


Chelator Iminodisuccinic Acid Regulates Reactive Oxygen Species Accumulation and Improves Maize (Zea mays L.) Seed Germination under Pb Stress

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
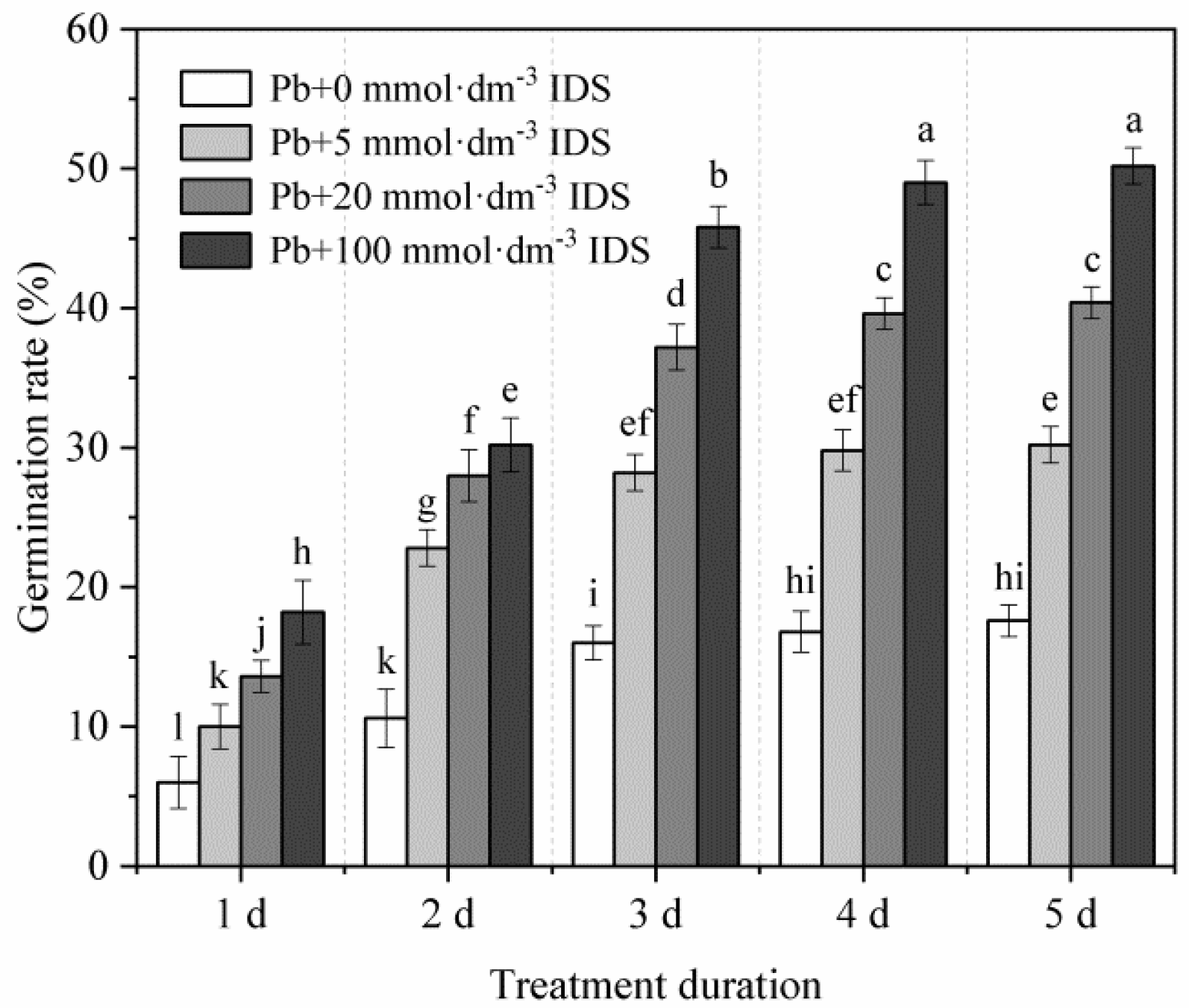
2.1. Effects of IDS and Pb on Maize Seed Germination
2.2. Effects of Pb and IDS on Seedling Growth
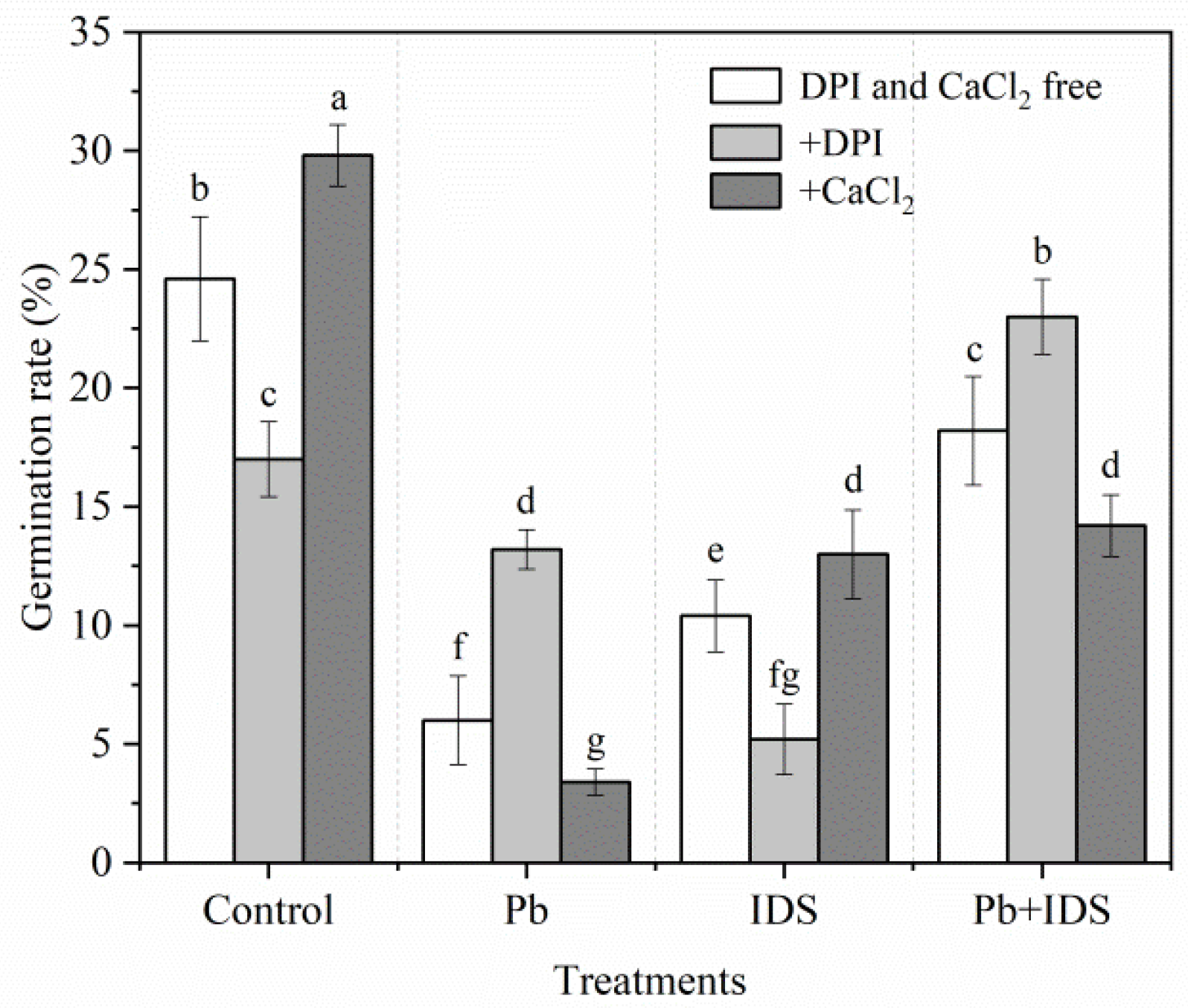
2.3. Effects of ROS-Related Reagents on Seed Germination
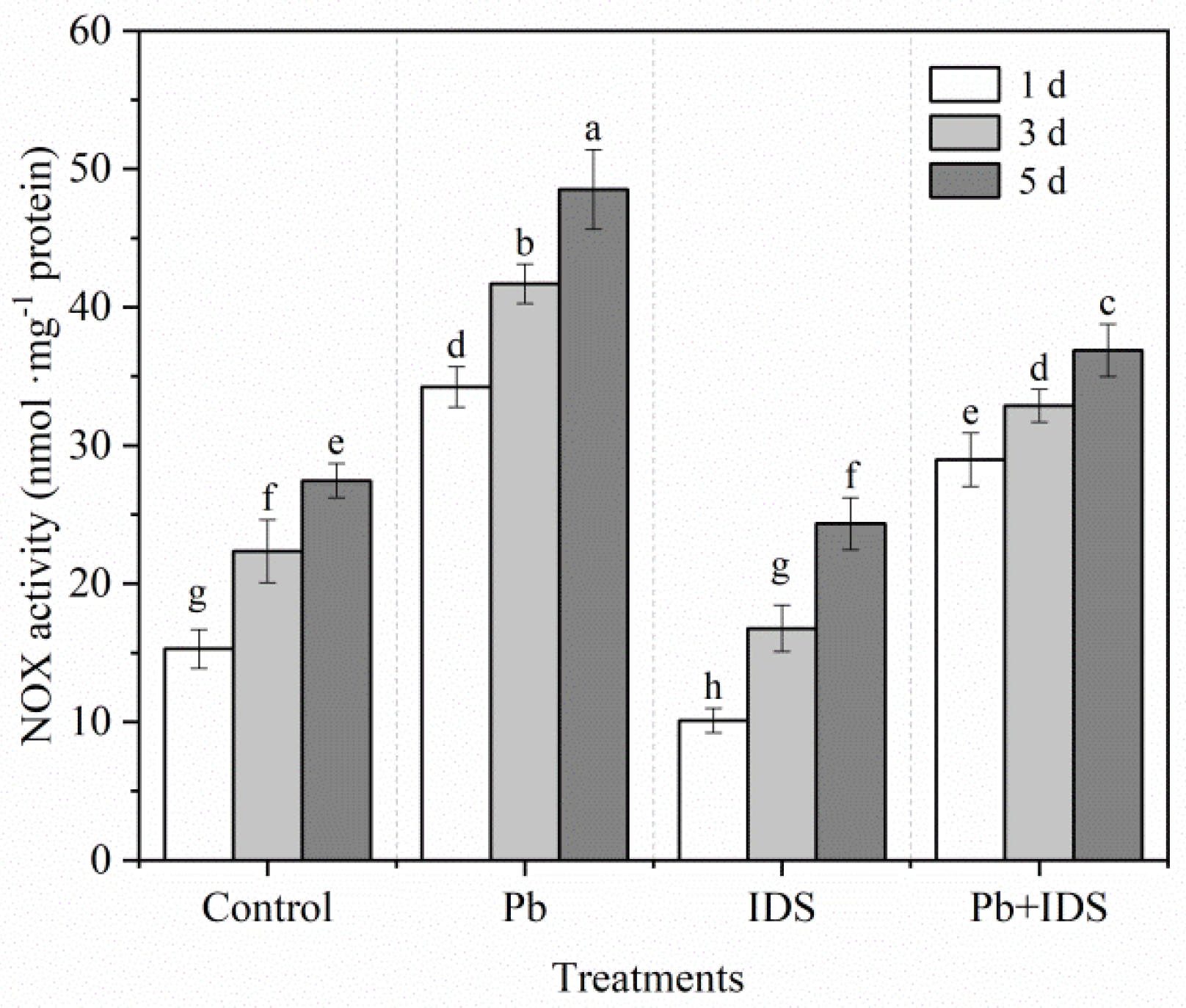
2.4. Effects of Pb and IDS on H2O2 Content and NOX Activity
2.5. Effects of Pb and IDS on Antioxidant Enzyme Activities
3. Discussion
4. Materials and Methods
4.1. Reagent Preparation
4.2. Seed Treatment Groups
- Group 1: The swollen seeds were sown in a plastic box containing quartz stones moistened with 0, 5, 20, or 100 mmol·dm−3 IDS solution. The GR and seedling growth were monitored after sowing.
- Group 2: In accordance with the results of our previous study [45], we used 20 mmol·dm−3 PbCl2 as the Pb stress treatment condition. The swollen seeds were sown in a plastic box containing quartz stones moistened with 20 mmol·dm−3 PbCl2 + 0 mmol·dm−3 IDS, 20 mmol·dm−3 PbCl2 + 5 mmol·dm−3 IDS, 20 mmol·dm−3 PbCl2 + 20 mmol·dm−3 IDS, or 20 mmol·dm−3 PbCl2 + 100 mmol·dm−3 IDS solution. GR and seedling growth were monitored after sowing.
- Group 3: The swollen seeds were sown in a plastic box containing quartz stones moistened with distilled water (control), PbCl2 (20 mmol·dm−3), IDS (100 mmol·dm−3), or PbCl2 (20 mmol·dm−3) + IDS (100 mmol·dm−3) solution. Subsequently, DMTU (a specific scavenger of ROS, 10 mmol·dm−3), H2O2 (10 mmol·dm−3), DPI (a specific inhibitor of NOX, 0.1 mmol·dm−3), CaCl2 (0.1 mmol·dm−3), DDC (a specific inhibitor of SOD, 1 mmol·dm−3), and ATZ (a specific inhibitor of CAT, 1 mmol·dm−3) were applied separately. One milliliter of each ROS-related reagent solution was uniformly added to the quartz stones moistened with the distilled water, PbCl2, IDS, or PbCl2+IDS solutions, respectively.
4.3. Determination of GR
4.4. Determination of Seedling Growth
4.5. Determination of H2O2 Content
4.6. Determination of Antioxidant Enzyme Activities
4.7. NADPH Oxidase Enzyme Assay
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamran, M.; Danish, M.; Saleem, M.H.; Malik, Z.; Parveen, A.; Abbasi, G.H.; Jamil, M.; Ali, S.; Afzal, S.; Riaz, M.; et al. Application of abscisic acid and 6-benzylaminopurine modulated morpho-physiological and antioxidative defense responses of tomato (Solanum lycopersicum L.) by minimizing cobalt uptake. Chemosphere 2021, 263, 128169. [Google Scholar] [CrossRef]
- Saleem, M.H.; Ali, S.; Rehman, M.; Rana, M.S.; Rizwan, M.; Kamran, M.; Imran, M.; Riaz, M.; Soliman, M.H.; Elkelish, A.; et al. Influence of phosphorus on copper phytoextraction via modulating cellular organelles in two jute (Corchorus capsularis L.) varieties grown in a copper mining soil of Hubei Province, China. Chemosphere 2020, 248, 126032. [Google Scholar] [CrossRef] [PubMed]
- Zaheer, I.E.; Ali, S.; Saleem, M.H.; Yousaf, H.S.; Malik, A.; Abbas, Z.; Rizwan, M.; Abualreesh, M.H.; Alatawi, A.; Wang, X. Combined application of zinc and iron-lysine and its effects on morpho-physiological traits, antioxidant capacity and chromium uptake in rapeseed (Brassica napus L.). PLoS ONE 2022, 17, e0262140. [Google Scholar] [CrossRef] [PubMed]
- Afzal, J.; Wang, X.; Saleem, M.H.; Sun, X.; Hussain, S.; Khan, I.; Rana, M.S.; Ahmed, S.; Awan, S.A.; Fiaz, S.; et al. Application of ferrous sulfate alleviates negative impact of cadmium in rice (Oryza sativa L.). Biocell 2021, 45, 1631–1649. [Google Scholar] [CrossRef]
- Ghani, M.A.; Abbas, M.M.; Ali, B.; Aziz, R.; Qadri, R.W.K.; Noor, A.; Azam, M.; Bahzad, S.; Saleem, M.H.; Abualreesh, M.H.; et al. Alleviating role of gibberellic acid in enhancing plant growth and stimulating phenolic compounds in carrot (Daucus carota L.) under lead stress. Sustainability 2021, 13, 12329. [Google Scholar] [CrossRef]
- Saleem, M.H.; Wang, X.; Ali, S.; Zafar, S.; Nawaz, M.; Adnan, M.; Fahad, S.; Shah, A.; Alyemeni, M.N.; Hefft, D.I.; et al. Interactive effects of gibberellic acid and NPK on morpho-physio-biochemical traits and organic acid exudation pattern in coriander (Coriandrum sativum L.) grown in soil artificially spiked with boron. Plant Physiol. Biochem. 2021, 167, 884–900. [Google Scholar] [CrossRef]
- Kamal, A.; Saleem, M.H.; Alshaya, H.; Okla, M.K.; Chaudhary, H.J.; Munis, M.F.H. Ball-milled synthesis of maize biochar-ZnO nanocomposite (MB-ZnO) and estimation of its photocatalytic ability against different organic and inorganic pollutants. J. Saudi Chem. Soc. 2022, 26, 101445. [Google Scholar] [CrossRef]
- Tariq, M.; Ahmad, B.; Adnan, M.; Mian, I.A.; Khan, S.; Fahad, S.; Saleem, M.H.; Ali, M.; Mussarat, M.; Ahmad, M.; et al. Improving boron use efficiency via different application techniques for optimum production of good quality potato (Solanum tuberosum L.) in alkaline soil. PLoS ONE 2022, 17, e0259403. [Google Scholar] [CrossRef]
- Zaheer, I.E.; Ali, S.; Saleem, M.H.; Ashraf, M.A.; Ali, Q.; Abbas, Z.; Rizwan, M.; El-Sheikh, M.A.; Alyemeni, M.N.; Wijaya, L. Zinc-lysine supplementation mitigates oxidative stress in rapeseed (Brassica napus L.) by preventing phytotoxicity of chromium, when irrigated with tannery wastewater. Plants 2020, 9, 1145. [Google Scholar] [CrossRef]
- Hussain, I.; Saleem, M.H.; Mumtaz, S.; Rasheed, R.; Ashraf, M.A.; Maqsood, F.; Rehman, M.; Yasmin, H.; Ahmed, S.; Ishtiaq, M.; et al. Choline chloride mediates chromium tolerance in spinach (Spinacia oleracea L.) by restricting its uptake in relation to morpho-physio-biochemical attributes. J. Plant Growth Regul. 2022, 41, 1594–1614. [Google Scholar] [CrossRef]
- Khan, A.A.; Gul, J.; Naqvi, S.R.; Ali, I.; Farooq, W.; Liaqat, R.; AlMohamadi, H.; Štěpanec, L.; Juchelková, D. Recent progress in microalgae-derived biochar for the treatment of textile industry wastewater. Chemosphere 2022, 306, 135565. [Google Scholar] [CrossRef] [PubMed]
- Ajmal, A.W.; Saroosh, S.; Mulk, S.; Hassan, M.N.; Yasmin, H.; Jabeen, Z.; Nosheen, A.; Shah, S.M.U.; Naz, R.; Hasnain, Z.; et al. Bacteria isolated from wastewater irrigated agricultural soils adapt to heavy metal toxicity while maintaining their plant growth promoting traits. Sustainability 2021, 13, 7792. [Google Scholar] [CrossRef]
- Kazamias, G.; Zorpas, A.A. Drill cuttings waste management from oil & gas exploitation industries through end-of-waste criteria in the framework of circular economy strategy. J. Clean. Prod. 2021, 322, 129098. [Google Scholar] [CrossRef]
- Sanzone, D.M.; Vinhaeiro, N.; Neff, J. Environmental Fates and Effects of Ocean Discharge of Drill Cuttings and Associated Drilling Fluids from Offshore Oil and Gas Operations [IOGP Report]; International Association of Oil & Gas Producers: London, UK, 2016; Volume 543. [Google Scholar]
- Leonard, S.A.; Stegemann, J.A. Stabilization/solidification of petroleum drill cuttings. J. Hazard. Mater. 2010, 174, 463–472. [Google Scholar] [CrossRef]
- De Almeida, P.C.; de Queiroz Fernandes Araújo, O.; de Medeiros, J.L. Managing offshore drill cuttings waste for improved sustainability. J. Clean. Prod. 2017, 165, 143–156. [Google Scholar] [CrossRef]
- Fernandes, I.; Pascoal, C.; Cássio, F. Intraspecific traits change biodiversity effects on ecosystem functioning under metal stress. Oecologia 2011, 166, 1019–1028. [Google Scholar] [CrossRef]
- Xu, T.; Wang, L.; Wang, X.; Li, T.; Zhan, X. Heavy metal pollution of oil-based drill cuttings at a shale gas drilling field in Chongqing, China: A human health risk assessment for the workers. Ecotoxicol. Environ. Saf. 2018, 165, 160–163. [Google Scholar] [CrossRef]
- Thewys, T.; Witters, N.; Van Slycken, S.; Ruttens, A.; Meers, E.; Tack, F.M.G.; Vangronsveld, J. Economic viability of phytoremediation of a cadmium contaminated agricultural area using energy maize. Part I: Effect on the farmer’s income. Int. J. Phytoremediation 2010, 12, 650–662. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; ur Rehman, M.Z.; Rinklebe, J.; Tsang, D.C.W.; Bashir, A.; Maqbool, A.; Tack, F.M.G.; Ok, Y.S. Cadmium phytoremediation potential of Brassica crop species: A review. Sci. Total Environ. 2018, 631–632, 1175–1191. [Google Scholar] [CrossRef]
- Van Slycken, S.; Witters, N.; Meers, E.; Peene, A.; Michels, E.; Adriaensen, K.; Ruttens, A.; Vangronsveld, J.; Du Laing, G.; Wierinck, I.; et al. Safe use of metal-contaminated agricultural land by cultivation of energy maize (Zea mays). Environ. Pollut. 2013, 178, 375–380. [Google Scholar] [CrossRef]
- Rizwan, M.; Ali, S.; Qayyum, M.F.; Ok, Y.S.; Zia-ur-Rehman, M.; Abbas, Z.; Hannan, F. Use of maize (Zea mays L.) for phytomanagement of Cd-contaminated soils: A critical review. Environ. Geochem. Health 2017, 39, 259–277. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Zhang, Y.; Hu, N.; Shi, Y.; Li, T.; Zhao, Z. Differential responses of 23 maize cultivar seedlings to an arbuscular mycorrhizal fungus when grown in a metal-polluted soil. Sci. Total Environ. 2021, 789, 148015. [Google Scholar] [CrossRef] [PubMed]
- Meers, E.; Ruttens, A.; Hopgood, M.; Lesage, E.; Tack, F.M.G. Potential of Brassic rapa, Cannabis sativa, Helianthus annuus and Zea mays for phytoextraction of heavy metals from calcareous dredged sediment derived soils. Chemosphere 2005, 61, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Vigliotta, G.; Matrella, S.; Cicatelli, A.; Guarino, F.; Castiglione, S. Effects of heavy metals and chelants on phytoremediation capacity and on rhizobacterial communities of maize. J. Environ. Manag. 2016, 179, 93–102. [Google Scholar] [CrossRef]
- Wang, A.; Wang, M.; Liao, Q.; He, X. Characterization of Cd translocation and accumulation in 19 maize cultivars grown on Cd-contaminated soil: Implication of maize cultivar selection for minimal risk to human health and for phytoremediation. Environ. Sci. Pollut. Res. Int. 2016, 23, 5410–5419. [Google Scholar] [CrossRef]
- Maitra, S.; Singh, V. Invited review on ‘maize in the 21st century’ Emerging trends of maize biorefineries in the 21st century: Scientific and technological advancements in biofuel and bio-sustainable market. J. Cereal Sci. 2021, 101, 103272. [Google Scholar] [CrossRef]
- Seneviratne, M.; Rajakaruna, N.; Rizwan, M.; Madawala, H.M.S.P.; Ok, Y.S.; Vithanage, M. Correction to: Heavy metal-induced oxidative stress on seed germination and seedling development: A critical review. Environ. Geochem. Health 2019, 41, 1635. [Google Scholar] [CrossRef]
- Soares, T.F.S.N.; dos Santos Dias, D.C.F.; Oliveira, A.M.S.; Ribeiro, D.M.; dos Santos Dias, L.A. Exogenous brassinosteroids increase lead stress tolerance in seed germination and seedling growth of Brassica juncea L. Ecotoxicol. Environ. Saf. 2020, 193, 110296. [Google Scholar] [CrossRef]
- Watanabe, M.E. Phytoremediation on the brink of commericialization. Environ. Sci. Technol. 1997, 31, 182A–186A. [Google Scholar] [CrossRef]
- Ahmad, M.; Ahmad, I.; Nazli, F.; Mumtaz, M.Z.; Latif, M.; Al-Mosallam, M.S.; Alotaibi, F.S.; Dewidar, A.Z.; Mattar, M.A.; El-Shafei, A.A. Lead-Tolerant Bacillus Strains Promote Growth and Antioxidant Activities of Spinach (Spinacia oleracea) Treated with Sewage Water. Agronomy 2021, 11, 2482. [Google Scholar] [CrossRef]
- Agency for Toxic Substances and Disease Registry. 2022. Available online: https://www.atsdr.cdc.gov/ (accessed on 8 August 2021).
- Jayasri, M.A.; Suthindhiran, K. Effect of zinc and lead on the physiological and biochemical properties of aquatic plant Lemna minor: Its potential role in phytoremediation. Appl. Water Sci. 2017, 7, 1247–1253. [Google Scholar] [CrossRef]
- Fatemi, H.; Pour, B.E.; Rizwan, M. Isolation and characterization of lead (Pb) resistant microbes and their combined use with silicon nanoparticles improved the growth, photosynthesis and antioxidant capacity of coriander (Coriandrum sativum L.) under Pb stress. Environ. Pollut. 2020, 266, 114982. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Yu, F.; Li, Y.; Niu, W.; Li, J.; Yang, J.; Liu, K. Comparative analysis of the seed germination of pakchoi and its phytoremediation efficacy combined with chemical amendment in four polluted soils. Int. J. Phytoremediation 2020, 22, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.F.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.G.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Van Breusegem, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. BioEssays 2006, 28, 1091–1101. [Google Scholar] [CrossRef]
- Wojtyla, Ł.; Lechowska, K.; Kubala, S.; Garnczarska, M. Different modes of hydrogen peroxide action during seed germination. Front. Plant Sci. 2016, 7, 66. [Google Scholar] [CrossRef]
- Gomes, M.P.; Carneiro, M.M.L.C.; Nogueira, C.O.G.; Soares, A.M.; Garcia, Q.S. The system modulating ROS content in germinating seeds of two Brazilian savanna tree species exposed to As and Zn. Acta Physiol. Plant 2013, 35, 1011–1022. [Google Scholar] [CrossRef]
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative signaling in seed germination and dormancy. Plant Signal. Behav. 2008, 3, 175–182. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Thakur, S.; Singh, L.; Zularisam, A.W.; Sakinah, M.; Din, M.F.M. Lead induced oxidative stress and alteration in the activities of antioxidative enzymes in rice shoots. Biologia Plant. 2017, 61, 595–598. [Google Scholar] [CrossRef]
- Kaurin, A.; Gluhar, S.; Tilikj, N.; Lestan, D. Soil washing with biodegradable chelating agents and EDTA: Effect on soil properties and plant growth. Chemosphere 2020, 260, 127673. [Google Scholar] [CrossRef] [PubMed]
- Staszak, A.M.; Małecka, A.; Ciereszko, I.; Ratajczak, E. Differences in stress defence mechanisms in germinating seeds of Pinus sylvestris exposed to various lead chemical forms. PLoS ONE 2020, 15, e0238448. [Google Scholar] [CrossRef]
- Zhang, Y.; Deng, B.; Li, Z. Inhibition of NADPH oxidase increases defense enzyme activities and improves maize seed germination under Pb stress. Ecotoxicol. Environ. Saf. 2018, 158, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ma, P.; Zhao, Z.; Zhao, G.; Tian, B.; Wang, J.; Wang, G. Mapping QTL controlling maize deep-seeding tolerance-related traits and confirmation of a major QTL for mesocotyl length. Theor. Appl. Genet. 2012, 124, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Virk, S.S.; Fulton, J.P.; Porter, W.M.; Pate, G.L. Row-crop planter performance to support variable-rate seeding of maize. Precis. Agric. 2020, 21, 603–619. [Google Scholar] [CrossRef]
- Rostami, S.; Azhdarpoor, A. The application of plant growth regulators to improve phytoremediation of contaminated soils: A review. Chemosphere 2019, 220, 818–827. [Google Scholar] [CrossRef]
- Menhas, S.; Hayat, K.; Niazi, N.K.; Zhou, P.; Amna; Bundschuh, J.; Naeem, M.; Munis, M.F.H.; Yang, X.; Chaudhary, H.J. Microbe-EDTA mediated approach in the phytoremediation of lead-contaminated soils using maize (Zea mays L.) plants. Int. J. Phytoremediation 2021, 23, 585–596. [Google Scholar] [CrossRef]
- Kołodyńska, D. Application of a new generation of complexing agents in removal of heavy metal ions from different wastes. Environ. Sci. Pollut. Res. Int. 2013, 20, 5939–5949. [Google Scholar] [CrossRef]
- Kołodyńska, D. Iminodisuccinic acid as a new complexing agent for removal of heavy metal ions from industrial effluents. Chem. Eng. J. 2009, 152, 277–288. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef]
- Boussen, S.; Soubrand, M.; Bril, H.; Ouerfelli, K.; Abdeljaouad, S. Transfer of lead, zinc and cadmium from mine tailings to wheat (Triticum aestivum) in carbonated Mediterranean (Northern Tunisia) soils. Geoderma 2013, 192, 227–236. [Google Scholar] [CrossRef]
- Guo, J.; Lv, X.; Jia, H.; Hua, L.; Ren, X.; Muhammad, H.; Wei, T.; Ding, Y. Effects of EDTA and plant growth-promoting rhizobacteria on plant growth and heavy metal uptake of hyperaccumulator Sedum alfredii Hance. J. Environ. Sci. 2020, 88, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Budelsky, R.A.; Galatowitsch, S.M. Effects of moisture, temperature, and time on seed germination of five wetland Carices: Implications for restoration. Restor. Ecol. 1999, 7, 86–97. [Google Scholar] [CrossRef]
- Jurado, E.; Flores, J. Is seed dormancy under environmental control or bound to plant traits? J. Veg. Sci. 2005, 16, 559–564. [Google Scholar] [CrossRef]
- Kranner, I.; Colville, L. Metals and seeds: Biochemical and molecular implications and their significance for seed germination. Environ. Exp. Bot. 2011, 72, 93–105. [Google Scholar] [CrossRef]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. C. R. Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef]
- Ziegelstein, R.C.; Zweier, J.L.; Mellits, E.D.; Younes, A.; Lakatta, E.G.; Stern, M.D.; Silverman, H.S. Dimethylthiourea, an oxygen radical scavenger, protects isolated cardiac myocytes from hypoxic injury by inhibition of Na(+)-Ca2+ exchange and not by its antioxidant effects. Circ. Res. 1992, 70, 804–811. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Tawaratsumida, T.; Zheng, S.; Yuasa, T.; Iwaya-Inoue, M. NADPH oxidases act as key enzyme on germination and seedling growth in barley (Hordeum vulgare L.). Plant Prod. Sci. 2010, 13, 45–52. [Google Scholar] [CrossRef]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox. Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef]
- Kim, S.Y.; Lee, S.M.; Park, J. Antioxidant enzyme inhibitors enhance singlet oxygen-induced cell death in HL-60 cells. Free Radic. Res. 2006, 40, 1190–1197. [Google Scholar] [CrossRef]
- Tabatabaie, T.; Floyd, R.A. Susceptibility of glutathione peroxidase and glutathione reductase to oxidative damage and the protective effect of spin trapping agents. Arch. Biochem. Biophys. 1994, 314, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Escobar, J.A.; Rubio, M.A.; Lissi, E.A. Sod and catalase inactivation by singlet oxygen and peroxyl radicals. Free Radic. Biol. Med. 1996, 20, 285–290. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, B.; Xu, Z.; Shi, Z.; Chen, S.; Huang, X.; Chen, J.; Wang, X. Involvement of reactive oxygen species in endosperm cap weakening and embryo elongation growth during lettuce seed germination. J. Exp. Bot. 2014, 65, 3189–3200. [Google Scholar] [CrossRef] [PubMed]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Grace, S.C.; Logan, B.A. Acclimation of foliar antioxidant systems to growth irradiance in three broad-leaved evergreen species. Plant Physiol. 1996, 112, 1631–1640. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Sun, Y.; Li, W.; Li, J.; Xu, R.; Du, J.; Li, Z.; Li, G.; Yang, K. Chelator Iminodisuccinic Acid Regulates Reactive Oxygen Species Accumulation and Improves Maize (Zea mays L.) Seed Germination under Pb Stress. Plants 2022, 11, 2487. https://doi.org/10.3390/plants11192487
Zhang Y, Sun Y, Li W, Li J, Xu R, Du J, Li Z, Li G, Yang K. Chelator Iminodisuccinic Acid Regulates Reactive Oxygen Species Accumulation and Improves Maize (Zea mays L.) Seed Germination under Pb Stress. Plants. 2022; 11(19):2487. https://doi.org/10.3390/plants11192487
Chicago/Turabian StyleZhang, Yifei, Yishan Sun, Weiqing Li, Jiayu Li, Rongqiong Xu, Jiarui Du, Zesong Li, Guibin Li, and Kejun Yang. 2022. "Chelator Iminodisuccinic Acid Regulates Reactive Oxygen Species Accumulation and Improves Maize (Zea mays L.) Seed Germination under Pb Stress" Plants 11, no. 19: 2487. https://doi.org/10.3390/plants11192487
APA StyleZhang, Y., Sun, Y., Li, W., Li, J., Xu, R., Du, J., Li, Z., Li, G., & Yang, K. (2022). Chelator Iminodisuccinic Acid Regulates Reactive Oxygen Species Accumulation and Improves Maize (Zea mays L.) Seed Germination under Pb Stress. Plants, 11(19), 2487. https://doi.org/10.3390/plants11192487