
The Sunflower WRINKLED1 Transcription Factor Regulates Fatty Acid Biosynthesis Genes through an AW Box Binding Sequence with a Particular Base Bias

 ,
, 
Abstract
:1. Introduction
2. Results
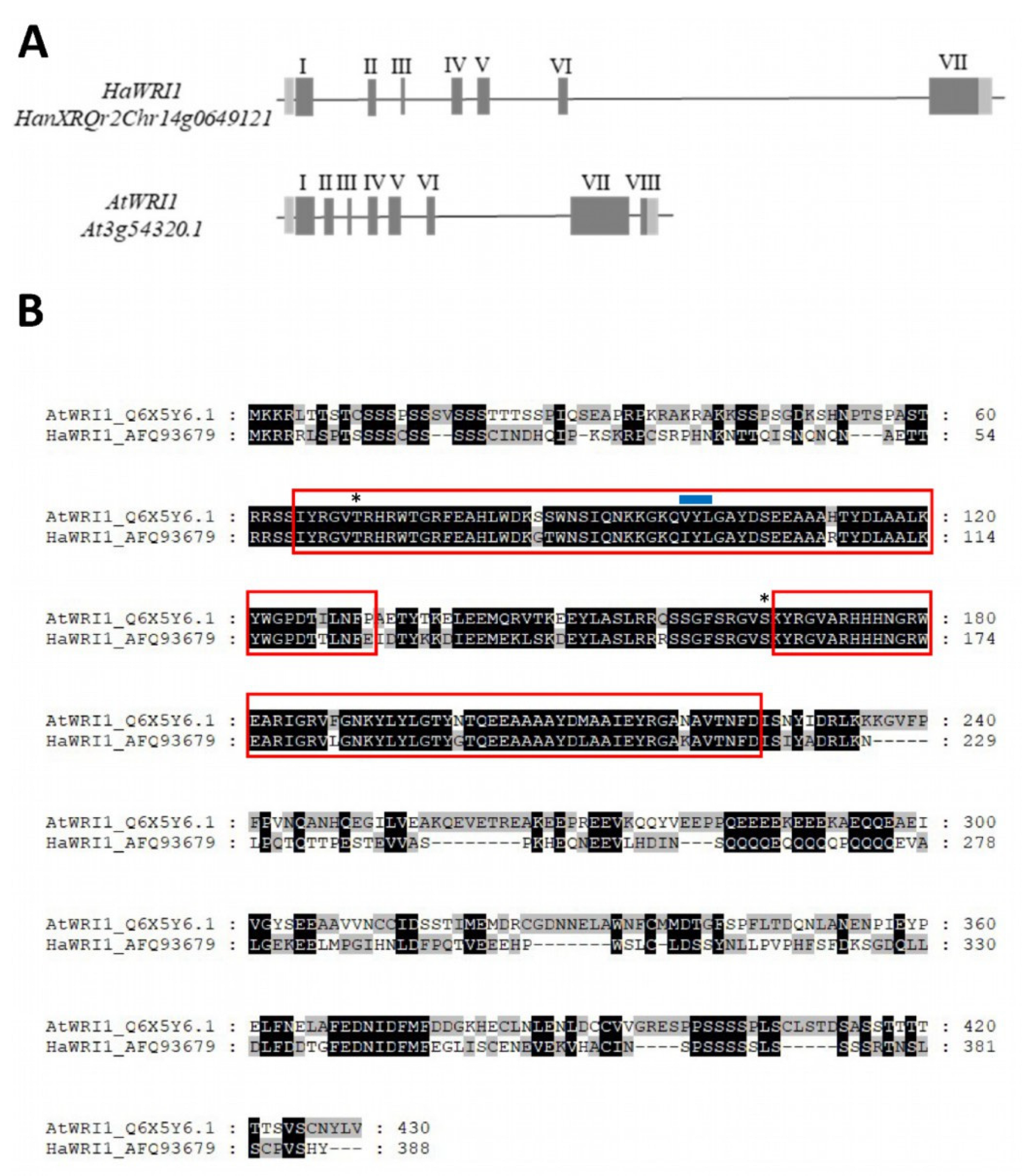
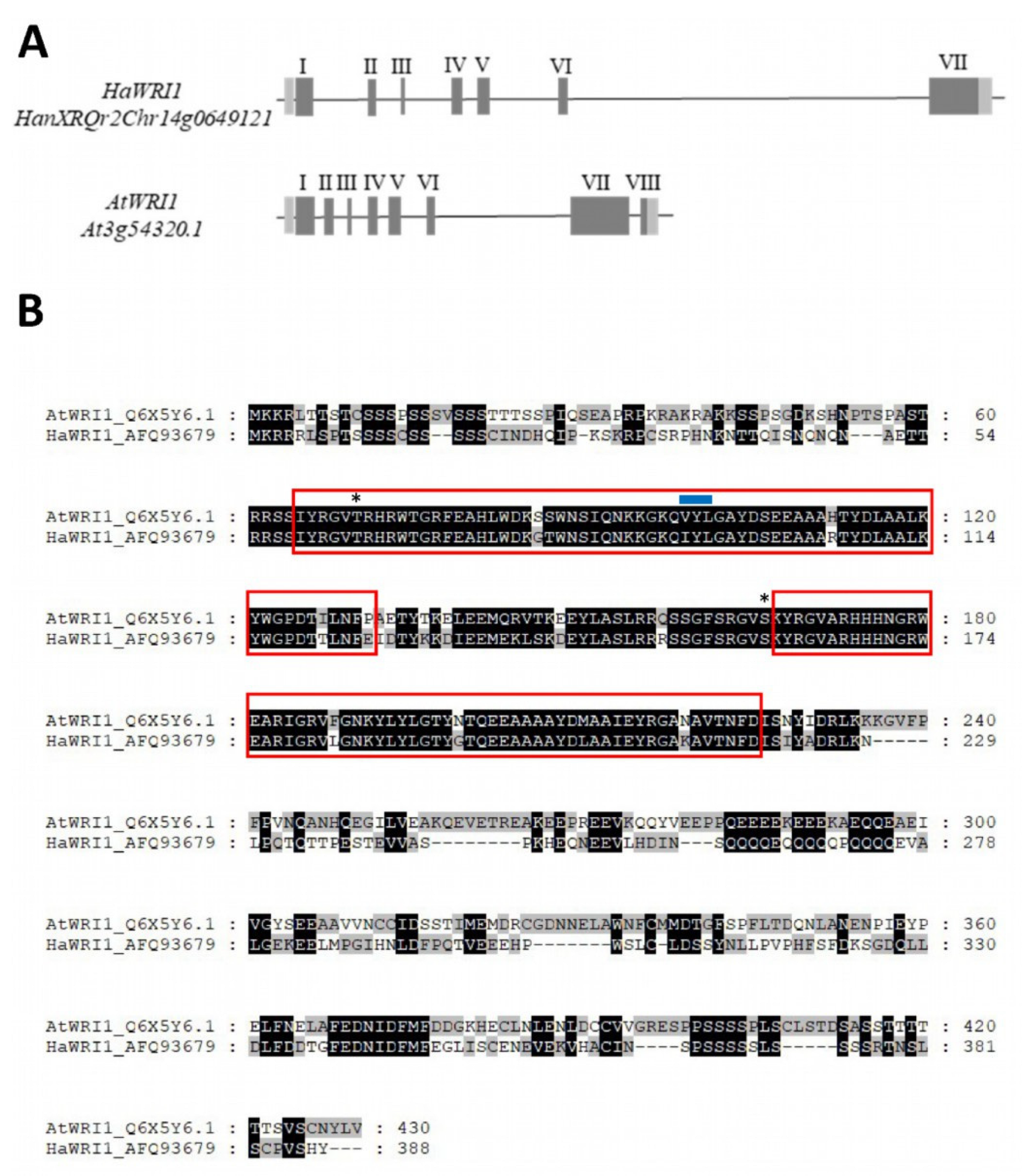
2.1. The Sunflower Genome Contains One Single WRI1 Gene
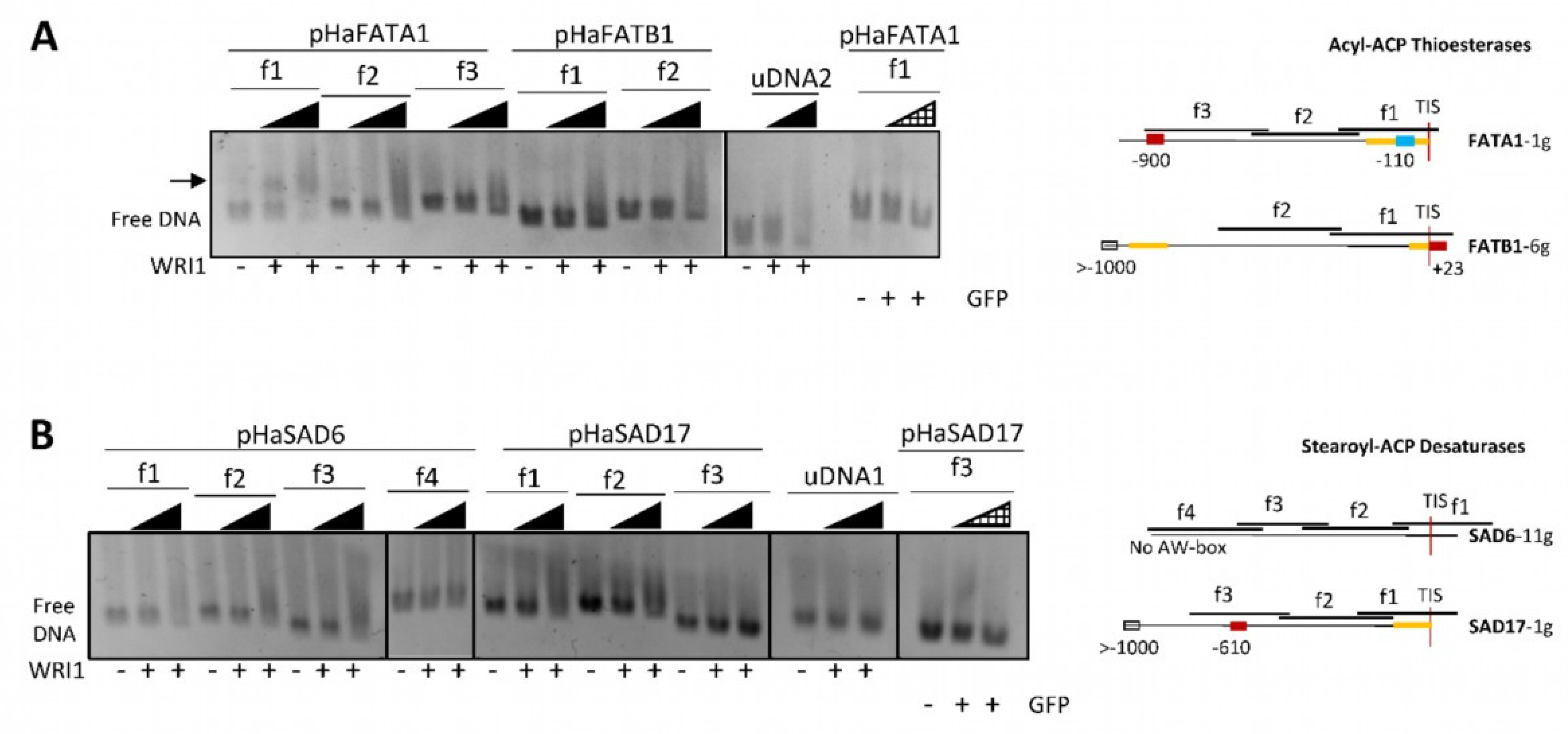
2.2. Sunflower WRI1 Binds to the AW Box Motif Present in the Acyl-ACP Thioesterase FATA1 Gene
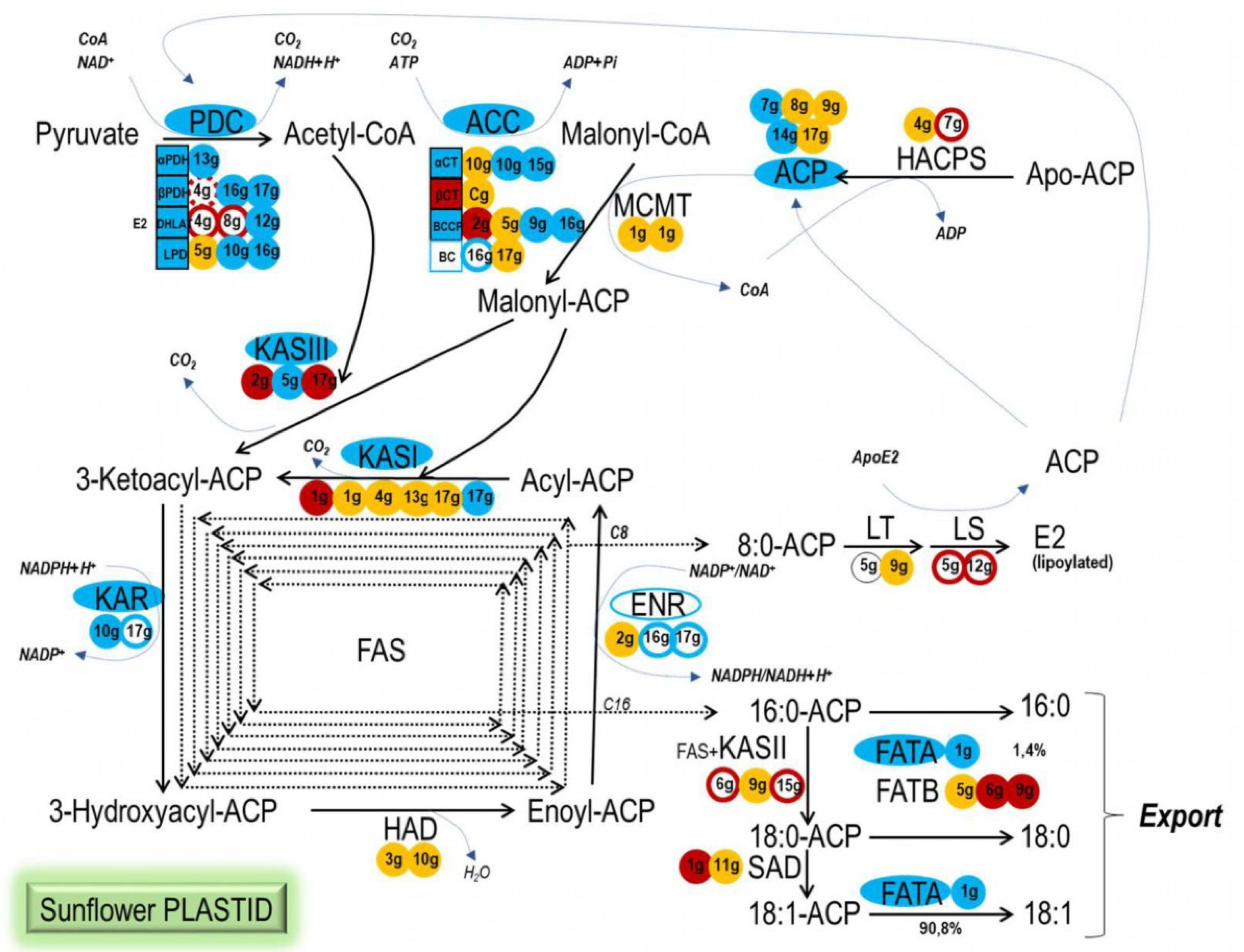
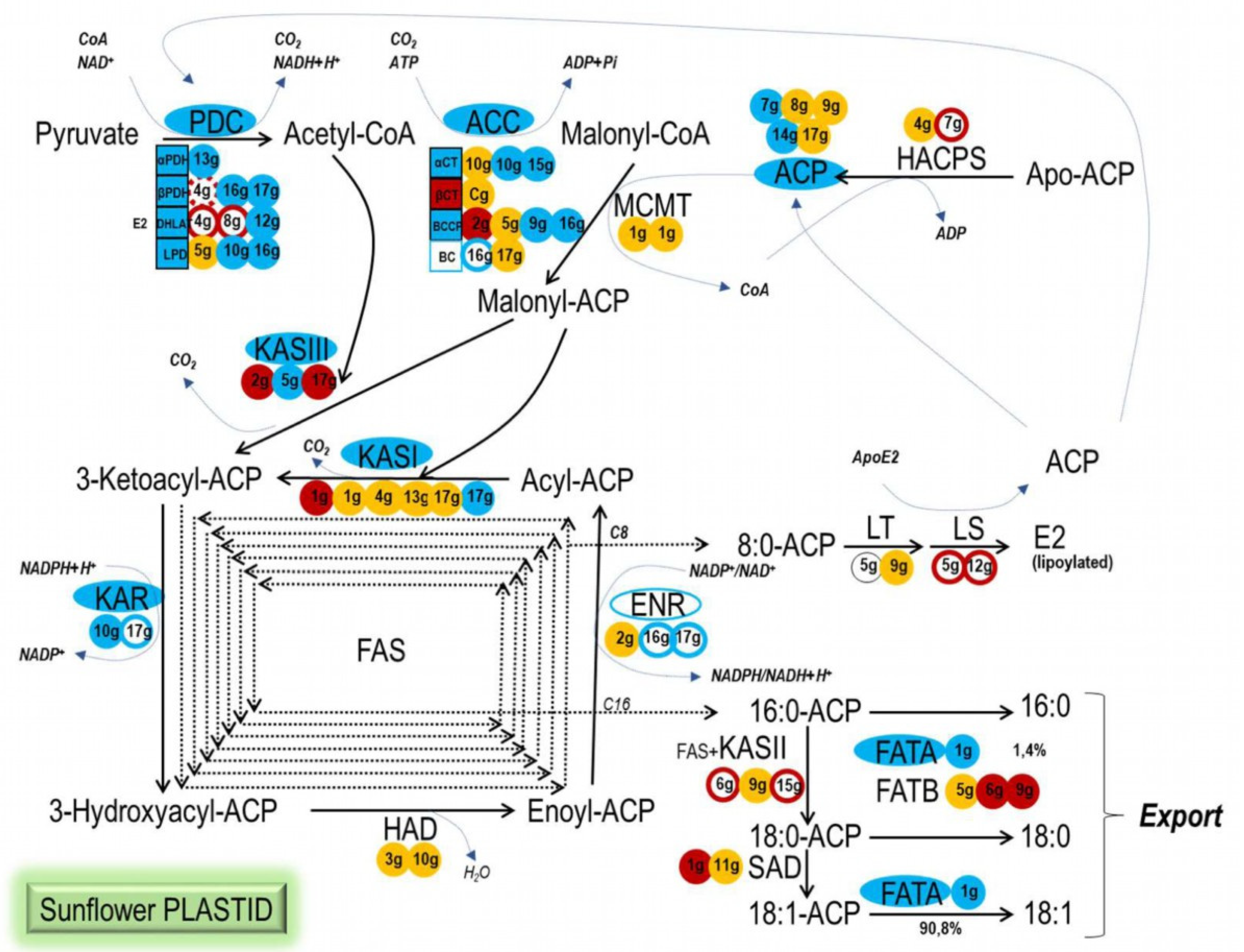
2.3. Sunflower WRI1 Regulates Plastidial Fatty Acid Synthesis Mainly at Early Steps of the Pathway
2.4. Sunflower WRI1 Can Bind to a Non-Canonical AW Box Motif
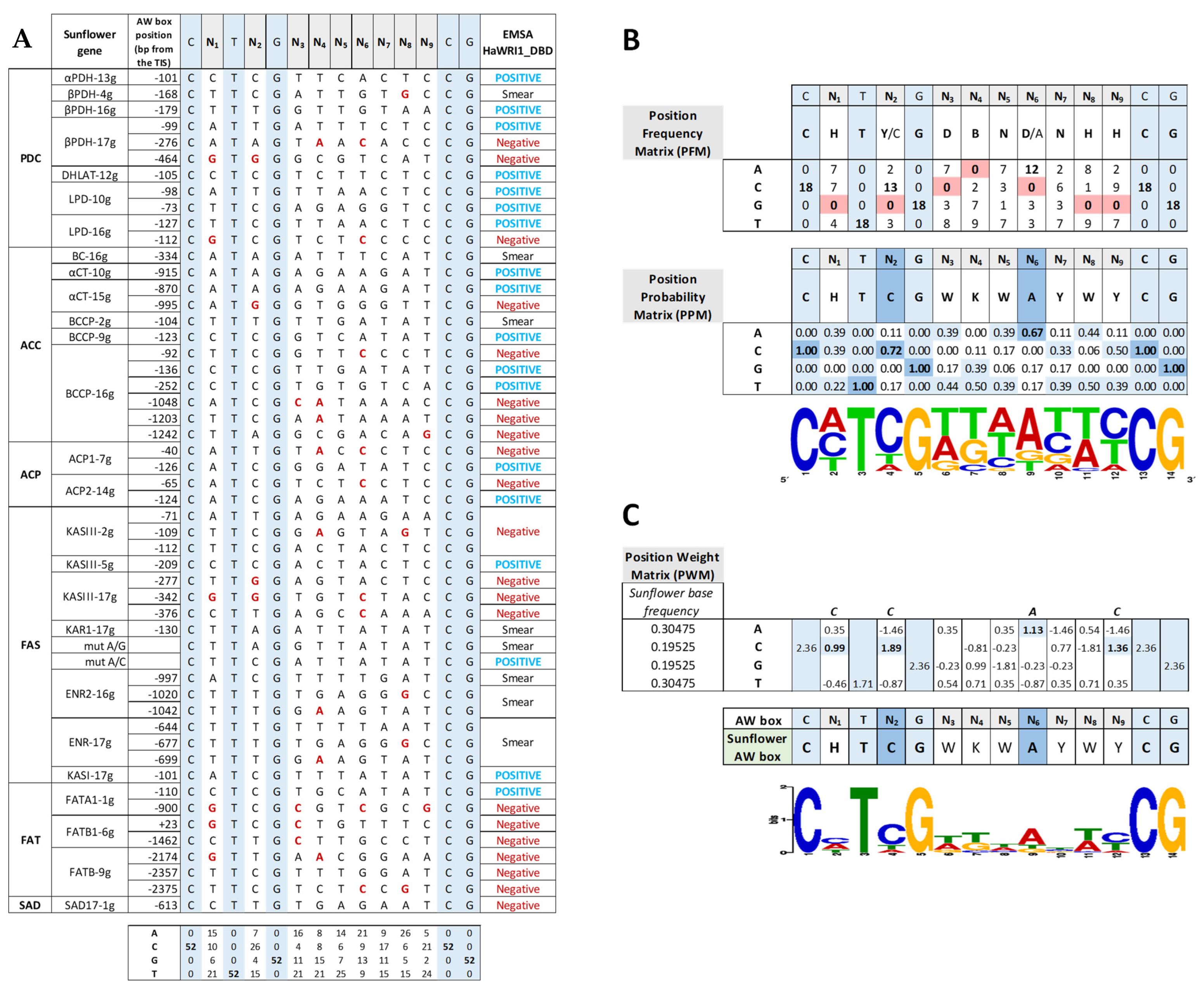
2.5. The Sunflower AW Box Motif That Regulates Transcripts Involved in FA Synthesis Shows Base Bias in Non-Conservative Motifs
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Growth Conditions
5.2. HaWRI1-DNA Binding Domain Cloning, Expression in Escherichia coli and Purification
5.3. Promoter Cloning, DNA Fragment Amplification and Purification, and Site-Directed Mutagenesis by Overlap Extension PCR
5.4. Agarose Gel Electrophoretic Mobility Shift Assays (EMSAs)
5.5. Acrylamide Gel Electrophoretic Mobility Shift Assays (EMSAs)
5.6. Bioinformatics Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapman, K.D.; Ohlrogge, J.B. Compartmentation of triacylglycerol accumulation in plants. J. Biol. Chem. 2012, 287, 2288–2294. [Google Scholar] [CrossRef]
- Ohlrogge, J.; Chapman, K. The seeds of green energy: Expanding the contribution of plant oils as biofuels. Biochemist 2011, 33, 34–38. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; Debono, A.; Durrett, T.P.; et al. Acyl-lipid metabolism. In Arabidopsis Book; American Society of Plant Biologists: Rockville, MD, USA, 2013; Volume 3, p. e0161. [Google Scholar] [CrossRef]
- Bates, P.D.; Stymne, S.; Ohlrogge, J. Biochemical pathways in seed oil synthesis. Curr. Opin. Plant Biol. 2013, 16, 358–364. [Google Scholar] [CrossRef]
- Baud, S.; Lepiniec, L. Physiological and developmental regulation of seed oil production. Prog. Lipid Res. 2010, 49, 235–249. [Google Scholar] [CrossRef]
- Schmid, M.; Davison, T.S.; Henz, S.R.; Pape, U.J.; Demar, M.; Vingron, M.; Schölkopf, B.; Weigel, D.; Lohmann, J.U. A gene expression map of Arabidopsis thaliana development. Nat. Genet. 2005, 37, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Penouilh-Suzette, C.; Pomiès, L.; Duruflé, H.; Blanchet, N.; Bonnafous, F.; Dinis, R.; Brouard, C.; Gody, L.; Grassa, C.; Heudelot, X.; et al. RNA expression dataset of 384 sunflower hybrids in field condition. OCL 2020, 27, 36. [Google Scholar] [CrossRef]
- Mentzen, W.I.; Peng, J.L.; Ransom, N.; Nikolau, B.J.; Wurtele, E.S. Articulation of three core metabolic processes in Arabidopsis: Fatty acid biosynthesis, leucine catabolism and starch metabolism. BMC Plant Biol. 2008, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Girke, T.; Todd, J.; Ruuska, S.; White, J.; Benning, C.; Ohlrogge, J. Microarray analysis of developing Arabidopsis seeds. Plant Physiol. 2000, 124, 1570–1581. [Google Scholar] [CrossRef] [PubMed]
- Ruuska, S.A.; Girke, T.; Benning, C.; Ohlrogge, J.B. Contrapuntal networks of gene expression during Arabidopsis seed filling. Plant Cell 2002, 14, 1191–1206. [Google Scholar] [CrossRef] [PubMed]
- Baud, S.; Lepiniec, L. Regulation of de novo fatty acid synthesis in maturing oilseeds of Arabidopsis. Plant Physiol. Biochem. 2009, 47, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Baud, S.; Mendoza, M.S.; To, A.; Harscoet, E.; Lepiniec, L.; Dubreucq, B. WRINKLED1 specifies the regulatory action of LEAFY COTYLEDON2 towards fatty acid metabolism during seed maturation in Arabidopsis. Plant J. 2007, 50, 825–838. [Google Scholar] [CrossRef]
- Baud, S.; Wuillème, S.; To, A.; Rochat, C.; Lepiniec, L. Role of WRINKLED1 in the transcriptional regulation of glycolytic and fatty acid biosynthetic genes in Arabidopsis. Plant J. 2009, 60, 933–947. [Google Scholar] [CrossRef] [PubMed]
- Maeo, K.; Tokuda, T.; Ayame, A.; Mitsui, N.; Kawai, T.; Tsukagoshi, H.; Ishiguro, S.; Nakamura, K. An AP2-type transcription factor, WRINKLED1, of Arabidopsis thaliana binds to the AW-box sequence conserved among proximal upstream regions of genes involved in fatty acid synthesis. Plant J. 2009, 60, 476–487. [Google Scholar] [CrossRef]
- Lepiniec, L.; Devic, M.; Roscoe, T.J.; Bouyer, D.; Zhou, D.X.; Boulard, C.; Baud, S.; Dubreucq, B. Molecular and epigenetic regulations and functions of the LAFL transcriptional regulators that control seed development. Plant Reprod. 2018, 31, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Cernac, A.; Benning, C. WRINKLED1 encodes an AP2/EREB domain protein involved in the control of storage compound biosynthesis in Arabidopsis. Plant J. 2004, 40, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Focks, N.; Benning, C. Wrinkled1: A novel, low-seed-oil mutant of Arabidopsis with a deficiency in the seed-specific regulation of carbohydrate metabolism. Plant Physiol. 1998, 118, 91–101. [Google Scholar] [CrossRef]
- Pouvreau, B.; Baud, S.; Vernoud, V.; Morin, V.; Py, C.; Gendrot, G.; Pichon, J.-P.; Rouster, J.; Paul, W.; Rogowsky, P.M. Duplicate maize wrinkled1 transcription factors activate target genes involved in seed oil biosynthesis. Plant Physiol. 2011, 156, 674–686. [Google Scholar] [CrossRef]
- To, A.; Joubès, J.; Barthole, G.; Lécureuil, A.; Scagnelli, A.; Jasinski, S.; Lepiniec, L.; Baud, S. WRINKLED transcription factors orchestrate tissue-specific regulation of fatty acid biosynthesis in Arabidopsis. Plant Cell 2012, 24, 5007–5023. [Google Scholar] [CrossRef]
- Fukuda, N.; Ikawa, Y.; Aoyagi, T.; Kozaki, A. Expression of the genes coding for plastidic acetyl-CoA carboxylase subunits is regulated by a location-sensitive transcription factor binding site. Plant Mol. Biol. 2013, 82, 473–483. [Google Scholar] [CrossRef]
- Li, Q.; Shao, J.; Tang, S.; Shen, Q.; Wang, T.; Chen, W.; Hong, Y. Wrinkled1 accelerates flowering and regulates lipid homeostasis between oil accumulation and membrane lipid anabolism in Brassica napus. Front. Plant Sci. 2015, 6, 1015. [Google Scholar] [CrossRef]
- Liu, H.; Zhai, Z.; Kuczynski, K.; Keereetaweep, J.; Schwender, J.; Shanklin, J. WRINKLED1 regulates Biotin Attachment Domain-Containing proteins that inhibit fatty acid synthesis. Plant Physiol. 2019, 181, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Kuczynski, C.; McCorkle, S.; Keereetaweep, J.; Shanklin, J.; Schwender, J. An expanded role for WRINKLED1 metabolic control based on combined phylogenetic and biochemical analyses. BioRxiv 2020, preprint. [Google Scholar] [CrossRef]
- Kazaz, S.; Barthole, G.; Domergue, F.; Ettaki, H.; To, A.; Vasselon, D.; De Vos, D.; Belcram, K.; Lepiniec, L.; Baud, S. Differential activation of partially redundant Δ9 stearoyl-ACP desaturase genes is critical for omega-9 monounsaturated fatty acid biosynthesis during seed development in Arabidopsis. Plant Cell 2020, 32, 3613–3637. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hua, W.; Zhan, G.M.; Wei, F.; Wang, X.F.; Liu, G.H.; Wang, H.Z. Increasing seed mass and oil content in transgenic Arabidopsis by the overexpression of wri1- like gene from Brassica napus. Plant Physiol. Biochem. 2010, 48, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Allen, W.B.; Zheng, P.Z.; Li, C.J.; Glassman, K.; Ranch, J.; Nubel, D.; Tarczynski, M.C. Expression of ZmLEC1 and ZmWRI1 increases seed oil production in maize. Plant Physiol. 2010, 153, 980–987. [Google Scholar] [CrossRef]
- Ma, W.; Kong, Q.; Arondel, V.; Kilaru, A.; Bates, P.D.; Thrower, N.A.; Benning, C.; Ohlrogge, J.B. WRINKLED1, a ubiquitous regulator in oil accumulating tissues from Arabidopsis embryos to oil Palm mesocarp. PLoS ONE 2013, 8, e68887. [Google Scholar] [CrossRef]
- Yang, Y.; Munz, J.; Cass, C.; Zienkiewicz, A.; Kong, Q.; Ma, W.; Sanjaya; Sedbrook, J.; Benning, C. Ectopic expression of WRINKLED1 affects fatty acid homeostasis in Brachypodium distachyon vegetative tissues. Plant Physiol. 2015, 69, 1836–1847. [Google Scholar] [CrossRef]
- Chen, B.B.; Zhang, G.Y.; Li, P.H.; Yang, J.H.; Guo, L.; Benning, C.; Wang, X.M.; Zhao, J. Multiple GmWRI1s are redundantly involved in seed filling and nodulation by regulating plastidic glycolysis, lipid biosynthesis and hormone signalling in soybean (Glycine max). Plant Biotechnol. J. 2020, 18, 155–171. [Google Scholar] [CrossRef]
- Mano, F.; Aoyanagi, T.; Kozaki, A. Atypical splicing accompanied by skipping conserved micro-exons produces unique WRINKLED1, an AP2 domain transcription factor in rice plants. Plants 2019, 8, 207. [Google Scholar] [CrossRef] [PubMed]
- Fei, W.; Yang, S.; Hu, J.; Yang, F.; Qu, G.; Peng, D.; Zhou, B. Research advances of WRINKLED1 (WRI1) in plants. Funct. Plant Biol. 2020, 47, 185–194. [Google Scholar] [CrossRef]
- Kong, Q.; Yang, Y.Z.; Guo, L.; Yuan, L.; Ma, W. Molecular basis of plant oil biosynthesis: Insights gained from studying the WRINKLED1 transcription factor. Front. Plant Sci. 2020, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Miray, R.; Kazaz, S.; To, A.; Baud, S. Molecular control of oil metabolism in the endosperm of seeds. Int. J. Mol. Sci. 2021, 22, 1621. [Google Scholar] [CrossRef]
- Zambelli, A.; León, A.; Garcés, R. Mutagenesis in Sunflower. In Sunflower Oilseed. Chemistry, Production, Processing and Utilization; Martínez-Force, E., Dunford, N.T., Salas, J.J., Eds.; AOCS Press: Urbana, IL, USA, 2015; Volume 7, pp. 27–52. [Google Scholar] [CrossRef]
- Salas, J.J.; Martínez-Force, E.; Harwood, J.L.; Venegas-Calerón, M.; Aznar-Moreno, J.A.; Moreno-Pérez, A.J.; Ruíz-López, N.; Serrano-Vega, M.J.; Graham, I.A.; Mullen, R.T.; et al. Biochemistry of high stearic sunflower, a new source of saturated fats. Prog. Lipid Res. 2014, 55, 30–42. [Google Scholar] [CrossRef]
- Venegas-Calerón, M.; Troncoso-Ponce, M.A.; Martínez-Force, E. Sunflower Oil and Lipids Biosynthesis. In Sunflower Oilseed. Chemistry, Production, Processing and Utilization; Martínez-Force, E., Dunford, N.T., Salas, J.J., Eds.; AOCS Press: Urbana, IL, USA, 2015; Volume 7, pp. 259–295. [Google Scholar] [CrossRef]
- Martins-Noguerol, R.; Moreno-Pérez, A.J.; Acket, S.; Makni, S.; Garcés, R.; Troncoso-Ponce, A.; Salas, J.J.; Thomasset, B.; Martínez-Force, E. Lipidomic analysis of plastidial octanoyltransferase mutants of Arabidopsis thaliana. Metabolites 2019, 9, 209. [Google Scholar] [CrossRef] [PubMed]
- Martins-Noguerol, R.; Moreno-Pérez, A.J.; Acket, S.; Troncoso-Ponce, M.A.; Garcés, R.; Thomasset, B.; Salas, J.J.; Martínez-Force, E. Impact of sunflower (Helianthus annuus L.) plastidial lipoyl synthases genes expression in glycerolipids composition of transgenic Arabidopsis plants. Sci. Rep. 2020, 10, 3749. [Google Scholar] [CrossRef] [PubMed]
- Aznar-Moreno, J.A.; Venegas-Calerón, M.; Du, Z.Y.; Garcés, R.; Tanner, J.A.; Chye, M.L.; Martínez-Force, E.; Salas, J.J. Characterization of a small acyl-CoA-binding protein (ACBP) from Helianthus annuus L. and its binding affinities. Plant Physiol. Biochem. 2016, 102, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Aznar-Moreno, J.A.; Venegas-Calerón, M.; Martínez-Force, E.; Garcés, R.; Salas, J.J. Acyl carrier proteins from sunflower (Helianthus annuus L.) seeds and their influence on FatA and FatB acyl-ACP thioesterase activities. Planta 2016, 244, 479–490. [Google Scholar] [CrossRef]
- Aznar-Moreno, J.A.; Sánchez, R.; Gidda, S.K.; Martínez-Force, E.; Moreno-Pérez, A.J.; Venegas-Calerón, M.; Garcés, R.; Mullen, R.T.; Salas, J.J. New insights into sunflower (Helianthus annuus L.) FatA and FatB thioesterases, their regulation, structure and distribution. Front. Plant Sci. 2018, 9, 1496. [Google Scholar] [CrossRef]
- Aznar-Moreno, J.A.; Venegas-Calerón, M.; Du, Z.Y.; Garcés, R.; Tanner, J.A.; Chye, M.L.; Martínez-Force, E.; Salas, J.J. Characterization and function of a sunflower (Helianthus annuus L.) Class II acyl-CoA-binding protein. Plant Sci. 2020, 300, 110630. [Google Scholar] [CrossRef]
- González-Thuillier, I.; Venegas-Calerón, M.; Garcés, R.; von Wettstein-Knowles, P.; Martínez-Force, E. Sunflower (Helianthus annuus) fatty acid synthase complex: Enoyl-[acyl carrier protein]-reductase genes. Planta 2015, 241, 43–56. [Google Scholar] [CrossRef]
- González-Thuillier, I.; Venegas-Calerón, M.; Sánchez, R.; Garcés, R.; von Wettstein-Knowles, P.; Martínez-Force, E. Sunflower (Helianthus annuus) fatty acid synthase complex: β-hydroxyacyl-[acyl carrier protein] dehydratase genes. Planta 2016, 243, 397–410. [Google Scholar] [CrossRef] [PubMed]
- González-Thuillier, I.; Venegas-Calerón, M.; Moreno-Pérez, A.J.; Salas, J.J.; Garcés, R.; von Wettstein-Knowles, P.; Martínez-Force, E. Sunflower (Helianthus annuus) fatty acid synthase complex: β-Ketoacyl-[acyl carrier protein] reductase genes. Plant Physiol. Biochem. 2021, 166, 689–699. [Google Scholar] [CrossRef]
- González-Mellado, D.; von Wettstein-Knowles, P.; Garcés, R.; Martínez-Force, E. The role of beta-ketoacyl-acyl carrier protein synthase III in the condensation steps of fatty acid biosynthesis in sunflower. Planta 2010, 231, 1277–1289. [Google Scholar] [CrossRef] [PubMed]
- González-Mellado, D.; Salas, J.J.; Venegas-Calerón, M.; Moreno-Pérez, A.J.; Garcés, R.; Martínez-Force, E. Functional characterization and structural modelling of Helianthus annuus (sunflower) ketoacyl-CoA synthases and role in seed oil composition. Planta 2019, 249, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Pérez, A.J.; Santos-Pereira, J.M.; Martins-Noguerol, R.; DeAndrés-Gil, C.; Troncoso-Ponce, M.A.; Venegas-Calerón, M.; Sánchez, R.; Garcés, R.; Salas, J.J.; Tena, J.J.; et al. Genome-wide mapping of histone H3 lysine 4 trimethylation (H3K4me3) and its involvement in fatty acid biosynthesis in sunflower developing seeds. Plants 2021, 10, 706. [Google Scholar] [CrossRef]
- Serrano-Vega, M.J.; Venegas-Calerón, M.; Garcés, R.; Martínez-Force, E. Cloning and expression of fatty acids biosynthesis key enzymes from sunflower (Helianthus annuus L.) in Escherichia coli. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2003, 786, 221–228. [Google Scholar] [CrossRef]
- Serrano-Vega, M.J.; Garcés, R.; Martínez-Force, E. Cloning, characterization and structural model of a FatA-type thioesterase from sunflower seeds (Helianthus annuus L.). Planta 2005, 221, 868–880. [Google Scholar] [CrossRef]
- Hongtrakul, V.; Slabaugh, M.B.; Knapp, S.J. DFLP, SSCP, and SSR markers for delta 9-stearoyl-acyl carrier protein desaturases strongly expressed in developing seeds of sunflower: Intron lengths are polymorphic among elite inbred lines. Mol. Breed. 1998, 4, 195–203. [Google Scholar] [CrossRef]
- Gill, N.; Buti, M.; Kane, N.; Bella, A.; Helmsletter, N.; Berges, H.; Rieseberg, L.H. Sequence-based analysis of structural organization and composition of the cultivated sunflower (Helianthus annuus L.) genome. Biology 2014, 3, 295–319. [Google Scholar] [CrossRef]
- An, D.; Kim, H.; Ju, S.; Go, Y.S.; Kim, H.U.; Suh, M.C. Expression of Camelina WRINKLED1 isoforms rescue the seed phenotype of the Arabidopsis wri1 mutant and increase the triacylglycerol content in tobacco leaves. Front. Plant Sci. 2017, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Tajima, D.; Kaneko, A.; Sakamoto, M.; Ito, Y.; Hue, N.; Miyazaki, M.; Ishibashi, Y.; Yuasa, T.; Iwaya-Inoue, M. Wrinkled 1 (WRI1) homologs, AP2-type transcription factors involving master regulation of seed storage oil synthesis in castor bean (Ricinus communis L.). Am. J. Plant Sci. 2013, 4, 333–339. [Google Scholar] [CrossRef]
- Ji, X.J.; Mao, X.; Hao, Q.T.; Liu, B.L.; Xue, J.A.; Li, R.Z. Splice variants of the castor WRI1 gene upregulate fatty acid and oil biosynthesis when expressed in tobacco leaves. Int. J. Mol. Sci. 2018, 19, 146. [Google Scholar] [CrossRef] [PubMed]
- Krizek, B.A. AINTEGUMENTA utilizes a mode of DNA recognition distinct from that used by proteins containing a single AP2 domain. Nucleic Acids Res. 2003, 31, 1859–1868. [Google Scholar] [CrossRef] [PubMed]
- Zeinalzadehtabrizi, H.; Hosseinpour, A.; Aydin, M.; Haliloglu, K. A modified genomic DNA extraction method from leaves of sunflower for PCR based analyzes. J. Biodivers. Environ. Sci. 2015, 7, 222–225. [Google Scholar]
- Heckman, K.L.; Pease, L.R. Gene splicing and mutagenesis by PCR-driven overlap extension. Nat. Protoc. 2007, 2, 924–932. [Google Scholar] [CrossRef]
- Berardini, T.Z.; Reiser, L.; Li, D.; Mezheritsky, Y.; Muller, R.; Strait, E.; Huala, E. The Arabidopsis Information Resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 2015, 53, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Badouin, H.; Gouzy, J.; Grassa, C.J.; Murat, F.; Staton, S.E.; Cottret, L.; Lelandais-Brière, C.; Owens, G.L.; Carrère, S.; Mayjonade, B.; et al. The sunflower genome provides insights into oil metabolism, flowering and Asterid evolution. Nature 2017, 546, 148–152. [Google Scholar] [CrossRef]
- Almagro-Armenteros, J.J.; Sønderby, C.K.; Sønderby, S.K.; Nielsen, H.; Winther, O. DeepLoc: Prediction of protein subcellular localization using deep learning. Bioinformatics 2017, 33, 3387–3395. [Google Scholar] [CrossRef]
- Sperschneider, J.; Catanzariti, A.-M.; DeBoer, K.; Petre, B.; Gardiner, D.M.; Singh, K.B.; Dodds, P.N.; Taylor, J.M. LOCALIZER: Subcellular localization prediction of both plant and effector proteins in the plant cell. Sci. Rep. 2017, 7, 44598. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.C.; Lee, T.Y.; Huang, H.D.; Huang, H.Y.; Pan, R.L. PlantPAN: Plant promoter analysis navigator, for identifying combinatorial cis-regulatory elements with distance constraint in plant gene groups. BMC Genom. 2008, 9, 561. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Gribskov, M. Combining evidence using p-values: Application to sequence homology searches. Bioinformatics 1998, 14, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
 160–640 ng); GFP, 6–His–TRX–GFP fusion protein (
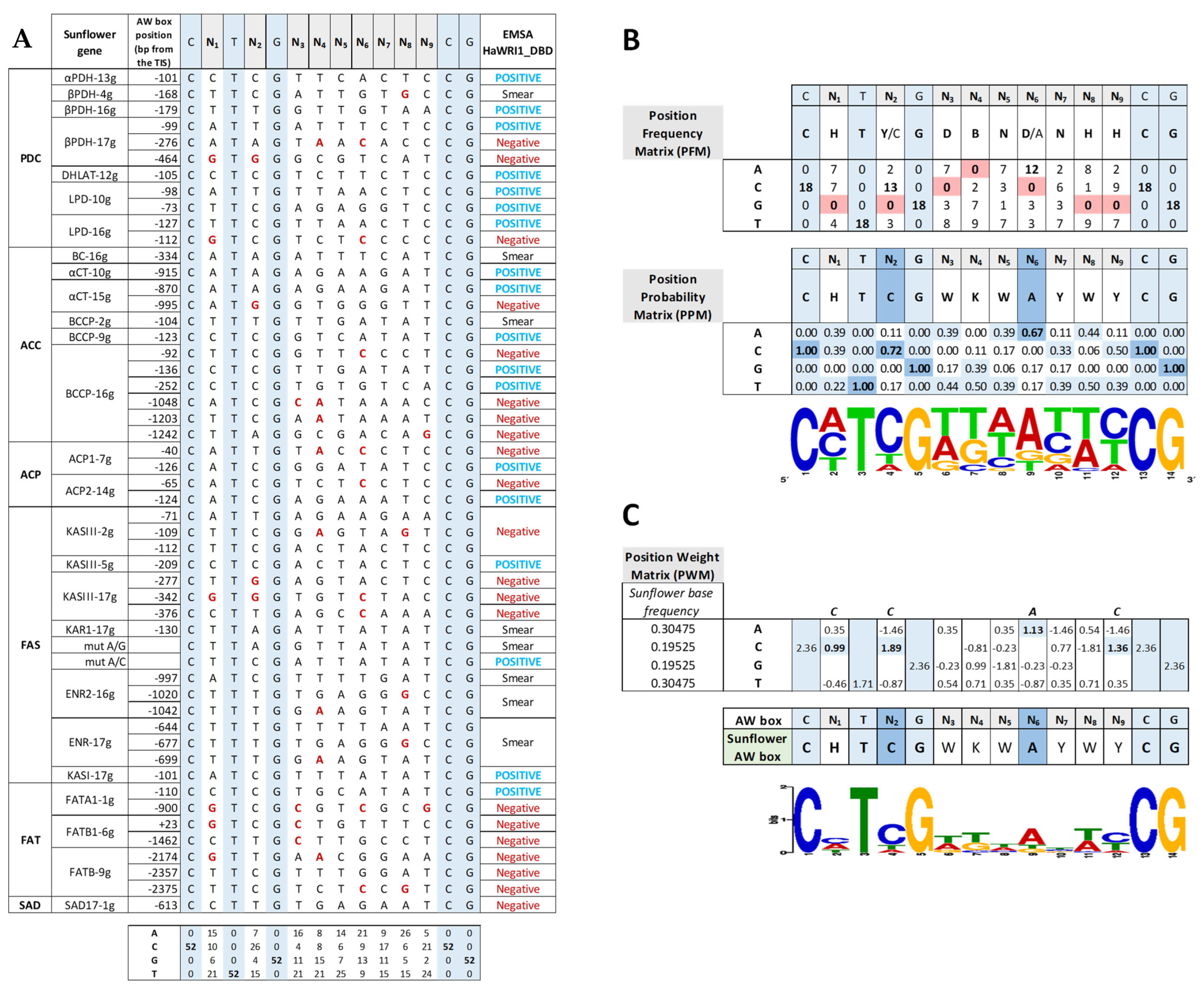
160–640 ng); GFP, 6–His–TRX–GFP fusion protein ( 160–640 ng); uDNA1, non–specific DNA1 (HacPGK2); uDNA2, non–specific DNA2 (HaCWI3). Overlapping DNA fragments (f1 to f4, 200–300 ng) tested in each gene in agarose EMSA are detailed on the right. The boxes indicate the AW box motifs, and the numbers indicate the distance from the ATG codon (bp). Binding is indicated as negative (red boxes) or positive (blue boxes), while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location. TIS, Translational Initiation Site (ATG).
160–640 ng); GFP, 6–His–TRX–GFP fusion protein ( 160–640 ng); uDNA1, non–specific DNA1 (HacPGK2); uDNA2, non–specific DNA2 (HaCWI3). Overlapping DNA fragments (f1 to f4, 200–300 ng) tested in each gene in agarose EMSA are detailed on the right. The boxes indicate the AW box motifs, and the numbers indicate the distance from the ATG codon (bp). Binding is indicated as negative (red boxes) or positive (blue boxes), while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location. TIS, Translational Initiation Site (ATG).
160–640 ng); uDNA1, non–specific DNA1 (HacPGK2); uDNA2, non–specific DNA2 (HaCWI3). Overlapping DNA fragments (f1 to f4, 200–300 ng) tested in each gene in agarose EMSA are detailed on the right. The boxes indicate the AW box motifs, and the numbers indicate the distance from the ATG codon (bp). Binding is indicated as negative (red boxes) or positive (blue boxes), while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location. TIS, Translational Initiation Site (ATG).
160–640 ng); GFP, 6–His–TRX–GFP fusion protein ( 160–640 ng); uDNA1, non–specific DNA1 (HacPGK2); uDNA2, non–specific DNA2 (HaCWI3). Overlapping DNA fragments (f1 to f4, 200–300 ng) tested in each gene in agarose EMSA are detailed on the right. The boxes indicate the AW box motifs, and the numbers indicate the distance from the ATG codon (bp). Binding is indicated as negative (red boxes) or positive (blue boxes), while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location. TIS, Translational Initiation Site (ATG).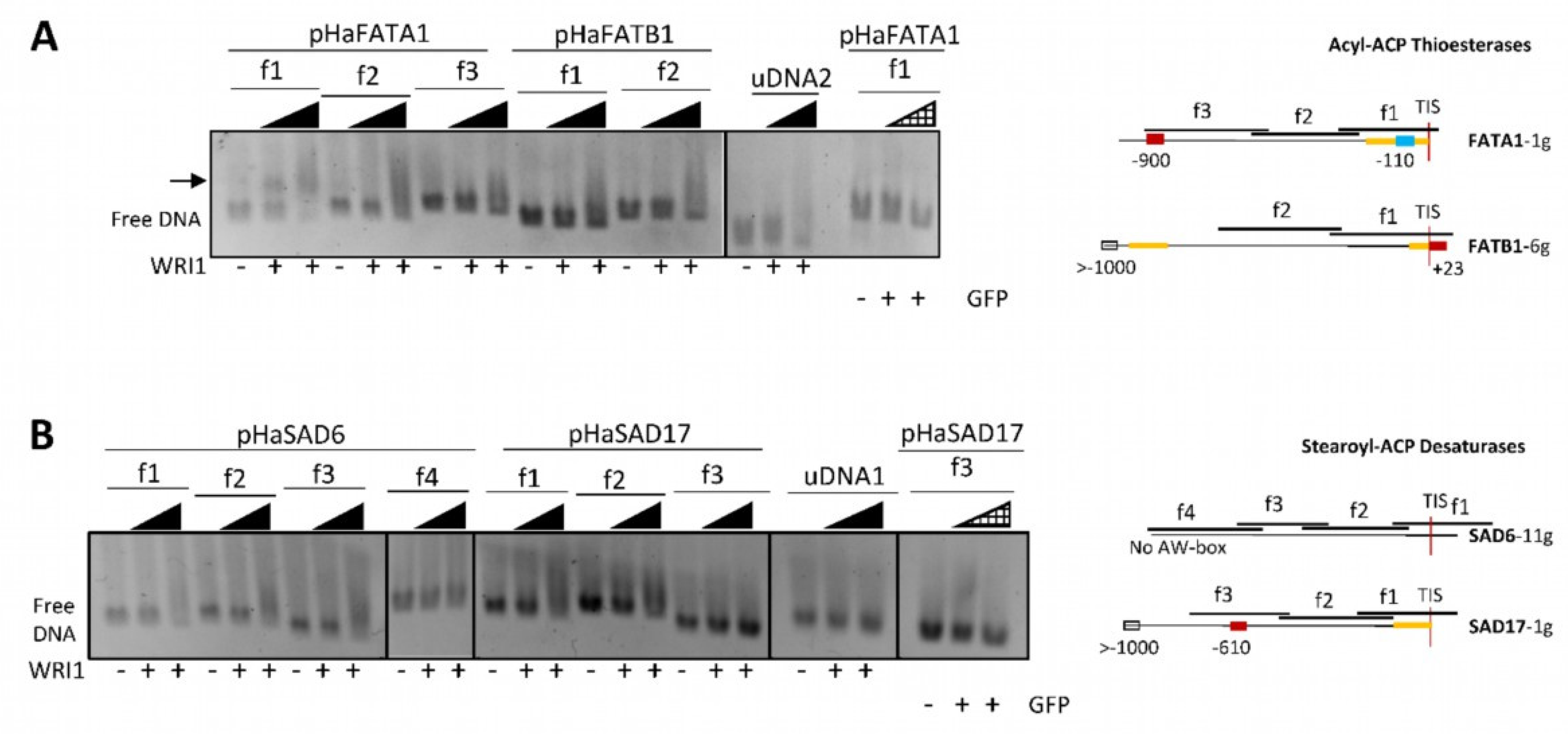
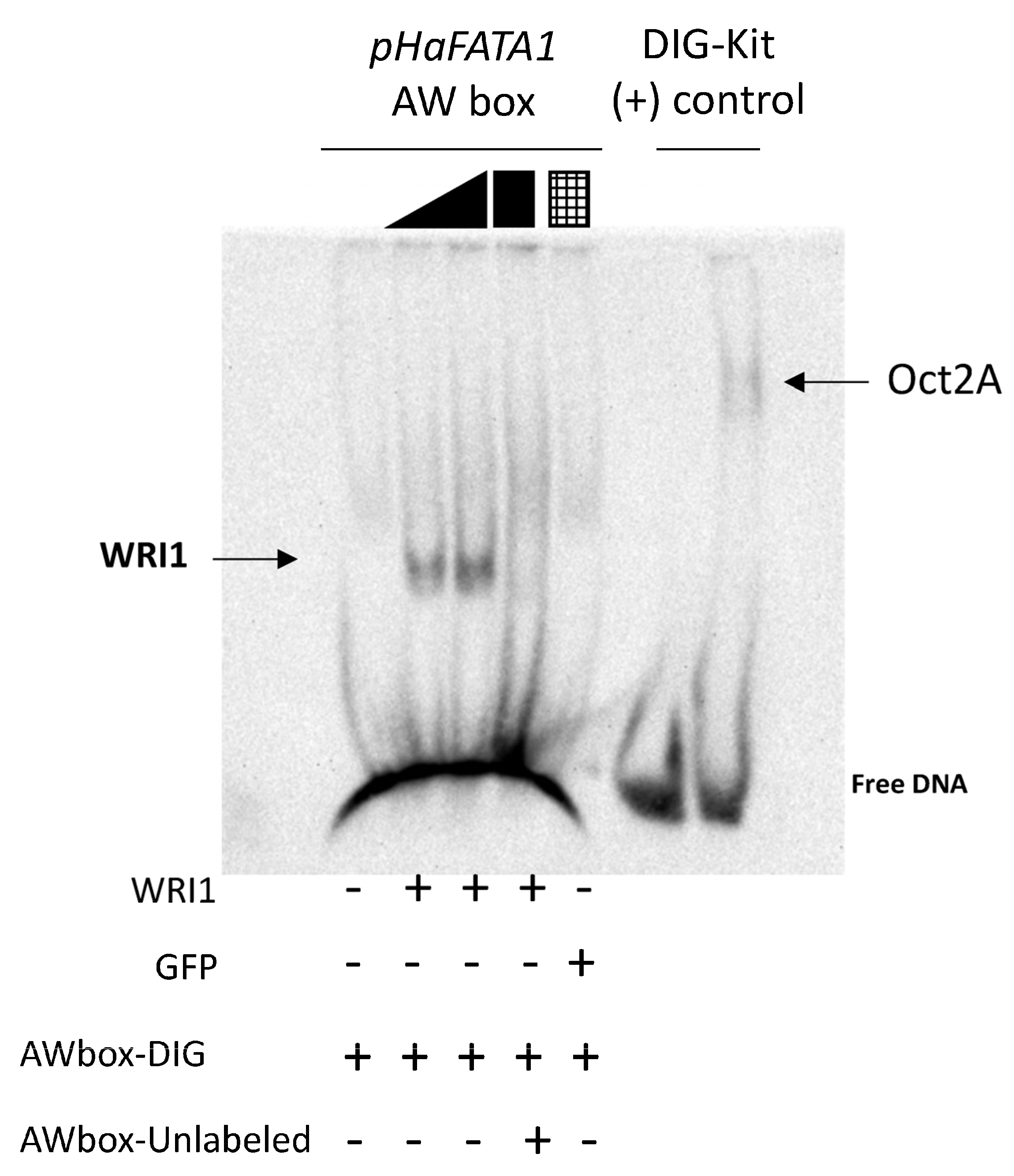
 cold competitor reaction) dsDNA (24 bp, 30 fmol) contains the 14 bp AW box motif. WRI1, 6–His–TRX–WRI1_DBD fusion protein (
cold competitor reaction) dsDNA (24 bp, 30 fmol) contains the 14 bp AW box motif. WRI1, 6–His–TRX–WRI1_DBD fusion protein ( 500–1000 ng); GFP, 6–His–TRX–GFP fusion protein (
500–1000 ng); GFP, 6–His–TRX–GFP fusion protein ( 1000 ng); Oct2A, Octamer–binding factor 2A (75 ng).
cold competitor reaction) dsDNA (24 bp, 30 fmol) contains the 14 bp AW box motif. WRI1, 6–His–TRX–WRI1_DBD fusion protein ( 500–1000 ng); GFP, 6–His–TRX–GFP fusion protein ( 1000 ng); Oct2A, Octamer–binding factor 2A (75 ng).
1000 ng); Oct2A, Octamer–binding factor 2A (75 ng).
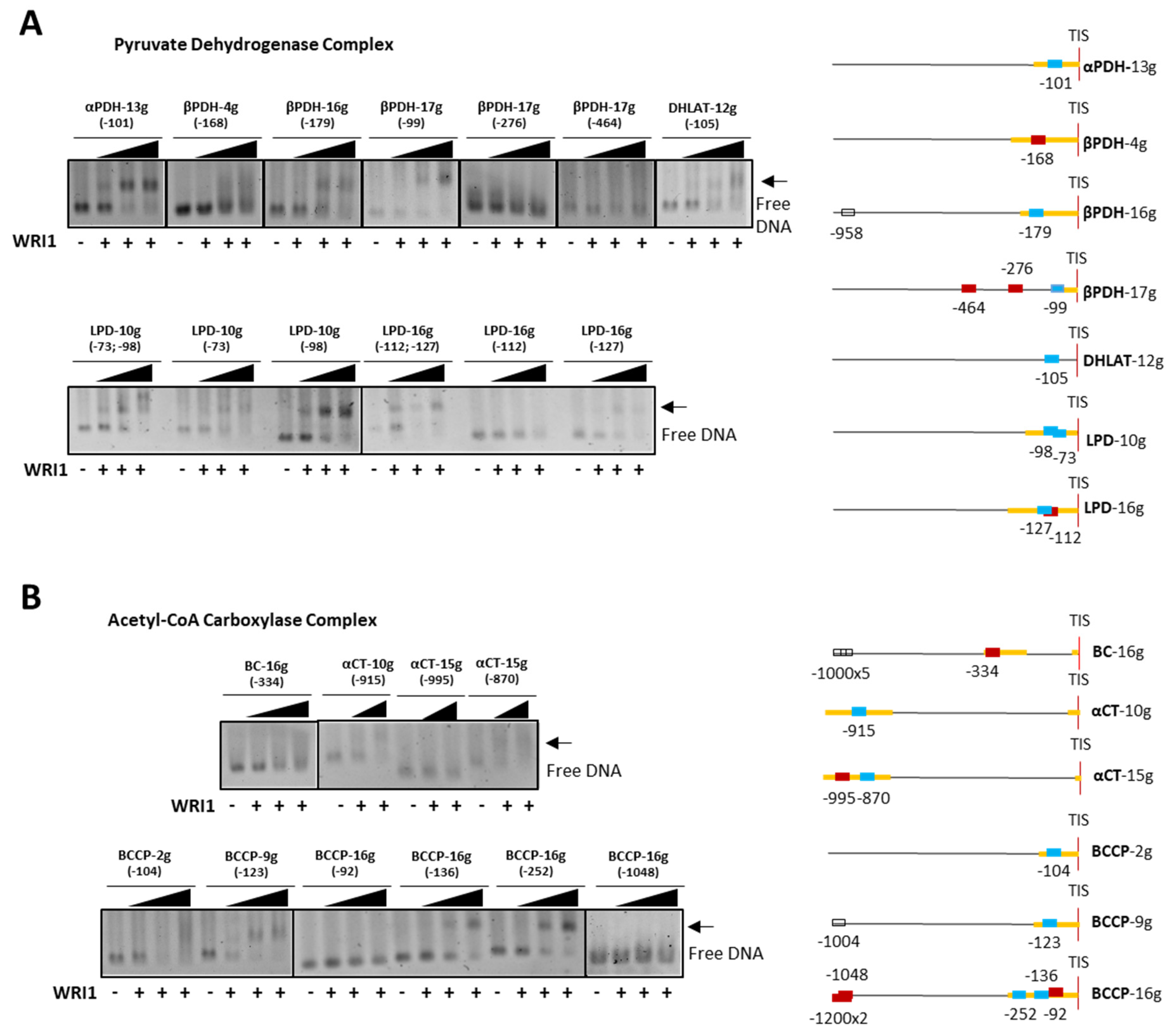
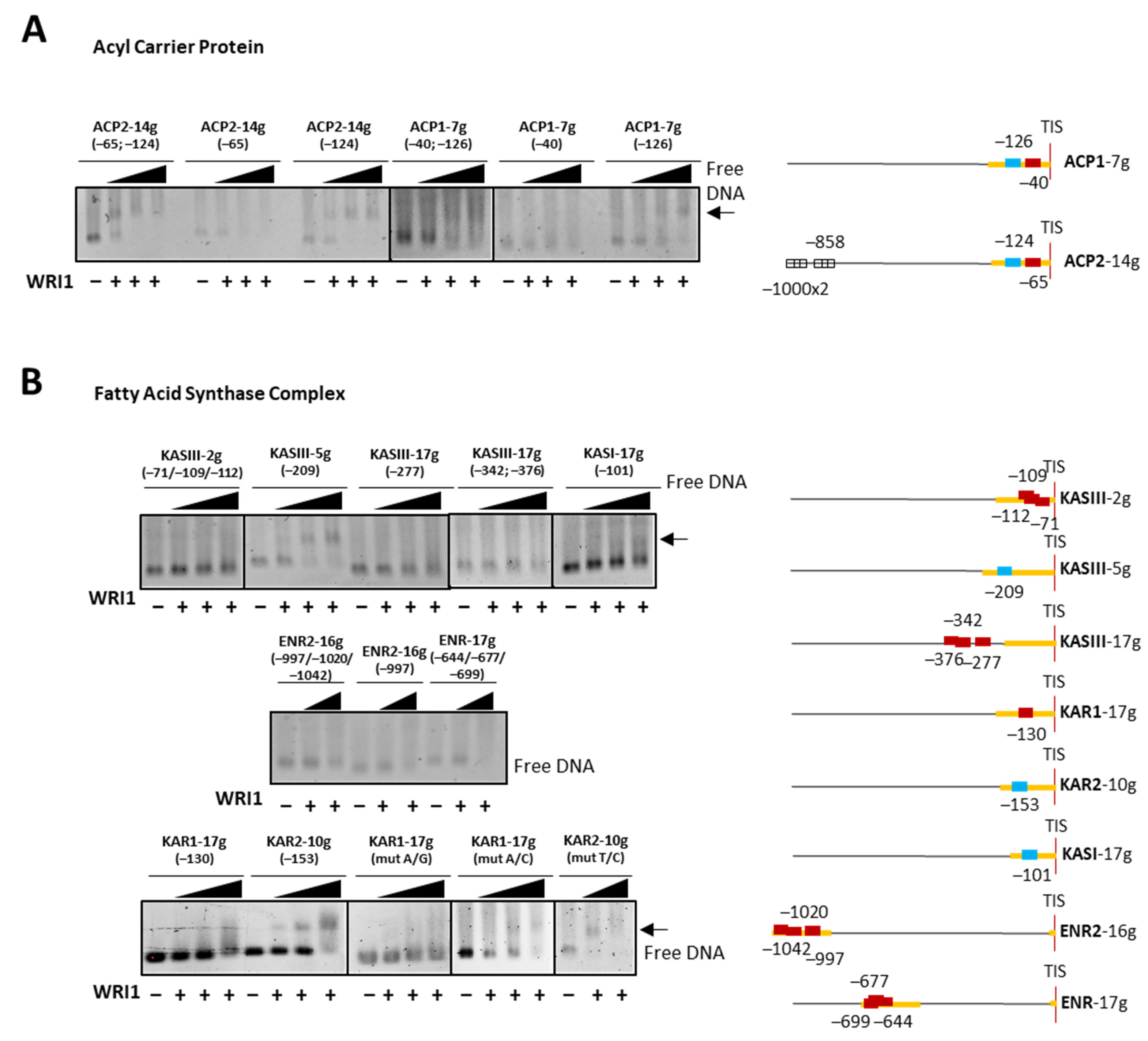
cold competitor reaction) dsDNA (24 bp, 30 fmol) contains the 14 bp AW box motif. WRI1, 6–His–TRX–WRI1_DBD fusion protein ( 500–1000 ng); GFP, 6–His–TRX–GFP fusion protein ( 1000 ng); Oct2A, Octamer–binding factor 2A (75 ng).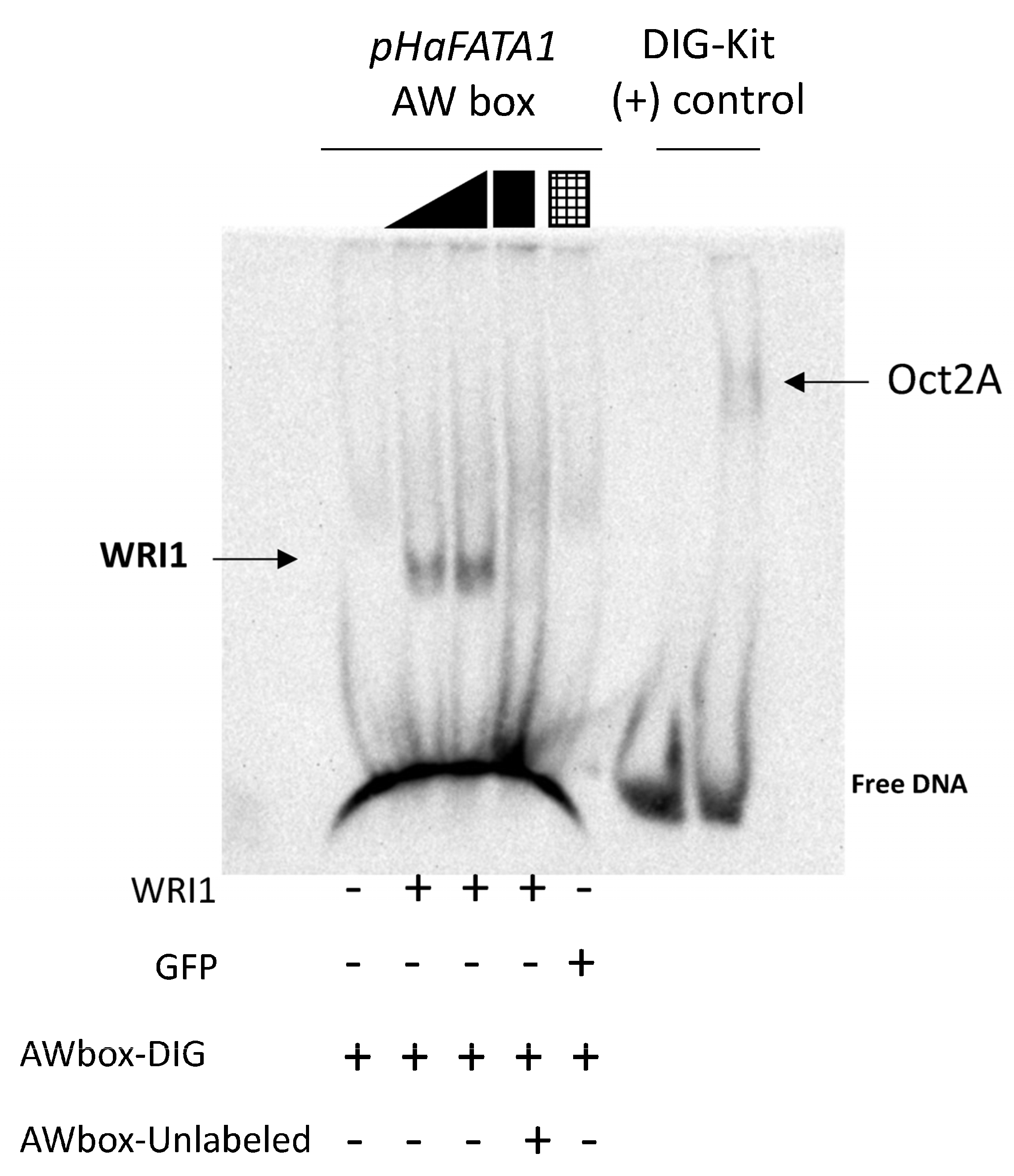 50–150–300 ng). The locations of the AW box motifs relative to the ATG codon in each DNA promoter region analyzed by EMSA (300 ng) are shown on the right. The boxes indicate AW box and the numbers indicate the location from the ATG (bp). Red boxes indicate negative binding and blue for positive binding, while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location: TIS, Translational Initiation Site (ATG).
50–150–300 ng). The locations of the AW box motifs relative to the ATG codon in each DNA promoter region analyzed by EMSA (300 ng) are shown on the right. The boxes indicate AW box and the numbers indicate the location from the ATG (bp). Red boxes indicate negative binding and blue for positive binding, while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location: TIS, Translational Initiation Site (ATG).
50–150–300 ng). The locations of the AW box motifs relative to the ATG codon in each DNA promoter region analyzed by EMSA (300 ng) are shown on the right. The boxes indicate AW box and the numbers indicate the location from the ATG (bp). Red boxes indicate negative binding and blue for positive binding, while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location: TIS, Translational Initiation Site (ATG).
50–150–300 ng). The locations of the AW box motifs relative to the ATG codon in each DNA promoter region analyzed by EMSA (300 ng) are shown on the right. The boxes indicate AW box and the numbers indicate the location from the ATG (bp). Red boxes indicate negative binding and blue for positive binding, while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location: TIS, Translational Initiation Site (ATG).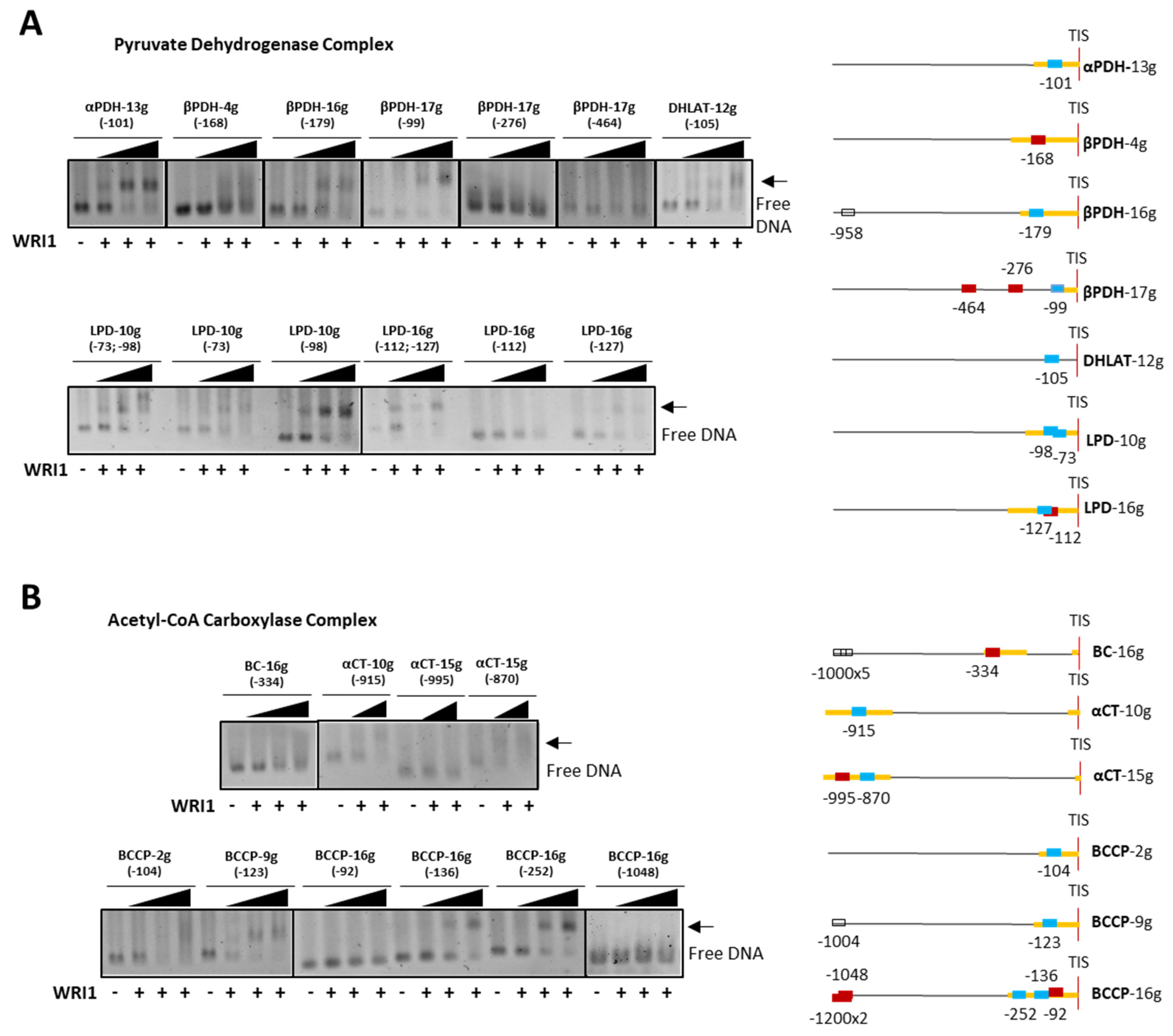 50–150–300 ng). The positions of the AW box motifs relative to ATG codon in each DNA promoter region analyzed by EMSA (300 ng) are shown on the right. The boxes indicate the AW box motifs, and the numbers indicate the positions relative to the ATG (bp). The binding data are indicated as red for negative and blue for positive, while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location: TIS, Translational Initiation Site (ATG).
50–150–300 ng). The positions of the AW box motifs relative to ATG codon in each DNA promoter region analyzed by EMSA (300 ng) are shown on the right. The boxes indicate the AW box motifs, and the numbers indicate the positions relative to the ATG (bp). The binding data are indicated as red for negative and blue for positive, while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location: TIS, Translational Initiation Site (ATG).
50–150–300 ng). The positions of the AW box motifs relative to ATG codon in each DNA promoter region analyzed by EMSA (300 ng) are shown on the right. The boxes indicate the AW box motifs, and the numbers indicate the positions relative to the ATG (bp). The binding data are indicated as red for negative and blue for positive, while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location: TIS, Translational Initiation Site (ATG).
50–150–300 ng). The positions of the AW box motifs relative to ATG codon in each DNA promoter region analyzed by EMSA (300 ng) are shown on the right. The boxes indicate the AW box motifs, and the numbers indicate the positions relative to the ATG (bp). The binding data are indicated as red for negative and blue for positive, while a colorless AW box means it was not tested. The 5′–UTR is in orange. The sunflower gene names are based on previous publications, and they are followed by the number indicating the chromosome location: TIS, Translational Initiation Site (ATG).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heliagene ID (v2020) | Sunflower Gene | Strand | AW Box Sequence (5′−3′) | AW Box Position (bp from the TIS) |
|---|---|---|---|---|
| HanXRQr2Chr01g0022981 | FATA1−1g | + | CCTCGTGCATATCG | −110/−97 |
| + | CGTCGCGTCGCGCG | −900/−887 | ||
| HanXRQr2Chr06g0263811 | FATB1−6g | + | CGTCGCTGTTTCCG | +23/+36 |
| − | CCTTGCTTGCCTCG | −1419/−1406 | ||
| HanXRQr2Chr01g0035021 | SAD17−1g | − | CCTTGTGAGAATCG | −610/−597 |
| − | CCTTGATATCATCG | −1159/−1146 | ||
| HanXRQr2Chr11g0503601 | SAD6−11g | + | CATAGGCAACTTACG * | −670/−656 |
| Plastidial Fatty Acid Synthesis Gene | Arabidopsis Gene | Sunflower Gene Heliagene ID-v2020 | Sunflower Gene Shorter Name | Number of AW Boxes | Subcelullar Localization | Gene Expression in SunFlower Seed Y/N-rpkm (Heliagene) | Gene Expression in SunFlower Seed (CAS-6 Line) | |
|---|---|---|---|---|---|---|---|---|
| PDC | α-PDH | At1g01090 | HanXRQr2Chr13g0619071 | α-PDH-13g | 1 | Plastid | Y | |
| β-PDH | At1g30120 | HanXRQr2Chr04g0172911 | β-PDH-4g | 1 | Plastid | Y-0.02 | ||
| HanXRQr2Chr16g0768891 | β-PDH-16g | 2 | Plastid | Y-55 | ||||
| HanXRQr2Chr17g0823581 | β-PDH-17g | 3 | Plastid | Y-35 | ||||
| LPD | At3g16950 | HanXRQr2Chr05g0217331 | 0 | Plastid | Y-2 | |||
| HanXRQr2Chr10g0440741 | LPD-10g | 2 | Plastid | Y-13 | ||||
| HanXRQr2Chr16g0749581 | LPD-16g | 2 | Plastid | Y-11 | ||||
| DHLAT | At3g25860 | HanXRQr2Chr04g0156631 | DHLAT-4g | 2 | Plastid | Y-35 | ||
| HanXRQr2Chr08g0324561 | DHLAT-8g | 1 | Plastid | Y-30 | ||||
| HanXRQr2Chr12g0535191 | DHLAT-12g | 1 | Plastid | Y | ||||
| ACC | α-CT | At2g38040 | HanXRQr2Chr10g0429681 | α-CT-10g | 4 | Plastid | Y-3 | |
| HanXRQr2Chr10g0429871 | 0 | Plastid | Y | |||||
| HanXRQr2Chr15g0671211 | α-CT-15g | 2 | Plastid | Y-5 | D.N.S. | |||
| β-CT | AtCg00500 | HanXRQr2CPg0836391 | 0 | Plastid | Y-0.8 | D.N.S. | ||
| BC | At5g35360 | HanXRQr2Chr16g0770591 | BC-16g | 6 | Plastid | Y-35 | D.N.S. | |
| HanXRQr2Chr17g0824731 | 0 | Plastid | Y-7 | |||||
| BCCP | At5g15530 | HanXRQr2Chr02g0061841 | BCCP-2g | 1 | Plastid | Y-18 | D.N.S. | |
| HanXRQr2Chr05g0231761 | 0 | Plastid | Y-3 | |||||
| HanXRQr2Chr09g0408331 | BCCP-9g | 2 | Plastid | Y-85 | ||||
| HanXRQr2Chr16g0729021 | BCCP-16g | 6 | Plastid | Y-23 | D.N.S. | |||
| MCMT | At2g30200 | HanXRQr2Chr01g0018151 | 0 | Plastid | Y-51 | |||
| HanXRQr2Chr01g0018201 | 0 | Plastid | Y-0.05 | |||||
| HACPS | At3g11470 | HanXRQr2Chr04g0161011 | HACPS-4g | 0 | Mitochondrion | Y-1 | ||
| HanXRQr2Chr07g0300321 | HACPS-7g | 1 | Mitochondrion | Y-1.6 | ||||
| ACP | At4g25050 | HanXRQr2Chr07g0290651 (ACP1) | ACP-7g | 2 | Plastid | Y-0.11 | [40] | |
| HanXRQr2Chr08g0343921 (ACP3) | 0 | Plastid | Y-370 | [40] | ||||
| HanXRQr2Chr09g0412861 | 0 | Plastid | Y | |||||
| HanXRQr2Chr14g0647771 (ACP2) | ACP-14g | 6 | Plastid | Y-1.5 | [40] | |||
| HanXRQr2Chr17g0819021 | 0 | Mitochondrion | Y-8 | |||||
| FAS | KASI | At5g46290 | HanXRQr2Chr01g0024181 | KASI-1g | 1 | Plastid | N.D. | |
| HanXRQr2Chr01g0027641 | 0 | Plastid | N.D. | |||||
| HanXRQr2Chr04g0179841 | 0 | Plastid | Y-38 | |||||
| HanXRQr2Chr13g0603901 | 0 | Plastid | Y-0.1 | |||||
| HanXRQr2Chr17g0822771 | KASI-17g | 1 | Plastid | Y-90 | ||||
| HanXRQr2Chr17g0780601 | 0 | Peroxisome | N.D. | |||||
| KASII | At1g74960 | HanXRQr2Chr06g0259761 | KASII-6g | 2 | Peroxisome | N.D. | ||
| HanXRQr2Chr09g0361681 | 0 | Plastid | Y-3 | |||||
| HanXRQr2Chr15g0722271 | KASII-15g | 2 | Plastid | Y-1 | ||||
| KASIII | At1g62640 | HanXRQr2Chr02g0073211 (KASIII) | KASIII-2g | 3 | Plastid | Y-50 | [46] | |
| HanXRQrChr05g0148701# | KASIII-5g | 1 | Plastid | Y-55 | ||||
| HanXRQr2Chr17g0801431 | KASIII-17g | 3 | Plastid | Y-5 | ||||
| KAR | At1g24360 | HanXRQr2Chr10g0464751 (KAR2) | 0 | Plastid | Y | [45] | ||
| HanXRQr2Chr17g0781961 (KAR1) | KAR-17g | 1 | Plastid | Y-100 | [45] | |||
| HAD | At2g22230 | HanXRQr2Chr03g0102091 (HAD2) | 0 | Plastid | Y-25 | [44] | ||
| HanXRQr2Chr10g0443511 (HAD1) | 0 | Plastid | Y-0.03 | [44] | ||||
| ENR | At2g05990 | HanXRQr2Chr02g0082991 (ENR1) | 0 | Plastid | Y-7 | [43] | ||
| HanXRQr2Chr16g0768471 (ENR2) | ENR-16g | 3 | Plastid | Y-20 | [43] | |||
| HanXRQr2Chr17g0822551 | ENR-17g | 3 | Plastid | Y-28 | ||||
| LS | At5g08415 | HanXRQr2Chr05g0201611 (LIP1p1) | LS-5g | 1 | Plastid | Y-1.5 | [38] | |
| HanXRQr2Chr12g0526011 (LIP1p2) | LS-12g | 1 | Plastid | Y-0.02 | [38] | |||
| LT | At4g31050 | HanXRQr2Chr05g0205061 (LIP2p) | LT-5g | 1 | Plastid | Y-4 | ||
| HanXRQr2Chr09g0406031 | 0 | Plastid | N.D. | |||||
| FAT | FATA | At3g25110 | HanXRQr2Chr01g0022981 (FATA1) | FATA-1g | 2 | Plastid | Y-73 | [41] |
| FATB | At1g08510 | HanXRQr2Chr05g0207531 | 0 | Plastid | Y-1 | |||
| HanXRQr2Chr06g0263811 (FATB1) | FATB-6g | 2 | Plastid | Y-0.02 | [42] | |||
| HanXRQr2Chr09g0365831 | FATB-9g | 3 | Plastid | Y-13 | D.N.S. | |||
| SAD | At2g43710 | HanXRQr2Chr01g0035021 (SAD17) | SAD-1g | 2 | Plastid | Y-310 | ||
| HanXRQr2Chr11g0503601 (SAD6) | 0 | Plastid | Y-120 |
| Sunflower Gene | Strand | AW Box Sequence (5′-3′) | AW Box Position (bp from the TIS) | AW Box in the 5′-UTR | Sunflower Gene | Strand | AW Box Sequence (5′-3′) | AW Box Position (bp from the TIS) | AW Box in the 5′-UTR |
|---|---|---|---|---|---|---|---|---|---|
| αPDH-13g | + | CCTCGTTCACTCCG | −101/−88 | Y | ACP1-7g | + | CATTGTACCCTCCG | −40/−27 | Y |
| βPDH-4g | − | CTTCGATTGTGCCG | −168/−155 | Y | − | CATCGGGATATCCG | −126/−113 | Y | |
| βPDH-16g | − | CTTTGGTTGTAACG | −179/−166 | Y | ACP2-14g | + | CATCGTCTCCTCCG | −65/−52 | Y |
| − | CATTGCGTTTTACG | −958/−944 | − | CATCGAGAAATCCG | −124/−110 | Y | |||
| βPDH-17g | + | CATTGATTTCTCCG | −99/−86 | Y | + | CGTCGTTGCCGACG | −859/−846 | ||
| − | CATAGTAACACCCG | −276/−263 | − | CGTCGGCAACGACG | −859/−846 | ||||
| − | CGTGGGCGTCATCG | −464/−451 | − | CCTGGGTGATAACG | −882/−869 | ||||
| LPD-10g | − | CTTCGAGAGGTCCG | −73/−60 | Y | − | CGTCGTTTCCATCG | −1195/−1182 | ||
| + | CATTGTTAACTCCG | −98/−85 | Y | KASI-1g | + | CGTTGTTATTTACG | −991/−978 | ||
| LPD-16g | + | CGTCGTCTCCCCCG | −112/−99 | Y | KASI-17g | + | CATCGTTTATATCG | −101/−87 | Y |
| + | CTTCGTTAACTCCG | −127/−114 | Y | KASII-15g | + | CCTGGAAACAAACG | −1240/−1226 | ||
| DHLAT-4g | − | CGTTGGTTAGCGCG | −641/−628 | + | CTTTGGGTTTTTCG | −1290/−1277 | |||
| − | CGTCGTATCTACCG | −675/−622 | KASIII-2g | − | CATTGAGAAGAACG | −71/−58 | Y | ||
| DHLAT-8g | − | CTTAGGCCTTAACG | −703/−690 | − | CTTCGGAGTAGTCG | −109/−96 | Y | ||
| DHLAT-12g | + | CCTCGTCTTCTCCG | −105/−92 | Y | + | CTTCGACTACTCCG | −112/−99 | Y | |
| αCT-10g | − | CATAGAGAAGATCG | −915/−902 | Y | KASIII-5g | + | CCTCGACTACTCCG | −209/−196 | Y |
| + | CATAGAACTCATCG | −1095/−1082 | KASIII-17g | + | CTTGGAGTACTCCG | −277/−264 | |||
| − | CGTCGAAAATCCCG | −1383/−1370 | + | CGTGGTGTCTACCG | −342/−329 | ||||
| − | CTTGGAATTTTTCG | −1406/−1393 | + | CCTTGAGCCAAACG | −376/−363 | ||||
| αCT-15g | − | CATAGAGAAGATCG | −870/−857 | Y | KAR1-17g | + | CTTAGATTATATCG | −130/−117 | Y |
| − | CATGGGTGGGTTCG | −995/−982 | Y | mut A/G | CTTAGGTTATATCG | ||||
| BC-16g | + | CATAGATTTCATCG | −334/−321 | Y | mut A/C | CTTCGATTATATCG | |||
| + | CGTTGTCTAAATCG | −1042/−1029 | KAR2-10g | + | CTTTAGGTTATATCG # | −153/−139 | |||
| − | CGTCGAAAATCCCG | −1217/−1204 | mut T/C | CTTCAGGTTATATCG | |||||
| − | CTTGGAATTTTTCG | −1240/−1227 | ENR2-16g | + | CATCGTTTTGATCG | −997/−984 | Y | ||
| − | CCTTGAATTTTTCG | −1262/−1249 | − | CTTTGTGAGGGCCG | −1020/−1007 | Y | |||
| − | CTTGGAATTTTTCG | −1285/−1272 | − | CTTTGGAAGTATCG | −1042/−1029 | Y | |||
| BCCP-2g | + | CTTTGTTGATATCG | −104/−91 | Y | ENR-17g | + | CTTTGTTTTAATCG | −644/−631 | Y |
| BCCP-9g | + | CCTCGGTCATATCG | −123/−110 | Y | − | CTTTGTGAGGGCCG | −677/−664 | Y | |
| − | CTTTGTTACAGGCG | −1004/−991 | − | CTTTGGAAGTATCG | −699/−686 | Y | |||
| BCCP-16g | + | CTTCGGTTCCCTCG | −92/−79 | Y | FATA1-1g | + | CCTCGTGCATATCG | −110/−97 | Y |
| + | CCTCGTTGATATCG | −136/−123 | Y | + | CGTCGCGTCGCGCG | −900/−887 | |||
| − | CCTCGTGTGTCACG | −252/−239 | Y | FATB1-6g | − | CCTTGCTTGCCTCG | −1419/−1406 | ||
| + | CATCGCATAAACCG | −1048/−1035 | FATB-9g | − | CGTTGAACGGAACG | −2174/−2161 | |||
| + | CTTCGAATAAATCG | −1203/−1190 | − | CTTCGTTTGGATCG | −2357/−2344 | ||||
| − | CTTAGGCGACAGCG | −1242/−1229 | − | CTTCGTCTCCGTCG | −2375/−2362 | ||||
| (LS) LIP1p1-5g | − | CCTTGTCAAGTACG | −867/−854 | SAD17-1g | − | CCTTGTGAGAATCG | −610/−597 | ||
| (LS) LIP1p2-12g | − | CATGGCTGTCTACG | −570/−557 | − | CCTTGATATCATCG | −1159/−1146 | |||
| (LT) LIP2p-5g | + | CTTCGGTCACTCCG | −42/−23 | Y |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, R.; González-Thuillier, I.; Venegas-Calerón, M.; Garcés, R.; Salas, J.J.; Martínez-Force, E. The Sunflower WRINKLED1 Transcription Factor Regulates Fatty Acid Biosynthesis Genes through an AW Box Binding Sequence with a Particular Base Bias. Plants 2022, 11, 972. https://doi.org/10.3390/plants11070972
Sánchez R, González-Thuillier I, Venegas-Calerón M, Garcés R, Salas JJ, Martínez-Force E. The Sunflower WRINKLED1 Transcription Factor Regulates Fatty Acid Biosynthesis Genes through an AW Box Binding Sequence with a Particular Base Bias. Plants. 2022; 11(7):972. https://doi.org/10.3390/plants11070972
Chicago/Turabian StyleSánchez, Rosario, Irene González-Thuillier, Mónica Venegas-Calerón, Rafael Garcés, Joaquín J. Salas, and Enrique Martínez-Force. 2022. "The Sunflower WRINKLED1 Transcription Factor Regulates Fatty Acid Biosynthesis Genes through an AW Box Binding Sequence with a Particular Base Bias" Plants 11, no. 7: 972. https://doi.org/10.3390/plants11070972
APA StyleSánchez, R., González-Thuillier, I., Venegas-Calerón, M., Garcés, R., Salas, J. J., & Martínez-Force, E. (2022). The Sunflower WRINKLED1 Transcription Factor Regulates Fatty Acid Biosynthesis Genes through an AW Box Binding Sequence with a Particular Base Bias. Plants, 11(7), 972. https://doi.org/10.3390/plants11070972