
Chromosome and Molecular Analyses Reveal Significant Karyotype Diversity and Provide New Evidence on the Origin of Aegilops columnaris

 ,
,  ,
,
Abstract
1. Introduction
2. Results
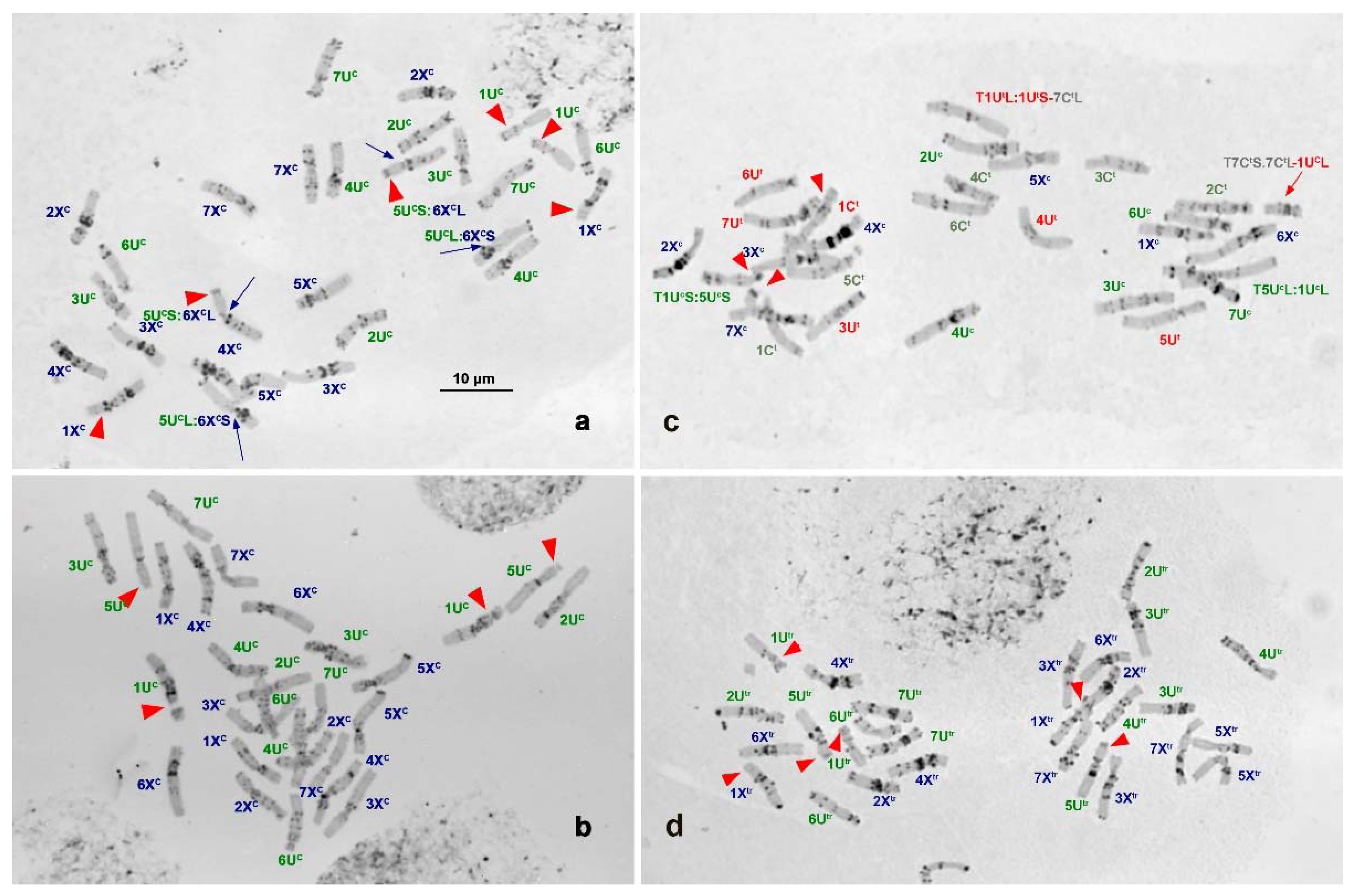
2.1. C-Banding Analysis of Ae. columnaris
2.2. FISH Analysis of Ae. columnaris
2.3. Analysis of Gliadin Spectra of Ae. columnaris
2.4. Variability of the U-Genome Specific U31 Nuclear Fragment in Ae. columnaris and Ae. neglecta
2.5. Variability of Three Plastome Intergenic Spacers in Ae. columnaris and Ae. neglecta
3. Discussion
- The chromosome 6Xt of Ae. neglecta possesses a minor 45S rDNA locus, which probably pre-existed in the progenitor Aegilops species; however, this locus is absent in Ae. columnaris.
- Ae. columnaris is characterized by chromosome instability expressed in a higher proportion and broader diversity of chromosomal rearrangements (20 variants in more than 55% of accessions). Chromosome instability is an essential factor of speciation [49,50] and is usually more expressed in phylogenetically new species. In addition, we found significant intraspecific polymorphism in Ae. columnaris plastome, although the only low variation of the chloroplast DNA sequences was recorded in Triticum and Aegilops species [51,52].
4. Materials and Methods
4.1. DNA Probes
- pSc119.2—a 120 bp long sequence isolated from rye [57];
- The oligo-(GTT)9 probe labeled at the 3′-end with fluorescein−12-dUTP was synthesized in the laboratory of biological microchips at the Engelhardt Institute of Molecular Biology, Moscow, Russia.
- The oligo-(GAA)10 probe labeled at the 3′-end with fluorescein−12-dUTP or Cy3 was synthesized in the laboratory of biological microchips at the Engelhardt Institute of Molecular Biology, Moscow, Russia.
- The oligo-(ACT)10 probe labeled at the 3′-end with Cy3 was synthesized in the laboratory of biological microchips at the Engelhardt Institute of Molecular Biology, Moscow, Russia.
4.2. Giemsa C-Banding Method
4.3. Fluorescence In Situ Hybridization
4.4. Seed Storage Protein (Gliadin) Analysis
4.5. DNA Extraction, PCR Amplification, and DNA Sequencing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Slageren, M.W. Wild Wheats: A Monograph of Aegilops L. and Amblyopyrum (Jaub. et Spach) Eig (Poaceae); Wageningen Agricultural University: Wageningen, The Netherlands, 1994; p. 514. [Google Scholar]
- Kilian, B.; Mammen, K.; Millet, E.; Sharma, R.; Graner, A.; Salamini, F.; Hammer, K.; Özkan, H. Aegilops. Wild Crop Relatives: Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–76. [Google Scholar]
- Witcombe, J.R. A Guide to the Species of Aegilops l. Their Taxonomy, Morphology and Distribution; IBPGR Secretariat: Rome, Italy, 1983; p. 74. [Google Scholar]
- Abdolmalaki, Z.; Mirzaghaderi, G.; Mason, A.S.; Badaeva, E.D. Molecular cytogenetic analysis reveals evolutionary relationships between polyploid Aegilops species. Plant Syst. Evol. 2019, 305, 459–475. [Google Scholar] [CrossRef]
- Zhukovsky, P.M. A critical-systematical survey of the species of the genus Aegilops L. Bull. Appl. Bot. Genet. Plant Breed. 1928, 18, 417–609. (In Russian) [Google Scholar]
- Eldarov, M.; Aminov, N.; van Slageren, M. Distribution and ecological diversity of Aegilops L. in the Greater and Lesser Caucasus Regions of Azerbaijan. Genet. Resour. Crop. Evol. 2015, 62, 265–273. [Google Scholar] [CrossRef]
- Haruntyunyan, M.; Dulloo, M.E.; Yeritsyan, N.; Danielyan, A. Red List assessment of nine Aegilops species in Armenia. Genet. Resour. Crop. Evol. 2010, 57, 1177–1189. [Google Scholar] [CrossRef]
- Ohta, S.; Iwasaki, R.; Mori, N.; Ozkan, H. Geographical Distribution of Two Varieties of Aegilops neglecta and Ae. columnaris in Southern Turkey Revealed by the Field Researches from 2003 to 2005; Fukui Prefectural University: Eiheiji, Japan, 2006; pp. 38–43. [Google Scholar]
- Goryunova, S.V.; Chikida, N.N.; Kochieva, E.Z. RAPD analysis of the intraspecific and interspecific variation and phylogenetic relationships of Aegilops L. species with the U genome. Russ. J. Genet. 2010, 46, 841–854. [Google Scholar] [CrossRef]
- Kimber, G.; Feldman, M. Wild Wheat, An Introduction; Special Report 353; College of Agriculture University of Missouri: Columbia, SC, USA, 1987; p. 142. [Google Scholar]
- Resta, P.; Zhang, H.-B.; Dubcovsky, J.; Dvorak, J. The Origins of the Genomes of Triticum biunciale, T. ovatum, T. neglectum, T. columnare, and T. rectum (Poaceae) Based on Variation in Repeated Nucleotide Sequences. Am. J. Bot. 1996, 83, 1556. [Google Scholar] [CrossRef]
- Edet, O.U.; Gorafi, Y.S.A.; Nasuda, S.; Tsujimoto, H. DArTseq-based analysis of genomic relationships among species of tribe Triticeae. Sci. Rep. 2018, 8, 16397. [Google Scholar] [CrossRef] [PubMed]
- Badaeva, E.D.; Amosova, A.V.; Samatadze, T.E.; Zoshchuk, S.A.; Shostak, N.G.; Chikida, N.N.; Zelenin, A.V.; Raupp, W.J.; Friebe, B.; Gill, B.S. Genome differentiation in Aegilops. 4. Evolution of the U-genome cluster. Plant Syst. Evol. 2004, 246, 45–76. [Google Scholar] [CrossRef]
- Cui, L.; Ren, Y.; Murray, T.D.; Yan, W.; Guo, Q.; Niu, Y.; Sun, Y.; Li, H. Development of Perennial Wheat Through Hybridization Between Wheat and Wheatgrasses: A Review. Engineering 2018, 4, 507–513. [Google Scholar] [CrossRef]
- Kihara, H. Genomanalyse bei Triticum und Aegilops IX. Cytologia 1949, 14, 135–144. [Google Scholar] [CrossRef]
- Kihara, H. Considerations on the Evolution and Distribution of Aegilops Species Based on the Analyser-method. Cytologia 1954, 19, 336–357. [Google Scholar] [CrossRef]
- Kihara, H. Interspecific relationship in Triticum and Aegilops. Seiken Ziho 1963, 15, 1–12. [Google Scholar]
- Kimber, G.; Yen, Y. Hybrids involving wheat relatives and autotetraploid Triticum umbellulatum. Genome 1989, 32, 1–5. [Google Scholar] [CrossRef]
- Chennaveeraiah, M.S. Karyomorphologic and cytotaxonomic studies in Aegilops. Acta Hort. Gotobg. 1960, 23, 85–186. [Google Scholar]
- Badaeva, E.D.; Ruban, A.S.; Shishkina, A.A.; Sibikeev, S.N.; Druzhin, A.E.; Surzhikov, S.A.; Dragovich, A.Y. Genetic classification of Aegilops columnaris Zhuk. (2n = 4x = 28, UcUcXcXc) chromosomes based on FISH analysis and substitution patterns in common wheat × Ae. columnaris introgressive lines. Genome 2018, 61, 131–143. [Google Scholar] [CrossRef]
- Ogihara, Y.; Tsunewaki, K. Molecular basis of the genetic diversity of the cytoplasm in Triticum and Aegilops. I. Diversity of the chloroplast genome and its lineage revealed by the restriction pattern of ct-DNAs. Jpn. J. Genet. 1982, 57, 371–396. [Google Scholar] [CrossRef]
- Tsunewaki, K. Plasmon analysis as the counterpart of genome analysis. In Methods of Genome Analysis in Plant: Their Merits and Pitfalls; Jauhar, P.P., Ed.; CRC Press: Boca Raton, FL, USA, 1996; pp. 271–299. [Google Scholar]
- Kadosumi, S.; Kawahara, T.; Sasanuma, T. Multiple origins of U genome in two UM genome tetraploid Aegilops species, Ae. columnaris and Ae. triaristata, revealed based on the polymorphism of a genome-specific PCR fragment. Genes Genet. Syst. 2005, 80, 105–111. [Google Scholar] [CrossRef][Green Version]
- Tsunewaki, K.; Mukai, Y.; Endo, T.R.; Tsuji, S.; Murata, M. Genetic diversity of the cytoplasm in Triticum and Aegilops. V. Classification of 23 cytoplasms into eight plasma types. Jpn. J. Genet. 1976, 51, 175–191. [Google Scholar] [CrossRef]
- Kimber, G.; Tsunewaki, K. Genome Symbols and Plasma Types in the Wheat Group, Proceedings of the 7th International Wheat Genetics Symposium, Cambridge, UK, 13−19 July 1988; Miller, T.E., Koebner, R.M.D., Eds.; Institute of Plant Science Research: Cambridge, UK, 1988; pp. 1209–1210. [Google Scholar]
- Dvořák, J. Genome Analysis in the Triticum-Aegilops Alliance. In Proceedings of the 9th International Wheat Genetics Symposium, Saskatoon, Saskatchewan, 2−7 August 1998; pp. 8–11. [Google Scholar]
- Novoselskaya-Dragovich, A.Y.; Yankovskaya, A.A.; Badaeva, E.D. Alien introgressions and chromosomal rearrangements do not affect the activity of gliadin-coding genes in hybrid lines of Triticum aestivum L. × Aegilops columnaris Zhuk. Vavilov J. Genet. Breed. 2018, 22, 507–514. [Google Scholar] [CrossRef]
- Metakovsky, E.; Branlard, G.; Graybosch, R. Chapter 2—Gliadins of Common Wheat: Polymorphism and Genetics. In Gliadin and Glutenin: The Unique Balance of Wheat Quality; Wrigley, C., Békés, F., Bushuk, W., Eds.; AACC Internat: Saint Paul, MN, USA, 2006; pp. 35–84. [Google Scholar]
- Metakovsky, E.; Graybosch, R. Chapter 3—Gliadin Alleles in Wheat: Identification and Applications. In Gliadin and Glutenin: The Unique Balance of Wheat Quality; Wrigley, C., Békés, F., Bushuk, W., Eds.; AACC Internat: Saint Paul, MN, USA, 2006; pp. 85–114. [Google Scholar]
- Dudnikov, A.J. Polymorphism of gliadins in Aegilops tauschii Coss. local populations in two primary habitats in Dagestan. Genet. Resour. Crop. Evol. 2017, 65, 845–854. [Google Scholar] [CrossRef]
- Yan, Y.M.; Hsam, S.L.K.; Yu, J.Z.; Jiang, Y.; Zeller, F.J. Genetic polymorphisms at Gli-Dt gliadin loci in Aegilops tauschii as revealed by acid polyacrylamide gel and capillary electrophoresis. Plant Breed. 2003, 122, 120–124. [Google Scholar] [CrossRef]
- Konarev, V.G. Wheat Proteins; Kolos: Moscow, Russia, 1980; p. 351. (In Russian) [Google Scholar]
- Cole, E.W.; Fullington, J.G.; Kasarda, D.D. Grain protein variability among species of Triticum and Aegilops: Quantitative SDS-PAGE studies. Theor. Appl. Genet. 1981, 60, 17–30. [Google Scholar] [CrossRef]
- Medouri, A.; Bellil, L.; Khelifi, D. The genetic diversity of gliadins in Aegilops geniculata from Algeria. Czech J. Genet. Plant Breed. 2016, 51, 9–15. [Google Scholar] [CrossRef]
- Metakovsky, E.V. Gliadin allele identification in common wheat. II. Catalogue of gliadin alleles in common wheat. J. Genet. Breed 1991, 45, 325–344. [Google Scholar]
- Badaeva, E.D.; Friebe, B.; Gill, B.S. Genome differentiation in Aegilops. 2. Physical mapping of 5S and 18S–26S ribosomal RNA gene families in diploid species. Genome 1996, 39, 1150–1158. [Google Scholar] [CrossRef]
- Badaeva, E.D.; Amosova, A.V.; Muravenko, O.V.; Samatadze, T.E.; Chikida, N.N.; Zelenin, A.V.; Friebe, B.; Gill, B.S. Genome differentiation in Aegilops. 3. Evolution of the D-genome cluster. Plant Syst. Evol. 2002, 231, 163–190. [Google Scholar] [CrossRef]
- Yamamoto, M. Distribution of Ribosomal RNA Genes in Aegilops and Triticum Chromosomes; Bulletin of Kansai Women’s College: Osaka, Japan, 1992; Volume 2, pp. 25–37. (In Japanese) [Google Scholar]
- Yamamoto, M. Detection of Ribosomal RNA Genes in Aegilops by in Situ Hybridization; Bulletin of Osaka Private College Association: Osaka, Japan, 1992; Volume 29, pp. 77–82. (In Japanese) [Google Scholar]
- Badaeva, E.D.; Chikida, N.N.; Belousova, M.K.; Ruban, A.S.; Surzhikov, S.A.; Zoshchuk, S.A. A new insight on the evolution of polyploid Aegilops species from the complex Crassa: Molecular-cytogenetic analysis. Plant Syst. Evol. 2021, 307, 1–18. [Google Scholar] [CrossRef]
- Friebe, B.; Jiang, J.; Tuleen, N.; Gill, B.S. Standard karyotype of Triticum umbellulatum and the characterization of derived chromosome addition and translocation lines in common wheat. Theor. Appl. Genet. 1995, 90, 150–156. [Google Scholar] [CrossRef]
- Molnár, I.; Molnár-Láng, M. Visualization of U and M genome chromosomes by multicolour genomic in situ hybridization in Aegilops biuncialis and Triticum aestivum-Ae. biuncialis amphiploids. Acta Agron. Hung. 2010, 58, 195–202. [Google Scholar] [CrossRef]
- Molnár, I.; Kubaláková, M.; Šimková, H.; Cseh, A.; Molnár-Láng, M.; Doležel, J. Chromosome Isolation by Flow Sorting in Aegilops umbellulata and Ae. comosa and Their Allotetraploid Hybrids Ae. biuncialis and Ae. geniculata. PLoS ONE 2011, 6, e27708. [Google Scholar] [CrossRef]
- Molnár, M.L.; Vrána, J.; Burešová, V.; Cápal, P.; Farkas, A.; Darkó, É.; Cseh, A.A.; Kubaláková, M.; Molnár-Láng, M.; Doležel, J. Dissecting the U, M, S and C genomes of wild relatives of bread wheat (Aegilops spp.) into chromosomes and exploring their synteny with wheat. Plant J. 2016, 88, 452–467. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Dai, S.; Bao, T.; Zuo, Y.; Xiang, Q.; Li, J.; Liu, G.; Yan, Z. Analysis of Structural Genomic Diversity in Aegilops umbellulata, Ae. markgrafii, Ae. comosa, and Ae. uniaristata by Fluorescence In Situ Hybridization Karyotyping. Front. Plant Sci. 2020, 11, 710. [Google Scholar] [CrossRef] [PubMed]
- Badaeva, E.D.; Friebe, B.; Gill, B.S. Genome differentiation in Aegilops. 1. Distribution of highly repetitive DNA sequences on chromosomes of diploid species. Genome 1996, 39, 293–306. [Google Scholar] [CrossRef]
- Kawahara, T. Morphological and isozyme variation in genebank accessions of Aegilops umbellulata Zhuk., a wild relative of wheat. Genet. Resour. Crop. Evol. 2002, 49, 89–94. [Google Scholar] [CrossRef]
- Stebbins, L. Chromosomal Evolution in Higher Plants; Addison-Wesley: London, UK, 1971; p. 216. [Google Scholar]
- Dion-Côté, A.-M.; Barbash, D.A. Beyond speciation genes: An overview of genome stability in evolution and speciation. Curr. Opin. Genet. Dev. 2017, 47, 17–23. [Google Scholar] [CrossRef]
- Fishman, L.; Stathos, A.; Beardsley, P.M.; Williams, C.F.; Hill, J.P. Chromosomal Rearrangements and the Genetics of Reproductive Barriers Inmimulus (Monkey Flowers). Evolution 2013, 67, 2547–2560. [Google Scholar] [CrossRef] [PubMed]
- Gornicki, P.; Zhu, H.; Wang, J.; Challa, G.S.; Zhang, Z.; Gill, B.S.; Li, W. The chloroplast view of the evolution of polyploid wheat. New Phytol. 2014, 204, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, N.; Brassac, J.; Kilian, B.; Blattner, F.R. Dated tribe-wide whole chloroplast genome phylogeny indicates recurrent hybridizations within Triticeae. BMC Evol. Biol. 2017, 17, 1–16. [Google Scholar] [CrossRef]
- Gerlach, W.L.; Bedbrook, J.R. Cloning and characterization of ribosomal RNA genes from wheat and barley. Nucleic Acids Res. 1979, 7, 1869–1885. [Google Scholar] [CrossRef]
- Tang, Z.; Yang, Z.; Fu, S. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J. Appl. Genet. 2014, 55, 313–318. [Google Scholar] [CrossRef]
- Gerlach, W.; Dyer, T. Sequence organization of the repeating units in the nucleus of wheat which contain 5S rRNA genes. Nucleic Acids Res. 1980, 8, 4851–4865. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, H.; Xu, Y.; Li, Y.; Lang, T.; Yang, Z.; Li, G. Characterization of Chromosomal Rearrangement in New Wheat—Thinopyrum intermedium Addition Lines Carrying Thinopyrum—Specific Grain Hardness Genes. Agronomy 2019, 9, 18. [Google Scholar] [CrossRef]
- Bedbrook, J.; Jones, J.; O’Dell, M.; Thompson, R.; Flavell, R. A molecular description of telomeric heterochromatin in Secale species. Cell 1980, 19, 545–560. [Google Scholar] [CrossRef]
- Rayburn, A.L.; Gill, B.S. Isolation of a D-genome specific repeated DNA sequence from Aegilops squarrosa. Plant Mol. Biol. Rep. 1986, 4, 102–109. [Google Scholar] [CrossRef]
- Komuro, S.; Endo, R.; Shikata, K.; Kato, A. Genomic and chromosomal distribution patterns of various repeated DNA sequences in wheat revealed by a fluorescence in situ hybridization procedure. Genome 2013, 56, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Badaeva, E.D.; Badaev, N.S.; Gill, B.S.; Filatenko, A.A. Intraspecific karyotype divergence in Triticum araraticum (Poaceae). Plant Syst. Evol. 1994, 192, 117–145. [Google Scholar] [CrossRef]
- Badaeva, E.D.; Ruban, A.S.; Aliyeva-Schnorr, L.; Municio, C.; Hesse, S.; Houben, A. In Situ Hybridization to Plant Chromosomes. In Fluorescence In Situ Hybridization (FISH) Application Guide; Liehr, T., Ed.; Springer: Berlin, Germany, 2017; pp. 477–494. [Google Scholar]
- Metakovsky, E.V.; Novoselskaya, A.Y. Gliadin allele identification in common wheat 1. Methodological aspects of the analysis of gliadin patterns by one-dimensional polyacrylamide gel electrophoresis. J. Genet. Breed. 1991, 45, 317–324. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Accession # | Duplicates | Country of Origin | Collection Site | Latitude (N) | Longitude (E) | Alt. (h, m) | Analyzed by FISH |
|---|---|---|---|---|---|---|---|---|
| 1 1 | K−1178 | - | Armenia | Aznaburt vil., near Dash Agl mountain | 39.4333 | 45.2833 | 1600 | - |
| 2 1 | K−1193 | - | Armenia | Abovyan, near village of Shor-Bulakh | 40.1333 | 45.6333 | 1300 | + |
| 3 1 | K−1495 | IG 48026 | Armenia | Naxchivan, Djulfinskii reg., N of Arbakunis vil. | 39.1167 | 45.6333 | 1350 | - |
| 4 | K−1512 | AE 1188 | Armenia | near Erevan, valley of Razdan river | 40.2 | 44.5333 | 136 | - |
| 5 1 | K−2344 | - | Armenia | Ekhegnadzor reg., along Elpin-Agavnadzor road | 39.7833 | 45.1833 | 776 | - |
| 6 1 | K−4224 (3) | IG 48738 | Armenia | Erevan region, 2 km SE of Jrvezh | 40.1667 | 44.6 | 780 | - |
| 7 1,2 | k−4228 | IG 48757 | Armenia | Shorap, 20 km W of Erevan | 40.25 | 44.3333 | 1350 | - |
| 8 | K−4229 | IG 126249 | Armenia | Abovyan reg., Erebuni Natural Reserve | 40.2833 | 44.6333 | 1072 | - |
| 9 1,2 | K−4225 | IG 48740 | Armenia | Outskirt NE Erevan | 40.1167 | 44.5167 | 1400.0 | - |
| 10 1 | K−4366 | IG 48745 | Armenia | Abovyan reg., NE of Erevan, N. Dzervesh; Gegadir; Muchavan | 40.2833 | 44.6333 | 1045 | - |
| 11 1 | K−4551 | - | Armenia | Vanadzor, after Gadzor | 39.7833 | 45.3667 | 363 | - |
| 12 | K−564 | - | Armenia | Azizbekovskii reg., around vil. Khandzorut | 39.55 | 45.35 | 1685 | + |
| 13 | NAZ (3) | - | Armenia | near the village of Urznadzor | - | - | - | - |
| 14 | PI 499258 | - | unknown | obtained from China | - | - | - | - |
| 15 | PI 574457 | K−512; AE111 | Azerbaijan | unknown | - | - | - | - |
| 16 | IG 48818 | - | Iran | Damavand | 35.7333 | 52.0667 | 2474 | - |
| 17 1 | K−4240 | IG 49138 | Iran | 10 km SW Horand from Ahar | 38.75 | 47.1667 | 1110 | - |
| 18 1,2 | K−4413 * | IG 49087 | Iran | 20 km W Takestan to Zia Abad road to Zanjan | 36.0333 | 49.5 | 1320 | - |
| 19 | K−4418 | IG 49107 | Iran | 31 km Urumiyeh to Oshnaviyeh Kazem Lo Valley | 37.25 | 45.1333 | 1380 | - |
| 20 | K−3899 II | IG 49010 | Iraq | Ninawa; Jebel Maqloub near Deir Matti | 36.5 | 43.4167 | 850 | - |
| 21 | IG 49067 | - | Lebanon | Rachaiya, 1 km E of Aita Al Foukhar | 33.6333 | 35.9 | 1350 | - |
| 22 | K−4003 | IG 48072 | Lebanon | Terbol region W slope Anti Lebanon Mts. (zone A) | 33.9 | 36.1 | - | - |
| 23 | K−4004 | IG 48091 | Lebanon | Sanin region E slope Lebanon Mts. (zone B) | 33.9333 | 35.8333 | - | - |
| 24 | K−4007 | IG 48107 | Lebanon | Irsal region; W slope Anti Lebanon Mts. (zone C) | 34.25 | 36.6667 | 0 | - |
| 25 1 | K−4406 | IG 49047 | Lebanon | Baalbek 4 km W Baalbek road to Bcharre laat vil. | 34.0333 | 36.1667 | 1050 | - |
| 26 1 | K−4241a | i−611188 | Lebanon | Sanin region E slope Lebanon Mts. (zone B) | 33.9333 | 35.8333 | - | - |
| 27 1 | K−4241b | i−611189 | Lebanon | Al Alia; 40 km N of Karak | 31.95 | 35.9333 | 800 | - |
| 28 | K−4407 | IG 49047 | Lebanon | Baalbek 4 km W Baalbek road to Bcharre Iaat vil. | 34.0333 | 36.1667 | 1050 | - |
| 29 2 | K−4409 | IG 49053 | Lebanon | 3 km from Deir Ahmar road to Ain Ata | 34.1333 | 36.1 | 1370 | - |
| 30 1 | K−2680 | PI 487198 | Syria | 7 km from Atareb to Qalaat Samaan, Aleppo | 36.2022 | 36.7758 | 460 | - |
| 31 | K−4009 | i−571713 | Syria | Al Hasakah; just N of Jabal Abd El-Aziz | 36.4667 | 40.3333 | 600 | + |
| 32 1 | K−4362 | IG 48729 | Syria | Damascus May Saloun; 4 km before Tukeya | 33.6 | 36.0667 | 1468 | - |
| 33 1 | K−4372 | IG 48800 | Syria | 2 km NE of Sa’an road from Shabki | 32.7 | 36.8417 | 1400 | - |
| 34 | PI 487196 | - | Syria | Aleppo Province | 36.1667 | 36.8333 | 450 | - |
| 35 | CIae 34 | - | Turkey | - | - | - | - | - |
| 36 | K−4002 | IG 47875 | Turkey | 14 km NW Keskin | 39.7167 | 33.4333 | 520 | - |
| 37 | #1 | 2006−6−25−8−2 | Turkey | 132 km NW from Nevşehir | 39.1622 | 33.9325 | 1060 | - |
| 38 | #10 | 2006−6−21−5−1 | Turkey | 50 km NE from Kilis to Gaziantep | 37.315 | 37.7347 | 510 | - |
| 39 | #2 | 2006−6−17−7−2 | Turkey | 29 km NE from Kilis to Gaziantep | 36.9242 | 37.0786 | 730 | - |
| 40 | #3 | 2006−6−21−12−1 | Turkey | 57 km NE from Kilis to Gaziantep | 37.2678 | 37.5208 | 700 | - |
| 41 | #4 | 2006−7−12−2 | Turkey | 34 km NE from Kilis to Gaziantep | 37.09667 | 37.0406 | 970 | - |
| 42 | #6 | 2006−6−21−9−1 | Turkey | 54 km NE from Kilis to Gaziantep | 37.3739 | 37.8458 | 700 | - |
| 43 | #7 | 2006−6−17−10−2 | Turkey | 32 km NE from Kilis to Gaziantep | 36.9944 | 37.9664 | 950 | - |
| 44 | #8 | 2006−6−25−6−3 | Turkey | 114 km NW from Nevşehir | 39.0442 | 34.0414 | 880 | - |
| 45 | #9 | 2006−6−21−1−2 | Turkey | 46 km NE from Kilis to Gaziantep | 37.2139 | 37.4903 | 831 | - |
| 46 1 | i−570045 | PI 554184 | Turkey | Kars, 5 km S Sivas Malatya border | 39.7333 | 37.05 | 1500 | + |
| 47 | PI 276968 | - | Turkey | Konya | 37.8333 | 32.5 | - | - |
| 48 | PI 486281 | IG 46886 | Turkey | 42 km southeast of Ercis-Karayollari Bakimevi | 38.9167 | 43.6 | 1700 | + |
| 491 | PI 542171 | - | Turkey | 19 km north of Gaziantep toward Yavuzeli | 37.1833 | 37.4667 | 800 | + |
| 50 1,2 | PI 542191 II | - | Turkey | Aegean Agric. Research Inst. Gene Bank, Menemen | - | - | 30 | + |
| 51 | PI 554178 | IG 47040 | Turkey | 22 km north of Van | 38.7 | 43.3333 | 1734 | + |
| 52 | PI 554180 | IG 46997 | Turkey | 35 km west of Tuzluca | 40.15 | 43.3667 | 1010 | - |
| 53 | PI 554181 | IG 47042 | Turkey | Aydin, 10 km north of Kusadasi | 37.9167 | 27.2833 | 130 | + |
| 54 | PI 554182 II | IG 47048 | Turkey | Icel, 24 km southwest of Erdemli, Mersin across from Boy Scout Recreation Center | 36.4667 | 34.1333 | 30 | - |
| 55 | PI 554185 | IG 47117 | Turkey | 2 km southeast of Van on route to Gurpinar | 38.5 | 43.3667 | 1790 | - |
| 56 1,2 | PI 554186 | IG 47166 | Turkey | Van, 6 km southeast of Van | 38.4667 | 43.3833 | 1990 | - |
| 57 1 | PI 554187 (2) | IG 47125 | Turkey | Van, 2 km north of Van | 38.5333 | 43.3333 | 1710 | - |
| 58 | PI 554188 | - | Turkey | Van, 29 km north of Van | 38.75 | 43.3667 | 1790 | - |
| 59 | PI 554190 | IG 47170 | Turkey | Van, 29 km north of Van | 38.75 | 43.3667 | - | - |
| 60 | PI 560506 | - | Turkey | Roadside along Lake Van. About 3 km W of Ermisler | 38.8667 | 43.4667 | 1630 | - |
| 61 | PI 560507 | - | Turkey | Van, About 2 km N of village of Yalnizagac | 38.7 | 43.5 | 1837 | - |
| 62 2 | PI 564179 II | - | Turkey | 23 km southeast of Manavgat, near Okucalar village | 36.6833 | 31.6333 | 50 | + |
| 631,2 | PI 564180 II | - | Turkey | 58 km southwest of Silifke; 4 km west of Ovacik, Mersin | 36.18333 | 33.6333 | 250 | + |
| 64 1,2 | PI 564181 II | - | Turkey | 49 km southwest of Silifke, Mersin | 36.2 | 33.7 | 150 | + |
| 65 1,2 | TA 2084 II | - | Turkey | 1 km N of Iskenderum (Alexandretta) | 36.6001 | 36.1969 | 50 | + |
| 66 | TA 2106 | KU11−2 | Turkey | Konya, collected by Dr. Johnson in 1965. | 37.8667 | 32.4833 | 1030 | - |
| 67 | AE 1521 | - | unknown | - | - | - | - | - |
| 68 | AE 1607 (2) | - | unknown | obtained from UK | - | - | - | + |
| 69 | TX 01 | - | unknown, | provided by Dr. M. Feldman | - | - | - | - |
| Aegilops neglecta | ||||||||
| 70 | PI 564182 * | - | Turkey | 9 km southeast of Ayvacik | 39.583333 | 26.483333 | 420 | + |
| 71 1 | K−4553 * | IG 126975 | Armenia | Kapan distr. road from Kapan to Charaten | 39.1903 | 46.43 | 970 | - |
| 72 2 | PI 170209 | - | Turkey | 17 km south of Canakkale | 40.033333 | 26.35 | 100 | + |
| 73 2 | AE 646 | - | Algeria | unknown | - | - | - | + |
| Aegilops umbellulata | ||||||||
| 74 2 | AE 155 | K−1234 | Azerbaijan | unknown | - | - | - | + |
| 75 2 | AE 820 | - | Turkey | 3 km E Kemalpasa | - | - | - | - |
| 76 2 | AE 1339 | - | Greece | Kreta | - | - | - | + |
| No. | Trans. Code | Translocation Type | Structure of Translocated Chromosomes | Accessions | Origin |
|---|---|---|---|---|---|
| 1 | inv1 | perInv2Xc | - | PI 542171; AE 1607b | Turkey |
| 2 | inv2 | perInv 6Xc | - | K−4406 | Lebanon |
| 3 | inv3 | perInv 7Uc | - | i−570045; CIae 34; K−4002; TA2106; AE 1521; H−8; TX01 | Turkey |
| 4 | T1 | perInv7Uc + T4Uc:4Xc | 4UcS:4XcS + 4UcL:4XcL | H−2 | Turkey |
| 5 | T2 | perInv2Xc−2 + T6Uc:7Xc | perInv2Xc−2 + 6UcS.6UcL−7XcS +T6UcL−7XcS.7XcL | AE 1607a | unknown |
| 6 | T3 | T3Uc:4Xc | 3UcS:4XcL + 3UcL:4XcS | K−4224c; K−564 | Armenia |
| 7 | T4 | T3Xc:7Xc | 3XcL.3XcS−7XcS + 3XcS−7XcS.7XcL | PI 276968 PI 574457 (K−512) | Turkey Azerbaijan |
| 8 | T5 | T5Uc:6Xc | 5UcS:6XcL + 5UcS:6XcL | K−4418 | Iran |
| 9 | T6 | T2Uc:2Xc | 2UcS.2UcL−2XcL + 2UcL−2XcL.2XcS | PI 554185 | Turkey |
| 10 | T7 | T2Uc:4Xc | 2UcS:4XcL + 2UcS:4XcL | PI 554187t | Turkey |
| 11 | T8 | T3Uc:1Xc | 3UcS.3UcL−1Xc + 3UcL−1XcS.1XcL | K−560506 | Turkey |
| 12 | T9 | T4Uc:5Uc | 4UcL.4UcS−5UcL + 4UcS−5UcL.5UcS | K−4002 | Turkey |
| 13 | T10 | T7Uc:2Xc | 7UcS. 7UcL−2Xc + 7UcL−2XcL.2XcS | K−4003; K−4407 | Lebanon |
| 14 | T11 | T1Uc:5Uc | 1UcS:5UcS + 1UcL:5UcL | PI 499258; K−1178; K−1495; K−4224; K−4366 | Armenia |
| 15 | T12 | T1Uc:5Uc + T4Uc:6Uc | 1UcS:5UcS + 1UcL:5UcL + 4UcL.4UcS−6UcS + 4UcS−6UcS.6UcL | K−1193 | Armenia |
| 16 | T13 | T1Uc:5Uc + T3Uc:1Xc | 1UcS:5UcS + 1UcL:5UcL + 3UcS.3UcL−1Xc + 3UcL−1XcS.1XcL | PI 554180 | Turkey |
| 17 | T14 | T2Xc:4Xc | 2XcS:4XcS + 2XcL:4XcL | PI 560507 | Turkey |
| 18 | T15 | T2Xc:4Xc:6Xc | 2XcS:4XcS + 2XcL:6XcS + 6XcL:4XcL | K−4240 | Iran |
| 19 | T16 | T1Uc:5Uc + T3Uc:5Xc | 1UcS:5UcS + 1UcL:5UcL + 3UcS:5XcL + 3UcL:5XcS | K−4224B; K−4228 | Armenia |
| 20 | T17 | T1Uc:5Uc + T7Uc:3Xc + T3Uc:4Uc | 1UcS:5UcS + 1UcL:5UcL + 7UcS:3XcS + 7UcL:3XcL + 3UcS:4UcS + 3UcL:4UcL | K−4225; K−4551 | Armenia |
| 21 | T18 | T2Uc:4Xc + T4Uc:6Xc | 2UcS:4XcS + 2UcL:4XcL + 4UcS:6XcS + 4UcL:6XcL | K−4372 | Syria |
| 22 | T19 | T6Uc:4Xc + T7Uc:3Xc | 6UcS:4XcS + 6UcL:4XcL + 7UcS:3XcL + 7UcL:3XcS | PI 486281; PI 554181 | Turkey |
| 23 | T20 | T2Uc:5Xc + T4Uc:2Xc | 2UcL.2UcS−5Xc + 2UcS−5XcL.5XcS + 4UcS:2XcL + 4UcL:2XcS | K−4362 | Syria |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badaeva, E.D.; Chikida, N.N.; Fisenko, A.N.; Surzhikov, S.A.; Belousova, M.K.; Özkan, H.; Dragovich, A.Y.; Kochieva, E.Z. Chromosome and Molecular Analyses Reveal Significant Karyotype Diversity and Provide New Evidence on the Origin of Aegilops columnaris. Plants 2021, 10, 956. https://doi.org/10.3390/plants10050956
Badaeva ED, Chikida NN, Fisenko AN, Surzhikov SA, Belousova MK, Özkan H, Dragovich AY, Kochieva EZ. Chromosome and Molecular Analyses Reveal Significant Karyotype Diversity and Provide New Evidence on the Origin of Aegilops columnaris. Plants. 2021; 10(5):956. https://doi.org/10.3390/plants10050956
Chicago/Turabian StyleBadaeva, Ekaterina D., Nadezhda N. Chikida, Andrey N. Fisenko, Sergei A. Surzhikov, Maria K. Belousova, Hakan Özkan, Alexandra Y. Dragovich, and Elena Z. Kochieva. 2021. "Chromosome and Molecular Analyses Reveal Significant Karyotype Diversity and Provide New Evidence on the Origin of Aegilops columnaris" Plants 10, no. 5: 956. https://doi.org/10.3390/plants10050956
APA StyleBadaeva, E. D., Chikida, N. N., Fisenko, A. N., Surzhikov, S. A., Belousova, M. K., Özkan, H., Dragovich, A. Y., & Kochieva, E. Z. (2021). Chromosome and Molecular Analyses Reveal Significant Karyotype Diversity and Provide New Evidence on the Origin of Aegilops columnaris. Plants, 10(5), 956. https://doi.org/10.3390/plants10050956