
A Pan-Global Study of Bacterial Leaf Spot of Chilli Caused by Xanthomonas spp.


Abstract
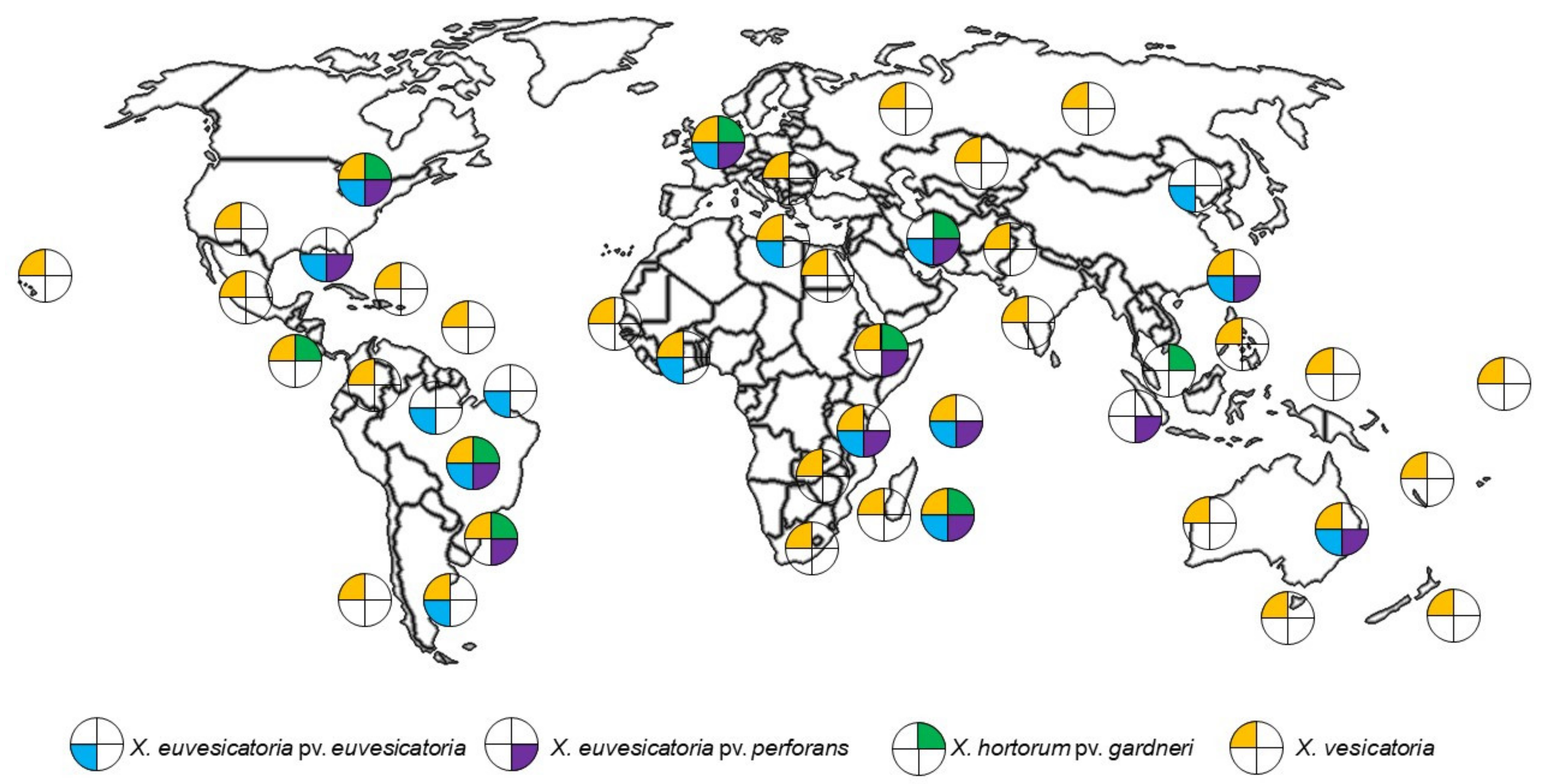
1. Introduction
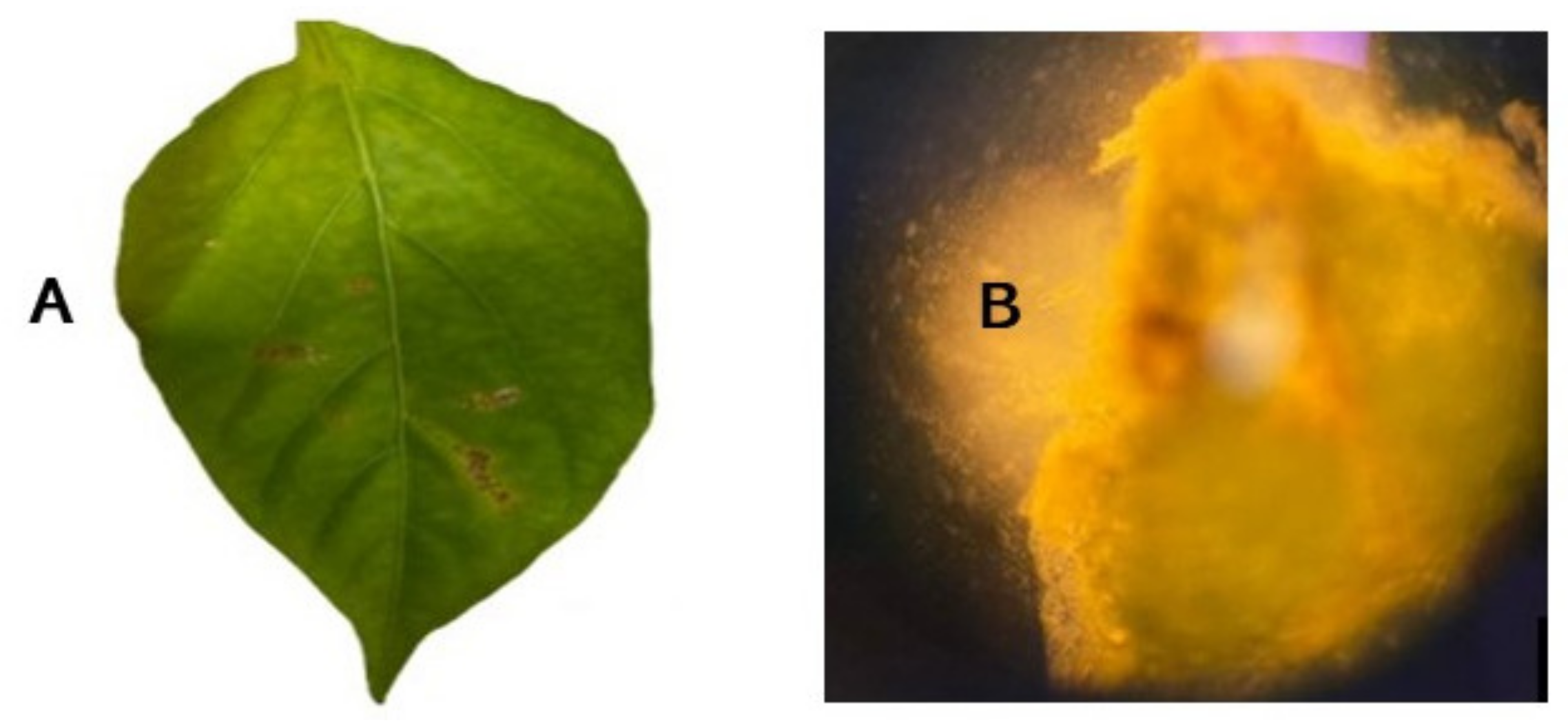
2. Bacterial Leaf Spot
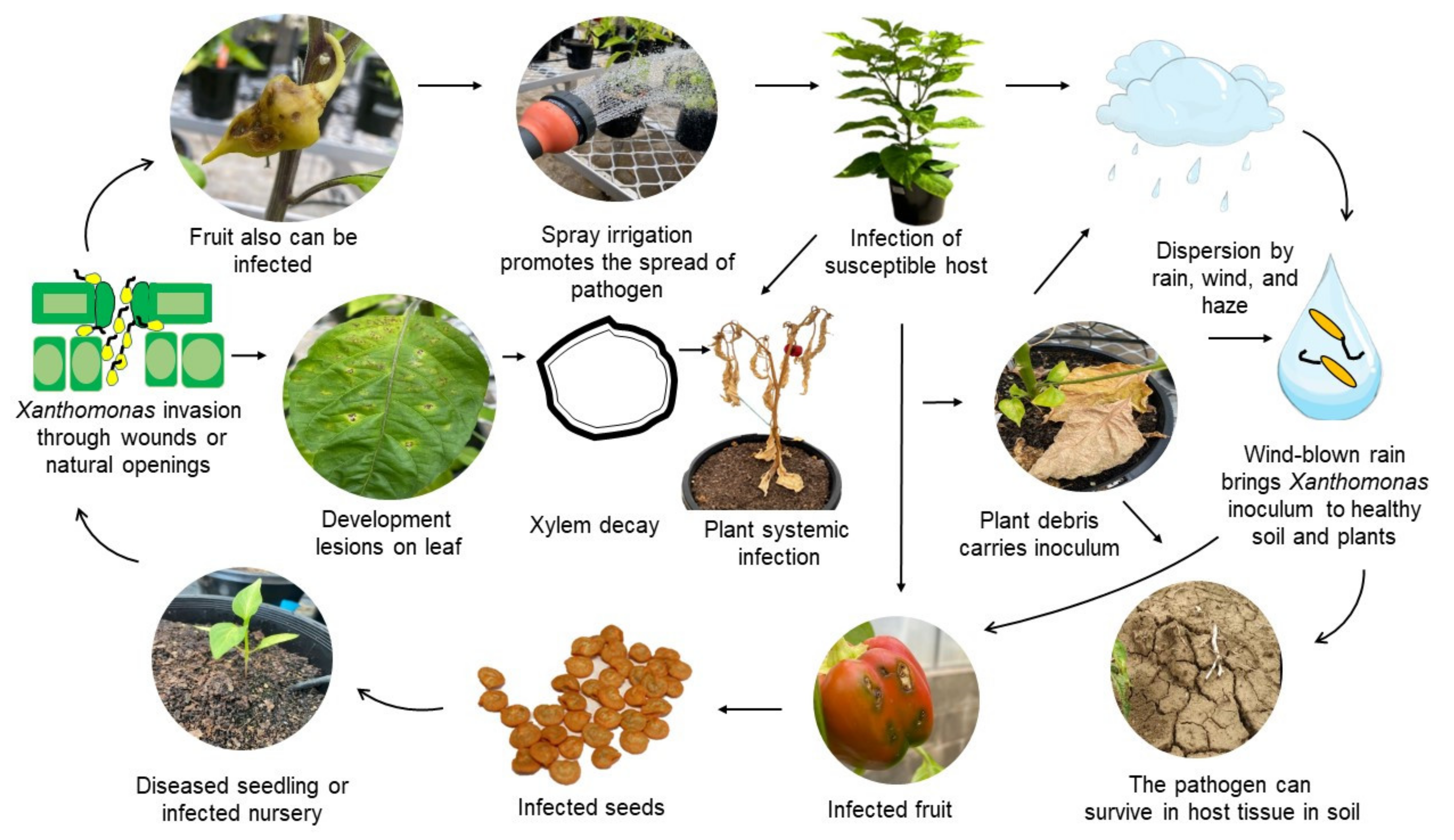
3. Infection Process and Life Cycle
4. Disease Detection
5. Management and Control
5.1. Chemical Control
5.2. Biological Control
6. Host–Pathogen Interactions
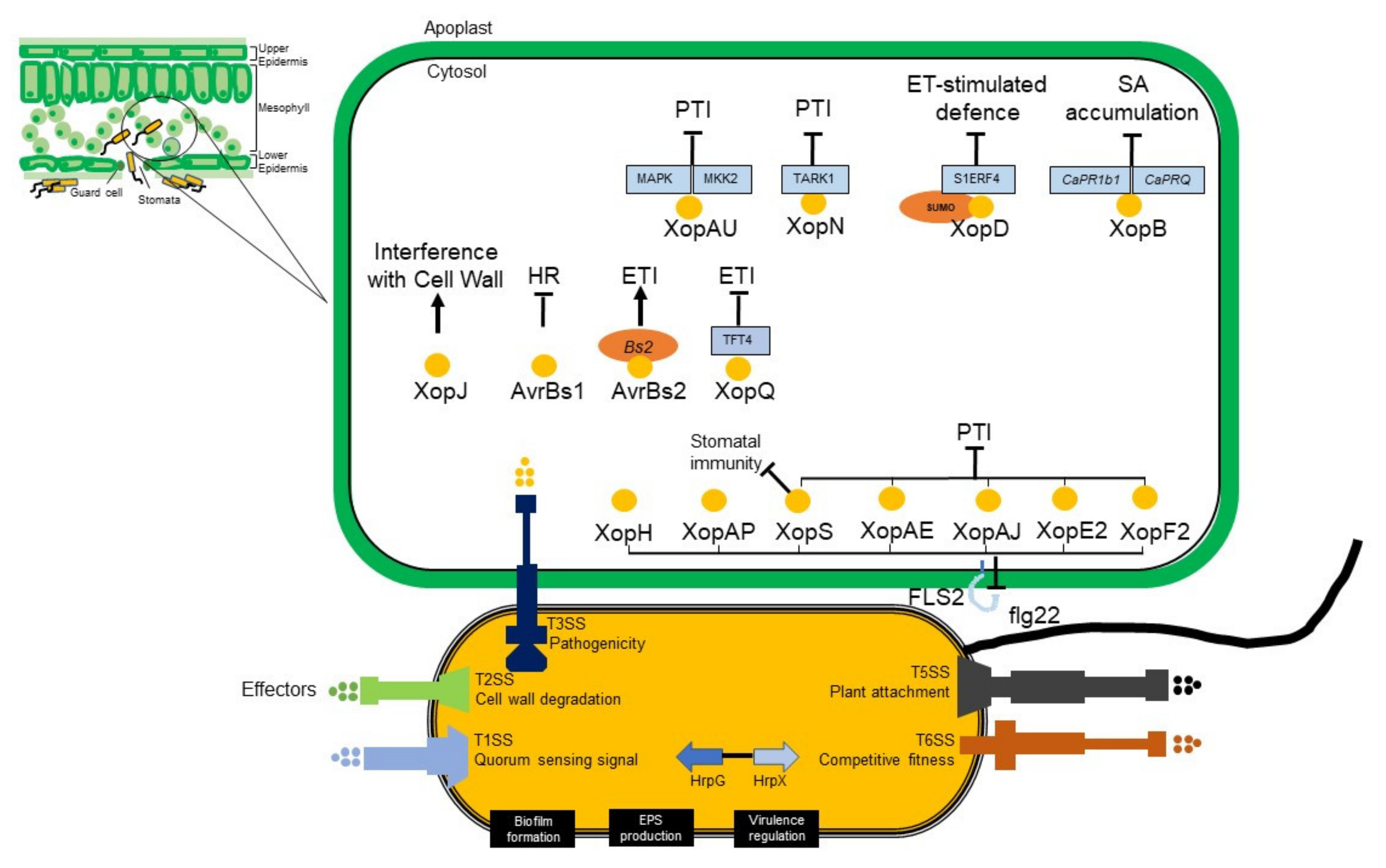
7. Type III Effector Biology and Action
8. Resistance Genes
9. Genes Involved in Plant Defenses
10. Conclusions and Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bosland, P.W.; Votava, E.J. Peppers: Vegetable and Spice Capsicums; CABI: Oxford, UK, 2012. [Google Scholar]
- Pickersgill, B. Relationships between weedy and cultivated forms in some species of chilli peppers (Genus Capsicum). Evolution 1971, 25, 683–691. [Google Scholar]
- Ramchiary, N.; Kole, C. The Capsicum Genome; Springer: New Delhi, India, 2019. [Google Scholar]
- Nunn, N.; Qian, N. The Columbian Exchange: A History of Disease, Food, and Ideas. J. Econ. Perspect. 2010, 24, 163–188. [Google Scholar] [CrossRef]
- Shaffer, M. Pepper: A History of the World’s Most Influential Spice; St. Martin’s Press: New York, NY, USA, 2013. [Google Scholar]
- Naves, E.R.; Silva, L.A.; Sulpice, R.; Araujo, W.L.; Nunes-Nesi, A.; Peres, L.E.P.; Zsogon, A. Capsaicinoids: Pungency beyond Capsicum. Trends Plant Sci. 2019, 24, 109–120. [Google Scholar] [CrossRef]
- Barbero, G.F.; Liazid, A.; Azaroual, L.; Palma, M.; Barroso, C.G. Capsaicinoid contents in peppers and pepper-related spicy foods. Int. J. Food Prop. 2016, 19, 485–493. [Google Scholar] [CrossRef]
- Tewksbury, J.J.; Nabhan, G.P. Directed deterrence by capsaicin in chillies. Nature 2001, 412, 403–404. [Google Scholar] [CrossRef]
- Mankowski, C.; Poole, C.D.; Ernault, E.; Thomas, R.; Berni, E.; Currie, C.J.; Treadwell, C.; Calvo, J.I.; Plastira, C.; Zafeiropoulou, E.; et al. Effectiveness of the capsaicin 8% patch in the management of peripheral neuropathic pain in European clinical practice: The ASCEND study. BMC Neurol. 2017, 17, 80. [Google Scholar] [CrossRef]
- Cserhati, T.; Forgacs, E.; Morais, M.H.; Mota, T.; Ramos, A. Separation and quantitation of colour pigments of chili powder (Capsicum frutescens) by high-performance liquid chromatography–diode array detection. J. Chromatogr. A 2000, 896, 69–73. [Google Scholar] [CrossRef]
- Li, B.; Yang, M.; Shi, R.; Ye, M. Insecticidal activity of natural capsaicinoids against several agricultural insects. Nat. Prod. Commun. 2019, 14, 1–7. [Google Scholar] [CrossRef]
- Kantar, M.B.; Anderson, J.E.; Lucht, S.A.; Mercer, K.; Bernau, V.; Case, K.A.; Le, N.C.; Frederiksen, M.K.; DeKeyser, H.C.; Wong, Z.; et al. Vitamin variation in Capsicum spp. provides opportunities to improve nutritional value of human diets. PLoS ONE 2016, 11, e0161464. [Google Scholar] [CrossRef]
- Olatunji, T.L.; Afolayan, A.J. The suitability of chili pepper (Capsicum annuum L.) for alleviating human micronutrient dietary deficiencies: A review. Food Sci. Nutr. 2018, 6, 2239–2251. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Crops and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 13 July 2022).
- Food and Agriculture Organization of the United Nations. FAOSTAT. Available online: http://faostat.fao.org/site/567/default.aspx#ancor (accessed on 2 January 2022).
- Abewoy, D. Review on impacts of climate change on vegetable production and its management practices. Adv. Crop Sci. 2018, 6, 330. [Google Scholar] [CrossRef]
- Cook, A.A.; Stall, R.E. Differentiation of pathotypes among isolates of Xanthomonas vesicatoria. Plant Dis. Rep. 1969, 53, 617–622. [Google Scholar]
- Goode, M.; Sasser, M. Prevention-The key to controlling bacterial spot and bacterial speck of tomato. Plant Dis. 1980, 64, 831–834. [Google Scholar] [CrossRef]
- Pajcin, I.S.; Vlaijkov, V.R.; Cvetkovic, D.D.; Ignjatov, M.V.; Grahovac, M.S.; Vucurovic, D.G.; Grahovac, J.A. Selection of antagonists for biocontrol of Xanthomonas euvesicatoria. Acta Period. Technol. 2020, 51, 181–189. [Google Scholar] [CrossRef]
- Roach, R.; Mann, R.; Gambley, C.G.; Shivas, R.G.; Rodoni, B. Identification of Xanthomonas species associated with bacterial leaf spot of tomato, capsicum, and chilli crops in eastern Australia. Eur. J. Plant Pathol. 2018, 150, 595–608. [Google Scholar] [CrossRef]
- Schwartz, A.R.; Potnis, N.; Timilsina, S.; Wilson, M.; Patane, J. Phylogenomics of Xanthomonas field strains infecting pepper and tomato reveals diversity in effector repertoires and identifies determinants of host specificity. Front. Microbiol. 2015, 6, 535. [Google Scholar] [CrossRef]
- Noble, T.J.; Young, A.J.; Barrerro, R.; Douglas, C.A.; Kelly, L.A.; Williams, B.; Mundree, S. Characterisation of the Pseudomonas savastanoi pv. phaseolicola population found in Eastern Australia associated with halo blight disease in Vigna radiata. Australas. Plant Pathol. 2020, 49, 515–524. [Google Scholar] [CrossRef]
- Doidge, E.M. A tomato canker. Ann. Appl. Biol. 1921, 7, 407–430. [Google Scholar] [CrossRef]
- Gardner, M.W.; Kendrick, J.B. Bacterial spot of tomato. J. Agric. Res. 1921, 21, 123–156. [Google Scholar]
- Dowson, W.J. On the systematic position and generic names of the gram-negative bacterial plant pathogens. Zent. Fur Bakteriol. Parasitenkd. Und Infekt. 1939, 2, 177–193. [Google Scholar]
- Young, J.M.; Dye, D.W.; Bradbury, J.F.; Panagopoulos, C.G.; Robbs, C.F. A proposed nomenclature and classification for plant pathogenic bacteria. N. Z. J. Agric. Res. 1978, 21, 153–177. [Google Scholar] [CrossRef]
- Vauterin, L.; Hoste, B.; Kersters, K.; Swings, J. Reclassification of Xanthomonas. Int. Syst. Ecol. Microbiol. 1995, 45, 472–489. [Google Scholar] [CrossRef]
- Šutic, D. Bakterioze crvenog patlidzana (Tomato Bacteriosis); Posebna Izd Inst Zastitu Bilja Beograd (Special Edition); Institut za zaštitu bilja: Beograd, Serbia, 1957; Volume 6, pp. 1–65. [Google Scholar]
- Jones, J.B.; Lacy, G.H.; Bouzar, H.; Stall, R.E.; Shcaad, N.W. Reclassification of the xanthomonads associated with bacterial spot disease of tomato and pepper. Syst. Appl. Microbiol. 2004, 27, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Timilsina, S.; Kara, S.; Jacques, M.A.; Potnis, N.; Minsavage, G.V.; Vallad, G.E.; Jones, J.B.; Saux, M.F. Reclassification of Xanthomonas gardneri (ex Šutič 1957) Jones et al. 2006 as a later heterotypic synonym of Xanthomonas cynarae Trébaol et al. 2000 and description of X. cynarae pv. cynarae and X. cynarae pv. gardneri based on whole genome analyses. Microbiol. Soc. 2018, 69, 792–806. [Google Scholar] [CrossRef]
- Moriniere, L.; Burlet, A.; Rosenthal, E.R.; Nesme, X.; Portier, P.; Bull, C.T.; Lavire, C.; Saux, M.F.; Bertolla, F. Clarifying the taxonomy of the causal agent of bacterial leaf spot of lettuce through a polyphasic approach reveals that Xanthomonas cynarae Trébaol et al. 2000 emend. Timilsina et al. 2019 is a later heterotypic synonym of Xanthomonas hortorum Vauterin et al. 1995. Syst. Appl. Microbiol. 2020, 43, 126087. [Google Scholar] [CrossRef] [PubMed]
- Stall, R.E.; Jones, J.B.; Minsavage, G.V. Durability of resistance in tomato and pepper to Xanthomonads causing bacterial spot. Ann. Rev. Phytopathol. 2009, 47, 265–284. [Google Scholar] [CrossRef]
- Keen, N.; Kobayashi, D.; Tamaki, S.; Shen, H.; Stayton, M.; Lawrence, D.; Sharma, A.; Midland, S.; Smith, M.; Sims, J. Avirulence gene D from Pseudomonas syringae pv. tomato and its interaction with resistance gene Rpg4 in soybean. In Current Plant Science and Biotechnology in Agriculture; Hennecke, H., Verma, D.P., Eds.; Springer-Science: Davis, CA, USA, 1991; pp. 37–44. [Google Scholar]
- Cook, A.A.; Guevara, Y.G. Hypersensitivity in Capsicum chacoense to race 1 or the bacterial spot pathogen of pepper. Plant Dis. 1984, 68, 329–330. [Google Scholar] [CrossRef]
- Sahin, F.; Miller, S.A. Resistance in Capsicum pubescens to Xanthomonas campestris pv. vesicatoria pepper race 6. Plant Dis. 1998, 82, 794–799. [Google Scholar] [CrossRef]
- Kousik, C.S.; Ritchie, D.F. Isolation of pepper races 4 and 5 of Xanthomonas campestris pv. vesicatoria from diseased pepper in southeastern U.S fields. Plant Dis. 1995, 79, 540. [Google Scholar] [CrossRef]
- Gambley, C. Management and detection of bacterial leaf spot in capsicum and chili crops. Hort Innovation. Available online: https://www.horticulture.com.au/globalassets/laserfiche/assets/project-reports/vg14010/vg14010---final-report-complete.pdf (accessed on 4 April 2022).
- Ivey, M.L.L.; Strayer, A.; Sidhu, J.K.; Minsavage, G.V. Bacterial leaf spot of bell pepper (Capsicum annuum) in Louisiana is caused by Xanthomonas euvesicatoria Pepper Races 1 and 3. Dis. Not. 2016, 100, 853. [Google Scholar] [CrossRef]
- Burlakoti, R.R.; Hsu, C.; Chen, J.; Wang, J. Population dynamics of Xanthomonads associated with bacterial spot of tomato and pepper during 27 years across Taiwan. Plant Dis. 2018, 102, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Garton, J.E. Evaluation of Race and Copper Tolerant Strains of Xanthomonas axonopodis pv. vesicatoria, Causal Agent of Bacterial Leaf Spot of Bell Pepper in Georgia. Master’s Thesis, University of Georgia, Athens, GA, USA, 2009. [Google Scholar]
- An, S.; Potnis, N.; Dow, M.; Vorholter, F.; He, Y.; Becker, A.; Teper, D.; Li, Y.; Wang, N.; Bleris, L.; et al. Mechanistic insights into host adaptation, virulence and epidemiology of the phytopathogen Xanthomonas. FEMS Microbiol. Rev. 2020, 44, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Hugouvieux, V.; Barber, C.E.; Daniels, M.J. Entry of Xanthomonas campestris pv. Campestris into Hydathodes of Arabidopsis thaliana Leaves: A System for Studying Early Infection Events in Bacterial Pathogenesis. Mol. Plant Microbe Interact. 1998, 11, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Almeida, R.P.P.; Lindow, S. Living in two worlds: The plant and insect lifestyles of Xylella fastidiosa. Annu. Rev. Phytopathol. 2008, 46, 243–271. [Google Scholar] [CrossRef]
- Ryan, R.P.; Vorholter, F.; Potnis, N.; Jones, J.B.; Sluys, M.V.; Bogdanove, A.J.; Dow, J.M. Pathogenomics of Xanthomonas: Understanding bacterium-plant interactions. Nat. Rev. Microbiol. 2011, 9, 344–355. [Google Scholar] [CrossRef]
- Petrocelli, S.; Tondo, M.L.; Daurelio, L.D.; Orellano, E.G. Modifications of Xanthomonas axonopodis pv. citri lipopolysaccharide affect the basal response and the virulence process during citrus canker. PLoS ONE 2012, 7, e40051. [Google Scholar] [CrossRef]
- Silipo, A.; Molinaro, A.; Lanzetta, R.; Parrilli, M. The structures of the lipid A moieties from the lipopolysaccharides of two phytopathogenic bacteria, Xanthomonas campestris pv. pruni and Xanthomonas fragariae. Eur. J. Org. Chem. 2004, 6, 1336–1343. [Google Scholar] [CrossRef]
- Castiblanco, L.F.; Sundin, G.W. New insights on molecular regulation of biofilm formation in plant-associated bacteria. J. Integr. Plant Biol. 2016, 58, 362–372. [Google Scholar] [CrossRef]
- Flemming, H.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Vojnov, A.A.; Slater, H.; Daniels, M.J.; Dow, J.W. Expression of the gum operon directing xanthan biosynthesis in Xanthomonas campestris and its regulation in planta. Mol. Plant Microbe Interact. 2001, 14, 768–774. [Google Scholar] [CrossRef]
- Katzen, F.; Becker, A.; Zorreguieta, A.; Puhler, A.; Lelpi, L. Promoter analysis of the Xanthomonas campestris pv. campestris gum operon directing biosynthesis of the xanthan polysaccharide. J. Bacteriol. 1996, 178, 4313–4318. [Google Scholar] [CrossRef] [PubMed]
- Branda, S.S.; Vik, A.; Friedman, L.; Kolter, R. Biofilms: The matrix revisited. Trends Microbiol. 2005, 13, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.M.; Facincani, A.P.; Ferreira, C.B.; Ferreira, R.M.; Ferro, M.I.T.; Gozzo, F.C.; de Oliveira, J.C.F.; Ferro, J.A.; Soares, M.R. Chemotactic signal transduction and phosphate metabolism as adaptive strategies during citrus canker induction by Xanthomonas citri. Funct. Integr. Genom. 2015, 2, 197–210. [Google Scholar] [CrossRef]
- Bashan, Y.; Diab, S.; Okon, Y. Survival of Xanthomonas campestris pv. vesicatoria in pepper seeds and roots in symptomless and dry leaves in non-host plants and in the soil. Plant Soil. 1982, 68, 161–170. [Google Scholar]
- Osdaghi, E.; Jones, J.B.; Sharma, A.; Goss, E.M.; Abrahamian, P.; Newberry, E.A.; Potnis, N.; Carvalho, R.; Choudhary, M.; Paret, M.L.; et al. A centenary for bacterial spot of tomato and pepper. Mol. Plant Pathol. 2021, 22, 1500–1519. [Google Scholar] [PubMed]
- European and Mediterranean Plant Protection Organization. PM 7/110 (1) Xanthomonas spp. (Xanthomonas euvesicatoria, Xanthomonas gardneri, Xanthomonas perforans, Xanthomonas vesicatoria) causing bacterial spot of tomato and sweet pepper. Bull. OEPP/EPPO Bull. 2013, 43, 7–20. [CrossRef]
- Young, A.J.; Marney, T.S.; Herrington, M.; Hutton, D.; Gomez, A.O.; Villiers, A.; Campbell, P.R.; Geering, A.D.W. Outbreak of angular leaf spot in an Australian strawberry germplasm collection. Australas. Plant Pathol. 2011, 40, 286–292. [Google Scholar]
- Bouzar, H.; Jones, J.B.; Stall, R.E.; Somodi, G.C.; Kelly, R.O.; Daouzli, N. Phenotypic characterization of Xanthomonas campestris pv. vesicatoria strains from the Caribbean and Central America. Phytopathology 1994, 84, 1069. [Google Scholar]
- Jones, J.B.; Somodi, G.C.; Scott, J.W. Increased ELISA sensitivity using a modified extraction buffer for detection of Xanthomonas campestris pv. vesicatoria in leaf tissue. J. Appl. Microbiol. 1997, 83, 397–401. [Google Scholar] [CrossRef]
- Massomo, S.M.S.; Nielsen, H.; Mabagala, R.B.; Mansfeld-Giese, K.; Hockenhull, J.; Mortensen, C.N. Identification and characterisation of Xanthomonas campestris pv. campestris strains from Tanzania by pathogenicity tests, Biolog, rep-PCR and fatty acid methyl ester analysis. Eur. J. Plant Pathol. 2003, 109, 775–789. [Google Scholar] [CrossRef]
- Bouzar, H.; Jones, J.B.; Minsavage, G.V.; Stall, R.E.; Scoot, J.W. Proteins unique to phenotypically distinct groups of Xanthomonas campestris pv. vesicatoria revealed by silver staining. Phytopathology 1994, 84, 39–44. [Google Scholar] [CrossRef]
- Jones, J.B.; Bouzar, H.; Stall, R.E.; Almira, E.C.; Roberts, P.D.; Bowen, B.W.; Sudberry, J.; Strickler, P.M.; Chun, J. Systematic analysis of xanthomonads (Xanthomonas spp.) associated with pepper and tomato lesions. Int. J. Syst. Evol. 2000, 50, 1211–1219. [Google Scholar] [CrossRef] [PubMed]
- Leite, R.P.; Minsavage, G.V.; Bonas, U.; Stall, R.E. Detection and identification of phytopathogenic Xanthomonas strains by amplification of DNA sequences related to the hrp genes of Xanthomonas campestris pv. vesicatoria. Appl. Environ. Microbiol. 1994, 60, 1068–1077. [Google Scholar] [CrossRef] [PubMed]
- Moretti, C.; Amatulli, M.T.; Buonaurio, R. PCR-based assay for the detection of Xanthomonas euvesicatoria causing pepper and tomato bacterial spot. Lett. Appl. Microbiol. 2009, 49, 466–471. [Google Scholar] [CrossRef]
- Park, D.S.; Shim, J.K.; Kim, J.S.; Lim, C.K.; Shrestha, R.; Hahn, J.H.; Kim, H.G. Sensitive and specific detection of Xanthomonas campestris pv. vesicatoria by PCR using pathovar-specific primers based on rhs family gene sequences. Microbiol. Res. 2009, 164, 36–42. [Google Scholar] [CrossRef]
- Koenraadt, H.; Betteray, B.; Germain, R.; Hiddink, G.; Jones, J.B.; Oosterhof, J.; Rijlaarsdam, A.; Roorda, P.; Woudt, B. Development of specific primers for the molecular detection of bacterial spot of pepper and tomato. Acta Hortic. 2009, 808, 99–102. [Google Scholar] [CrossRef]
- Strayer-Scherer, A.; Jones, J.B.; Paret, M.L. Recombinase polymerase amplification assay for field detection of tomato bacterial spot pathogens. Phytopathology 2019, 109, 690–700. [Google Scholar] [CrossRef]
- Larrea-Sarmiento, A.; Dhakal, U.; Boluk, G.; Fatdal, L.; Alvarez, A.; Strayer-Scherer, A.; Paret, M.; Jones, J.; Jenkins, D.; Arif, M. Development of a genome-informed loop-mediated isothermal amplification assay for rapid and specific detection of Xanthomonas euvesicatoria. Sci. Rep. 2018, 8, 14298. [Google Scholar] [CrossRef]
- Stehlikova, D.; Beran, P.; Cohen, S.P.; Curn, V. Development of real-time and colorimetric loop mediated isothermal amplification assay for detection of Xanthomonas gardneri. Microorganisms 2020, 8, 1301. [Google Scholar] [CrossRef]
- Pecenka, J.; Kacanova, M.; Baranek, M.; Gazdik, F.; Ragasova, L.; Penazova, E.; Cechova, J.; Beran, P.; Eichmeier, A. Species-specific PCR primers for the detection of poorly distinguishable Xanthomonas euvesicatoria. Crop Prot. 2020, 127, 104978. [Google Scholar] [CrossRef]
- Liu, H.; Dong, C.; Zhao, T.; Han, J.; Wang, T.; Wen, X.; Huang, Q. Functional analysis of the ferric uptake regulator gene fur in Xanthomonas vesicatoria. PLoS ONE 2016, 11, e0149280. [Google Scholar] [CrossRef]
- Beran, P.; Mraz, I. Species-specific PCR primers for detection of Xanthomonas vesicatoria. Crop Prot. 2012, 43, 213–215. [Google Scholar] [CrossRef]
- Albuquerque, P.; Caridade, C.M.R.; Rodrigues, A.S.; Marcal, A.R.S.; Cruz, J.; Cruz, L.; Santos, C.L.; Mendes, M.V.; Tavares, F. Evolutionary and experimental assessment of novel markers for detection of Xanthomonas euvesicatoria in plant samples. PLoS ONE 2012, 7, e37836. [Google Scholar] [CrossRef]
- Boudon, S.; Manceau, C.; Notteghem, J. Structure and origin of Xanthomonas arboricola pv. pruni populations causing bacterial spot of stone fruit trees in Western Europe. APS 2005, 95, 1081–1088. [Google Scholar] [CrossRef]
- Hamza, A.A.; Robene-Soustrade, I.; Jouen, E.; Lefeuvre, P.; Chiroleu, F.; Fisher-Le Saux, M.; Gagnevin, L.; Provust, O. Multilocus sequence analysis and amplified fragment length polymorphism-based characterization of xanthomonads associated with bacterial spot of tomato and pepper and their relatedness to Xanthomonas species. Syst. Appl. Microbiol. 2012, 35, 183–190. [Google Scholar] [CrossRef]
- Dutta, B.; Gitaitis, R.; Sanders, H.; Booth, C.; Smith, S.; Langston, D.B., Jr. Role of blossom colonization in pepper seed infestation by Xanthomonas euvesicatoria. Phytopathology 2014, 104, 232–239. [Google Scholar] [CrossRef]
- Dutta, B.; Langston, D.B.; Luo, X.; Carlson, S.; Kichler, J.; Gitaitis, R. A risk assessment model for bacterial leaf spot of pepper (Capsicum annuum), caused by Xanthomonas euvesicatoria, based on concentrations of macronutrients, micronutrients, and micronutrient ratios. Phytopathology 2017, 107, 1331–1338. [Google Scholar] [CrossRef]
- Johnson, G.F. The early history of copper fungicides. Agric. Hist. 1935, 9, 67–79. [Google Scholar]
- Conover, R.A.; Gerhold, N.R. Mixtures of copper and maneb or mancozeb for control of bacterial spot of tomato and their compatibility for control fungus diseases. Proc. Annu. Meet. Fla. State Hort. Soc. 1981, 94, 154–156. [Google Scholar]
- Marco, G.M.; Stall, R.E. Control of bacterial spot of pepper initiated by strains of Xanthomonas campestris pv. vesicatoria that differ in sensibility to copper. Plant Dis. 1983, 67, 779–781. [Google Scholar] [CrossRef]
- Li, J.; Wang, N. Foliar application of biofilm formation-inhibiting compounds enhances control of citrus canker caused by Xanthomonas citri subsp. citri. Phytopathology 2014, 104, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, S.; Huang, Y.; Jones, J. Evaluation of a small molecule compound 3-indolylacetonitrile for control of bacterial spot-on tomato. Crop Prot. 2019, 120, 7–12. [Google Scholar] [CrossRef]
- Qiao, K.; Liu, Q.; Huang, Y.; Xia, Y.; Zhang, S. Management of bacterial spot of tomato caused by copper-resistant Xanthomonas perforans using a small molecule compound carvacrol. Crop Prot. 2020, 132, 105114. [Google Scholar] [CrossRef]
- Abbasi, P.A.; Soltani, N.; Cuppels, D.A.; Lazarovits, G. Reduction of bacterial spot disease severity on tomato and pepper plants with foliar applications of ammonium lignosulfonate and potassium phosphate. Plant Dis. 2002, 86, 1232–1236. [Google Scholar]
- Carvalho, R.; Duman, K.; Jones, J.B.; Paret, M.L. Bactericidal activity of copper-ziinc hybrid nanoparticles on copper-tolerant Xanthomonas perforans. Sci. Rep. 2019, 9, 20124. [Google Scholar] [CrossRef]
- Srivastava, V.; Deblais, L.; Kathayat, D.; Rotondo, F.; Helmy, Y.A.; Miller, S.A.; Rajashekara, G. Novel small molecule growth inhibitors of Xanthomonas spp. causing bacterial spot of tomato. Phytopathology 2021, 111, 940–953. [Google Scholar] [CrossRef]
- He, D.C.; He, M.H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological control of plant diseases: An evolutionary and eco-economic consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef]
- Pajcin, I.; Vlajkov, V.; Frohme, M.; Grebinyk, S.; Grahovac, M.; Mojicevic, M.; Grahovac, J. Pepper bacterial spot control by Bacillus velezensis: Bioprocess solution. Microorganisms 2020, 8, 1463. [Google Scholar] [CrossRef]
- Yi, H.; Yang, J.; Ryu, C. ISR meets SAR outside: Additive action of the endophyte Bacillus pumilus INR7 and the chemical inducer, benzothiadiazole, on induced resistance against bacterial spot in field-grown pepper. Front. Plant Sci. 2013, 14, 122. [Google Scholar] [CrossRef]
- Da Silva, A.P.S.; Olivares, F.L.; Sudre, C.P.; Peres, L.E.P.; Canellas, N.A.; da Silva, R.M.; Cozzolino, V.; Canellas, L.P. Attenuations of bacterial spot disease Xanthomonas euvesicatoria on tomato plants treated with biostimulants. Chem. Biol. Technol. 2021, 8, 42. [Google Scholar] [CrossRef]
- Le, K.D.; Kim, J.; Yu, N.H.; Kim, B.; Lee, C.W.; Kim, J. Biological control of tomato bacterial wilt, kimchi cabbage soft rot, and red pepper bacterial leaf spot using Paenibacillus elgii JCK-5075. Front. Plant Sci. 2020, 11, 775. [Google Scholar] [CrossRef] [PubMed]
- Moss, W.P.; Byrne, J.M.; Campbell, H.L. Biological control of bacterial spot of tomato using hrp mutants of Xanthomonas campestris pv. vesicatoria. Biol. Control. 2007, 41, 199–206. [Google Scholar] [CrossRef]
- Moineau, S. Bacteriophage. In Brenner’s Encyclopedia of Genetics, 2nd ed.; Maloy, S., Hughes, K., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 280–283. [Google Scholar]
- Flaherty, J.E.; Jones, J.B.; Harbaugh, B.K.; Somodi, G.C.; Jackson, L.E. Control of bacterial spot on tomato in the greenhouse and field with H-mutant bacteriophages. J. Am. Soc. Hortic. Sci. 2000, 35, 882–884. [Google Scholar] [CrossRef]
- Gasic, K.; Ivanovic, M.M.; Ignjatov, M.; Calic, A.; Obradovic, A. Isolation and characterization of Xanthomonas euvesicatoria bacteriophages. Plant Pathol. J. 2011, 93, 415–423. [Google Scholar]
- Gasic, K.; Kuzmanovic, N.; Ivanovic, M.; Prokic, A.; Sevic, M.; Obradovic, A. Complete genom of the Xanthomonas euvesicatoria specific bacteriophage KФ1, its survival and potential in control of pepper bacterial spot. Front. Microbiol. 2018, 9, 2021. [Google Scholar] [CrossRef]
- Solis-Sanchez, G.A.; Quinones-Aguilar, E.E.; Fraire-Velasquez, S.; Vega-Arreguin, J.; Rincon-Enriquez, G. Complete genome sequence of XaF13, a novel bacteriophage of Xanthomonas vesicatoria from Mexico. Microbiol. Resour. Announc. 2020, 9, e01371-19. [Google Scholar] [CrossRef]
- Rios-Sandoval, M.; Quinones-Aguilar, E.E.; Solis-Sanchez, G.A.; Enriquez-Vara, J.N.; Rincon-Enriquez, G. Complete genome sequence of Xanthomonas vesicatoria bacteriophage ФXaF18, a contribution to the biocontrol of bacterial spot of pepper in Mexico. Microbiol. Resour. Announc. 2020, 9, e00213-20. [Google Scholar] [CrossRef]
- Omnilytics. Find a Natural, Safe, organic, Eco-Friendly, and Effective Pesticide. Available online: https://www.agriphage.com/product-info/ (accessed on 10 January 2022).
- Sevic, M.; Gasic, K.; Ignjatov, M.; Mijatovic, M.; Prokic, A.; Obradovic, A. Integration of biological and conventional treatments in control of pepper bacterial spot. Crop Prot. 2019, 119, 46–51. [Google Scholar] [CrossRef]
- Restrepo, M.; Babu, B.L.; Reyes, L.F.; Chalmers, J.D.; Soni, N.J.; Sibila, O.; Faverio, P.; Cilloniz, C.; Rodriguez-Cintron, W.; Aliberti, S. Burden and risk factors for Pseudomonas aeruginosa community-acquired pneumonia: A multinational point prevalence study of hospitalised patients. Eur. Respir. J. 2018, 52, 1701190. [Google Scholar] [CrossRef]
- Keswani, C.; Prakash, O.; Bharti, N.; Vílchez, J.I.; Sansinenea, E.; Lally, R.D.; Borriss, R.; Singh, S.P.; Gupta, V.K.; Fraceto, L.F.; et al. Re-addressing the biosafety issues of plant growth promoting rhizobacteria. Sci. Total Environ. 2019, 690, 841–852. [Google Scholar] [CrossRef]
- Vilchez, J.I.; Navas, A.; Gonzales-Lopez, J.; Arcos, S.C.; Manzanera, M. Biosafety test for plant growth-promoting bacteria: Proposed environmental and human safety index (EHSI) protocol. Front. Microbiol. 2016, 6, 1514. [Google Scholar] [CrossRef] [PubMed]
- Pena, R.T.; Blasco, L.; Ambroa, A.; González-Pedrajo, B.; Fernández-García, L.; López, M.; Bleriot, I.; Bou, G.; García-Contreras, R.; Wood, T.K.; et al. Relationship between quorum sensing and secretion systems. Front. Microbiol. 2019, 10, 1100. [Google Scholar] [CrossRef] [PubMed]
- Timilsina, S.; Potnis, N.; Newberry, E.A.; Liyanapathiranage, P.; Iruegas-Bocardo, F.; White, F.F.; Goss, E.M.; Jones, J.B. Xanthomonas diversity, virulence and plant–pathogen interactions. Nat. Rev. Microbiol. 2020, 18, 415–427. [Google Scholar] [CrossRef]
- Abrahamian, P.; Timilsina, S.; Minsavage, G.V.; Suhmita, K.C.; Goss, E.M.; Jones, J.B.; Vallad, G.E. The type III effector AvrBst enhance Xanthomonas perforans fitness in field-grown tomato. Phytopathology 2018, 108, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Potnis, N.; Krasileva, K.; Chow, V.; Almeida, N.F.; Patil, P.B.; Ryan, R.P.; Sharlach, M.; Berhlau, F.; Dow, J.M.; Momol, M.T.; et al. Comparative genomics reveals diversity among xanthomonads infecting tomato and pepper. BMC Genom. 2011, 12, 146. [Google Scholar] [CrossRef]
- Liyanapathiranage, P.; Wagner, N.; Avram, O.; Pupko, T.; Potnis, N. Phylogenetic distribution and evolution of type VI secretion system in the Genus Xanthomonas. Front. Microbiol. 2022, 13, 840308. [Google Scholar] [CrossRef]
- Sole, M.; Scheibner, F.; Hoffmeister, A.; Hartman, N.; Hause, G.; Rother, A.; Jordan, M.; Lautier, M.; Arlat, M.; Buttner, D. Xanthomonas campestris pv. vesicatoria secretes proteases and xylanases via the Xps type II secretion system and outer membrane vesicles. J. Bacteriol. 2015, 197, 2879–2893. [Google Scholar] [CrossRef]
- Szczesny, R.; Jordan, M.; Schramm, C.; Schulz, S.; Cogez, V.; Bonas, U.; Buttner, D. Functional characterization of the Xcs and Xps type II secretion systems from the plant pathogenic bacterium Xanthomonas campestris pv. vesicatoria. New Pathologist. 2010, 187, 983–1002. [Google Scholar] [CrossRef]
- Barak, J.D.; Vancheva, T.; Lefeuvre, P.; Jones, J.B.; Timilsina, S.; Minsavage, G.V.; Vallad, G.E.; Koebnik, E. Whole-genome sequences of Xanthomonas euvesicatoria strains clarify taxonomy and reveal a stepwise erosion of Type 3 Effectors. Front. Plant Sci. 2016, 7, 1805. [Google Scholar] [CrossRef]
- Buttner, D.; Bonas, U. Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol.-Microbiol. Rev. 2010, 34, 107–133. [Google Scholar] [CrossRef]
- Lu, H.; Patil, P.; Sluys, M.V.; White, F.F.; Ryan, R.P.; Dow, J.M.; Rabinowicz, P.; Salzberg, S.L.; Leach, J.E.; Sonti, R.; et al. Acquisition and evolution of plant pathogenesis-associated gene clusters and candidate determinants of tissue-specificity in Xanthomonas. PLoS ONE 2008, 3, e3828. [Google Scholar] [CrossRef]
- Dow, J.M.; Feng, J.; Barber, C.E.; Tang, J.; Daniels, M.J. Novel genes involved in the regulation of pathogenicity factor production within the rpf gene cluster of Xanthomonas campestris. Microbiology 2000, 146, 885–891. [Google Scholar] [CrossRef][Green Version]
- Teper, D.; Salomon, D.; Sunitha, S.; Kim, J.; Mudgett, M.B.; Sessa, G. Xanthomonas euvesicatoria type III effector XopQ interacts with tomato and pepper 13-4-4 isoforms to suppress effector-triggered immunity. Plant J. 2014, 77, 297–309. [Google Scholar] [CrossRef]
- Marois, E.; den Ackerveken, G.V.; Bonas, U. The Xanthomonas type III effector protein AvrBs3 modulates plant gene expression and induces cell hypertrophy in the susceptible host. Mol. Plant Microbe Interact. 2002, 15, 637–646. [Google Scholar] [CrossRef]
- Szurek, B.; Rossier, O.; Hause, G.; Bonas, U. Type III-dependent translocation of the Xanthomonas AvrBs3 protein into the plant cell. Mol. Microbiol. 2002, 46, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Figueiredo, F.; Jones, J.; Wang, N. HrpG and HrpX play global roles in coordinating different virulence traits of Xanthomonas axonopodis pv.citri. Mol. Plant Microbe Interact. 2011, 24, 649–661. [Google Scholar] [CrossRef]
- Popov, G.; Fraiture, M.; Brunner, F.; Sessa, G. Multiple Xanthomonas euvesicatoria type III effectors inhibit flg22-triggered immunity. Mol. Plant-Microbe. Interact. 2016, 29, 651–660. [Google Scholar] [CrossRef]
- Raffeiner, M.; Ustun, S.; Guerra, T.; Spinti, D.; Fitzner, M.; Sonnewald, S.; Baldermann, S.; Bornke, F. The Xanthomonas type-III effector XopS stabilizes CaWRKY40a to regulate defense responses and stomatal immunity in pepper (Capsicum annuum). Plant. Cell 2022, 34, 1684–1708. [Google Scholar] [CrossRef] [PubMed]
- Schulze, S.; Kay, S.; Buttner, D.; Egler, M.; Eschen-Lippold, L.; Hause, G.; Kruger, A.; Lee, J.; Muller, O.; Scheel, D.; et al. Analysis of new type III effectors from Xanthomonas uncovers XopB and XopS as suppressors of plant immunity. New Phytol. 2012, 195, 894–911. [Google Scholar] [CrossRef] [PubMed]
- Szczesny, R.; Büttner, D.; Escolar, L.; Schulze, S.; Seiferth, A.; Bonas, U. Suppression of the AvrBs1-specific hypersensitive response by the YopJ effector homolog AvrBsT from Xanthomonas depends on a SNF1-related kinase. New Phytolog. 2010, 187, 1058–1074. [Google Scholar] [CrossRef]
- Zhao, B.; Dahlbeck, D.; Krasileva, K.V.; Fong, R.W.; Staskawicz, B.J. Computational and biochemical analysis of the Xanthomonas effector avrbs2 and its role in the modulation of Xanthomonas type three effector delivery. PLoS Pathog. 2011, 7, e1002408. [Google Scholar] [CrossRef]
- Ki, J.; Stork, W.; Mudget, M.B. Xanthomonas type iii effector xopd desumoylates tomato transcription factor slerf4 to suppress ethylene responses and promote pathogen growth. Cell Host Microbe 2013, 13, 143–154. [Google Scholar] [CrossRef]
- Teper, D.; Girija, A.M.; Bosis, E.; Popov, G.; Savidor, A.; Sessa, G. The Xanthomonas euvesicatoria type III effector XopAU is an active protein kinase that manipulates plant MAP kinase signaling. PLoS Pathog. 2018, 14, e1006880. [Google Scholar] [CrossRef] [PubMed]
- Priller, J.P.R.; Reid, S.; Konein, P.; Dietrich, P.; Sonnewald, S. The Xanthomonas campestris pv. vesicatoria Type-3 Effector XopB Inhibits plant defence responses by interfering with ROS production. PLoS ONE 2016, 11, e0159107. [Google Scholar] [CrossRef]
- Kim, J.; Li, X.; Roden, J.A.; Taylor, K.W.; Aakre, C.D.; Su, B.; Lalonde, S.; Kirik, A.; Chen, Y.; Baranage, G.; et al. Xanthomonas T3S effector Xopn suppresses pamp-triggered immunity and interacts with a tomato atypical receptor-like kinase and TFT1. Plant Cell. 2009, 21, pp. 1305–1323. Available online: www.plantcell.org/cgi/doi/10.1105/tpc.108.063123 (accessed on 1 April 2022).
- Bartetzko, V.; Sonnewald, S.; Vogel, F.; Hartner, K.; Stadler, R.; Hammes, U.Z.; Bornke, F. The Xanthomonas campestris pv. vesicatoriaType III Effector Protein XopJ Inhibits Protein Secretion: Evidence for Interference with Cell Wall–Associated Defense Responses. Mol. Plant Microbe Interact. 2009, 22, 655–664. [Google Scholar] [CrossRef]
- Kousik, C.S.; Ritchie, D.F. Response of bell pepper cultivars to bacterial spot pathogen races that individually overcome major resistance genes. Plant Dis. 1998, 82, 181–186. [Google Scholar] [CrossRef]
- Horvath, D.M.; Pauly, M.H.; Hutton, S.F.; Vallad, G.E.; Scott, J.W.; Jones, J.B.; Stall, R.E.; Dahlbeck, D.; Staskawicz, B.J.; Tricoli, D.; et al. The pepper BS2 gene confers effective field resistance to bacterial leaf spot and yield enhancement in Florida tomatoes. Acta Hortic. 2015, 1069, 47–51. [Google Scholar] [CrossRef]
- Tai, T.H.; Dahlbeck, D.; Clark, E.T.; Gajiwala, P.; Pasion, R.; Whalen, M.C.; Stall, R.E.; Staskawicz, B.J. Expression of the Bs2 pepper gene confers resistance to bacterial spot disease in tomato. Proc. Natl. Acad. Sci. USA 1999, 96, 14153–14158. [Google Scholar] [CrossRef]
- Jordan, T.; Romer, P.; Meyer, A.; Szczesny, R.; Pierre, M.; Piffanelli, P.; Bendahmane, A.; Bonas, U.; Lahaye, T. Physical delimitation of the pepper Bs3 resistance gene specifying recognition of the AvrBs3 protein from Xanthomonas campestris pv. vesicatoria. Theor. Appl. Genet. 2006, 113, 896–905. [Google Scholar] [CrossRef]
- Adhikari, P.; Adhikari, T.B.; Louws, F.J.; Panthee, D.R. Advances and challenges in bacterial spot resistance breeding in tomato (Solanum lycopersicum L.). Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef]
- Vallejos, C.E.; Jones, V.; Stall, R.E.; Jones, J.B.; Minsavage, G.V.; Schultz, D.C.; Rodrigues, E.; Olsen, L.E.; Mazourek, M. Characterization of two recessive gene controlling resistance to all races of bacterial spot in peppers. Theor. Appl. Genet. 2010, 121, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Hwang, B.K. An important role of the pepper phenylalanine ammonia-lyase gene (PAL1) in salicylic acid-dependent signalling of the defence response to microbial pathogens. J. Exp. Bot. 2014, 65, 2295–2306. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Kim, S.Y.; Paek, K.H.; Choi, D.; Park, J.M. Suppression of a CaCYP1, a novel cytochrome P450 gene, compromises the basal pathogen defense response of pepper plants. Biochem. Biophys. Res. Commun. 2006, 345, 638–645. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Hwang, B.K. The pepper receptor-like cytoplasmic protein kinase CaPIK1 is involved in plant signalling of defence and cell-death responses. Plant J. 2011, 66, 642–655. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Races | References |
|---|---|---|
| X. euvesicatoria pv. euvesicatoria | 0–10 | [34,35,36,37,38,39,40] |
| X. euvesicatoria pv. perforans | ||
| X.hortorum pv. gardneri | ||
| X. vesicatoria |
| X. euvesicatoria pv. euvesicatoria | X. euvesicatoria pv. perforans | X. hortorum pv. gardneri | X. vesicatoria | |
|---|---|---|---|---|
| Group | A | C | D | B |
| Reference strains | ATCC11633T (NCPPB2968 | ATCC BAA-983T (NCPPB4321) | ATCC19865T (NCPPB881) | ATCC35937T (NCPPB422) |
| Protein unique size (kDA) | 32–35 | 25–27 | 25–27 | 25–27 |
| Amylolytic activity | - | + | - | + |
| Pectate hydrolysis | - | + | - | + |
| mAb * reaction | 1, 21 | 30 | 8 | 8, 15 |
| Utilization of: | ||||
| Dextrin | + | + | - | + |
| Glycogen | + | v | - | v |
| N-acetyl-D-glucosamine | + | + | - | v |
| D-galactose | + | + | - | v- |
| Gentibiose | + | + | - | v |
| α-D-lactose lactulose | v | + | - | v- |
| Acetic acid | v | + | - | - |
| Cis-aconitic acid | + | v | - | - |
| Malonic acid | + | + | - | v |
| Propionic acid | v- | + | - | v |
| D-alanine | v | + | - | v |
| Glycyl-L-aspartic acid | - | + | - | v- |
| L-threonine | v | + | - | v- |
| Bacterium | Target | (Assay)Primer Name | Sequence 5′-3′ | Amplicon Size | Ref. |
|---|---|---|---|---|---|
| (PCR) | |||||
| X. euvesciatoria pv. euvesicatoria | Zn-dependent oxidoreductase | ZnDoF | GGTGACAAACCGTCAGGAATAG | 100 bp | [69] |
| ZnDoR | CGCACTGGCACGTTATCA | ||||
| (LAMP) | |||||
| X. hortorum pv. gardneri | hrpB | F3 | CGGGGTGCAGGTCAGC | a/n | [68] |
| B3 | ACCGGCACCGCCAAG | ||||
| FIP | CCACCTCGGCACGTTGCAGGCGAGGTATGCGAGTTGC | ||||
| BIP | GCCGCCATCTCGCCTTGCGCCCCGATCCGATCACG | ||||
| LB | CGAGCTGGTGGGCTTGT | ||||
| (RPA) | |||||
| X. hortorum pv. gardneri,X. euvesicatoria pv. euvesicatoria,X. euvesicatoria pv. perforans,X. vesicatoria | hrcN | X. hortorum pv. gardneri exo probe | TCTCGCCTTGCTGGCGCCGTTTGGCGAGC-dT-FAM-HG-dT-BHQ1-GGGCTTGTCGCGC | a/n | [66] |
| XGF | GCACGCTGTTGCAACGTGCCGAGGTGGTGG | ||||
| XGR | CGTCCGCCGGCTCACCCAGGCCATCGAGTA | ||||
| X. euvesicatoria pv. euvesicatoria pathovar exo probe | CGGGCAAGGCGCAATCGCCTGTGACACC-dT-FAM-GHG-dT-BHQ1-GCCGATCCAGGCG | ||||
| X. euvesicatoria pv. perforans exo probe | CGGGCAAGGAGCCATCGCCTGTGACACC-dT-FAM-GHG-dT-BHQ1-GCCGATCCAGGCG | ||||
| FP1 | GTTGGACCGGCCTTGCTGGGCCGCGTGCTC | ||||
| RP1 | GTCGGCATGGGGTGTTCGATCAGCCGCCGA | ||||
| X. vesicatoria exo probe | GCCTTGCTGGCGCCGTTTGGCGAATTGG-dT-FAM-GHGG-dT-BHQ1-TGTCGCGCGAAA | ||||
| XVF | ATGGCACGCTGTTGCAGCGCGCCGAGGTGGTGG | ||||
| XVR | GCACCGCCAATGGGCGACCGGATCCGATCA | ||||
| (LAMP) | |||||
| X. euvesicatoria pv. euvesicatoria | ATP-dependent DNA helicase (recG) | XeRec-F3 | CCATGTAGGGCTTGTTGACG | a/n | [67] |
| XeRec-B3 | GGTGGTCGCATCTTCATTGG | ||||
| XeRec-FIP | ACCCGCTCACGGAAAACGTGCC-TTCAGCGATGGACAGC | ||||
| XeRec-BIP | GAGGCCACGTTGGCGATGAG-GTGAACGACGACGGTTCG | ||||
| XeRec-LF | ACCCGGCAGGCACGGTGCT | ||||
| XeRec-LB | AGCAACGTCGGCGCCGGATA | ||||
| (PCR) | |||||
| X. vesicatoria | Ferric uptake regulator (fur) | fur1 | GAATTCATCGGTCCTGGGAGTC | 1572 bp | [70] |
| fur2 | AAGCTTCGGCGTGGAAGTGA | ||||
| (PCR) | |||||
| X. vesicatoria | ATP synthase | XV1F | CAGTCCTCCAGCACCGAAC | 365 bp | [71] |
| XV1R | TCTCGTCGCGGAAGTACTCA | ||||
| (PCR) | |||||
| X. euvesicatoria pv. euvesicatoria | XCV0215 | XV4F | ATCAATGAGCCTTGGGATGTGACGA | 230 bp | [72] |
| XV4R | GCATAGGTCAGGGCTTGCTTTAGCG | ||||
| XCV0217 | XV5F | GCCTAAGAATGCGGAGCCTTGGCT | 210 bp | ||
| XV5R | ATCTTCGGAGGCGTGTACGGCGTA | ||||
| XCV3374 | XV6F | AATGTGATCTTTTTGACGAGCGCA | 169 bp | ||
| XV6R | GCAACCTCGTCTGTTTCATTCTCAT | ||||
| XCV3818 | XV7F | CATTTCCATCACGCGTCATGCCG | 179 bp | ||
| XV7R | TGTTGCTCGGAATCGGTGGACCACC | ||||
| XCV3902 | XV8F | TGTCTCAAGCCGCGCTTAAC | 123 bp | ||
| XV8R | AACCGAAGAACAGGAACGATCTC | ||||
| XCV0217 | XV10F | GCGTTGGCACAATGTCGACC | 805 bp | ||
| XV10R | TTCGTCTAGCTCTCCACGGACCTG | ||||
| XCV0655 | XV11F | GCGACTGCGCTGGTATGAGCTCTA | 631 bp | ||
| XV11R | TGGCGTGTAGACACCCACTGTCGAG | ||||
| XCV1116 | XV12F | GGAGCCGTCTGCTGGTAAGCTGAT | 638 bp | ||
| XV12R | GCTGTATCAAACGAGATCCGCTG | ||||
| XV1853 | XV14F | TGGTTCACGTCATCGTTGTCGGA | 713 bp | ||
| XV14R | TAGAGCTCGCTCAAAGCCCTTCGG | ||||
| (MLSA-AFLP) | |||||
| X. euvesicatoria pv. euvesicatoria | atpD | atpD-F | GGGCAAGATCGTTCAGAT | 756 bp | [73,74] |
| atpD-R | GCTCTTGGTCGAGGTGAT | ||||
| (AFLP-PCR) | |||||
| X. euvesicatoria pv. euvesicatoria | Bs-XeF | CATGAAGAACTCGGCGTATCG | 173 bp | [65] | |
| Bs-XeR | GTCGGACATAGTGGACACATAC | ||||
| X. vesicatoria | Bs-XvF | CCATGTGCCGTTGAAATACTTG | 138 bp | ||
| Bs-XvR | ACAAGAGATGTTGCTATGATTTGC | ||||
| X. hortorum pv. gardneri | Bs-XgF | TCAGTGCTTAGTTCCTCATTGTC | 154 bp | ||
| Bs-XgR | TGACCGATAAAGACTGCGAAAG | ||||
| X. euvesicatoria pv. perforans | Bs-XpF | GTCGTGTTGATGGAGCGTTC | 197 bp | ||
| Bs-XpR | GTGCGAGTCAATTATCAGAATGTGG | ||||
| (PCR) | |||||
| X. euvesicatoria pv. euvesicatoria | rhs family | XCVF | AGAAGCAGTCCTTGAAGGCA | 517 bp | [64] |
| XCVR | AATGACCTCGCCAGTTGAGT | ||||
| (PCR) | |||||
| X. euvesicatoria pv. euvesicatoria | hypothetical protein XCV3137 | Xeu2.4 | CTGGGAAACTCATTCGCAGT | 208 bp | [63] |
| Xeu2.5 | TTGTGGCGCTCTTATTTCCT | ||||
| (PCR) | |||||
| X. euvesicatoria pv. euvesicatoria | hypersensitive reaction and pathogenicity (hrp) | RST9 | CACTATGCAATGACTG | 355 bp | [62] |
| RST10 | AATACGCTGGAACTGCTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Utami, D.; Meale, S.J.; Young, A.J. A Pan-Global Study of Bacterial Leaf Spot of Chilli Caused by Xanthomonas spp. Plants 2022, 11, 2291. https://doi.org/10.3390/plants11172291
Utami D, Meale SJ, Young AJ. A Pan-Global Study of Bacterial Leaf Spot of Chilli Caused by Xanthomonas spp. Plants. 2022; 11(17):2291. https://doi.org/10.3390/plants11172291
Chicago/Turabian StyleUtami, Desi, Sarah Jade Meale, and Anthony Joseph Young. 2022. "A Pan-Global Study of Bacterial Leaf Spot of Chilli Caused by Xanthomonas spp." Plants 11, no. 17: 2291. https://doi.org/10.3390/plants11172291
APA StyleUtami, D., Meale, S. J., & Young, A. J. (2022). A Pan-Global Study of Bacterial Leaf Spot of Chilli Caused by Xanthomonas spp. Plants, 11(17), 2291. https://doi.org/10.3390/plants11172291