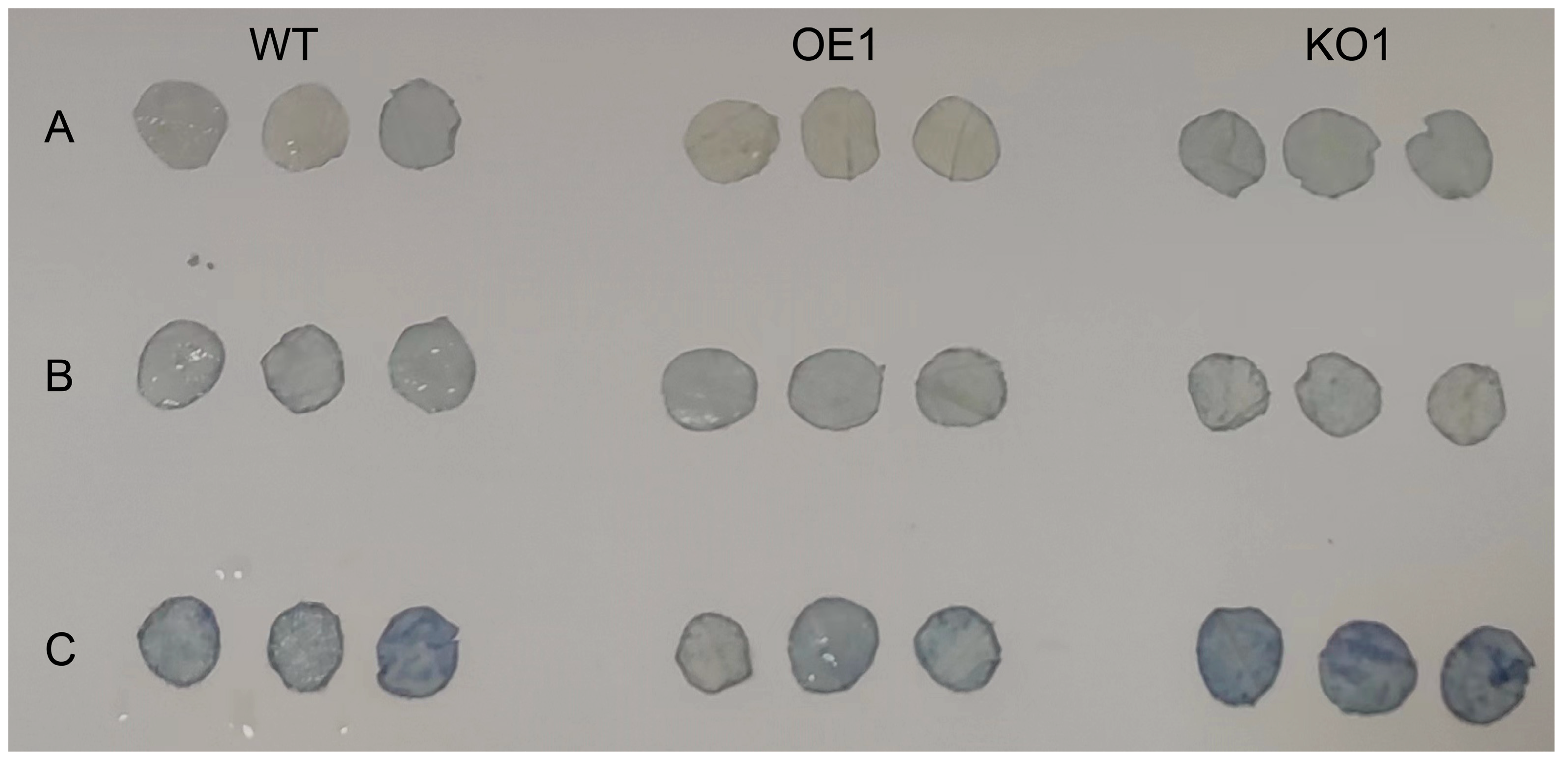
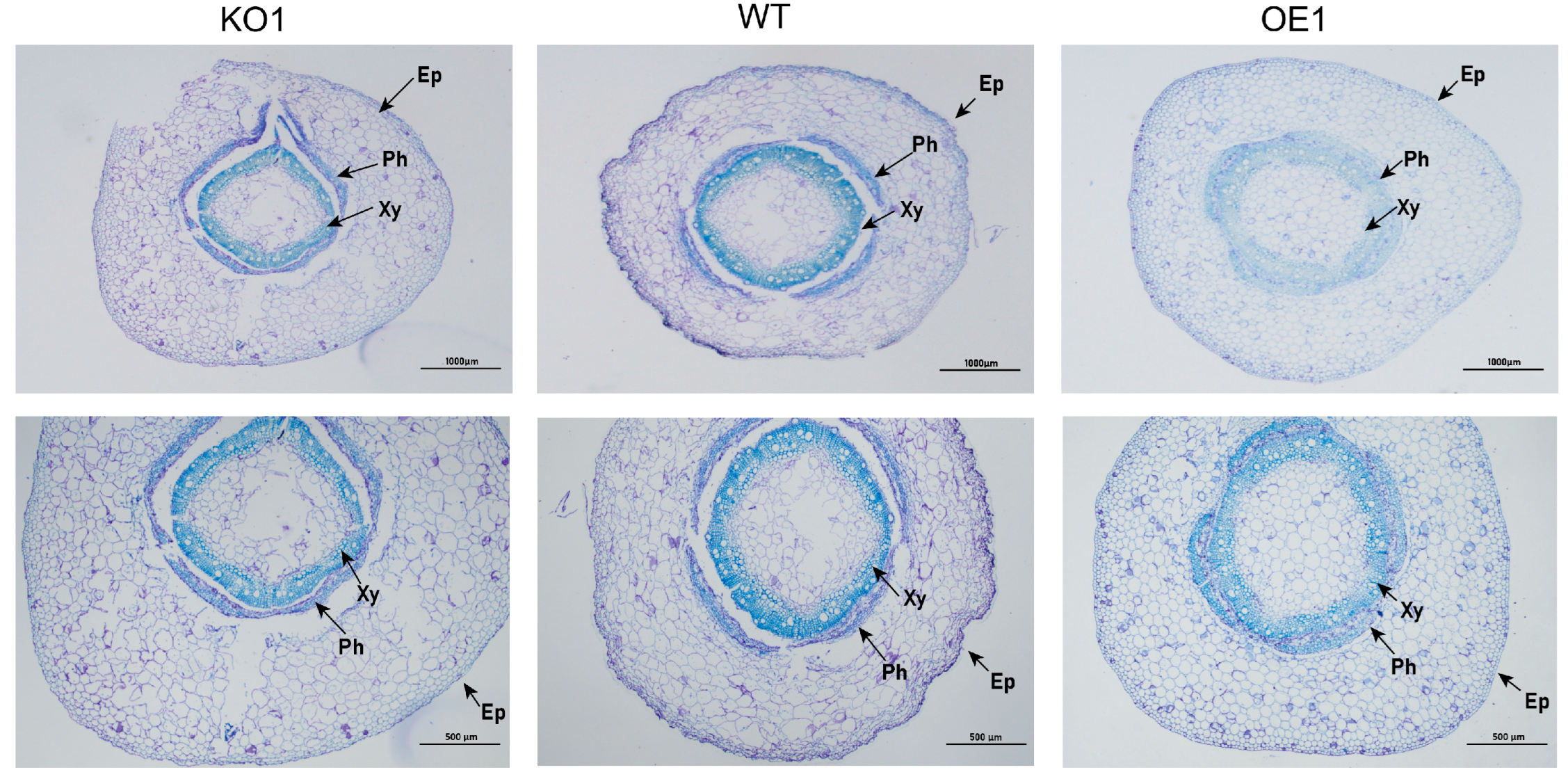
2.9. Effects of Salt Stress at Seedling Stage (V1 Stage) on Soybean Phenotype and Microstructure
3. Discussion
4. Materials and Methods
4.1. Test Materials
4.2. Cloning and Subcellular Localization of Genes
4.3. Construction of Plant Expression Vector
4.4. Genetic Transformation and Detection of Positive Plants
4.5. Germination and Seedling Test
4.6. Determination of O2− Content and Physiological and Biochemical Indexes in Soybean Plants
4.7. Real-Time Quantitative PCR (RT-qPCR)
4.8. Phenotypic Analysis of Soybean under Different Degrees of Salt Stress
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, Q.; Gan, Z.; Wang, Y.; Lu, S.; Hou, Z.; Li, H.; Xiang, H.; Liu, B.; Kong, F.; Dong, L. The Soybean Gene J Contributes to Salt Stress Tolerance by Up-Regulating Salt-Responsive Genes. Front. Plant Sci. 2020, 11, 272. [Google Scholar] [CrossRef]
- Chele, K.H.; Tinte, M.M.; Piater, L.A.; Dubery, I.A.; Tugizimana, F. Soil Salinity, a Serious Environmental Issue and Plant Responses: A Metabolomics Perspective. Metabolites 2021, 11, 724. [Google Scholar] [CrossRef]
- Song, X.; Su, Y.; Zheng, J.; Zhang, Z.; Liang, Z.; Tang, Z. Study on the Effects of Salt Tolerance Type, Soil Salinity and Soil Characteristics on the Element Composition of Chenopodiaceae Halophytes. Plants 2022, 11, 1288. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, X.; Zhang, H.; Yuan, X.; Gu, H.; Cui, X.; Chen, X. Advances in Salinity Tolerance of Soybean: Genetic Diversity, Heredity, and Gene Identification Contribute to Improving Salinity Tolerance. J. Integr. Agric. 2018, 17, 2215–2221. [Google Scholar] [CrossRef]
- Guan, R.; Yu, L.; Liu, X.; Li, M.; Chang, R.; Gilliham, M.; Qiu, L. Selection of the Salt Tolerance Gene GmSALT3 During Six Decades of Soybean Breeding in China. Front. Plant Sci. 2021, 12, 794241. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Tang, J.; Zhang, J. Effects of Salt Stress on the Morphology, Growth and Physiological Parameters of Juglansmicrocarpa L. Seedlings. Plants 2022, 11, 2381. [Google Scholar] [CrossRef] [PubMed]
- Shelke, D.B.; Pandey, M.; Nikalje, G.C.; Zaware, B.N.; Suprasanna, P.; Nikam, T.D. Salt Responsive Physiological, Photosynthetic and Biochemical Attributes at Early Seedling Stage for Screening Soybean Genotypes. Plant Physiol. Biochem. 2017, 118, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ni, Y.; Qiao, T.; Ji, X.; Xu, J.; Li, B.; Sun, Q. Overexpression of VvASMT1 from Grapevine Enhanced Salt and Osmotic Stress Tolerance in Nicotiana Benthamiana. PLoS ONE 2022, 17, e0269028. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Maximova, E.; Fuggi, A.; Carillo, P. Durum Wheat Roots Adapt to Salinity Remodeling the Cellular Content of Nitrogen Metabolites and Sucrose. Front. Plant Sci. 2017, 7, 2035. [Google Scholar] [CrossRef]
- Park, J.S.; Kim, H.J.; Cho, H.S.; Jung, H.W.; Cha, J.-Y.; Yun, D.-J.; Oh, S.-W.; Chung, Y.-S. Overexpression of AtYUCCA6 in Soybean Crop Results in Reduced ROS Production and Increased Drought Tolerance. Plant Biotechnol. Rep. 2019, 13, 161–168. [Google Scholar] [CrossRef]
- Zhang, W.; Hu, S.; Yin, J.-J.; He, W.; Lu, W.; Ma, M.; Gu, N.; Zhang, Y. Prussian Blue Nanoparticles as Multienzyme Mimetics and Reactive Oxygen Species Scavengers. J. Am. Chem. Soc. 2016, 138, 5860–5865. [Google Scholar] [CrossRef]
- Yao, S.-H.; Guo, Y.; Wang, Y.-Z.; Zhang, D.; Xu, L.; Tang, W.-H. A Cytoplasmic Cu-Zn Superoxide Dismutase SOD1 Contributes to Hyphal Growth and Virulence of Fusarium Graminearum. Fungal Genet. Biol. 2016, 91, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, H.; Yu, F.; Hu, B.; Jia, Y.; Sha, H.; Zhao, H. Differential Activity of the Antioxidant Defence System and Alterations in the Accumulation of Osmolyte and Reactive Oxygen Species under Drought Stress and Recovery in Rice (Oryza sativa L.) Tillering. Sci. Rep. 2019, 9, 8543. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Li, Y.; Wang, X.; Yan, C.; Ma, C.; Liu, J.; Dong, S. Effects of Different Drought Degrees on Physiological Characteristics and Endogenous Hormones of Soybean. Plants 2022, 11, 2282. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.H.; Vu, L.T.K.; Nguyen, L.T.N.; Pham, N.T.T.; Nguyen, Y.T.H.; Le, S.V.; Chu, M.H. Overexpression of the GmDREB6 Gene Enhances Proline Accumulation and Salt Tolerance in Genetically Modified Soybean Plants. Sci. Rep. 2019, 9, 19663. [Google Scholar] [CrossRef]
- Jin, T.; Sun, Y.; Zhao, R.; Shan, Z.; Gai, J.; Li, Y. Overexpression of Peroxidase Gene GsPRX9 Confers Salt Tolerance in Soybean. Int. J. Mol. Sci. 2019, 20, 3745. [Google Scholar] [CrossRef]
- Zhang, X.; Xue, Y.; Wang, H.; Nisa, Z.; Jin, X.; Yu, L.; Liu, X.; Yu, Y.; Chen, C. Genome-Wide Identification and Characterization of NHL Gene Family in Response to Alkaline Stress, ABA and MEJA Treatments in Wild Soybean (Glycine soja). PeerJ 2022, 10, e14451. [Google Scholar] [CrossRef]
- Wang, J.; Wu, R.; Shangguan, T.; Chen, G.; Zheng, Y.; Tao, X.; Li, S.; Wang, Y.; Xu, S. NDR1/HIN1-like Genes May Regulate Glycine Max Seed Germination under Chilling Stress through the ABA Pathway. Plant Growth Regul. 2022, 98, 613–624. [Google Scholar] [CrossRef]
- Wu, N.; Lu, Q.; Wang, P.; Zhang, Q.; Zhang, J.; Qu, J.; Wang, N. Construction and Analysis of GmFAD2-1A and GmFAD2-2A Soybean Fatty Acid Desaturase Mutants Based on CRISPR/Cas9 Technology. Int. J. Mol. Sci. 2020, 21, 1104. [Google Scholar] [CrossRef]
- Muvunyi, B.; Yan, Q.; Wu, F.; Min, X.; Yan, Z.; Kanzana, G.; Wang, Y.; Zhang, J. Mining Late Embryogenesis Abundant (LEA) Family Genes in Cleistogenes Songorica, a Xerophyte Perennial Desert Plant. Int. J. Mol. Sci. 2018, 19, 3430. [Google Scholar] [CrossRef]
- Song, W.; Cheng, Z.; Guo, X.; Yu, C.; Wang, H.; Wang, J.; Li, B.; Zhang, H.; Bao, Y. Overexpression of NHL6 Affects Seed Production in Transgenic Arabidopsis Plants. Plant Growth Regul. 2019, 88, 41–47. [Google Scholar] [CrossRef]
- Liu, C.; Peng, H.; Li, X.; Liu, C.; Lv, X.; Wei, X.; Zou, A.; Zhang, J.; Fan, G.; Ma, G.; et al. Genome-Wide Analysis of NDR1/HIN1-like Genes in Pepper (Capsicum Annuum L.) and Functional Characterization of CaNHL4 under Biotic and Abiotic Stresses. Hortic. Res. 2020, 7, 93. [Google Scholar] [CrossRef] [PubMed]
- Samaradivakara, S.P.; Chen, H.; Lu, Y.; Li, P.; Kim, Y.; Tsuda, K.; Mine, A.; Day, B. Overexpression of NDR1 Leads to Pathogen Resistance at Elevated Temperatures. New Phytol. 2022, 235, 1146–1162. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Klessig, D.F.; Nürnberger, T. A Harpin Binding Site in Tobacco Plasma Membranes Mediates Activation of the Pathogenesis-Related Gene HIN1 Independent of Extracellular Calcium but Dependent on Mitogen-Activated Protein Kinase Activity. Plant Cell 2001, 13, 1079–1093. [Google Scholar] [CrossRef] [PubMed]
- Cacas, J.-L.; Petitot, A.-S.; Bernier, L.; Estevan, J.; Conejero, G.; Mongrand, S.; Fernandez, D. Identification and Characterization of the Non-Race Specific Disease Resistance 1 (NDR1) Orthologous Protein in Coffee. BMC Plant Biol. 2011, 11, 144. [Google Scholar] [CrossRef]
- Century, K.S.; Shapiro, A.D.; Repetti, P.P.; Dahlbeck, D.; Holub, E.; Staskawicz, B.J. NDR1, a Pathogen-Induced Component Required for Arabidopsis Disease Resistance. Science 1997, 278, 1963–1965. [Google Scholar] [CrossRef]
- Gayatri, G.; Agurla, S.; Kuchitsu, K.; Anil, K.; Podile, A.R.; Raghavendra, A.S. Stomatal Closure and Rise in ROS/NO of Arabidopsis Guard Cells by Tobacco Microbial Elicitors: Cryptogein and Harpin. Front. Plant Sci. 2017, 8, 1096. [Google Scholar] [CrossRef]
- Li, Y.; Xian, X.; Guo, L.; Zhang, J.; Gan, C.; Wang, Z.; Li, H.; Li, X.; Yuan, X.; Zhang, N.; et al. CsbZIP50 Binds to the G-Box/ABRE Motif in CsRD29A Promoter to Enhance Drought Tolerance in Cucumber. Environ. Exp. Bot. 2022, 199, 104884. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, Q.; Shao, Y.; Zhang, B.; Feng, A.; Meng, F.; Li, W. GmLEA2-1, a Late Embryogenesis Abundant Protein Gene Isolated from Soybean (Glycine max (L.) Merr.), Confers Tolerance to Abiotic Stress. Acta Biol. Hung. 2018, 69, 270–282. [Google Scholar] [CrossRef]
- Wang, L.; Wen, S.; Wang, R.; Wang, C.; Gao, B.; Lu, M. PagWOX11/12a Activates PagCYP736A12 Gene That Facilitates Salt Tolerance in Poplar. Plant Biotechnol. J. 2021, 19, 2249–2260. [Google Scholar] [CrossRef]
- Silva, B.R.S.; Batista, B.L.; Lobato, A.K.S. Anatomical Changes in Stem and Root of Soybean Plants Submitted to Salt Stress. Plant Biol. J. 2021, 23, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Liu, Z.; Zou, X.; Xu, Y.; Peng, L.; Hu, J.; Lin, H. Silencing of Rice PPR Gene PPS1 Exhibited Enhanced Sensibility to Abiotic Stress and Remarkable Accumulation of ROS. J. Plant Physiol. 2021, 258–259, 153361. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Lv, J.; Qiu, N.; Bai, Y.; Yang, N.; Dong, W. The Constitutive Expression of Alfalfa MsMYB2L Enhances Salinity and Drought Tolerance of Arabidopsis Thaliana. Plant Physiol. Biochem. 2019, 141, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Karikari, B.; Wang, Z.; Zhou, Y.; Yan, W.; Feng, J.; Zhao, T. Identification of Quantitative Trait Nucleotides and Candidate Genes for Soybean Seed Weight by Multiple Models of Genome-Wide Association Study. BMC Plant Biol. 2020, 20, 404. [Google Scholar] [CrossRef]
- Breygina, M.; Voronkov, A.; Galin, I.; Akhiyarova, G.; Polevova, S.; Klimenko, E.; Ivanov, I.; Kudoyarova, G. Dynamics of Endogenous Levels and Subcellular Localization of ABA and Cytokinins during Pollen Germination in Spruce and Tobacco. Protoplasma 2023, 260, 237–248. [Google Scholar] [CrossRef]
- Matveeva, T.V. Agrobacterium-Mediated Transformation in the Evolution of Plants. In Agrobacterium Biology; Gelvin, S.B., Ed.; Current Topics in Microbiology and Immunology; Springer International Publishing: Cham, Switzerland, 2018; Volume 418, pp. 421–441. ISBN 978-3-030-03256-2. [Google Scholar]
- Li, T.; Sun, J.; Bi, Y.; Peng, Z. Overexpression of an MYB-Related Gene FvMYB1 from Fraxinus Velutina Increases Tolerance to Salt Stress in Transgenic Tobacco. J. Plant Growth Regul. 2016, 35, 632–645. [Google Scholar] [CrossRef]
- Zhou, X.; Tian, Y.; Qu, Z.; Wang, J.; Han, D.; Dong, S. Comparing the Salt Tolerance of Different Spring Soybean Varieties at the Germination Stage. Plants 2023, 12, 2789. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Fu, J.; Du, Y.; Qu, J.; Song, Y.; Wang, P. The GmXTH1 Gene Improves Drought Stress Resistance of Soybean Seedlings. Mol. Breed. 2022, 42, 3. [Google Scholar] [CrossRef]
- Fryer, M.J.; Oxborough, K.; Mullineaux, P.M.; Baker, N.R. Imaging of Photo-Oxidative Stress Responses in Leaves. J. Exp. Bot. 2002, 53, 1249–1254. [Google Scholar]
- Wang, C.; Chen, N.; Liu, J.; Jiao, P.; Liu, S.; Qu, J.; Guan, S.; Ma, Y. Overexpression of ZmSAG39 in Maize Accelerates Leaf Senescence in Arabidopsis Thaliana. Plant Growth Regul. 2022, 98, 451–463. [Google Scholar] [CrossRef]
- Nielsen, S.L.; Hansen, B.W. Evaluation of the Robustness of Optical Density as a Tool for Estimation of Biomass in Microalgal Cultivation: The Effects of Growth Conditions and Physiological State. Aquac. Res. 2019, 50, 2698–2706. [Google Scholar] [CrossRef]
- Manna, M.; Thakur, T.; Chirom, O.; Mandlik, R.; Deshmukh, R.; Salvi, P. Transcription Factors as Key Molecular Target to Strengthen the Drought Stress Tolerance in Plants. Physiol. Plant. 2021, 172, 847–868. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Chen, L.; Liu, D.; Sun, D.; Shi, G.; Yin, Y.; Wen, D.; Wang, Z.; Ai, J. Enhancement of Photosynthetic Capacity in Spongy Mesophyll Cells in White Leaves of Actinidia Kolomikta. Front. Plant Sci. 2022, 13, 856732. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Determination of Indicators | Normal | ||||
| JN74 | OE1 | OE2 | KO1 | KO2 | |
| GR (%) | 98.45 ± 1.67 | 99.12 ± 2.16 * | 98.33 ± 1.67 * | 98.33 ± 1.67 | 98.33 ± 1.67 |
| GI | 9.21 ± 0.03 | 9.57 ± 0.10 * | 9.34 ± 0.27 * | 9.51 ± 0.18 | 9.21 ± 0.14 |
| GE (%) | 78.45 ± 0.12 | 77.61 ± 0.16 * | 77.82 ± 0.28 * | 78.56 ± 1.01 | 77.27 ± 1.12 |
| VI | 6.47 ± 0.12 | 7.39 ± 0.12 * | 8.71 ± 0.21 * | 7.01 ± 0.13 | 7.12 ± 0.14 * |
| Determination of Indicators | 100 mmol L−1 NaCl | ||||
| JN74 | OE1 | OE2 | KO1 | KO2 | |
| GR (%) | 98.44 ± 1.14 | 98.05 ± 1.70 ** | 98.69 ± 2.89 ** | 96.67 ± 1.67 ** | 97.33 ± 2.67 ** |
| GI | 7.73 ± 0.17 | 8.61 ± 0.12 ** | 9.08 ± 0.18 ** | 6.71 ± 0.21 ** | 7.03 ± 0.18 ** |
| GE (%) | 65.05 ± 0.15 | 68.15 ± 0.16 ** | 66.16 ± 0.95 ** | 63.54 ± 0.85 ** | 62.96 ± 0.75 ** |
| VI | 5.38 ± 0.14 | 6.56 ± 0.13 ** | 6.81 ± 0.12 ** | 4.55 ± 0.14 ** | 3.83 ± 0.12 ** |
| Determination of Indicators | 200 mmol L−1 NaCl | ||||
| JN74 | OE1 | OE2 | KO1 | KO2 | |
| GR (%) | 68.33 ± 2.01 | 86.76 ± 2.26 ** | 90.00 ± 1.77 ** | 69.67 ± 1.41 ** | 61.67 ± 1.28 ** |
| GI | 4.08 ± 0.25 | 5.71 ± 0.18 ** | 5.19 ± 0.23 ** | 3.95 ± 0.17 ** | 3.47 ± 0.22 ** |
| GE (%) | 47.45 ± 0.23 | 59.38 ± 0.41 ** | 54.36 ± 0.36 ** | 46.84 ± 0.46 ** | 47.35 ± 0.67 ** |
| VI | 1.83 ± 0.12 | 2.24 ± 0.12 ** | 2.32 ± 0.11 ** | 1.71 ± 0.15 ** | 1.12 ± 0.11 ** |
| Normal | |||||
| JN74 | OE1 | OE2 | KO1 | KO2 | |
| The dry weight (g) | 0.32 ± 0.15 | 0.35 ± 0.23 | 0.37 ± 0.26 | 0.33 ± 2.31 | 0.31 ± 0.76 |
| Total root length (cm) | 129 ± 1.21 | 172.41 ± 0.77 | 158.42 ± 0.91 | 102.51 ± 1.73 | 99.71 ± 2.41 |
| 100 mmol L−1 NaCl | |||||
| JN74 | OE1 | OE2 | KO1 | KO2 | |
| The dry weight (g) | 0.39 ± 0.17 | 0.45 ± 0.25* | 0.47 ± 1.31 * | 0.35 ± 1.74 * | 0.34 ± 2.14 * |
| Total root length (cm) | 131.65 ± 0.26 | 175.53 ± 2.61** | 160.13 ± 0.89 ** | 105.74 ± 2.97 ** | 100.76 ± 3.45 ** |
| 200 mmol L−1 NaCl | |||||
| JN74 | OE1 | OE2 | KO1 | KO2 | |
| The dry weight (g) | 0.43 ± 2.01 | 0.56 ± 2.26 * | 0.61 ± 1.77 * | 0.39 ± 1.41 * | 0.38 ± 1.28 * |
| Total root length (cm) | 135.61 ± 1.11 ** | 178.45 ± 0.76 ** | 166.41 ± 2.35 ** | 107.41 ± 0.87 ** | 103.28 ± 1.01 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Wang, J.; Zhang, Q.; Sun, T.; Wang, P. Cloning of the Soybean GmNHL1 Gene and Functional Analysis under Salt Stress. Plants 2023, 12, 3869. https://doi.org/10.3390/plants12223869
Liu L, Wang J, Zhang Q, Sun T, Wang P. Cloning of the Soybean GmNHL1 Gene and Functional Analysis under Salt Stress. Plants. 2023; 12(22):3869. https://doi.org/10.3390/plants12223869
Chicago/Turabian StyleLiu, Lu, Jiabao Wang, Qi Zhang, Tingting Sun, and Piwu Wang. 2023. "Cloning of the Soybean GmNHL1 Gene and Functional Analysis under Salt Stress" Plants 12, no. 22: 3869. https://doi.org/10.3390/plants12223869
APA StyleLiu, L., Wang, J., Zhang, Q., Sun, T., & Wang, P. (2023). Cloning of the Soybean GmNHL1 Gene and Functional Analysis under Salt Stress. Plants, 12(22), 3869. https://doi.org/10.3390/plants12223869