
Metabolic Variation among Fruits of Different Chili Cultivars (Capsicum spp.) Using HPLC/MS






Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction of Sugars, Organic Acids and Ascorbic Acid
2.2. Analysis of Sugars, Organic Acids and Ascorbic Acid
2.3. Extraction of Total Phenolics and Capsaicinoids
2.4. Analysis of Total Phenolic
2.5. Analysis of Capsaicinoids
2.6. Chemicals
2.7. Statistical Analysis
3. Results
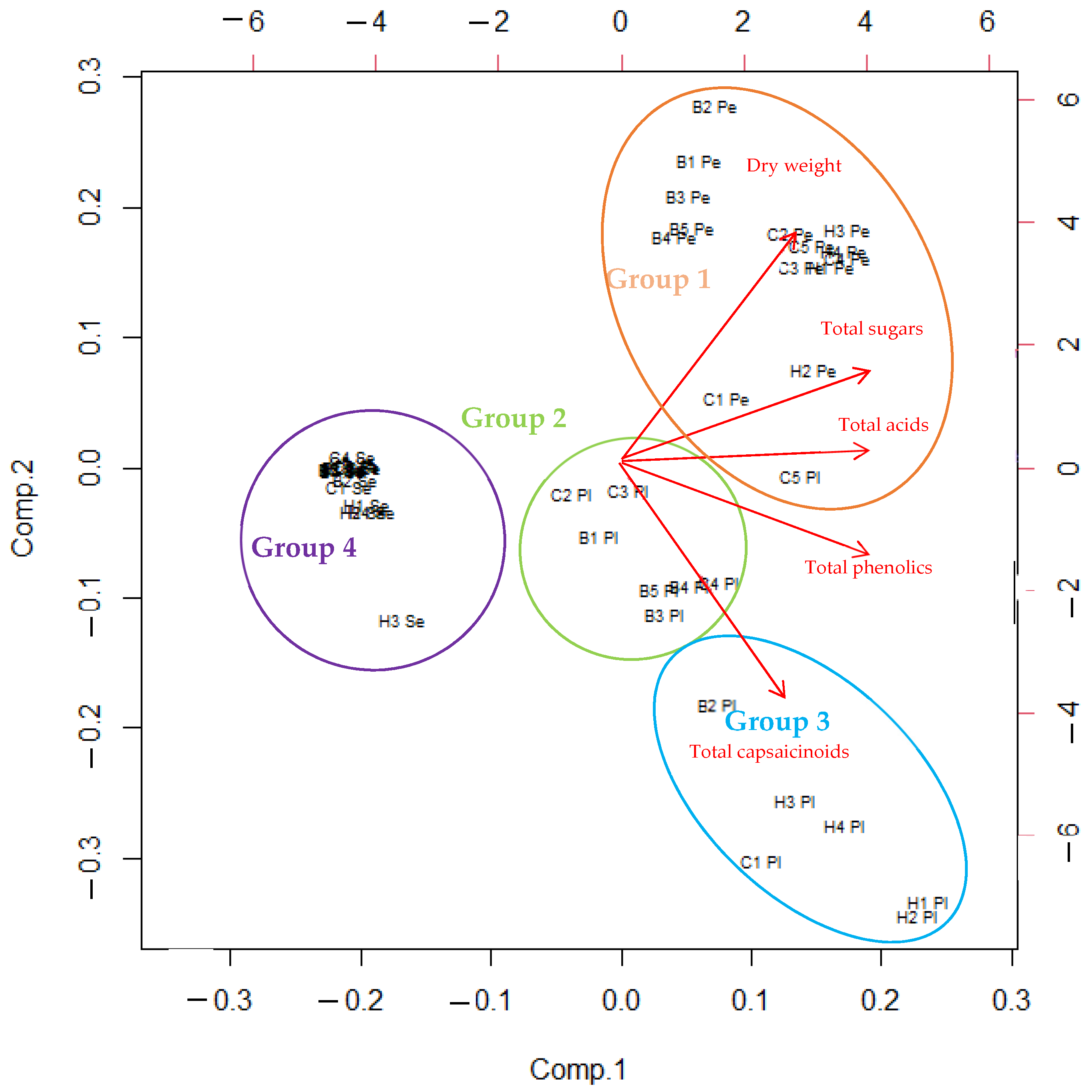
3.1. Multivariate Analysis of All Data
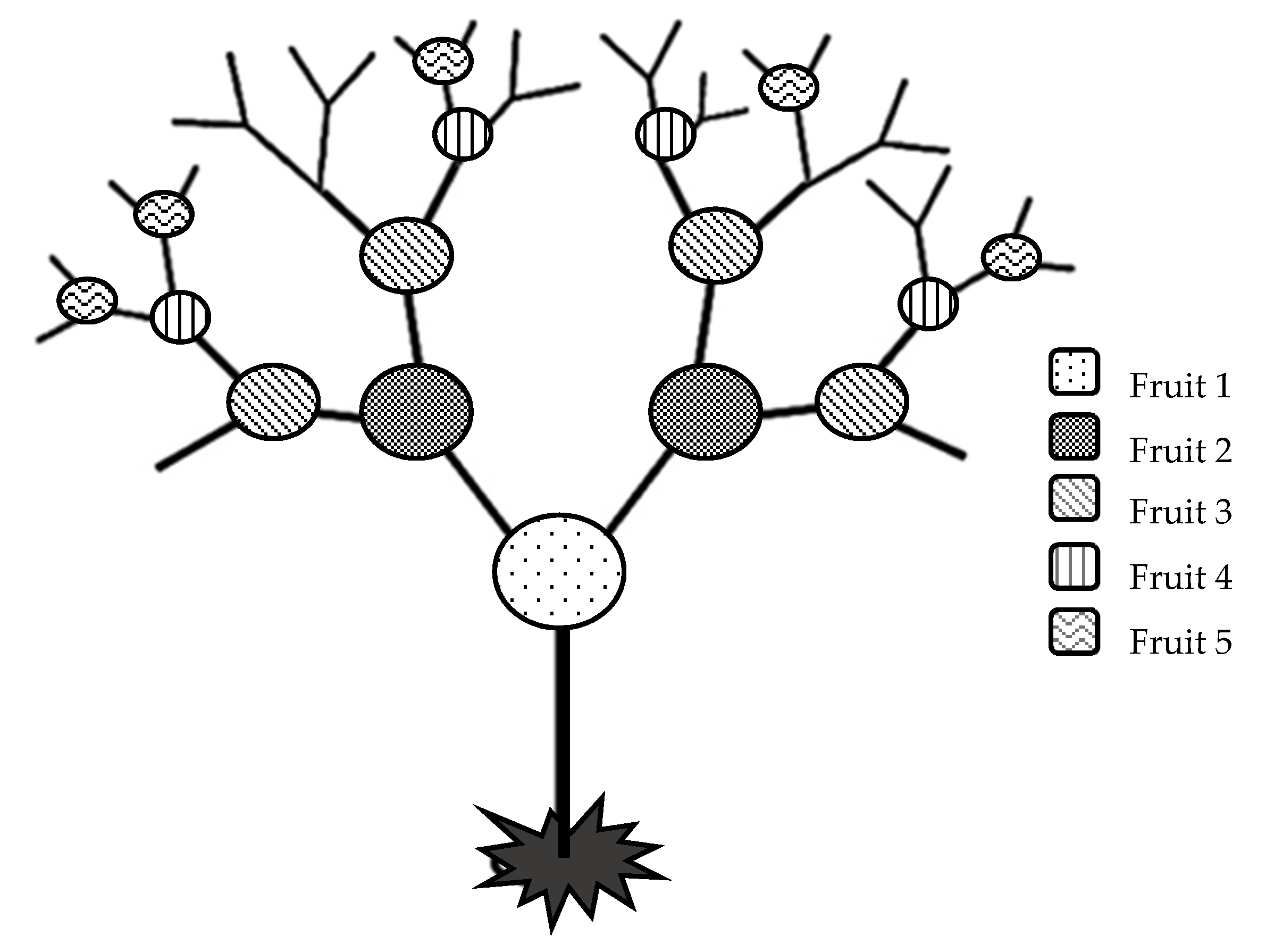
3.2. Fruit Weight
3.3. Sugars
3.4. Organic Acids
3.5. Total Phenolic Contents and Capsaicnoid Contents
3.5.1. Total Phenolic Contents
3.5.2. Capsaicinoids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Welbaum, G. Vegetable Production and Practices; CAB International: Wallingforth, UK, 2015; 486p. [Google Scholar]
- Fett, D.D. Botanical briefs: Capsicum peppers. Cutis 2003, 72, 21–23. [Google Scholar]
- Al Othman, Z.A.; Ahmed, Y.B.; Habila, M.A.; Ghafar, A.A. Determination of capsaicin and dihydrocapsaicin in Capsicum fruit samples using high performance liquid chromatography. Molecules 2011, 16, 8919–8929. [Google Scholar] [CrossRef] [Green Version]
- Zewdie, Y.; Bosland, P. Pungency of Chile (Capsicum annuum L.) Fruit is Affected by Node Position. HortScience 2000, 35, 1174. [Google Scholar] [CrossRef]
- Erickson, A.; Markhart, A.H. Flower developmental stage and organ sensitivity of bell pepper (Capsicum annuum L.) to elevated temperature. Plant Cell Environ. 2002, 25, 123–130. [Google Scholar] [CrossRef]
- Orobiyi, A. Capsaicin and Ascorbic Acid Content in the High Yielding Chili Pepper (Capsicum annuum L.) Landraces of Northern Benin. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 394–403. [Google Scholar]
- Verma, N.; Shukla, S. Impact of various factors responsible for fluctuation in plant secondary metabolites. J. Appl. Res. Med. Aromat. Plants 2015, 2, 105–113. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [Green Version]
- Fayos, O.; Ochoa-Alejo, N.; de la Vega, O.M.; Savirón, M.; Orduna, J.; Mallor, C.; Barbero, G.F.; Garcés-Claver, A. Assessment of Capsaicinoid and Capsinoid Accumulation Patterns during Fruit Development in Three Chili Pepper Genotypes (Capsicum spp.) Carrying Pun1 and pAMT Alleles Related to Pungency. J. Agric. Food Chem. 2019, 67, 12219–12227. [Google Scholar] [CrossRef]
- De Ávila Silva, L.; Condori-Apfata, J.A.; Marcelino, M.M.; Tavares, A.C.A.; Raimundi, S.C.J.; Martino, P.B.; Araújo, W.L.; Zsögön, A.; Sulpice, R.; Nunes-Nesi, A. Nitrogen differentially modulates photosynthesis, carbon allocation and yield related traits in two contrasting Capsicum chinense cultivars. Plant Sci. Int. J. Exp. Plant Biol. 2019, 283, 224–237. [Google Scholar] [CrossRef]
- Zamljen, T.; Veberic, R.; Hudina, M.; Slatnar, A. The Brown Marmorated Stink Bug (Halyomorpha halys Stål.) Influences Pungent and Non-Pungent Capsicum Cultivars’ Pre- and Post-Harvest Quality. Agronomy 2021, 11, 2252. [Google Scholar] [CrossRef]
- Zamljen, T.; Zupanc, V.; Slatnar, A. Influence of irrigation on yield and primary and secondary metabolites in two chilies species, Capsicum annuum L. and Capsicum chinense Jacq. Agric. Water Manag. 2020, 234, 106104. [Google Scholar] [CrossRef]
- Zamljen, T.; Hudina, M.; Veberič, R.; Slatnar, A. Biostimulative effect of amino acids and green algae extract on capsaicinoid and other metabolite contents in fruits of Capsicum spp. Chem. Biol. Technol. Agric. 2021, 8, 63. [Google Scholar] [CrossRef]
- Zamljen, T.; Jakopič, J.; Hudina, M.; Veberič, R.; Slatnar, A. Influence of intra and inter species variation in chilies (Capsicum spp.) on metabolite composition of three fruit segments. Sci. Rep. 2021, 11, 4932. [Google Scholar] [CrossRef]
- Medic, A.; Jakopic, J.; Hudina, M.; Solar, A.; Veberic, R. Identification and quantification of the major phenolic constituents in Juglans regia L. peeled kernels and pellicles, using HPLC–MS/MS. Food Chem. 2021, 352, 129404. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org (accessed on 15 December 2021).
- Alan, Ö.; Eser, B. Pepper seed yield and quality in relation to fruit position on the mother plant. PJBS 2008, 10, 4251–4255. [Google Scholar] [CrossRef]
- Krstić, B.; Tepić Horecki, A.; Nikolić, N.; Dj, G.; Tomicic, M. Chemical variability of inedible fruit parts in pepper varieties (Capsicum annuum L.). Bulg. J. Agric. Sci. 2013, 19, 490. [Google Scholar]
- Pandhair, V.; Sharma, S. Accumulation of Capsaicin in Seed, Pericarp and Placenta of Capsicum annuum L Fruit. J. Plant Biochem. Biotechnol. 2008, 17, 23–27. [Google Scholar] [CrossRef]
- Aldana-Iuit, J.G.; Sauri-Duch, E.; Miranda-Ham, M.D.L.; Castro-Concha, L.A.; Cuevas-Glory, L.F.; Vázquez-Flota, F.A. Nitrate Promotes Capsaicin Accumulation in Capsicum chinense Immobilized Placentas. BioMed Res. Int. 2015, 2015, 794084. [Google Scholar] [CrossRef] [Green Version]
- Gavilán Guillen, N.; Tito, R.; Gamarra Mendoza, N. Capsaicinoids and pungency in Capsicum chinense and Capsicum baccatum fruits1. Pesqui. Agropecu. Trop. 2018, 48, 237–244. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Pugliese, A.; Bonesi, M.; De Luca, D.; O’Brien, N.; Menichini, F.; Tundis, R. Influence of drying and cooking process on the phytochemical content, antioxidant and hypoglycaemic properties of two bell Capsicum annum L. cultivars. Food Chem. Toxicol. 2013, 53, 392–401. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Pugliese, A.; Bonesi, M.; Menichini, F.; Tundis, R. Evaluation of chemical profile and antioxidant activity of twenty cultivars from Capsicum annuum, Capsicum baccatum, Capsicum chacoense and Capsicum chinense: A comparison between fresh and processed peppers. LWT 2015, 64, 623–631. [Google Scholar] [CrossRef]
- Jarret, R.; Berke, T.; Baldwin, E.; Antonious, G. Variability for Free Sugars and Organic Acids in Capsicum chinense. Chem. Biodivers. 2009, 6, 138–145. [Google Scholar] [CrossRef]
- Iwai, K.; Suzuki, T.; Fujiwake, H.J.A. Formation and Accumulation of Pungent Principle of Hot Pepper Fruits, Capsaicin and Its Analogues, in Capsicum annuun var. annuun cv. Karayatsubusa at Different Growth Stages after Flowering. Agric. Biol. Chem. 1979, 43, 2493–2498. [Google Scholar]
- Lewallen, K.; Marini, R. Relationship between Flesh Firmness and Ground Color in Peach as Influenced by Light and Canopy Position. J. Am. Soc. Hortic. Sci. 2003, 128, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Willaume, M.; Lauri, P.-E.; Sinoquet, H. Light interception in apple trees influenced by canopy architecture manipulation. Trees 2004, 18, 705–713. [Google Scholar] [CrossRef]
- Ghasemnezhad, M.; Sherafati, M.; Payvast, G.A. Variation in phenolic compounds, ascorbic acid and antioxidant activity of five coloured bell pepper (Capsicum annum) fruits at two different harvest times. J. Funct. Foods 2011, 3, 44–49. [Google Scholar] [CrossRef]
- Contreras-Padilla, M.; Yahia, E.M. Changes in Capsaicinoids during Development, Maturation, and Senescence of Chile Peppers and Relation with Peroxidase Activity. J. Agric. Food Chem. 1998, 46, 2075–2079. [Google Scholar] [CrossRef]
- Estrada, B.; Bernal, M.A.; Díaz, J.; Pomar, F.; Merino, F. Capsaicinoids in vegetative organs of Capsicum annuum L. in relation to fruiting. J. Agric. Food Chem. 2002, 50, 1188–1191. [Google Scholar] [CrossRef]
- Coyago-Cruz, E.; Corell, M.; Moriana, A.; Hernanz, D.; Stinco, C.M.; Meléndez-Martínez, A.J. Effect of the fruit position on the cluster on fruit quality, carotenoids, phenolics and sugars in cherry tomatoes (Solanum lycopersicum L.). Food Res. Int. 2017, 100, 804–813. [Google Scholar] [CrossRef]
- Kalcsits, L.; Mattheis, J.; Giordani, L.; Reid, M.; Mullin, K.; Beres, B. Fruit canopy positioning affects fruit calcium and potassium concentrations, disorder incidence, and fruit quality for ‘Honeycrisp’ apple. Can. J. Plant Sci. 2019, 99, 761–771. [Google Scholar] [CrossRef]
- Feng, F.; Li, M.; Ma, F.; Cheng, L. Effects of location within the tree canopy on carbohydrates, organic acids, amino acids and phenolic compounds in the fruit peel and flesh from three apple (Malus × domestica) cultivars. Hortic. Res. 2014, 1, 14019. [Google Scholar] [CrossRef] [Green Version]
- Tundis, R.; Loizzo, M.R.; Menichini, F.; Bonesi, M.; Conforti, F.; De Luca, D.; Menichini, F. Air-dried Capsicum annuum var. acuminatum medium and big: Determination of bioactive constituents, antioxidant activity and carbohydrate-hydrolyzing enzymes inhibition. Food Res. Int. 2012, 45, 170–176. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Treatment | Dry Weight | Level 1 | Level 2 | Level 3 | Level 4 | Level 5 |
|---|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | ||
| C. annuum ‘Cayenne’ | Pericarp | 3.72 ± 0.21 | 11.75 ± 1.71 *** | 9.33 ± 0.36 * | 9.51 ± 0.67 * | 8.92 ± 0.88 * |
| Placenta | 0.26 ± 0.01 | 0.73 ± 0.16 * | 0.72 ± 0.06 ** | 0.74 ± 0.01 ** | 0.86 ± 0.12 *** | |
| Seed | 0.47 ± 0.01 | 1.34 ± 0.12 * | 1.36 ± 0.12 * | 1.44 ± 0.06 ** | 1.22 ± 0.16 | |
| Whole fruit | 4.14 ± 0.31 | 13.71 ± 1.23 *** | 11.35 ± 0.93 ** | 11.62 ± 0.86 ** | 10.95 ± 1.01 ** | |
| C. chinense ‘Habanero Orange’ | Pericarp | 12.61 ± 1.11 | 9.25 ± 0.87 ** | 14.71 ± 1.34 | 14.16 ± 1.11 | |
| Placenta | 1.82 ± 0.63 | 1.56 ± 0.26 | 2.42 ± 0.13 | 2.17 ± 0.46 | ||
| Seed | 0.82 ± 0.16 | 0.47 ± 0.02 | 0.81 ± 0.26 | 0.87 ± 0.13 | ||
| Whole fruit | 15.23 ± 1.32 | 11.19 ± 0.77 ** | 17.92 ± 1.45 | 17.08 ± 1.63 | ||
| C. baccatum ‘Bishop Crown’ | Pericarp | 10.17 ± 1.13 | 12.63 ± 1.08 ** | 9.30 ± 0.76 | 8.18 ± 0.33 | 7.53 ± 0.79 *** |
| Placenta | 0.57 ± 0.07 | 0.72 ± 0.13 | 0.47 ± 0.17 | 0.52 ± 0.12 | 0.58 ± 0.06 | |
| Seed | 0.82 ± 0.07 | 1.08 ± 0.12 | 0.72 ± 0.27 | 0.67 ± 0.22 | 0.61 ± 0.07 | |
| Whole fruit | 11.48 ± 0.52 | 14.32 ± 1.27 *** | 10.48 ± 0.92 | 9.22 ± 0.37 ** | 8.67 ± 0.52 *** |
| Species | Sugar | Fruit Part | Level 1 | Level 2 | Level 3 | Level 4 | Level 5 |
|---|---|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |||
| C. annuum ‘Cayenne’ | Sucrose | Pericarp | 17.26 ± 1.23 | 19.21 ± 1.89 | 14.11 ± 0.82 | 22.53 ± 0.36 | 27.10 ± 1.21 |
| Placenta | 4.22 ± 0.11 | 41.51 ± 3.20 *** | 16.94 ± 1.85 * | 17.89 ± 1.21 | 95.69 ± 7.61 *** | ||
| Glucose | Pericarp | 143.72 ± 3.71 | 134.96 ± 8.16 | 159.47 ± 1.76 ** | 166.77 ± 2.32 ** | 201.93 ± 4.12 *** | |
| Placenta | 23.34 ± 1.53 | 43.85 ± 2.64 | 70.20 ± 6.77 ** | 79.81 ± 6.83 ** | 138.31 ± 3.36 *** | ||
| Fructose | Pericarp | 147.28 ± 7.37 | 140.43 ± 2.91 | 159.22 ± 3.41 | 162.71 ± 5.37 | 179.31 ± 3.47 ** | |
| Placenta | 68.67 ± 4.98 | 79.42 ± 7.38 | 150.11± 3.61 *** | 155.94 ± 7.26 *** | 236.35 ± 7.12 *** | ||
| Total sugars | Pericarp | 308.12± 13.22 | 294.52± 10.86 | 332.79± 15.32 | 351.94± 14.35 | 408.30± 20.46 | |
| Placenta | 96.18± 8.34 | 164.74± 11.46 | 237.18± 16.33 | 253.59± 17.24 | 470.26± 19.68 | ||
| C. chinense ‘Habanero Orange’ | Sucrose | Pericarp | 21.81 ± 1.81 | 17.21 ± 1.72 | 19.80 ± 0.93 | 14.62 ± 1.31 | |
| Placenta | 15.91 ± 0.91 | 89.41 ± 9.31 | 11.33 ± 0.28 | 14.29 ± 1.42 | |||
| Glucose | Pericarp | 169.43 ± 8.23 | 163.46 ± 4.48 | 171.55 ± 4.36 | 149.46 ± 9.16 | ||
| Placenta | 136.99 ± 6.28 | 102.89 ± 9.20 | 88.77 ± 7.12 | 122.91 ± 4.75 | |||
| Fructose | Pericarp | 216.35 ± 9.17 | 199.13 ± 2.63 | 203.11 ± 0.71 | 203.39 ± 12.52 | ||
| Placenta | 193.11 ± 3.91 | 155.63 ± 8.84 | 114.46 ± 4.37 | 153.33 ± 2.81 | |||
| Total sugars | Pericarp | 407.57± 20.79 | 379.68±19.25 | 394.25± 19.92 | 367.30± 18.41 | ||
| Placenta | 345.96± 17.52 | 347.83± 18.31 | 214.37± 15.53 | 290.43± 20.42 | |||
| C. baccatum ‘Bishop Crown’ | Sucrose | Pericarp | 19.14 ± 0.58 | 31.61 ± 1.22 ** | 32.11 ± 1.42 ** | 13.95 ± 1.97 | 30.71 ± 2.27 |
| Placenta | 15.28 ± 2.50 | 48.57 ± 4.89 | 58.35 ± 2.07 | 120.77 ± 5.41 *** | 60.55 ± 1.47 | ||
| Glucose | Pericarp | 197.35 ± 3.62 | 181.48 ± 3.45 | 168.04 ± 1.09 | 153.32 ± 11.12 | 169.27 ± 5.82 | |
| Placenta | 69.42 ± 1.11 | 85.52 ± 3.65 * | 64.47 ± 1.92 | 71.41 ± 2.77 | 59.21 ± 3.58 | ||
| Fructose | Pericarp | 196.42 ± 11.14 | 170.82 ± 11.21 | 165.41 ± 15.91 | 161.38 ± 10.56 | 195.11 ± 9.31 | |
| Placenta | 130.70 ± 3.63 | 113.68 ± 2.57 | 166.11 ± 3.56 | 152.33 ± 3.52 | 139.06 ± 8.14 | ||
| Total sugars | Pericarp | 412.83± 22.41 | 383.84± 21.85 | 365.53± 25.36 | 328.59± 15.35 | 395.02± 17.94 | |
| Placenta | 215.30± 20.36 | 247.61± 18.94 | 288.84± 20.41 | 344.48± 19.09 | 258.73± 16.59 |
| Species | Organic Acid | Fruit Part | Level 1 | Level 2 | Level 3 | Level 4 | Level 5 |
|---|---|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |||
| C. annuum ‘Cayenne’ | Ascorbic acid | Pericarp | 18.11 ± 1.19 | 15.12 ± 1.50 | 13.40 ± 0.15 *** | 15.37 ± 0.54 | 16.89 ± 0.78 |
| Placenta | 4.27 ± 0.04 | 3.60 ± 0.03 | 0.70 ± 0.05 *** | 0.91 ± 0.03 *** | 2.81 ± 0.19 | ||
| Citric acid | Pericarp | 73.84 ± 6.62 | 63.91 ± 3.82 | 75.42 ± 5.55 | 88.96 ± 5.30 | 80.06 ± 4.42 | |
| Placenta | 55.82 ± 4.96 | 68.99 ± 6.22 | 63.52 ± 2.21 | 60.31 ± 3.22 | 98.93 ± 4.32 | ||
| Malic acid | Pericarp | 14.46 ± 1.72 | 15.02 ± 1.95 | 21.97 ± 2.03 ** | 24.74 ± 1.55 *** | 14.43 ± 0.93 | |
| Placenta | 35.76 ± 2.02 | 27.32 ± 1.84 | 32.04 ± 3.15 | 48.94 ± 3.74 | 45.96 ± 1.70 | ||
| Quinic acid | Pericarp | 11.32 ± 1.06 | 10.77 ± 0.92 | 15.32 ± 1.26 * | 18.37 ± 1.42 * | 12.72 ± 0.99 | |
| Placenta | 12.60 ± 0.56 | 9.84 ± 0.42 | 12.36 ± 1.02 | 14.82 ± 1.36 | 14.42 ± 1.66 | ||
| Succinic acid | Pericarp | 0.26 ± 0.01 | 0.29 ± 0.02 | 0.36 ±0.02 | 0.38 ± 0.01 | 0.40 ± 0.035 | |
| Placenta | 0.58 ± 0.04 | 0.75 ± 0.03 | 0.51 ± 0.01 | 0.76 ± 0.03 | 1.06 ± 0.09 | ||
| Fumaric acid | Pericarp | 0.04 ± 0.01 | 0.04 ± 0.01 | 0.07 ± 0.01 ** | 0.06 ± 0.01 | 0.04 ± 0.01 | |
| Placenta | 0.25 ± 0.02 | 0.16 ± 0.01 | 0.16 ± 0.01 | 0.21 ± 0.02 | 0.13 ± 0.01 | ||
| Oxalic acid | Pericarp | 1.49 ± 0.10 | 1.66 ± 0.15 | 2.74 ± 0.25 | 5.55 ± 0.29 *** | 2.96 ± 0.23 | |
| Placenta | 5.23 ± 0.43 | 3.74 ± 0.32 | 5.41 ± 0.35 | 9.49 ± 0.72 *** | 6.09 ± 0.59 | ||
| Total organic acids | Pericarp | 164.81 ± 10.46 | 149.61 ± 11.42 | 190.17 ± 12.52 | 226.51 ± 14.52 | 178.11 ± 13.24 | |
| Placenta | 165.12 ± 9.73 | 153.55 ± 10.52 | 163.82 ± 11.52 | 194.67 ± 14.42 | 227.02 ± 15.52 | ||
| C. chinense ‘Habanero Orange’ | Ascorbic acid | Pericarp | 12.95 ± 1.08 | 14.61 ± 0.94 | 15.68 ± 1.17 | 16.62 ± 1.29 | |
| Placenta | 1.54 ± 0.04 | 2.27 ± 0.07 | 1.39 ± 0.06 | 2.27 ± 0.11 | |||
| Citric acid | Pericarp | 36.32 ± 3.52 | 35.73 ± 4.03 | 38.47 ± 2.12 | 44.22 ± 2.32 | ||
| Placenta | 41.82 ± 3.68 | 33.07 ± 3.42 ** | 23.92 ± 1.62 *** | 36.62 ± 2.74 | |||
| Malic acid | Pericarp | 27.36 ± 2.56 | 27.12 ± 1.94 | 18.14 ± 1.37 ** | 16.06 ± 1.23 ** | ||
| Placenta | 44.37 ± 4.37 | 49.84 ± 4.23 | 32.73 ± 2.37 | 44.26 ± 4.63 | |||
| Quinic acid | Pericarp | 11.92 ± 0.92 | 11.86 ± 1.16 | 11.22 ± 0.72 | 12.04 ± 0.94 | ||
| Placenta | 16.73 ± 0.72 | 14.22 ± 1.84 | 8.47 ± 0.45 ** | 13.13 ± 1.06 | |||
| Succinic acid | Pericarp | 0.29 ± 0.01 | 0.38 ±0.02 | 0.40 ± 0.03 | 0.38 ± 0.02 | ||
| Placenta | 0.64 ± 0.02 | 0.59 ± 0.03 | 0.39 ± 0.01 | 0.64 ± 0.01 | |||
| Fumaric acid | Pericarp | 0.15 ± 0.01 | 0.12 ± 0.01 | 0.07 ± 0.01 | 0.18 ± 0.01 | ||
| Placenta | 0.37 ± 0.01 | 0.24 ± 0.01 | 0.28 ± 0.01 | 0.29 ± 0.01 | |||
| Oxalic acid | Pericarp | 2.34 ± 0.22 | 2.65 ± 0.21 | 2.36 ± 0.10 | 3.76 ± 0.32 | ||
| Placenta | 2.10 ± 0.20 | 2.03 ± 0.19 | 1.33 ± 0.03 | 2.06 ± 0.13 | |||
| Total organic acids | Pericarp | 138.79 ± 8.52 | 139.67 ± 9.42 | 131.04 ± 9.32 | 140.94 ± 9.15 | ||
| Placenta | 174.22 ± 10.52 | 158.82 ± 10.99 | 102.29 ± 7.24 | 151.56 ± 8.27 | |||
| C. baccatum ‘Bishop Crown’ | Ascorbic acid | Pericarp | 5.47 ± 0.07 | 5.65 ± 0.01 | 6.64 ± 0.03 | 7.09 ± 0.05 | 5.44 ± 0.03 |
| Placenta | 1.56 ± 0.06 | 1.35 ± 0.07 | 2.96 ± 0.14 | 1.59 ± 0.02 | 2.31 ± 0.15 | ||
| Citric acid | Pericarp | 53.23 ± 4.25 | 65.22 ± 5.99 | 64.27 ± 6.6 | 65.35 ± 2.95 | 70.02 ± 5.68 | |
| Placenta | 83.72 ± 7.22 | 70.12 ± 5.67 | 57.73 ± 3.54 *** | 87.23 ± 7.96 | 98.22 ± 7.15 | ||
| Malic acid | Pericarp | 8.77 ± 0.25 | 9.97 ± 0.66 | 11.54 ± 1.02 | 5.54 ± 0.44 | 5.73 ± 0.56 | |
| Placenta | 14.27 ± 1.33 | 16.83 ± 1.34 | 18.06 ± 1.72 | 11.88 ± 0.82 | 13.65 ± 1.02 | ||
| Quinic acid | Pericarp | 7.83 ± 0.62 | 7.72 ± 0.34 | 7.71 ± 0.49 | 8.63 ± 0.62 | 8.96 ± 0.22 | |
| Placenta | 6.43 ± 0.47 | 8.46 ± 0.72 | 7.57 ± 1.07 | 6.49 ± 0.55 | 5.62 ± 0.42 | ||
| Succinic acid | Pericarp | 0.60 ± 0.02 | 0.47 ± 0.03 | 0.42 ± 0.02 | 0.55 ±0.04 | 0.43 ± 0.02 | |
| Placenta | 0.82 ± 0.06 | 0.71 ± 0.04 | 0.95 ± 0.07 | 0.87 ± 0.08 | 0.69 ± 0.06 | ||
| Fumaric acid | Pericarp | 0.07 ± 0.01 | 0.09 ± 0.01 | 0.07 ± 0.01 | 0.06 ± 0.01 | 0.06 ± 0.01 | |
| Placenta | 0.10 ± 0.01 | 0.11 ± 0.01 | 0.09 ± 0.01 | 0.11 ± 0.01 | 0.08 ± 0.01 | ||
| Oxalic acid | Pericarp | 0.40 ± 0.03 | 0.33 ± 0.01 | 0.19 ± 0.01 *** | 0.17 ± 0.01 *** | 0.16 ± 0.01 *** | |
| Placenta | 1.55 ± 0.04 | 1.55 ± 0.12 | 2.08 ± 0.18 | 1.44 ± 0.12 | 1.31 ± 0.07 | ||
| Total organic acids | Pericarp | 107.69 ± 6.62 | 120.29 ± 7.10 | 121.32 ± 9.12 | 121.58 ± 8.42 | 126.25 ± 9.42 | |
| Placenta | 134.26 ± 8.48 | 132.62 ± 9.42 | 119.14 ± 9.22 | 135.20 ± 10.49 | 144.22 ± 10.85 |
| Species | Fruit Part | Level 1 | Level 2 | Level 3 | Level 4 | Level 5 |
|---|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | ||
| C. annuum ‘Cayenne’ | Pericarp | 20.00 ± 0.67 | 22.26 ± 0.14 | 19.99 ± 0.16 | 21.59 ± 0.12 | 19.83 ± 0.11 |
| Placenta | 27.85 ± 0.04 | 6.70 ± 0.03 *** | 10.50 ± 0.08 *** | 16.25 ± 0.10 | 14.69 ± 0.92 * | |
| Seed | 2.66 ± 0.02 | 2.81 ± 0.01 | 2.68 ± 0.01 | 2.38 ± 0.02 | 2.57 ± 0.01 | |
| Whole fruit | 10.76 ± 0.06 | 7.75 ± 0.02 *** | 7.82 ± 0.05 *** | 8.96 ± 0.06 * | 8.43 ± 0.04 ** | |
| C. chinense ‘Habanero Orange’ | Pericarp | 17.59 ± 0.06 | 20.25 ± 0.07 | 19.03 ± 0.04 | 19.70 ± 0.10 | |
| Placenta | 30.43 ± 0.19 | 30.28 ± 0.12 | 31.46 ± 0.09 | 25.89 ± 0.17 | ||
| Seed | 3.86 ± 0.03 | 3.73 ± 0.03 | 4.09 ± 0.03 | 4.34 ± 0.04 | ||
| Whole fruit | 12.19 ± 0.10 | 12.61 ± 0.11 | 12.82 ± 0.10 | 11.93 ± 0.09 | ||
| C. baccatum ‘Bishop Crown’ | Pericarp | 10.02 ± 0.08 | 9.75 ± 0.04 | 10.31 ± 0.06 | 11.42 ± 0.06 | 10.82 ± 0.09 |
| Placenta | 8.61 ± 0.04 | 19.01 ± 0.17 *** | 13.47 ± 0.11 * | 12.30 ± 0.12 | 12.12 ± 0.07 | |
| Seed | 2.08 ± 0.01 | 2.51 ± 0.01 | 1.87 ± 0.01 | 1.76 ± 0.01 | 1.69 ± 0.01 | |
| Whole fruit | 5.01 ± 0.03 | 6.44 ± 0.03 | 5.54 ± 0.04 | 5.63 ± 0.03 | 5.40 ± 0.05 |
| Species | Capsaicinoid | Fruit Part | Fruit 1 | Fruit 2 | Fruit 3 | Fruit 4 | Fruit 5 |
|---|---|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |||
| C. annuum ‘Cayenne’ | Capsaicin | Pericarp | 0.58 ± 0.03 | 1.15 ± 0.13 | 1.09 ± 0.12 | 0.92 ± 0.06 | 0.64 ± 0.13 |
| Placenta | 9.67 ± 1.36 | 2.10 ± 0.32 *** | 2.27 ± 0.63 *** | 4.23 ± 0.56 | 2.83 ± 0.05 * | ||
| Seeds | 0.62 ± 0.04 | 0.56 ± 0.04 | 0.67 ± 0.05 | 0.41 ± 0.04 | 0.37 ± 0.05 | ||
| Dihydrocapsaicin | Pericarp | 0.12 ± 0.67 | 0.20 ± 0.04 | 0.20 ± 0.01 | 0.15 ± 0.01 | 0.11 ± 0.02 | |
| Placenta | 2.70 ± 0.21 | 0.56 ± 0.02 *** | 0.42 ± 0.07 *** | 1.15 ± 0.07 | 1.04 ± 0.09 *** | ||
| Seeds | 0.10 ± 0.01 | 0.09 ± 0.05 | 0.01 ± 0.01 | 0.05 ± 0.01 | 0.04 ± 0.01 | ||
| Nordihydrocapsaicin | Pericarp | 0.11 ± 0.21 | 0.22 ± 0.05 | 0.20 ± 0.01 | 0.17 ± 0.02 | 0.18 ± 0.05 | |
| Placenta | 2.03 ± 0.04 | 0.43 ± 0.03 *** | 0.40 ± 0.07 *** | 0.79 ± 0.03 * | 0.53 ± 0.04 * | ||
| Seeds | 0.10 ± 0.01 | 0.11 ± 0.03 | 0.01 ± 0.01 | 0.07 ± 0.02 | 0.06 ± 0.02 | ||
| Homocapsaicin | Pericarp | 0.03 ± 0.01 | 0.06 ± 0.03 | 0.06 ± 0.01 | 0.06 ± 0.01 | 0.05 ± 0.01 | |
| Placenta | 0.69 ± 0.11 | 0.17 ± 0.01 ** | 0.13 ± 0.02 *** | 0.43 ± 0.06 | 0.34 ± 0.04 | ||
| Seeds | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.02 ± 0.01 | ||
| Homodihydrocapsaicin | Pericarp | 0.04 ± 0.01 | 0.11 ± 0.05 | 0.10 ± 0.01 | 0.09 ± 0.01 | 0.08 ± 0.02 | |
| Placenta | 1.04 ± 0.26 | 0.30 ± 0.01 | 0.21 ± 0.02 | 0.63 ± 0.04 | 0.58 ± 0.03 | ||
| Seeds | 0.05 ± 0.02 | 0.04 ± 0.01 | 0.06 ± 0.01 | 0.04 ± 0.01 | 0.03 ± 0.02 | ||
| Total capsaicinoids | Pericarp | 0.90 ± 0.32 | 1.76 ± 0.12 | 1.66 ± 0.15 | 1.41 ± 0.10 | 1.07 ± 0.25 | |
| Placenta | 16.15 ± 0.70 | 3.57 ± 0.40 *** | 3.45 ± 0.81 *** | 7.25 ± 0.52 | 5.34 ± 0.16 * | ||
| Seeds | 0.91 ± 0.07 | 0.84 ± 0.03 | 1.03 ± 0.08 | 0.62 ± 0.07 | 0.55 ± 0.09 | ||
| C. chinense ‘Habanero Orange’ | Capsaicin | Pericarp | 6.57 ± 0.59 | 6.51 ± 0.38 | 6.54 ± 0.25 | 6.31 ± 0.43 | |
| Placenta | 17.52 ± 0.18 | 17.27 ± 0.28 | 12.48 ± 0.39 | 15.27 ± 0.18 | |||
| Seeds | 1.70 ± 0.13 | 1.63 ± 0.06 | 2.72 ± 0.05 | 1.97 ± 0.03 | |||
| Dihydrocapsaicin | Pericarp | 0.58 ± 0.03 | 0.66 ± 0.07 | 0.67 ± 0.03 | 0.73 ± 0.11 | ||
| Placenta | 3.17 ± 0.18 | 3.35 ± 0.48 | 1.96 ± 0.53 | 2.46 ± 0.47 | |||
| Seeds | 0.16 ± 0.04 | 0.17 ± 0.06 | 0.30 ± 0.07 | 0.22 ± 0.04 | |||
| Nordihydrocapsaicin | Pericarp | 0.22 ± 0.02 | 0.19 ± 0.06 | 0.28 ± 0.02 | 0.27 ± 0.02 | ||
| Placenta | 1.50 ± 0.11 | 1.70 ± 0.34 | 0.96 ± 0.28 | 1.23 ± 0.09 | |||
| Seeds | 0.08 ± 0.02 | 0.09 ± 0.03 | 0.01 ± 0.02 | 0.01 ± 0.02 | |||
| Homocapsaicin | Pericarp | 0.10 ± 0.01 | 0.11 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.01 | ||
| Placenta | 0.53 ± 0.04 | 0.76 ± 0.11 | 0.34 ± 0.09 | 0.38 ± 0.01 | |||
| Seeds | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.05 ± 0.01 | 0.03 ± 0.01 | |||
| Homodihydrocapsaicin | Pericarp | 0.08 ± 0.02 | 0.10 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 | ||
| Placenta | 0.73 ± 0.04 | 0.82 ± 0.21 | 0.54 ± 0.0.1 | 0.77 ± 0.01 | |||
| Seeds | 0.02 ± 0.01 | 0.03 ± 0.01 | 0.06 ± 0.01 | 0.03 ± 0.01 | |||
| Total capsaicinoids | Pericarp | 7.56 ± 0.65 | 7.59 ± 0.40 | 7.75 ± 0.21 | 7.56 ± 0.58 | ||
| Placenta | 23.47 ± 0.23 | 23.92 ± 0.39 | 16.29 ± 0.47 | 20.13 ± 0.16 | |||
| Seeds | 2.00 ± 0.43 | 1.98 ± 0.76 | 3.30 ± 0.70 | 2.37 ± 0.43 | |||
| C. baccatum ‘Bishop Crown’ | Capsaicin | Pericarp | 0.44 ± 0.03 | 0.36 ± 0.05 | 0.47 ± 0.07 | 0.36 ± 0.11 | 0.23 ± 0.01 |
| Placenta | 4.58 ± 0.11 | 9.97 ± 0.15 ** | 7.05 ± 0.47 * | 6.84 ± 1.35 | 6.25 ± 0.41 | ||
| Seeds | 0.38 ± 0.10 | 1.09 ± 0.48 | 0.37 ± 0.08 | 0.43 ± 0.10 | 0.30 ± 0.06 | ||
| Dihydrocapsaicin | Pericarp | 0.08 ± 0.02 | 0.07 ± 0.01 | 0.09 ± 0.01 | 0.08 ± 0.01 | 0.04 ± 0.01 | |
| Placenta | 0.64 ± 0.01 | 1.41 ± 0.14 | 1.36 ± 0.21 | 1.23 ± 0.25 | 1.06 ± 0.15 | ||
| Seeds | 0.06 ± 0.01 | 0.22 ± 0.01 *** | 0.08 ± 0.01 | 0.09 ± 0.01 | 0.06 ± 0.01 | ||
| Nordihydrocapsaicin | Pericarp | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.03 ± 0.01 | 0.03 ± 0.01 | 0.01 ± 0.01 | |
| Placenta | 0.30 ± 0.05 | 0.80 ± 0.08 | 0.65 ± 0.01 | 0.64 ± 0.11 | 0.62 ± 0.07 | ||
| Seeds | 0.01 ± 0.00 | 0.04 ± 0.01 *** | 0.02 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | ||
| Homocapsaicin | Pericarp | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.02 ± 0.01 ** | 0.01 ± 0.01 ** | 0.01 ± 0.00 | |
| Placenta | 0.16 ± 0.04 | 0.32 ± 0.07 | 0.31 ± 0.06 | 0.38 ± 0.14 | 0.29 ± 0.03 | ||
| Seeds | 0.01 ± 0.01 | 0.01 ± 0.00 ** | 0.01 ± 0.00 | 0.01 ± 0.00 ** | 0.01 ± 0.00 | ||
| Homodihydrocapsaicin | Pericarp | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | |
| Placenta | 0.07 ± 0.01 | 0.27 ± 0.05 *** | 0.17 ± 0.02 | 0.19 ± 0.05 | 0.18 ± 0.03 | ||
| Seeds | 0.01 ± 0.00 | 0.01 ± 0.00 *** | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | ||
| Total capsaicinoids | Pericarp | 58.33 ± 5.90 | 48.66 ± 8.07 | 61.98 ± 11.12 | 50.35 ± 16.27 | 30.90 ± 1.56 | |
| Placenta | 5.78 ± 0.13 | 12.77 ± 1.82 | 9.56 ± 0.78 | 9.30 ± 0.19 | 8.42 ± 0.70 | ||
| Seeds | 0.48 ± 0.12 | 1.40 ± 0.61 | 0.49 ± 0.10 | 0.56 ± 0.13 | 0.40 ± 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zamljen, T.; Medič, A.; Veberič, R.; Hudina, M.; Jakopič, J.; Slatnar, A. Metabolic Variation among Fruits of Different Chili Cultivars (Capsicum spp.) Using HPLC/MS. Plants 2022, 11, 101. https://doi.org/10.3390/plants11010101
Zamljen T, Medič A, Veberič R, Hudina M, Jakopič J, Slatnar A. Metabolic Variation among Fruits of Different Chili Cultivars (Capsicum spp.) Using HPLC/MS. Plants. 2022; 11(1):101. https://doi.org/10.3390/plants11010101
Chicago/Turabian StyleZamljen, Tilen, Aljaž Medič, Robert Veberič, Metka Hudina, Jerneja Jakopič, and Ana Slatnar. 2022. "Metabolic Variation among Fruits of Different Chili Cultivars (Capsicum spp.) Using HPLC/MS" Plants 11, no. 1: 101. https://doi.org/10.3390/plants11010101
APA StyleZamljen, T., Medič, A., Veberič, R., Hudina, M., Jakopič, J., & Slatnar, A. (2022). Metabolic Variation among Fruits of Different Chili Cultivars (Capsicum spp.) Using HPLC/MS. Plants, 11(1), 101. https://doi.org/10.3390/plants11010101