
Exploring Relationships among Grapevine Chemical and Physiological Parameters and Leaf and Berry Mycobiome Composition

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
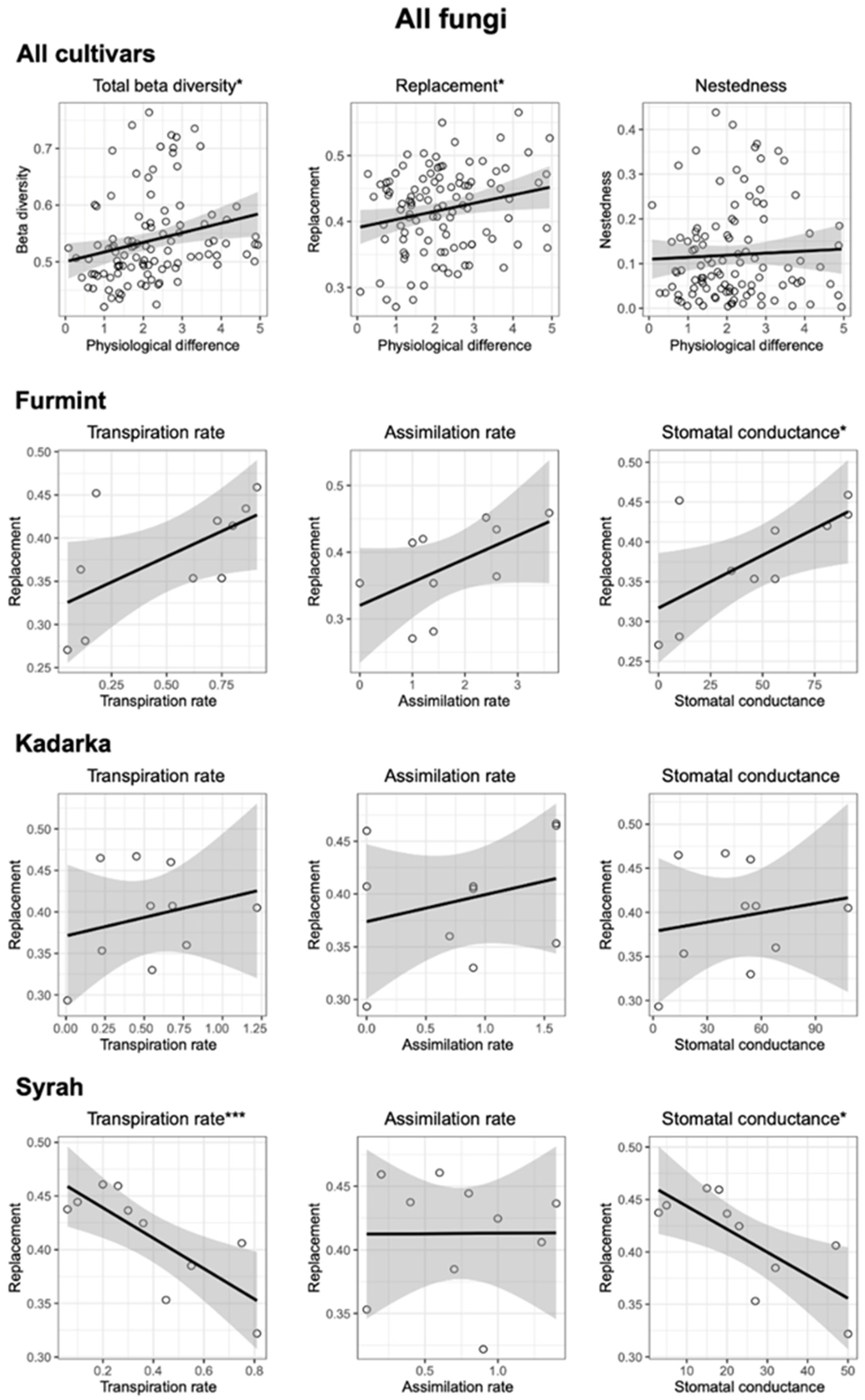
2.1. Meteorological and Physiological Data
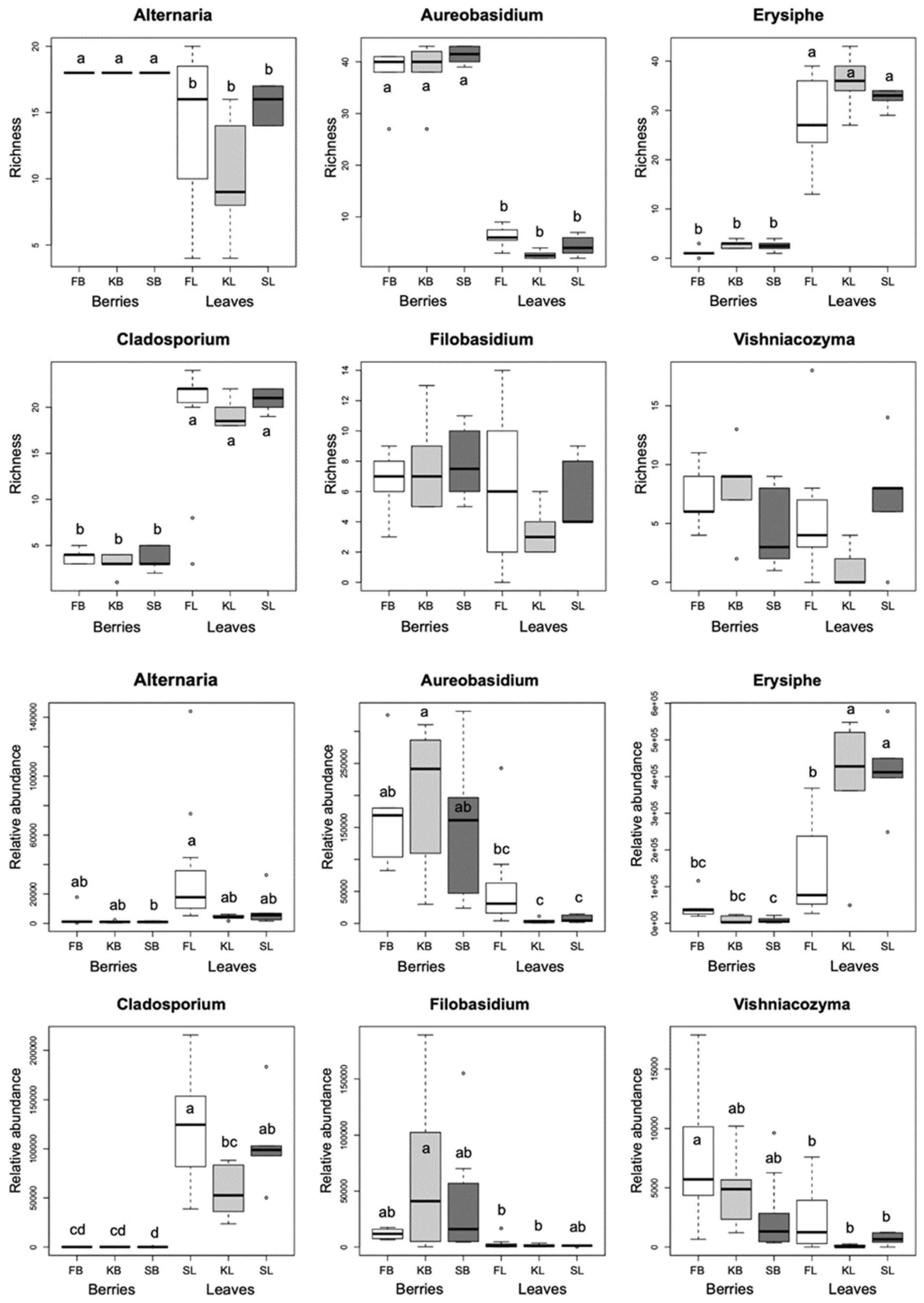
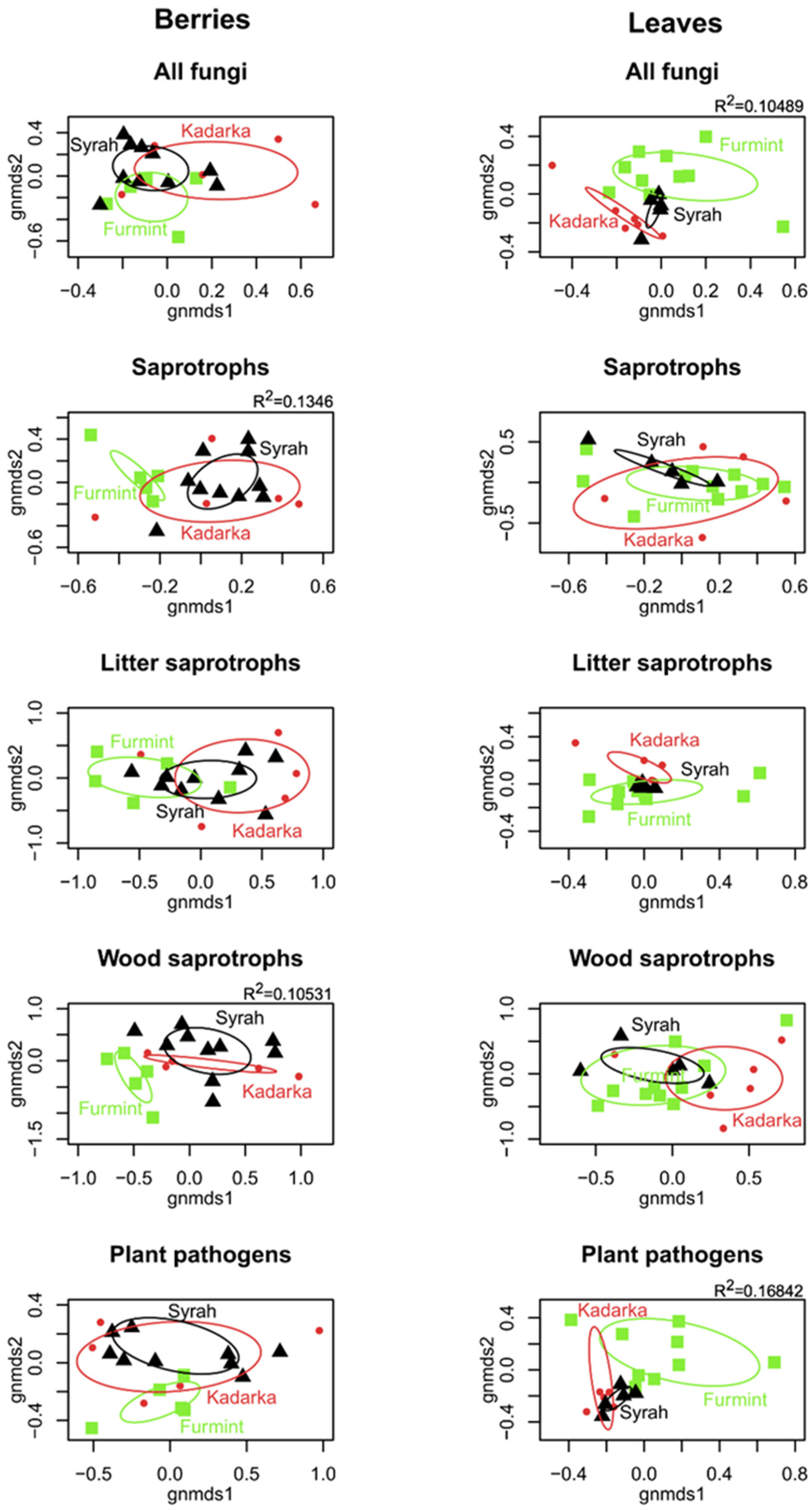
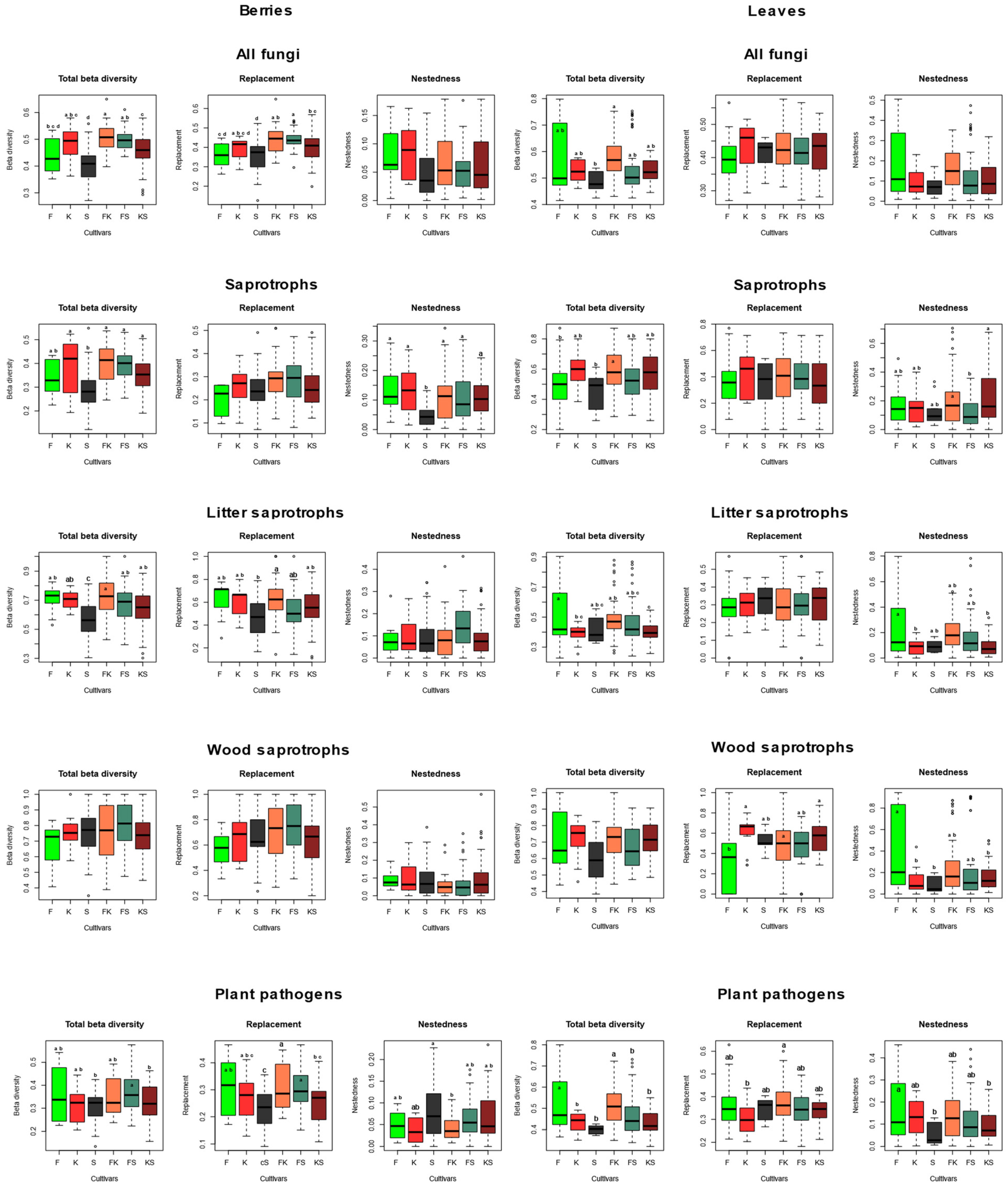
2.2. Grapevine Mycobiome
2.3. Chemical Parameters of Leaves and Berries
3. Discussion
4. Materials and Methods
4.1. Sampling Site
4.2. Leaf Gas-Exchange and Pre-Dawn Water Potential
4.3. Sample Collection and Metagenomic DNA Extraction
4.4. DNA Metabarcoding Data Analysis
4.5. Analytical Measurements
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Torregrosa, L.; Vialet, S.; Adivèze, A.; Iocco-Corena, P.; Thomas, M.R. Grapevine (Vitis vinifera L.). Methods Mol. Biol. 2015, 1224, 177–194. [Google Scholar] [PubMed]
- Pinto, C.; Pinho, D.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Unravelling the diversity of grapevine microbiome. PLoS ONE 2014, 9, e85622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, B.W.G.; Weingarten, E.A.; Jackson, C.R. The role of the phyllosphere microbiome in plant health hand function. Annu Plant Rev. Online 2018, 1, 533–556. [Google Scholar]
- van Niekerk, J.M.; Calitz, F.J.; Halleen, F.; Fourie, P.H. Temporal spore dispersal patterns of grapevine trunk pathogens in South Africa. Eur. J. Plant Pathol. 2010, 127, 375–390. [Google Scholar] [CrossRef]
- Zarraonaindia, I.; Owens, S.M.; Weisenhorn, P.; West, K.; Hampton-Marcell, J.; Lax, S.; Bokulich, N.A.; Mills, D.A.; Martin, G.; Taghavi, S.; et al. The soil microbiome influences grapevine-associated microbiota. mBio 2015, 6, e02527-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francesca, N.; Canale, D.E.; Settanni, L.; Moschetti, G. Dissemination of wine-related yeasts by migratory birds. Environ. Microbiol. Rep. 2012, 4, 105–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, C.E. Phyllosphere. In Encyclopedia of Life Sciences; Wiley: Chichester, UK, 2002. [Google Scholar] [CrossRef]
- Kinkel, L.L. Fungal community dynamics. In Microbial Ecology of Leaves; Andrews, J.H., Hirano, S.S., Eds.; Springer: New York, NY, USA, 1991; pp. 253–270. [Google Scholar]
- Pancher, M.; Ceol, M.; Corneo, P.E.; Longa, C.M.O.; Yousaf, S.; Pertot, I.; Campisano, A. Fungal endophytic communities in grapevines (Vitis vinifera L.) respond to crop management. Appl. Environ. Microbiol. 2012, 78, 4308–4317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perazzolli, M.; Antonielli, L.; Storari, M.; Puopolo, G.; Pancher, M.; Giovannini, O.; Pindo, M.; Pertot, I. Resilience of the natural phyllosphere microbiota of the grapevine to chemical and biological pesticides. Appl. Environ. Microbiol. 2014, 80, 3585–3596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setati, M.E.; Jacobson, D.; Bauer, F.F. Sequence-based analysis of the Vitis vinifera L. cv. Cabernet Sauvignon grape must mycobiome in three South African vineyards employing distinct agronomic systems. Front. Microbiol. 2015, 6, 1358. [Google Scholar] [CrossRef] [PubMed]
- Varanda, C.M.R.; Oliveira, M.; Materatski, P.; Landum, M.; Clara, M.I.E.; Félix, M.R. Fungal endophytic communities associated to the phyllosphere of grapevine cultivars under different types of management. Fungal Biol. 2016, 120, 1525–1536. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, L.E.; Miura, T.; Sánchez, R.; Barbosa, O. Effects of agricultural management on phyllosphere fungal diversity in vineyards and the association with adjacent native forests. PeerJ 2018, 6, e5715. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Santoni, S.; This, P.; Péros, J.P. Genotype-environment interaction shapes the microbial assemblage in grapevine’s phyllosphere and carposphere: An NGS approach. Microorganisms 2018, 6, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Gobbi, A.; Santoni, S.; Hansen, L.H.; This, P.; Péros, J.P. Assessing the impact of plant genetic diversity in shaping the microbial community structure of Vitis vinifera phyllosphere in the Mediterranean. Front. Life Sci. 2018, 11, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Diz, M.D.; Eichmeier, A.; Spetik, M.; Bujanda, R.; Díaz-Fernández, Á.; Díaz-Losada, E.; Gramaje, D. Grapevine pruning time affects natural wound colonization by wood-invading fungi. Fungal Ecol. 2020, 48, 100994. [Google Scholar] [CrossRef]
- Knapp, D.G.; Lázár, A.; Molnár, A.; Vajna, B.; Karácsony, Z.; Váczy, K.Z.; Kovács, G.M. Above-ground parts of white grapevine Vitis vinifera cv. Furmint share core members of the fungal microbiome. Environ. Microbiol. Rep. 2021, 13, 509–520. [Google Scholar] [CrossRef]
- Liu, D.; Howell, K. Community succession of the grapevine fungal microbiome in the annual growth cycle. Environ. Microbiol. 2021, 23, 1842–1857. [Google Scholar] [CrossRef] [PubMed]
- Geiger, A.; Karácsony, Z.; Golen, R.; Váczy, K.Z.; Geml, J. The compositional turnover of grapevine-associated plant pathogenic fungal communities is greater among intraindividual microhabitats and terroirs than among healthy and Esca-diseased plants. Phytopathology 2022, 112, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Ruppel, S. Progress in cultivation-independent phyllosphere microbiology. FEMS Microbiol. Ecol. 2014, 87, 2–17. [Google Scholar] [CrossRef]
- Balint, M.; Tiffin, P.; Hallström, B.; O’Hara, R.B.; Olson, M.S.; Fankhauser, J.D.; Piepenbring, M.; Schmitt, I. Host genotype shapes the foliar fungal microbiome of balsam poplar (Populus balsamifera). PLoS ONE 2013, 8, e53987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbonneau, A. Aspects qualitatifs. In Traite D’irrigation; Tiercelin, J.R., Ed.; Tec & Doc. Lavosier Ed: Paris, France, 1998; pp. 258–276. [Google Scholar]
- Deloire, A.; Carbonneau, A.; Wang, Z.; Ojeda, H. Vine and water: A short review. OENO One 2004, 38, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Adamo, I.; Ortiz-Malavasi, E.; Chazdon, R.; Chaverri, P.; ter Steege, H.; Geml, J. Soil fungal community composition correlates with site-specific abiotic factors, tree community structure, and forest age in regenerating tropical rainforests. Biology 2021, 10, 1120. [Google Scholar] [CrossRef] [PubMed]
- Fanin, N.; Lin, D.; Freschet, G.T.; Keiser, A.D.; Augusto, L.; Wardle, D.A.; Veen, G.F. Home-field advantage of litter decomposition: From the phyllosphere to the soil. New Phytol. 2021, 231, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, R.; Knorr, K.; Jørgensen, L.N.; O’Hanlon, K.A.; Nicolaisen, M. Host genotype is an important determinant of the cereal phyllosphere mycobiome. New Phytol. 2015, 207, 1134–1144. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Villarreal Ruiz, L.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Fungal biogeography. Global diversity and geography of soil fungi. Science 2014, 28, 346. [Google Scholar]
- Geml, J. Soil fungal communities reflect aspect-driven environmental structuring and vegetation types in a Pannonian forest landscape. Fungal Ecol. 2019, 39, 63–79. [Google Scholar] [CrossRef]
- Větrovský, T.; Kohout, P.; Kopecky, M.; Machac, A.; Man, M.; Bahnmann, B.D.; Brabcová, V.; Choi, J.; Mészárosová, L.; Human, Z.R.; et al. A meta-analysis of global fungal distribution reveals climate-driven patterns. Nat. Commun. 2019, 10, 5142. [Google Scholar] [CrossRef] [Green Version]
- Geml, J.; Arnold, A.E.; Semenova-Nelsen, T.A.; Nouhra, E.R.; Drechsler-Santos, E.R.; Góes-Neto, A.; Morgado, L.N.; Ódor, P.; Hegyi, B.; Grau, O.; et al. Community dynamics of soil-borne fungal communities along elevation gradients in neotropical and paleotropical forests. Mol. Ecol. 2022, 31, 2044–2060. [Google Scholar] [CrossRef]
- Gautier, A.T.; Merlin, I.; Doumas, P.; Cochetel, N.; Mollier, A.; Vivin, P.; Lauvergeat, V.; P’eret, B.; Cookson, S.J. Identifying roles of the scion and the rootstock in regulating plant development and functioning under different phosphorus supplies in grapevine. Environ. Exp. Bot. 2021, 185, 104405. [Google Scholar] [CrossRef]
- Verdenal, T.; Dienes-Nagy, Á.; Spangenberg, J.E.; Zufferey, V.; Spring, J.-L.; Viret, O.; Marin-Carbonne, J.; van Leeuwen, C. Understanding and managing nitrogen nutrition in grapevine: A review. OENO One 2021, 55, 1–43. [Google Scholar] [CrossRef]
- Lekberg, Y.; Arnillas, C.A.; Borer, E.T.; Bullington, L.S.; Fierer, N.; Kennedy, P.G.; Leff, J.W.; Luis, A.D.; Seabloom, E.W.; Henning, J.A. Nitrogen and phosphorus fertilization consistently favor pathogenic over mutualistic fungi in grassland soils. Nat. Commun. 2021, 12, 3484. [Google Scholar] [CrossRef]
- Simón, M.R.; Fleitas, M.C.; Castro, A.C.; Schierenbeck, M. How foliar fungal diseases affect nitrogen dynamics, milling, and end-use quality of wheat. Front. Plant Sci. 2020, 11, 569401. [Google Scholar] [CrossRef] [PubMed]
- Snoeijers, S.S.; Perez-Garcia, A.; Joosten, M.H.A.J.; De Wit, P.J.G.M. The effect of nitrogen on disease development and gene expression in bacterial and fungal plant pathogens. Eur. J. Plant Pathol. 2000, 106, 493–506. [Google Scholar] [CrossRef]
- Hoffland, E.; Jeger, M.J.; van Beusichem, M.L. Effect of nitrogen supply rate on disease resistance in tomato depends on the pathogen. Plant Soil 2000, 218, 239–247. [Google Scholar] [CrossRef]
- Neumann, S.; Paveley, N.D.; Beed, F.D.; Sylvester-Bradley, R. Nitrogen per unit leaf area affects the upper asymptote of Puccinia striiformis f. sp. tritici epidemics in winter wheat. Plant Pathol. 2004, 53, 725–732. [Google Scholar]
- Solomon, P.S.; Tan, K.C.; Oliver, R.P. The nutrient supply of pathogenic fungi; fertile field for study. Mol. Plant Pathol. 2003, 4, 203–210. [Google Scholar] [CrossRef]
- Zanetti, G.; Pandini, V. Ferredoxin. In Encyclopedia of Biological Chemistry II, 2nd ed.; Lennarz, W.J., Lane, M.D., Eds.; Academic Press, Elsevier Inc.: London, UK, 2013; pp. 296–298. [Google Scholar]
- Roy, A.; Kumar, A.; Baruah, D.; Tamuli, R. Calcium signaling is involved in diverse cellular processes in fungi. Mycology 2020, 12, 10–24. [Google Scholar] [CrossRef]
- Sala, F.; Blidariu, C. Macro- and micronutrient content in grapevine cordons under the influence of organic and mineral fertilization. Bulletin UASVM. Horticulture 2012, 69, 317–324. [Google Scholar]
- Capps, E.R. The Relationship between Mineral Nutrition and Late-Season Bunch Stem Necrosis of Cabernet Sauvignon (Vitis vinifera L.) Grapevines. Master’s Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VA, USA, 1999. [Google Scholar]
- Nagy, R.; Zsófi, Z.; Papp, I.; Földvári, M.; Kerényi, A.; Szabó, S. Evaluation of the relationship between soil erosion and the mineral composition of the soil: A case study from a cool climate wine region of Hungary. Carpathian J. Earth Environ. Sci. 2012, 7, 223–230. [Google Scholar]
- Kocsis, M.; Ayaydin, F.; Kőrösi, L.; Teszlák, P.; Radványi, L.; Jakab, G.; Hideg, É. Contrasting acclimation mechanisms of berry color variant grapevine cultivars (Vitis vinifera L. cv. Furmint) to natural sunlight conditions. Acta Physiol. Plant 2017, 39, 178. [Google Scholar] [CrossRef]
- Hajdu, E. Breeding of Kadarka. Horticulture 2010, 42, 27–37. [Google Scholar]
- Vouillamoz, J.F.; Grando, M.S. Genealogy of wine grape cultivars: ‘Pinot’ is related to ‘Syrah’. Heredity 2006, 97, 102–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Németh, M. Ampelográfiai Album—Termesztett Borszőlőfajták 1; Mezőgazdasági Kiadó: Budapest, Hungary, 1967. [Google Scholar]
- Bowers, J.E.; Siret, R.; Meredith, C.P.; This, P.; Boursiquot, J.M. A single pair of parents proposed for a group of grapevine varieties in Northeastern France. Acta Hortic. 2000, 528, 129–132. [Google Scholar] [CrossRef]
- Organisation Internationale de la Vigne et du Vin (OIV). OIV Descriptor List for Grape Varieties and Vitis Species, 2nd ed.; OIV: Paris, France, 2009. [Google Scholar]
- Scholander, P.F.; Hammel, H.T.; Bradstreet, E.D.; Hemmingsen, E.A. Sap pressure in vascular plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Ihrmark, K.; Bödeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandström-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region–evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef] [PubMed]
- White, T.M.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–321. [Google Scholar]
- Callahan, B.; McMurdie, P.; Rosen, M. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M. Towards a unified paradigm for sequence-based identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Põlme, S.; Abarenkov, K.; Nilsson, R.H.; Lindahl, B.D.; Clemmensen, K.; Kauserud, H.; Nguyen, N.; Kjoller, R.; Bates, S.T.; Baldrian, P. FungalTraits: A user-friendly traits database of fungi and fungus-like stramenopiles. Fungal Divers. 2020, 105, 1–16. [Google Scholar] [CrossRef]
- Sreenivasulu, V.; Kumar, N.S.; Dharmendra, V.; Asif, M.; Balaram, V.; Zhengxu, H.; Zhen, Z. Determination of Boron, Phosphorus, and Molybdenum Content in Biosludge Samples by Microwave Plasma Atomic Emission Spectrometry (MP-AES). Appl. Sci. 2017, 7, 264. [Google Scholar]
- Dharmendra, V. Total metals analysis of digested plant tissue using an Agilent 4200 Microwave. In Microwave Plasma Atomic Emission Spectroscopy (MP-AES). Application eHandbook; Agilent Technologies Plasma-AES: New Delhi, India, 2016; pp. 69–71. [Google Scholar]
- Liberato, C.G.; Barros, J.A.V.A.; Virgilio, A.; Machado, R.C.; Nogueira, A.R.A.; Nóbrega, J.A.; Schiavo, D. Determination of Macro and Micronutrients in Plants Using the Agilent 4200 MP AES; Agilent Technologies: New Delhi, India, 2017; pp. 1–5. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 25 February 2022).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Szoecs, E. Package ‘Vegan’. Community Ecology Package, Version 2. 2013. Available online: http://CRAN.R-project.org/package=vegan (accessed on 25 February 2022).
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species assemblages and indicator species: The need for a flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- De Cáceres, M.; Legendre, P.; Wiser, S.K.; Brotons, L. Using species combinations in indicator analyses. Methods Ecol. Evol. 2012, 3, 973–982. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ASV | p | SH | % | Matching Taxon | Class | Function |
|---|---|---|---|---|---|---|
| Berry | ||||||
| Furmint | ||||||
| ASV01064 | 0.016 | SH1646443.08FU | 95.4 | Agaricomycetes sp. | Agaricomycetes | |
| ASV00648 | 0.015 | SH1559713.08FU | 99.6 | Lophiostomataceae sp. | Dothideomycetes | |
| ASV00359 | 0.031 | SH1189171.08FU | 99.7 | Sporidiobolus sp. | Microbotryomycetes | MP |
| ASV00229 | 0.025 | SH1524265.08FU | 99.6 | Pleosporales sp. | Dothideomycetes | |
| ASV00731 | 0.023 | SH1517815.08FU | 99.7 | Hyphodontia quercina | Agaricomycetes | WOOD |
| ASV00470 | 0.032 | SH1185123.08FU | 99.7 | Rhodotorula glutinis | Microbotryomycetes | SAP |
| ASV00712 | 0.040 | SH1575129.08FU | 100 | Sporobolomyces ruberrimus | Microbotryomycetes | MP |
| ASV00279 | 0.037 | SH2232205.08FU | 99.7 | Briansuttonomyces eucalypti | Dothideomycetes | LITTER |
| ASV00441 | 0.037 | SH1646414.08FU | 98.8 | Agaricomycetes sp. | Agaricomycetes | |
| ASV00355 | 0.045 | SH2232205.08FU | 100 | Briansuttonomyces eucalypti | Dothideomycetes | LITTER |
| ASV00270 | 0.042 | SH1610160.08FU | 99.6 | Gibberella zeae | Sordariomycetes | PPATH |
| Kadarka | ||||||
| ASV00212 | 0.026 | SH1635391.08FU | 98.2 | Alternariaster helianthi | Dothideomycetes | PPATH |
| ASV00061 | 0.017 | SH1507512.08FU | 98.3 | Botryosphaeria dothidea | Dothideomycetes | PPATH |
| ASV00366 | 0.014 | SH1188671.08FU | 95.2 | Pyrenochaetopsis microspora | Dothideomycetes | COM |
| ASV00027 | 0.024 | SH1528219.08FU | 98.9 | Vishniacozyma sp. | Tremellomycetes | SAP |
| Syrah | ||||||
| ASV00266 | 0.024 | SH1564713.08FU | 98 | Keissleriella taminensis | Dothideomycetes | WOOD |
| Leaf | ||||||
| Furmint | ||||||
| ASV00142 | 0.013 | SH1609774.08FU | 99.6 | Dioszegia hungarica | Tremellomycetes | LITTER |
| ASV00012 | 0.016 | SH1515148.08FU | 99.3 | Aureobasidium pullulans | Dothideomycetes | SAP |
| ASV02203 | 0.046 | SH1575129.08FU | 100 | Sporobolomyces ruberrimus | Microbotryomycetes | MP |
| ASV00908 | 0.022 | SH1540544.08FU | 96.3 | Microstroma bacarum | Exobasidiomycetes | PPATH |
| ASV03652 | 0.038 | SH1615186.08FU | 96.7 | Ustilago crameri | Ustilaginomycetes | PPATH |
| Kadarka | ||||||
| ASV00352 | 0.001 | SH1562822.08FU | 100 | Erysiphe convolvuli | Leotiomycetes | PPATH |
| ASV00051 | 0.011 | SH1575708.08FU | 96.9 | Pleosporaceae sp. | Dothideomycetes | |
| ASV00257 | 0.016 | SH1505878.08FU | 96.8 | Comoclathris sp. | Dothideomycetes | WOOD |
| ASV00454 | 0.013 | SH1560734.08FU | 100 | Herpotrichiellaceae sp. | Eurotiomycetes | |
| ASV01091 | 0.028 | SH2337384.08FU | 99.6 | Septoria malagutii | Dothideomycetes | PPATH |
| ASV00010 | 0.001 | SH1171505.08FU | 99 | Ascomycota sp. | unidentified | |
| ASV00084 | 0.030 | SH2131097.08FU | 99.3 | Diplodia scrobiculata | Dothideomycetes | PPATH |
| Syrah | ||||||
| ASV00702 | 0.011 | SH1565276.08FU | 100 | Schizophyllum commune | Agaricomycetes | WOOD |
| ASV00947 | 0.006 | SH1155568.08FU | 98.7 | Clonostachys rosea | Sordariomycetes | WOOD |
| ASV00178 | 0.041 | SH2176172.08FU | 97.6 | Loratospora luzulae | Dothideomycetes | LITTER |
| ASV00477 | 0.002 | SH1246647.08FU | 99.4 | Coprinellus xanthothrix | Agaricomycetes | SAP |
| ASV00168 | 0.021 | SH1599074.08FU | 99.3 | Vuilleminia comedens | Agaricomycetes | WOOD |
| ASV00521 | 0.025 | SH2568050.08FU | 98.4 | Erysiphe necator | Leotiomycetes | PPATH |
| ASV02168 | 0.013 | SH1574527.08FU | 100 | Bulleromyces albus | Tremellomycetes | SAP |
| ASV00847 | 0.039 | SH1515148.08FU | 99 | Aureobasidium pullulans | Dothideomycetes | SAP |
| ASV04003 | 0.043 | SH1555460.08FU | 99 | Microdochium seminicola | Sordariomycetes | PPATH |
| ASV02439 | 0.038 | SH1643363.08FU | 99.6 | Muriphaeosphaeria viburni | Dothideomycetes | PPATH |
| ASV04868 | 0.043 | SH1572518.08FU | 99.7 | Melampsora laricis-populina | Pucciniomycetes | PPATH |
| ASV01105 | 0.039 | SH1539608.08FU | 98.5 | Myrmecridium phragmitis | Sordariomycetes | SAP |
| ASV01832 | 0.044 | SH1542846.08FU | 99.7 | Massaria anomia | Dothideomycetes | WOOD |
| ASV01602 | 0.045 | SH1509408.08FU | 99 | Ustilago nunavutica | Ustilaginomycetes | PPATH |
| Furmint | Kadarka | Syrah | |
|---|---|---|---|
| Leaves | |||
| P ** | 1627.80 ± 151.84 b | 1923.57 ± 84.79 a | 1407.17 ± 75.19 b |
| Fe * | 76.47 ± 11.42 a | 65.77 ± 3.20 ab | 53.43 ± 1.25 b |
| Zn | 151.43 ± 9.49 | 138.50 ± 18.79 | 132.47 ± 1.33 |
| Ca ** | 24,795.57 ± 1650.34 b | 23,770.10 ± 2531.14 b | 30,613.27 ± 967.30 a |
| Cu | 11.20 ± 4.62 | 9.57 ± 4.61 | 5.77 ± 1.03 |
| B | 115.20 ± 14.13 | 93.67 ± 19.61 | 94.33 ± 4.34 |
| K | 6897.20 ± 321.18 | 6903.80 ± 614.66 | 6083.83 ± 466.27 |
| Mn | 132.10 ± 26.69 | 163.73 ± 3.01 | 163.50 ± 5.09 |
| Mg ** | 4584.60 ± 176.31 b | 4110.17 ± 99.94 b | 5604.50 ± 597.11 a |
| Al | 162.20 ± 97.33 | 93.40 ± 20.20 | 122.13 ± 1.91 |
| Na | 975.07 ± 45.35 | 1003.90 ± 89.86 | 1075.10 ± 18.53 |
| N ** | 1.94 ± 0.07 b | 1.98 ± 0.08 b | 2.34 ± 0.01 a |
| C | 42.78 ± 0.53 | 42.38 ± 0.45 | 42.94 ± 0.33 |
| S | 0.19 ± 0.02 | 0.27 ± 0.06 | 0.19 ± 0.02 |
| Berries | |||
| Brix index *** | 24.50 ± 0.00 a | 23.53 ± 0.45 b | 21.00 ± 0.00 c |
| Total acid *** | 7.33 ± 0.06 a | 4.57 ± 0.12 b | 4.07 ± 0.06 c |
| Malic acid | 2.63 ± 0.90 | 3.37 ± 0.02 | 3.46 ± 0.01 |
| Alpha-amino nitrogen ** | 117.00 ± 2.65 b | 125.67 ± 4.93 ab | 133.33 ± 3.51 a |
| pH *** | 3.15 ± 0.01 c | 3.37 ± 0.02 b | 3.46 ± 0.01 a |
| Cultivar | Furmint | Kadarka | Syrah |
|---|---|---|---|
| Origin | convar. pontica subconvar. balcanica | convar. pontica subconvar. balcanica | convar. occidentalis subconvar. gallica |
| Time of bud burst (OIV 301) a | medium | late | early/medium |
| Time of full bloom (OIV 302) | late/very late | late | medium/late |
| Time of the onset of berry ripening (veraison (OIV 303) | medium | late | medium |
| Time of full physiological maturity of berry (OIV 304) | medium | late/very late | medium |
| Bunch: density (OIV 204) | dense | dense/very dense | medium/dense |
| Bunch: single bunch weight (OIV 502) | very low/low | medium/high | medium |
| Berry: single berry weight (OIV 503) | low | medium/high | low |
| Yield per m2 (OIV 504) | low/medium | high | high |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molnár, A.; Geml, J.; Geiger, A.; Leal, C.M.; Kgobe, G.; Tóth, A.M.; Villangó, S.; Mézes, L.; Czeglédi, M.; Lőrincz, G.; et al. Exploring Relationships among Grapevine Chemical and Physiological Parameters and Leaf and Berry Mycobiome Composition. Plants 2022, 11, 1924. https://doi.org/10.3390/plants11151924
Molnár A, Geml J, Geiger A, Leal CM, Kgobe G, Tóth AM, Villangó S, Mézes L, Czeglédi M, Lőrincz G, et al. Exploring Relationships among Grapevine Chemical and Physiological Parameters and Leaf and Berry Mycobiome Composition. Plants. 2022; 11(15):1924. https://doi.org/10.3390/plants11151924
Chicago/Turabian StyleMolnár, Anna, József Geml, Adrienn Geiger, Carla Mota Leal, Glodia Kgobe, Adrienn Mária Tóth, Szabolcs Villangó, Lili Mézes, Márk Czeglédi, György Lőrincz, and et al. 2022. "Exploring Relationships among Grapevine Chemical and Physiological Parameters and Leaf and Berry Mycobiome Composition" Plants 11, no. 15: 1924. https://doi.org/10.3390/plants11151924
APA StyleMolnár, A., Geml, J., Geiger, A., Leal, C. M., Kgobe, G., Tóth, A. M., Villangó, S., Mézes, L., Czeglédi, M., Lőrincz, G., & Zsófi, Z. (2022). Exploring Relationships among Grapevine Chemical and Physiological Parameters and Leaf and Berry Mycobiome Composition. Plants, 11(15), 1924. https://doi.org/10.3390/plants11151924