
Critical Determinants in ER-Golgi Trafficking of Enzymes Involved in Glycosylation



Abstract
1. Introduction
2. Main Components of the Secretory Pathway
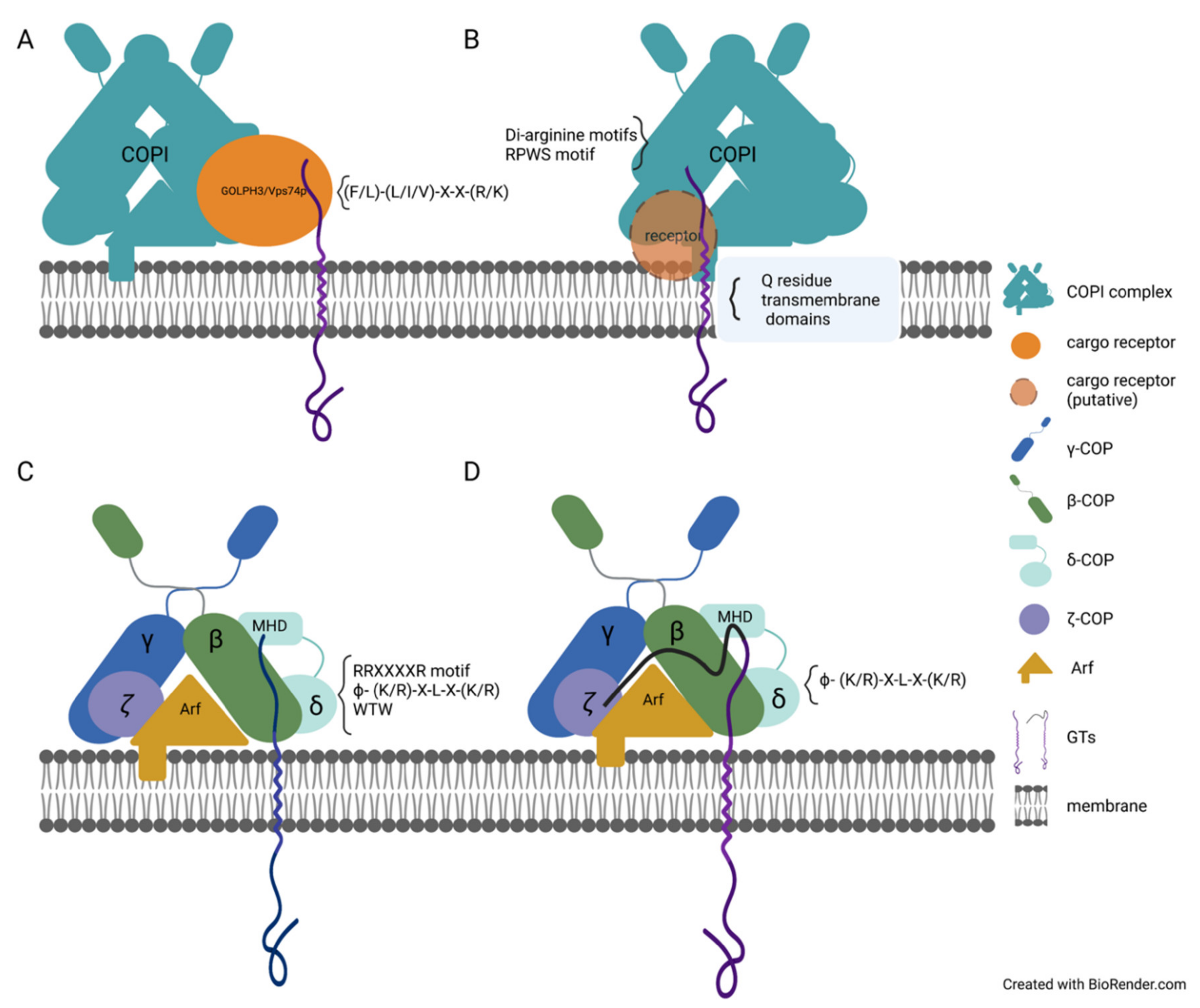
3. Specific Sequence Motifs Involved in GTs and Glycosidases Sorting and Trafficking
4. Other Protein Domains Essential for the Trafficking of Enzymes Involved in Glycosylation
5. Recycling of Glycosyltransferase and Glycosidases Involved in Glycosylation
6. Protein–Protein Interactions Contribute to GT Trafficking
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for Glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef] [PubMed]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S.G. Glycosyltransferases: Structures, functions, and mechanisms. Annu. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [PubMed]
- Zabotina, O.A.; Zhang, N.; Weerts, R. Polysaccharide Biosynthesis: Glycosyltransferases and Their Complexes. [published correction appears in Front. Plant Sci. 2021, 12, 720709]. Front. Plant Sci. 2021, 12, 625307. [Google Scholar] [CrossRef]
- Yuan, Y.; Barrett, D.; Zhang, Y.; Kahne, D.; Sliz, P.; Walker, S. Crystal structure of a peptidoglycan glycosyltransferase suggests a model for processive glycan chain synthesis. Proc. Natl. Acad. Sci. USA 2007, 104, 5348–5353. [Google Scholar] [CrossRef]
- Nagashima, Y.; von Schaewen, A.; Koiwa, H. Function of N-glycosylation in plants. Plant Sci. 2018, 274, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Cummings, R.D.; Esko, J.D.; Stanley, P.; Hart, G.W.; Aebi, M.; Darvill, A.G.; Kinoshita, T.; Packer, N.H.; Prestegard, H.H. (Eds.) Essentials of Glycobiology, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2015. [Google Scholar]
- Aebi, M. N-linked protein glycosylation in the ER. Biochim. Biophys. Acta 2013, 1833, 2430–2437. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Zhu, H.; Chen, X.; Tian, M.; Wei, Y.; Gong, Y.; Jiang, J. N-acetylglucosaminyltransferase I promotes glioma cell proliferation and migration through increasing the stability of the glucose transporter GLUT1. FEBS Lett. 2020, 594, 358–366. [Google Scholar] [CrossRef]
- Vanier, G.; Lucas, P.L.; Loutelier-Bourhis, C.; Vanier, J.; Plasson, C.; Walet-Balieu, M.-L.; Tchi-Song, P.C.; Remy-Jouet, I.; Richard, V.; Bernard, S.; et al. Heterologous expression of the N-acetylglucosaminyltransferase I dictates a reinvestigation of the N-glycosylation pathway in Chlamydomonas reinhardtii. Sci. Rep. 2017, 7, 10156. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Ko, K.S.; Vu, B.N.; Lee, Y.E.; Yoon, S.H.; Pham, T.T.; Kim, J.-T.; Lim, J.-M.; Kang, Y.J.; Hong, J.C.; et al. N-acetylglucosaminyltransferase II Is Involved in Plant Growth and Development under Stress Conditions. Front. Plant Sci. 2021, 12, 761064. [Google Scholar] [CrossRef]
- Kadirvelraj, R.; Yang, J.-Y.; Sanders, J.H.; Liu, L.; Ramiah, A.; Prabhakar, P.K.; Boons, G.-J.; Wood, Z.A.; Moremen, K.W. Human N-acetylglucosaminyltransferase II substrate recognition uses a modular architecture that includes a convergent exosite. Proc. Natl. Acad. Sci. USA 2018, 115, 4637–4642. [Google Scholar] [CrossRef]
- Wong, L.-J.; Sheu, K.-F.R.; Lee, S.-L.; Frey, P.A. Galactose-1-phosphate uridylyltransferase: Isolation and properties of a uridylyl-enzyme intermediate. Biochemistry 1977, 16, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Garnham, R.; Scott, E.; Livermore, K.E.; Munkley, J. ST6GAL1: A key player in cancer (Review). Oncol. Lett. 2019, 18, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Khoder-Agha, F.; Harrus, D.; Brysbaert, G.; Lensink, M.F.; Harduin-Lepers, A.; Glumoff, T.; Kellokumpu, S. Assembly of B4GALT1/ST6GAL1 heteromers in the Golgi membranes involves lateral interactions via highly charged surface domains. J. Biol. Chem. 2019, 294, 14383–14393. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Li, Z.; Li, Y.; Liu, G.; Sun, S.; Gu, J.; Kameyama, A.; Li, W.; Dong, W. Loss of core fucosylation in both ST6GAL1 and its substrate enhances glycoprotein sialylation in mice. Biochem. J. 2020, 477, 1179–1201. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Wang, H.; Liu, J.; Qin, H.; Liu, S.; Yan, Q. The sialyltransferase ST3Gal3 facilitates the receptivity of the uterine endometrium in vitro and in vivo. FEBS Lett. 2018, 592, 3696–3707. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, X.; Chen, M.; Zheng, S.; Li, J.; Lin, S.; Wang, X. ST3Gal3 confers paclitaxel-mediated chemoresistance in ovarian cancer cells by attenuating caspase-8/3 signaling. Mol. Med. Rep. 2019, 20, 4499–4506. [Google Scholar] [CrossRef] [PubMed]
- Khoder-Agha, F.; Sosicka, P.; Escriva Conde, M.; Hassinen, A.; Glumoff, T.; Olczak, M.; Kellokumpu, S. N-acetylglucosaminyltransferases and nucleotide sugar transporters form multi-enzyme-multi-transporter assemblies in golgi membranes in vivo. Cell. Mol. Life Sci. 2019, 76, 1821–1832. [Google Scholar] [CrossRef]
- Kellokumpu, S.; Hassinen, A.; Glumoff, T. Glycosyltransferase complexes in eukaryotes: Long-known, prevalent but still unrecognized. Cell. Mol. Life Sci. 2016, 73, 305–325. [Google Scholar] [CrossRef]
- van Tol, W.; Wessels, H.; Lefeber, D.J. O-glycosylation disorders pave the road for understanding the complex human O-glycosylation machinery. Curr. Opin. Struct. Biol. 2019, 56, 107–118. [Google Scholar] [CrossRef]
- Van den Steen, P.; Rudd, P.M.; Dwek, R.A.; Opdenakker, G. Concepts and Principles of O-Linked Glycosylation. Crit. Rev. Biochem. Mol. Biol. 1998, 33, 151–208. [Google Scholar] [CrossRef]
- Joshi, H.J.; Narimatsu, Y.; Schjoldager, K.T.; Tytgat, H.; Aebi, M.; Clausen, H.; Halim, A. SnapShot: O-Glycosylation Pathways across Kingdoms. Cell 2018, 172, 632. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Song, A.; Ao, M.; Xu, Y.; Zhang, H. Large-scale site-specific mapping of the O-GalNAc glycoproteome. Nat. Protoc. 2020, 15, 2589–2610. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.; Seifert, G.; Doblin, M.S.; Johnson, K.L.; Ruprecht, C.; Pfrengle, F.; Bacic, A.; Estevez, J.M. Cracking the “Sugar Code”: A Snapshot of N- and O-Glycosylation Pathways and Functions in Plants Cells. Front. Plant Sci. 2021, 12, 640919. [Google Scholar] [CrossRef] [PubMed]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. Arabidopsis β-COP Function Affects Golgi Structure, Plant Growth and Tolerance to Salt. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Polko, J.K.; Kieber, J.J. The Regulation of Cellulose Biosynthesis in Plants. Plant Cell 2019, 31, 282–296. [Google Scholar] [CrossRef]
- Kumar, M.; Turner, S. Plant cellulose synthesis: CESA proteins crossing kingdoms. Phytochemistry 2015, 112, 91–99. [Google Scholar] [CrossRef]
- Kumar, M.; Mishra, L.; Carr, P.; Pilling, M.; Gardner, P.; Mansfield, S.D.; Turner, S. Exploiting Cellulose Synthase (CESA) Class Specificity to Probe Cellulose Microfibril Biosynthesis. Plant Physiol. 2018, 177, 151–167. [Google Scholar] [CrossRef]
- Speicher, T.L.; Li, P.Z.; Wallace, I.S. Phosphoregulation of the Plant Cellulose Synthase Complex and Cellulose Synthase-Like Proteins. Plants 2018, 7, 52. [Google Scholar] [CrossRef]
- Zeng, W.; Lampugnani, E.R.; Picard, K.; Song, L.; Wu, A.; Farion, I.M.; Zhao, J.; Ford, K.; Doblin, M.S.; Bacic, A. Asparagus IRX9, IRX10, and IRX14A Are Components of an Active Xylan Backbone Synthase Complex that Forms in the Golgi Apparatus. Plant Physiol. 2016, 171, 93–109. [Google Scholar] [CrossRef]
- Wu, A.-M.; Hörnblad, E.; Voxeur, A.; Gerber, L.; Rihouey, C.; Lerouge, P.; Marchant, A. Analysis of the Arabidopsis IRX9/IRX9-L and IRX14/IRX14-L Pairs of Glycosyltransferase Genes Reveals Critical Contributions to Biosynthesis of the Hemicellulose Glucuronoxylan. Plant Physiol. 2010, 153, 542–554. [Google Scholar] [CrossRef]
- Urbanowicz, B.R.; Peña, M.J.; Moniz, H.A.; Moremen, K.W.; York, W.S. Two Arabidopsis proteins synthesize acetylated xylan in vitro. Plant J. 2014, 80, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Culbertson, A.T.; Ehrlich, J.J.; Choe, J.Y.; Honzatko, R.B.; Zabotina, O.A. Structure of xyloglucan xylosyltransferase 1 reveals simple steric rules that define biological patterns of xyloglucan polymers. Proc. Natl. Acad. Sci. USA 2018, 115, 6064–6069. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.H.; Pogorelko, G.; Zabotina, O.A. Xyloglucan xylosyltransferases XXT1, XXT2, and XXT5 and the glucan synthase CSLC4 form Golgi-localized multiprotein complexes. Plant Physiol. 2012, 159, 1355–1366. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Atmodjo, M.A.; Sakuragi, Y.; Zhu, X.; Burrell, A.J.; Mohanty, S.S.; Atwood, J.A.; Orlando, R.; Scheller, H.; Mohnen, D. Galacturonosyltransferase (GAUT)1 and GAUT7 are the core of a plant cell wall pectin biosynthetic homogalacturonan:galacturonosyltransferase complex. Proc. Natl. Acad. Sci. USA 2011, 108, 20225–20230. [Google Scholar] [CrossRef]
- Wang, J.; Chen, J.; Enns, C.A.; Mayinger, P. The First Transmembrane Domain of Lipid Phosphatase SAC1 Promotes Golgi Localization. PLoS ONE 2013, 8, e71112. [Google Scholar] [CrossRef]
- Béthune, J.; Wieland, F.T. Assembly of COPI and COPII Vesicular Coat Proteins on Membranes. Annu. Rev. Biophys. 2018, 47, 63–83. [Google Scholar] [CrossRef]
- Arakel, E.C.; Schwappach, B. Formation of COPI-coated vesicles at a glance. [published correction appears in J. Cell Sci. 2018, 131]. 37073 Göttingen, Germany. J. Cell Sci. 2018, 131, jcs209890. [Google Scholar] [CrossRef]
- Luo, P.M.; Boyce, M. Directing Traffic: Regulation of COPI Transport by Post-translational Modifications. Front. Cell Dev. Biol. 2019, 7, 190. [Google Scholar] [CrossRef]
- Sager, G.; Szul, T.; Lee, E.; Kawai, R.; Presley, J.F.; Sztul, E. Modeling the dynamic behaviors of the COPI vesicle formation regulators, the small GTPase Arf1 and its activating Sec7 guanine nucleotide exchange factor GBF1 on Golgi membranes. Mol. Biol. Cell 2021, 32, 446–459. [Google Scholar] [CrossRef]
- Bui, Q.T.; Golinelli-Cohen, M.P.; Jackson, C.L. Large Arf1 guanine nucleotide exchange factors: Evolution, domain structure, and roles in membrane trafficking and human disease. Mol. Genet. Genom. 2009, 282, 329–350. [Google Scholar] [CrossRef]
- Beck, R.; Adolf, F.; Weimer, C.; Bruegger, B.; Wieland, F.T. ArfGAP1 activity and COPI vesicle biogenesis. Traffic 2009, 10, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Shiba, Y.; Luo, R.; Hinshaw, J.E.; Szul, T.; Hayashi, R.; Sztul, E.; Nagashima, K.; Baxa, U.; Randazzo, P.A. ArfGAP1 promotes COPI vesicle formation by facilitating coatomer polymerization. Cell. Logist. 2011, 1, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Stagg, S.M.; LaPointe, P.; Razvi, A.; Gurkan, C.; Potter, C.S.; Carragher, B.; Balch, W.E. Structural Basis for Cargo Regulation of COPII Coat Assembly. Cell 2008, 134, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Peotter, J.; Kasberg, W.; Pustova, I.; Audhya, A. COPII-mediated trafficking at the ER/ERGIC interface. Traffic 2019, 20, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Fath, S.; Mancias, J.D.; Bi, X.; Goldberg, J. Structure and Organization of Coat Proteins in the COPII Cage. Cell 2007, 129, 1325–1336. [Google Scholar] [CrossRef]
- McCaughey, J.; Stephens, D.J. COPII-dependent ER export in animal cells: Adaptation and control for diverse cargo. Histochem. Cell Biol. 2018, 150, 119–131. [Google Scholar] [CrossRef]
- Melville, D.B.; Studer, S.; Schekman, R. Small sequence variations between two mammalian paralogs of the small GTPase SAR1 underlie functional differences in coat protein complex II assembly. J. Biol. Chem. 2020, 295, 8401–8412. [Google Scholar] [CrossRef]
- Sánchez-Simarro, J.; Bernat-Silvestre, C.; Gimeno-Ferrer, F.; Selvi-Martínez, P.; Montero-Pau, J.; Aniento, F.; Marcote, M.J. Loss of Arabidopsis β-COP Function Affects Golgi Structure, Plant Growth and Tolerance to Salt Stress. Front. Plant Sci. 2020, 11, 430. [Google Scholar] [CrossRef]
- Bernat-Silvestre, C.; De Sousa Vieira, V.; Sanchez-Simarro, J.; Pastor-Cantizano, N.; Hawes, C.; Marcote, M.J.; Aniento, F. p24 Family Proteins Are Involved in Transport to the Plasma Membrane of GPI-Anchored Proteins in Plants. Plant Physiol. 2020, 184, 1333–1347. [Google Scholar] [CrossRef]
- Sánchez-Simarro, J.; Bernat-Silvestre, C.; Aniento, F.; Marcote, M.J. ß-COP mutants show specific high sensitivity to chloride ions. Plant Signal. Behav. 2021, 16, 1858629. [Google Scholar] [CrossRef]
- Gimeno-Ferrer, F.; Pastor-Cantizano, N.; Bernat-Silvestre, C.; Selvi-Martínez, P.; Vera-Sirera, F.; Gao, C.; Perez-Amador, M.A.; Jiang, L.; Aniento, F.; Marcote, M.J. α2-COP is involved in early secretory traffic in Arabidopsis and is required for plant growth. J. Exp. Bot. 2017, 68, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Cantizano, N.; Bernat-Silvestre, C.; Marcote, M.J.; Aniento, F. Loss of Arabidopsis p24 function affects ERD2 trafficking and Golgi structure, and activates the unfolded protein response. J. Cell Sci. 2018, 131, jcs203802. [Google Scholar] [CrossRef] [PubMed]
- Cabada Gomez, D.A.; Chavez, M.I.; Cobos, A.N.; Gross, R.J.; Yescas, J.A.; Balogh, M.A.; Indriolo, E. COPI complex isoforms are required for the early acceptance of compatible pollen grains in Arabidopsis thaliana. Plant Reprod. 2020, 33, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.K.; Kang, Y.W.; Lim, H.M.; Hwang, I.; Pai, H.S. Physiological Functions of the COPI Complex in Higher Plants. Mol. Cells 2015, 38, 866–875. [Google Scholar] [CrossRef] [PubMed]
- Min, M.K.; Jang, M.; Lee, M.; Lee, J.; Song, K.; Lee, Y.; Choi, K.Y.; Robinson, D.G.; Hwang, I. Recruitment of Arf1-GDP to Golgi by Glo3p-type ArfGAPs is crucial for golgi maintenance and plant growth. Plant Physiol. 2013, 161, 676–691. [Google Scholar] [CrossRef]
- Pastor-Cantizano, N.; García-Murria, M.J.; Bernat-Silvestre, C.; Marcote, M.J.; Mingarro, I.; Aniento, F. N-Linked Glycosylation of the p24 Family Protein p24δ5 Modulates Retrograde Golgi-to-ER Transport of K/HDEL Ligands in Arabidopsis. Mol. Plant 2017, 10, 1095–1106. [Google Scholar] [CrossRef]
- Montesinos, J.C.; Pastor-Cantizano, N.; Robinson, D.G.; Marcote, M.J.; Aniento, F. Arabidopsis p24δ5 and p24δ9 facilitate Coat Protein I-dependent transport of the K/HDEL receptor ERD2 from the Golgi to the endoplasmic reticulum. Plant J. 2014, 80, 1014–1030. [Google Scholar] [CrossRef]
- Chung, K.P.; Zeng, Y.; Jiang, L. COPII Paralogs in Plants: Functional Redundancy or Diversity? Trends Plant Sci. 2016, 21, 758–769. [Google Scholar] [CrossRef]
- Chang, M.; Wu, S.Z.; Ryken, S.E.; O’Sullivan, J.E.; Bezanilla, M. COPII Sec23 proteins form isoform-specific endoplasmic reticulum exit sites with differential effects on polarized growth. Plant Cell 2022, 34, 333–350. [Google Scholar] [CrossRef]
- Liu, X.; Tong, M.; Zhang, A.; Liu, M.; Zhao, B.; Liu, Z.; Li, Z.; Zhu, X.; Guo, Y.; Li, R. COPII genes SEC31A/B are essential for gametogenesis and interchangeable in pollen development in Arabidopsis. Plant J. 2021, 105, 1600–1614. [Google Scholar] [CrossRef]
- Liang, X.; Li, S.W.; Gong, L.M.; Li, S.; Zhang, Y. COPII Components Sar1b and Sar1c Play Distinct yet Interchangeable Roles in Pollen Development. Plant Physiol. 2020, 183, 974–985. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Chung, K.P.; Li, B.; Lai, C.M.; Lam, S.K.; Wang, X.; Cui, Y.; Gao, C.; Luo, M.; Wong, K.-B.; et al. Unique COPII component AtSar1a/AtSec23a pair is required for the distinct function of protein ER export in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2015, 112, 14360–14365. [Google Scholar] [CrossRef] [PubMed]
- Aboulela, M.; Nakagawa, T.; Oshima, A.; Nishimura, K.; Tanaka, Y. The Arabidopsis COPII components, AtSEC23A and AtSEC23D, are essential for pollen wall development and exine patterning. J. Exp. Bot. 2018, 69, 1615–1633. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Shi, H.; Wang, W.; Liu, X.; Gao, H.; Wang, X.; Zhang, Y.; Yang, M.; Li, R.; Guo, Y. Secretory COPII Protein SEC31B Is Required for Pollen Wall Development. Plant Physiol. 2016, 172, 1625–1642. [Google Scholar] [CrossRef]
- Li, B.; Zeng, Y.; Cao, W.; Zhang, W.; Cheng, L.; Yin, H.; Wu, Q.; Wang, X.; Huang, Y.; Lau, W.C.Y.; et al. A distinct giant coat protein complex II vesicle population in Arabidopsis thaliana. Nat. Plants 2021, 7, 1335–1346. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.G.; Brandizzi, F.; Hawes, C.; Nakano, A. Vesicles versus Tubes: Is Endoplasmic Reticulum-Golgi Transport in Plants Fundamentally Different from Other Eukaryotes? Plant Physiol. 2015, 168, 393–406. [Google Scholar] [CrossRef] [PubMed]
- Mironov, A.A. ER-Golgi transport could occur in the absence of COPII vesicles. Nat. Rev. Mol. Cell Biol. 2014, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Uemura, T.; Nakano, A. The Golgi entry core compartment functions as a COPII-independent scaffold for ER-to-Golgi transport in plant cells. J. Cell Sci. 2018, 131, jcs203893. [Google Scholar] [CrossRef]
- Pagant, S.; Wu, A.; Edwards, S.; Diehl, F.; Miller, E.A. Sec24 is a coincidence detector that simultaneously binds two signals to drive ER export. Curr. Biol. 2015, 25, 403–412. [Google Scholar] [CrossRef]
- Chatterjee, S.; Choi, A.J.; Frankel, G. A systematic review of Sec24 cargo interactome. Traffic 2021, 22, 412–424. [Google Scholar] [CrossRef]
- Gomez-Navarro, N.; Miller, E. Protein sorting at the ER-Golgi interface. J. Cell Biol. 2016, 215, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Geva, Y.; Schuldiner, M. The back and forth of cargo exit from the endoplasmic reticulum. Curr. Biol. 2014, 24, R130–R136. [Google Scholar] [CrossRef] [PubMed]
- Sieben, C.; Mikosch, M.; Brandizzi, F.; Homann, U. Interaction of the K(+)-channel KAT1 with the coat protein complex II coat component Sec24 depends on a di-acidic endoplasmic reticulum export motif. Plant J. 2008, 56, 997–1006. [Google Scholar] [CrossRef]
- Mancias, J.D.; Goldberg, J. Structural basis of cargo membrane protein discrimination by the human COPII coat machinery. EMBO J. 2008, 27, 2918–2928. [Google Scholar] [CrossRef]
- Dancourt, J.; Barlowe, C. Protein sorting receptors in the early secretory pathway. Annu. Rev. Biochem. 2010, 79, 777–802. [Google Scholar] [CrossRef] [PubMed]
- Barlowe, C.K.; Miller, E.A. Secretory protein biogenesis and traffic in the early secretory pathway. Genetics 2013, 193, 383–410. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Nichols, C.D.; Guo, J.; Huang, W.; Lambert, N.A.; Wu, G. A triple arg motif mediates α(2B)-adrenergic receptor interaction with Sec24C/D and export. Traffic 2012, 13, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Otsu, W.; Kurooka, T.; Otsuka, Y.; Sato, K.; Inaba, M. A new class of endoplasmic reticulum export signal PhiXPhiXPhi for transmembrane proteins and its selective interaction with Sec24C. J. Biol. Chem. 2013, 288, 18521–18532. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, L.; Ma, T.; Wang, M.; Li, B.; Jiang, L.; Yan, Y.; Guo, Y. Molecular mechanisms that regulate export of the planar cell-polarity protein Frizzled-6 out of the endoplasmic reticulum. J. Biol. Chem. 2020, 295, 8972–8987. [Google Scholar] [CrossRef]
- Kumichel, A.; Kapp, K.; Knust, E. A Conserved Di-Basic Motif of Drosophila Crumbs Contributes to Efficient ER Export. Traffic 2015, 16, 604–616. [Google Scholar] [CrossRef]
- Woo, C.H.; Gao, C.; Yu, P.; Tu, L.; Meng, Z.; Banfield, D.K.; Yao, X.; Jiang, L. Conserved function of the lysine-based KXD/E motif in Golgi retention for endomembrane proteins among different organisms. Mol. Biol. Cell 2015, 26, 4280–4293. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.P.; Lewis, M.; Kent, H.M.; Edeling, M.A.; Evans, P.R.; Duden, R.; Owen, D.J. Molecular Basis for Recognition of Dilysine Trafficking Motifs by COPI. Dev. Cell 2012, 23, 1255–1262. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Cheng, X.; Sifers, R.N. Golgi-situated endoplasmic reticulum α-1, 2-mannosidase contributes to the retrieval of ERAD substrates through a direct interaction with γ-COP. Mol. Biol. Cell 2013, 24, 1111–1121. [Google Scholar] [CrossRef]
- Kappeler, F.; Klopfenstein, D.R.; Foguet, M.; Paccaud, J.P.; Hauri, H.P. The recycling of ERGIC-53 in the early secretory pathway. ERGIC-53 carries a cytosolic endoplasmic reticulum-exit determinant interacting with COPII. J. Biol. Chem. 1997, 272, 31801–31808. [Google Scholar] [CrossRef] [PubMed]
- Manzano-Lopez, J.; Perez-Linero, A.M.; Aguilera-Romero, A.; Martin, M.E.; Okano, T.; Silva, D.V.; Seeberger, P.H.; Riezman, H.; Funato, K.; Goder, V.; et al. COPII coat composition is actively regulated by luminal cargo maturation. Curr. Biol. 2015, 25, 152–162. [Google Scholar] [CrossRef]
- Malkus, P.; Jiang, F.; Schekman, R. Concentrative sorting of secretory cargo proteins into COPII-coated vesicles. J. Cell Biol. 2002, 159, 915–921. [Google Scholar] [CrossRef]
- Caldwell, S.R.; Hill, K.J.; Cooper, A.A. Degradation of endoplasmic reticulum (ER) quality control substrates requires transport between the ER and Golgi. J. Biol. Chem. 2001, 276, 23296–23303. [Google Scholar] [CrossRef]
- Belden, W.J.; Barlowe, C. Role of Erv29p in collecting soluble secretory proteins into ER-derived transport vesicles. Science 2001, 294, 1528–1531. [Google Scholar] [CrossRef]
- Barlowe, C.; Helenius, A. Cargo Capture and Bulk Flow in the Early Secretory Pathway. Annu. Rev. Cell Dev. Biol. 2016, 32, 197–222. [Google Scholar] [CrossRef]
- Herzig, Y.; Sharpe, H.J.; Elbaz, Y.; Munro, S.; Schuldiner, M. A systematic approach to pair secretory cargo receptors with their cargo suggests a mechanism for cargo selection by Erv14. PLoS Biol. 2012, 10, e1001329. [Google Scholar] [CrossRef]
- Bräuer, P.; Parker, J.L.; Gerondopoulos, A.; Zimmermann, I.; Seeger, M.A.; Barr, F.A.; Newstead, S. Structural basis for pH-dependent retrieval of ER proteins from the Golgi by the KDEL receptor. Science 2019, 363, 1103–1107. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Sato, M.; Nakano, A. Rer1p as common machinery for the endoplasmic reticulum localization of membrane proteins. Proc. Natl. Acad. Sci. USA 1997, 94, 9693–9698. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Sato, M.; Nakano, A. Rer1p, a retrieval receptor for ER membrane proteins, recognizes transmembrane domains in multiple modes. Mol. Biol. Cell 2003, 14, 3605–3616. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, T.; Hoe, M.H.; Slusarewicz, P.; Rabouille, C.; Watson, R.; Hunte, F.; Watzele, G.; Berger, E.G.; Warren, G. Kin recognition between medial Golgi enzymes in HeLa cells. EMBO J. 1994, 13, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Tu, L.; Tai, W.C.; Chen, L.; Banfield, D.K. Signal-mediated dynamic retention of glycosyltransferases in the Golgi. Science 2008, 321, 404–407. [Google Scholar] [CrossRef] [PubMed]
- Schoberer, J.; König, J.; Veit, C.; Vavra, U.; Liebminger, E.; Botchway, S.W.; Altmann, F.; Kriechbaumer, V.; Hawes, C.; Strasser, R. A signal motif retains Arabidopsis ER-α-mannosidase I in the cis-Golgi and prevents enhanced glycoprotein ERAD. Nat. Commun. 2019, 10, 3701. [Google Scholar] [CrossRef]
- Boulaflous, A.; Saint-Jore-Dupas, C.; Herranz-Gordo, M.-C.; Pagny-Salehabadi, S.; Plasson, C.; Garidou, F.; Kiefer-Meyer, M.-C.; Ritzenthaler, C.; Faye, L.; Gomord, V. Cytosolic N-terminal arginine-based signals together with a luminal signal target a type II membrane protein to the plant ER. BMC Plant Biol. 2009, 9, 144. [Google Scholar] [CrossRef]
- Saint-Jore-Dupas, C.; Nebenführ, A.; Boulaflous, A.; Follet-Gueye, M.-L.; Plasson, C.; Hawes, C.; Driouich, A.; Faye, L.; Gomord, V. PlantN-Glycan Processing Enzymes Employ Different Targeting Mechanisms for Their Spatial Arrangement along the Secretory Pathway. Plant Cell 2006, 18, 3182–3200. [Google Scholar] [CrossRef]
- Uemura, S.; Yoshida, S.; Shishido, F.; Inokuchi, J. The cytoplasmic tail of GM3 synthase defines its subcellular localization, stability, and in vivo activity. Mol. Biol. Cell 2009, 20, 3088–3100. [Google Scholar] [CrossRef]
- Schoberer, J.; Vavra, U.; Stadlmann, J.; Hawes, C.; Mach, L.; Steinkellner, H.; Strasser, R. Arginine/lysine residues in the cytoplasmic tail promote ER export of plant glycosylation enzymes. Traffic 2009, 10, 101–115. [Google Scholar] [CrossRef]
- Giraudo, C.G.; Maccioni, H.J. Endoplasmic reticulum export of glycosyltransferases depends on interaction of a cytoplasmic dibasic motif with Sar1. Mol. Biol. Cell 2003, 14, 3753–3766. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Doray, B.; Kornfeld, S. Recycling of Golgi glycosyltransferases requires direct binding to coatomer. Proc. Natl. Acad. Sci. USA 2018, 115, 8984–8989. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, D.; Jing, P.; Wu, Y.; Xia, Y.; Chen, M.; Hong, L. A Novel Golgi Retention Signal RPWS for Tumor Suppressor UBIAD1. PLoS ONE 2013, 8, e72015. [Google Scholar] [CrossRef] [PubMed]
- Schoberer, J.; Liebminger, E.; Vavra, U.; Veit, C.; Castilho, A.; Dicker, M.; Maresch, D.; Altmann, F.; Hawes, C.; Botchway, S.W.; et al. The transmembrane domain of N -acetylglucosaminyltransferase I is the key determinant for its Golgi subcompartmentation. Plant J. 2014, 80, 809–822. [Google Scholar] [CrossRef] [PubMed]
- Schoberer, J.; Liebminger, E.; Vavra, U.; Veit, C.; Grünwald-Gruber, C.; Altmann, F.; Botchway, S.W.; Strasser, R. The Golgi Localization of GnTI Requires a Polar Amino Acid Residue within Its Transmembrane Domain. Plant Physiol. 2019, 180, 859–873. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.L.; Tran, D.T.; Tabak, L.A. Members of the GalNAc-T family of enzymes utilize distinct Golgi localization mechanisms. Glycobiology 2018, 28, 841–848. [Google Scholar] [CrossRef]
- Franke, M.; Braulke, T.; Storch, S. Transport of the GlcNAc-1-phosphotransferase α/β-Subunit Precursor Protein to the Golgi Apparatus Requires a Combinatorial Sorting Motif. J. Biol. Chem. 2013, 288, 1238–1249. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.P.; Zeng, Y.; Li, Y.; Ji, C.; Xia, Y.; Jiang, L. Signal motif-dependent ER export of the Qc-SNARE BET12 interacts with MEMB12 and affects PR1 trafficking in Arabidopsis. J. Cell Sci. 2018, 131, jcs202838. [Google Scholar] [CrossRef]
- Eckert, E.S.; Reckmann, I.; Hellwig, A.; Röhling, S.; El-Battari, A.; Wieland, F.T.; Popoff, V. Golgi Phosphoprotein 3 Triggers Signal-mediated Incorporation of Glycosyltransferases into Coatomer-coated (COPI) Vesicles. J. Biol. Chem. 2014, 289, 31319–31329. [Google Scholar] [CrossRef]
- Rizzo, R.; Russo, D.; Kurokawa, K.; Sahu, P.; Lombardi, B.; Supino, D.; A Zhukovsky, M.; Vocat, A.; Pothukuchi, P.; Kunnathully, V.; et al. Golgi maturation-dependent glycoenzyme recycling controls glycosphingolipid biosynthesis and cell growth via GOLPH3. EMBO J. 2021, 40, e107238. [Google Scholar] [CrossRef]
- Welch, L.G.; Peak-Chew, S.Y.; Begum, F.; Stevens, T.J.; Munro, S. GOLPH3 and GOLPH3L are broad-spectrum COPI adaptors for sorting into intra-Golgi transport vesicles. J. Cell Biol. 2021, 220, e202106115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Nikolovski, N.; Sorieul, M.; Vellosillo, T.; McFarlane, H.E.; DuPree, R.; Kesten, C.; Schneider, R.; Driemeier, C.; Lathe, R.; et al. Golgi-localized STELLO proteins regulate the assembly and trafficking of cellulose synthase complexes in Arabidopsis. Nat. Commun. 2016, 7, 11656. [Google Scholar] [CrossRef]
- Huang, H.H.; Stanley, P. A testis-specific regulator of complex and hybrid N-glycan synthesis. J. Cell Biol. 2010, 190, 893–910. [Google Scholar] [CrossRef] [PubMed]
- Pothukuchi, P.; Agliarulo, I.; Pirozzi, M.; Rizzo, R.; Russo, D.; Turacchio, G.; Nüchel, J.; Yang, J.; Gehin, C.; Capolupo, L.; et al. GRASP55 regulates intra-Golgi localization of glycosylation enzymes to control glycosphingolipid biosynthesis. EMBO J. 2021, 40, e107766. [Google Scholar] [CrossRef] [PubMed]
- Chia, J.; Wang, S.C.; Wee, S.; Gill, D.J.; Tay, F.; Kannan, S.; Verma, C.S.; Gunaratne, J.; Bard, F.A. Src activates retrograde membrane traffic through phosphorylation of GBF1. eLife 2021, 10, e68678. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Wiemels, R.E.; Soya, A.; Whitley, R.; Held, M.; Faik, A. Composition, Assembly, and Trafficking of a Wheat Xylan Synthase Complex. Plant Physiol. 2016, 170, 1999–2023. [Google Scholar] [CrossRef]
- Hassinen, A.; Kellokumpu, S. Organizational interplay of Golgi N-glycosyltransferases involves organelle microenvironment-dependent transitions between enzyme homo- and heteromers. J. Biol. Chem. 2014, 289, 26937–26948. [Google Scholar] [CrossRef]
- Hassinen, A.; Pujol, F.M.; Kokkonen, N.; Pieters, C.; Kihlström, M.; Korhonen, K.; Kellokumpu, S. Functional organization of Golgi N- and O-glycosylation pathways involves pH-dependent complex formation that is impaired in cancer cells. J. Biol. Chem. 2011, 286, 38329–38340. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| COPI-Dependent Transportation | |||||
|---|---|---|---|---|---|
| Motif | Species | Interaction with | Position | GTs | Ref. |
| (F/L)-(L/I/V)-X-X-(R/K) | Yeast | Vps74p | Cytosolic tail | Kre2, Mnn5, Mnn9, Mnn2, Ktr6 | [96] |
| RRXXXXR | Mouse | β-and/or δ-COP (data not shown) | Cytosolic tail | M1-SAT-I | [100] |
| Di-arginine motifs/lumen domain | A.thaliana | Receptor (putative) | Cytosolic tail/ lumen domain | AtGCSI | [98] |
| ϕ- (K/R)-X-L-X-(K/R) | Human | β -, ζ-COP and MHD domain of δ-COP | Cytosolic tail | Ptase, C2GNT1, GALNT3, GALNT8 | [103] |
| ϕ- (K/R)-X-L-X-(K/R) | Human | β -COP and MHD domain of δ-COP | Cytosolic tail | GALNT6 | [103] |
| WTW | Human | β -COP and MHD domain of δ-COP | Cytosolic tail | GALNT4 | [103] |
| Q residue | N. benthamiana/ A.thaliana | TMD | GnTI | [105,106] | |
| Human | GOLPH3 | SiaTI, C2GnT | [110] | ||
| COPII-Dependent Transportation | |||||
| Motif | Species | Interaction with | Position | GTs | Ref. |
| RPWS/ first two TMDs | Human | Sar1 (putative) | Cytosolic tail/ TMDs | UBIAD1 | [104] |
| [RK](X)[RK] | Mouse/ Human | Sar1 Sec23p | Cytosolic tail | GalT2, GalNAcT | [102] |
| RLR | Rat | Cytosolic tail | β1,4GT | [102] | |
| RR, RTR | Chicken | Cytosolic tail | Sial-T2 | [102] | |
| R and K residues | N. tabacum/A.thaliana | Cytosolic tail | GnTI, XylT, GMII | [101] | |
| Di-leucine motif /[RK]X[RK] motifs | Human | N-terminus/C-terminus | Precursor protein of α/β-subunit of PT complex | [108] | |
| Others | |||||
| Motif | Species | Interaction with | Position | GTs | Ref. |
| LPYS | A.thaliana | Cytosolic tail | MNS3 | [97] | |
| Cytosolic tail or luminal stem and TMDs | Human | GalNAc-T7 | [107] | ||
| Luminal stem and TMDs | Human | GalNAc-T10 | [107] | ||
| Cytosolic tail and TMDs | Human | GalNAc-T1, GalNAc-T2 | [107] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Zabotina, O.A. Critical Determinants in ER-Golgi Trafficking of Enzymes Involved in Glycosylation. Plants 2022, 11, 428. https://doi.org/10.3390/plants11030428
Zhang N, Zabotina OA. Critical Determinants in ER-Golgi Trafficking of Enzymes Involved in Glycosylation. Plants. 2022; 11(3):428. https://doi.org/10.3390/plants11030428
Chicago/Turabian StyleZhang, Ning, and Olga A. Zabotina. 2022. "Critical Determinants in ER-Golgi Trafficking of Enzymes Involved in Glycosylation" Plants 11, no. 3: 428. https://doi.org/10.3390/plants11030428
APA StyleZhang, N., & Zabotina, O. A. (2022). Critical Determinants in ER-Golgi Trafficking of Enzymes Involved in Glycosylation. Plants, 11(3), 428. https://doi.org/10.3390/plants11030428