Abstract
Prostate cancer, a slow-growing tumor, develops through the over-proliferation of malignant cells in the prostate and is one of the most common types of cancer. Active surveillance, radical prostatectomy, external beam radiation, brachytherapy, cryotherapy, stereotactic body radiation therapy, hormone therapy, and chemotherapy are common treatment strategies for prostate cancer. However, resistance to treatment in advanced prostate cancer is a concerning issue in the use of these therapies. Immune checkpoint inhibitor (ICI) therapy for prostate cancer is an emerging strategy for the treatment of advanced prostate cancers but the resistance and limited efficacy to ICIs observed in metastatic castration-resistant prostate cancer (mCRPC) raises concerns. The ongoing clinical trials for combination therapies for mCRPC have provided some hope. This review concisely discusses the molecular and cellular mechanisms, immunotherapy, the limitations of ICIs, combination therapies, and the prospects of developing novel therapeutics for prostate cancer.
1. Introduction
According to the American Cancer Society, about 42 out of 100 men living in the United States will develop cancer at some point during their lifetime, and prostate cancer (PCa) is the most common cancer diagnosed in men, with one in eight men expected to develop PCa during their lifetime. The American Urological Association (AUA) reported approximately 288,300 newly diagnosed cases of PCa and 34,700 deaths related to PCa in the U.S. in 2023 [1]. As per the European Urology Association (EUA), the estimated age-standardized rate (ASR) of PCa was 31 per 100,000 males and the lifetime cumulative risk was 3.9%, with the highest all-age incidence ASR in Northern Europe (83 per 100,000), followed by Western Europe. In Europe, the all-age incidence ASR was 63 per 100,000 males in the year 2020, the lifetime cumulative risk was 16%, and all-age mortality ASR was 11 per 100,000 males varying in different regions of Europe [2]. PCa, which usually grows very slowly, is the fifth leading cause of overall death worldwide and the second leading cause of cancer-related death among men in the United States [3]. Aging is strongly related to the incidence and mortality rates of PCa, with >65 years of age associated with the highest incidence. African American men have more aggressive tumors and higher incidence compared to white men. In addition to aging, a diet rich in saturated animal fat and red meat, lower intake of fruits, vegetables, vitamins, and coffee, hyperglycemia, obesity, physical inactivity, chronic inflammation, infections, family history, endogenous hormones, and exposure to chemicals (insecticides, herbicides, and other organic compounds) or ionizing radiation are common risk factors for PCa [3,4]. A retrospective analysis including 955 patients (395 with PCa and 238 with advanced tumor) suggested metabolic syndrome and physical inactivity as risk factors for PCa. The study concluded that metabolic syndrome is an independent predictor of high-grade PCa [5]. However, the exact cause of PCa is largely unknown. The early stages of PCa are asymptomatic, while difficulty urinating, frequent urination, blood in the urine or semen, pain or discomfort during urination, and erectile dysfunction are common symptoms when they do occur at later stages. Digital rectal exam, prostate-specific antigen, and biopsy are the methods of diagnosis, while active surveillance (for less aggressive tumors), radical prostatectomy, external beam radiation, brachytherapy, cryotherapy, stereotactic body radiation therapy, hormone therapy, and chemotherapy are common treatment options for PCa [6,7].
Targeted therapies, including hormonal treatment, chemotherapy, radiotherapy, and immunotherapy, are used in advanced and metastatic prostate cancers. The use of hormone therapy and radiotherapy in prostate cancer treatment is associated with side effects including erectile dysfunction, urinary incontinence, decreased libido, hot flashes, bone loss, muscle loss, blood lipid changes, insulin resistance, and bowel problems [8]. Targeting prostate-specific membrane antigen (PSMA), a transmembrane glycoprotein that is specific and highly expressed in PCa, has shown significant progress in the last two decades. Radioligand therapy (177Lu-PSMA-RLT, 225Ac-PSMA-RLT), antibody–drug conjugates (MLN2704, PSMA-MMAE, MEDI3726), cellular immunotherapy (CAR-T, CAR/NK-92, PSMA-targeted BiTE), PSMA-targeted photodynamic therapy, PSMA-PET, PSMA-targeted RGS, and multimodal PSMA-targeted imaging guided therapy are common PSMA-targeted therapies with encouraging anti-tumor activity in PCa [9]. However, the side effects associated with PSMA targeted therapy warrant further research and clinical trials. Gonadotropin-releasing hormone (GnRH) agonists (Degarelix) and GnRH antagonists (buserelin, goserelin, leuprorelin, and triptorelin) are initial hormonal therapies for PCa but these are associated with an increased risk of adverse cardiovascular events. Cicione et al. [10] reported a higher association of cardiovascular events, including hypertension, thrombosis, and myocardial infarction, with GnRH antagonists compared to GnRH agonists. Thus, these drugs should be prescribed with precaution in PCa patients with cardiovascular complications.
Newer therapies, including high-intensity focused ultrasound, docetaxel, cabazetaxel (for advanced hormone-refractory prostate cancer), abiraterone (an oral androgen biosynthesis inhibitor for the treatment of advanced metastatic castrate-resistant prostate cancer), denosumab (a monoclonal antibody), olaparib (PARP (poly(ADP-ribose) polymerase) inhibitor; targeted therapy), and sipuleucel-T (a new vaccine; immunotherapy) are emerging therapies for metastatic and resistant tumors [6,7]. The randomized, double-blind, phase III KEYNOTE-921 trial reported no significantly improved efficacy after the addition of pembrolizumab to docetaxel for the treatment of metastatic castration-resistant prostate cancer (mCRPC) [11]. A recent study reported a long-term process for the recovery of total testosterone levels after androgen-deprivation therapy cessation [12]. The therapeutic response of systemic therapy in PCa is usually transient, and invasive disease is associated with high mortality rates. This suggests that even the newer therapies are associated with side effects or are not sufficient for refractory PCa or the recurrence of PCa, and there is a need for a more robust therapeutic strategy for treatment-resistant advanced PCa. The emergence of immunotherapy as an efficacious and non-toxic alternative treatment is because of its immune-targeted molecular traits, which can achieve the maximum clinical benefit. This review summarizes the recent developments in immunotherapy for PCa.
2. Molecular Mechanisms of PCa Progression and Immunosuppression
Understanding the molecular mechanisms underlying PCa is important because immunotherapy uses the body’s own immune system to fight cancer by stimulating or enhancing the immune system’s ability to recognize and destroy cancer cells. Inflammation and immune cells play a critical role in the pathogenesis of PCa. The role of CD3+, CD8+, and CD4+ T cells; B-cells; tumor-associated macrophage (TAM); dendritic cells (altered function due to the increased secretion of transforming growth factor (TGF)-β and interleukin (IL)-10)); immunosuppressive immune cells including T regulator cells (Treg), myeloid-derived suppressor cells (MDSC), Th17 cells, macrophages, mast cells, and neutrophils involving IL-2; interferon (IFN)-γ; IL-1β; IL-1α; IL-1RA; IKKα, a signal transducer and activator of transcription 3 (STAT3); BMI1, C-X-C chemokine ligand (CXCL)13; and C-C chemokine ligand (CCL)2 in the pathogenesis, progression, and aggressiveness of PCa has been discussed [13]. Tumor cells, stromal cells, and other immune cells in the tumor microenvironment (TME) secrete TGF-β, contributing to the attenuated function of Tregs, CD8+ T-cells, which activate ERK and promote epithelial–mesenchymal transition (EMT), leading to the growth and increased invasiveness of PCa (Figure 1).
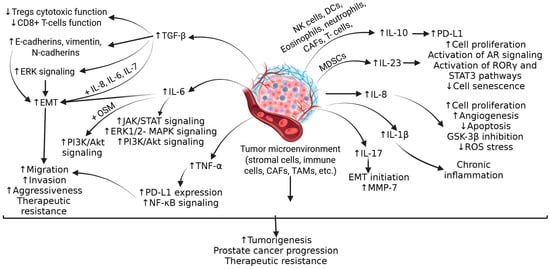
Figure 1.
The tumor microenvironment (TME) induces and contributes to an immunosuppressive environment in advanced prostate cancer. The cytokines, growth factors, and activation of downstream signaling pathways contribute to increased tumorigenesis, tumor progression, and therapeutic resistance. IL—interleukin; Tregs—T regulatory cells; ERK—extracellular signal-regulated kinase; EMT-epithelial–mesenchymal transition; OSM—oncostatin-M; PI3K—phosphoinositide 3-kinase; Akt—also known as protein kinase B; MAPK-mitogen activated protein kinase; JAK/STAT—Janus kinase, a signal transducer and activator of transcription; TGF-β—transforming growth factor beta; TNF-α—tumor necrosis factor alpha; PD-L1—programmed cell death ligand 1; NF-kB- nuclear factor kappa beta; CAFs—carcinoma-associated fibroblasts; TAMs—tumor associated macrophages; MDSCs—myeloid-derived stem cells; NK cells—natural killer cells; DC—dendritic cells; AR—androgen receptor; MMP—matrix metalloproteinase; ROS—reactive oxygen species; ROR γ—RAR-related orphan receptor gamma.”↑”—increased expression; “↓”—decreased expression.
IL-6 secreted by TME cells activates Janus kinase-signal transducer and activator of transcription (JAK/STAT), extracellular signal-regulated kinase (ERK)1/2-mitogen activated protein kinase (MAPK), and phosphoinositide 3-kinase (PI3K) pathways, and contributes to cell proliferation, EMT, tumor migration, increasing its growth and aggressiveness, and resistance to chemotherapy by inducing anti-apoptotic pathways. (Figure 1). IL-10, an anti-inflammatory cytokine secreted by various immune cells in TME cells, is correlated with poor prognosis in PCa and contributes to immunosuppression via increased expression of PD-L1. Another cytokine, IL-23, secreted by MDSCs, activates STAT3, RAR-related orphan receptor gamma (RORγ), and androgen receptor signaling, leading to cell proliferation, CRPC growth and metastasis, and treatment resistance to anti-androgen therapy (Figure 1). Cytokines such as IL-8, IL-1β, IL-17, IFN-γ, and tumor necrosis factor (TNF)-α play a critical role in cell proliferation, chronic inflammation, modulating apoptosis, angiogenesis, and the progression of PCa (Figure 1). It should be noted that the increased expression of TGF-β, IL-6, IL-10, and IL-23 is associated with poor prognosis in PCa [14,15]. Furthermore, along with these cytokines, CC and C-X-C chemokines including CCL2, CCL22, CCR4, CCL17, CXCL2, CXCR2, CXCL8, CXCL13, CXCL5, CXCR1, CXCR2, CXCL1, CXCL2, CXCL12, and CXCR4 play a critical role in pathogenesis, tumor heterogeneity, and immunosuppressive TME in PCa (Figure 2) [15].
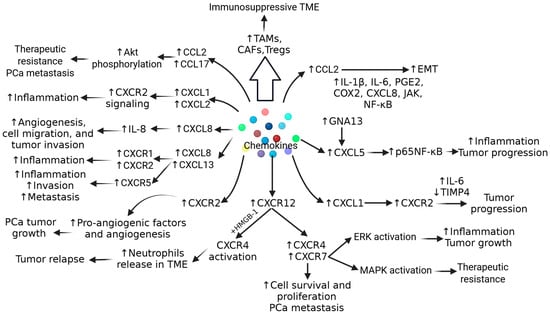
Figure 2.
Role of chemokines in progression, tumorigenesis, and therapeutic resistance in prostate cancer (PCa). IL—interleukin; Tregs—T regulatory cells; ERK—extracellular signal-regulated kinase; EMT—epithelial–mesenchymal transition; Akt—also known as protein kinase B; MAPK—mitogen activated protein kinase; JAK—Janus kinase; NF-kB—nuclear factor kappa beta; CAFs—carcinoma associated fibroblasts; TAMs—tumor-associated macrophages; TIMP—tissue inhibitor of matrix metalloproteinase; TME—tumor microenvironment; CXCL—C-X-C motif ligand; CXCR—C-X-C motif receptor; CCR—C-C motif chemokine receptor; CCL—C-C motif chemokine ligand; COX2—cyclooxygenase 2; PGE2—prostaglandin E2; HMGB1—high-mobility group box protein 1. “↑”—increased expression; “↓”—decreased expression.
In addition to immune cells, TAMs, MDSCs, and carcinoma-associated fibroblasts (CAFs) present in the tumor microenvironment (TME) also contribute to tumor progression and aggressiveness. Fibroblasts secrete basic fibroblast growth factor (bFGF), platelet-derived growth factor (PDGF), vascular endothelium growth factor (VEGF), TNF-α, and MMPs, increasing migration and angiogenesis in tumors [16]. CAFs are also associated with a phenotypic change in cancer cells, causing them to act as stem cells undergoing epithelial–mesenchymal transition (EMT). CAF also contributes to fibrous stroma formation, causing hypoxia and chronic inflammation in the tumor microenvironment. Activated Tregs contribute to immunosuppressive TME and its maintenance by attenuating the antitumor immune response mediated by T cells, tumor-infiltrating lymphocytes, natural killer (NK) cells, and neutrophils. MDSCs, after binding to CXCL5 and IL-8-mediated chemotaxis, are recruited to the TME and contribute to immunosuppression via the activation of arginase 1, nitrous oxide synthase, and indoleamine 2,3-dioxygenase. The predominance of M2-like macrophages in TME reduces the T-cell function and contributes to tumor progression and immunosuppression (Figure 3) [17]. The immunosuppressive environment and attenuation of antitumor immune response can also be mediated through the epithelial immune-like transition of cancer cells mediated by various cytokines, receptors, Ig motifs, transcription factors, and immune checkpoint molecules, including PD-1, PD-L1 (B7-H3, modulating T lymphocyte activity), B7-H4 (expressed on activated T-cells, B-cells, dendritic cells, and monocytes, modulating the inhibition of T cells’ proliferation and cytokine production), T-cell immunoglobulin 3 (TIM-3, negatively regulating Th1 cell activity), lymphocyte-activation gene 3 (LAG3, binds to HLA class II and inhibits T-cell function), and cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), restricting early T-cell activation [18] (Figure 3). Tregs play a critical role in the pathogenesis of PCa and are an attractive therapeutic target. Tregs suppress the function of naive T cells, B and T lymphocytes, dendritic cells, NK cells, and macrophages. The inhibition of immune response via Tregs is mediated through modulating and regulating cytokine release, apoptosis, antigen-presenting cells (APCs), and ATP degradation [19]. Tregs contribute to the growth, migration, and invasion of PCa via the increased secretion of IL-10, TGF-β, and IL-35, leading to the suppression of immune response, T cells’ exhaustion, and the increased expression of neoantigens such as lymphocyte activation gene-3 (LAG3), T-cell immunoglobulin and mucin domain 3 (TIM3), PD-1, and PD-L1 (Figure 3).
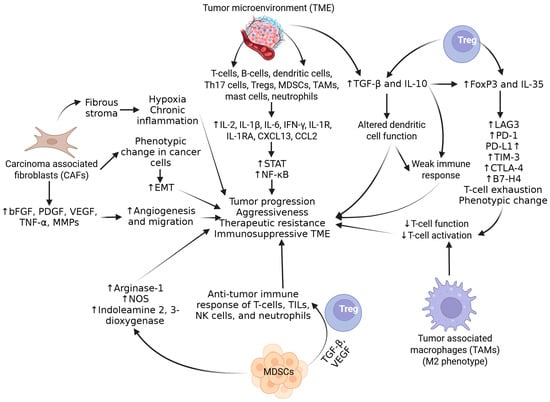
Figure 3.
Role of tumor microenvironment components in inducing immunosuppression and therapeutic resistance. IL—interleukin; Tregs—T regulatory cells; EMT—epithelial–mesenchymal transition; STAT—signal transducer and activator of transcription; TGF-β—transforming growth factor beta; TNF-α—tumor necrosis factor alpha; PD-1—programmed cell death protein 1; PD-L1—programmed cell death ligand 1; NF-kB—nuclear factor kappa beta; CAFs—carcinoma-associated fibroblasts; TAMs—tumor-associated macrophages; MDSCs—myeloid-derived stem cells; NK cells—natural killer cells; MMPs—matrix metalloproteinases; IFN γ—interferon-gamma; TME—tumor microenvironment; CXCL—C-X-C motif ligand; CCR—C-C motif chemokine receptor; TILs—tumor-infiltrating lymphocytes; NOS—nitric oxide synthetase; VEGF—vascular endothelial growth factor; PDGF—platelet-derived growth factor; bFGF—basic fibroblast growth factor; LAG3—lymphocyte activation gene-3; TIM3—T-cell immunoglobulin and mucin domain 3; CTLA-4—cytotoxic T-lymphocyte-associated protein 4. ”↑”—increased expression; “↓”—decreased expression.
DNA mutations cause abnormal cell growth and affect androgen receptor (AR) pathways, DNA repair pathways, and other pathways underlying PCa. Mutations, amplifications, and splicing variants in androgen receptor-related genes in AR pathways are common mechanisms underlying PCa. Failures in DNA repair can lead to genomic instability and tumor onset. Abnormalities in the cell cycle and PI3K pathway are associated with the onset and progression of PCa. Chromosomal alterations and rearrangements, the loss of phosphatase and tensin homolog (PTEN) and activation of phosphoinositide 3-Kinase/-mammalian target of rapamycin (PI3K/mTOR), the increased expression of interleukins and growth factors, global defects in apoptosis, and the loss of function of the prostate tumor-suppressor gene NKX3 are other molecular aberrations associated with PCa [20,21,22,23,24] (Figure 4). Co-regulator modification (e.g., GRHL2) [25], co-transcriptional R-loops-mediated epigenetic regulation [26], and intratumoral steroid synthesis, mainly androgens, after androgen deprivation therapy play a critical role in metastatic castration-resistant PCa (mCRPC) progression [27], and androgen promiscuity, the situation in which mutated androgen receptors may be activated by alternative ligands, including estrogen and other hormones [28,29,30], which contribute to the progression and pathogenesis of PCa (Figure 4). These mechanisms also contribute to resistance to treatment. Chromosomal abnormalities, including deletion, addition, and amplification, are frequently found in PCa. Further, a decreased copy number and loss of heterozygosity in chromosome 8p are also associated with PCa [31]. Telomere attrition also plays a role in the pathogenesis, greater genotype and phenotype heterogeneity, aggressiveness, decreased overall survival, increased biochemical recurrence, and increased risk of death in PCa [32,33].
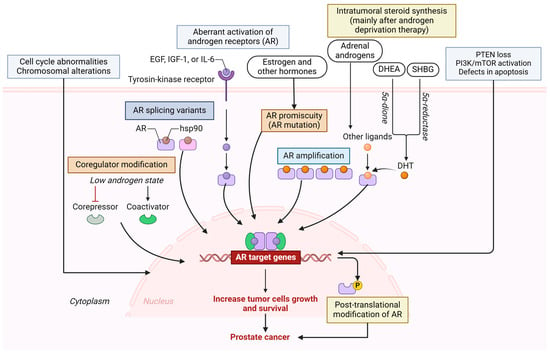
Figure 4.
Molecular mechanisms involved in pathogenesis and resistance to therapy in advanced PCa. Multiple mechanisms include aberrant androgen receptor (AR) expression, AR mutation, AR splice variants, AR amplification, cell cycle abnormalities, defective apoptosis, loss of PTEN, activation of PI3K/mTOR pathways, and other factors that increase the growth and survival of tumor cells and contribute to pathogenesis and resistance to treatment in advanced PCa. PTEN—phosphatase and tensin homolog; PI3K—phosphoinositide 3-Kinase; mTOR—mammalian target of rapamycin; HSP90—heat shock protein 90; EGF—epidermal growth factor; IL6—interleukin 6; IGF1—insulin-like growth factor 1; DHEA—dehydroepiandrosterone; SHBG—sex hormone binding globulin; DHT—dihydrotestosterone. Created in BioRender. Vikrant Rai. (2025) https://app.biorender.com/.
The role of TMPRSS2-ERG gene fusion, PTEN deletion, and MYC loss in the pathogenesis of PCa has been discussed. SLC45A3-ERG rearrangements co-occurring with TMPRSS2-ERG and in association with PTEN loss, the mutations in TP53 pathways, the mutation of the ORZAP1 gene, mismatch repair gene alteration at the level of MSH2 or MSH6 genes, germline DNA repair gene mutations (BRCA2, ATM, CHEK2, and BRCA1), CTNNB1 hot spot mutations, germline BRCA2 mutations, the genomic and epigenomic dysregulation of MED12L/MED12 axis, point mutations of TP53, SPOP, and FOXA1, the increased frequency of SPOP and ATM mutations, the loss of the retinoblastoma (RB) tumor-suppressor, the inactivation of LRF proto-oncogene, CDK12 deletion, and mutations in IDH1, MAP3K7, and CDH1 genes in the pathogenesis of PCa have been discussed [20,23]. Further, the rearrangement of MAG12 (PTEN inhibitor), BRAF, RAF, and CADM2, their overexpression and mutations in PIK3CA1, the deletion and downregulation of PHLPP1/2, a point mutation in Akt1, mutations in SPOP, CHD1, MLL2, ASH2L, and MED12, the overexpression of SPINK1, MYC, and NMYC, the elevated expression of EZH2 and BM1, the deletion of TAK1/MAP3K7, and the loss of TP53 also play a role in PCa pathogenesis [23].
Epigenetics plays a critical role in the pathogenesis of PCa. Dietary carcinogens (heterocyclic aromatic amines and polycyclic aromatic hydrocarbon due to red meat consumption), estrogens, and oxidants acting as a trigger for chronic inflammation promote PCa [24]. Hypermethylation in the promoter regions of tumor-suppressor genes leads to their inactivation in the pathogenesis of PCa. The most frequent somatic alteration in PCa is the aberrant methylation of the CpG island at the glutathione S transferase pi (GSTP1) locus in the presence of oxidative stress. However, this may also be positive in hyperplastic and normal tissue. GSTP1 polymorphisms have been associated with the risk of biochemical recurrence of PCa. Further, methylation of the Ras-association domain of the familial protein 1 isoform A (RASSF1A) located on chromosome 3p21 has also been reported [24]. Methylation has been reported in other genes, including androgen receptor, estrogen receptor 1 and 2, retinoic acid receptor β2, retinoic acid receptor responder 1, cyclin d2, cyclin-dependent kinase inhibitor 2a (p16), reprimo, stratifin (14-3-3 sigma), dickkopf 3, endothelin receptor type b, ras-association domain family protein 1 isoform a, runt-related transcription factor 3, secreted frizzled-related protein 1, familial adenomatous polyposis, caveolin 1, E-cadherin, cadherin 13, cluster-differentiation antigen 44, α-3 laminin, γ-3 laminin, TIMP metallopeptidase inhibitor 3, glutathione s-transferase m1, glutathione s-transferase p1, glutathione peroxidase 3, o-6-methylguanine DNA methyltransferase, apoptosis-associated speck-like protein containing a CARD, B cell lymphoma 2, death-associated kinase, multidrug-resistance receptor 1, and prostaglandin endoperoxidase synthase 2. Hypermethylation in genes involved in hormonal response, cell cycle control, signal transduction, tumor invasion, DNA damage repair, and apoptosis has also reported in PCa [34,35].
In addition to hypermethylation, post-transcriptional histone modifications involving histone deacetylases (HDACs, e.g., HDAC1), histone methyltransferases (HMTs, e.g., EZH2), and histone demethylases (HDMs, e.g., LSD1) also play a critical role in PCa. The enhancer of the zeste homolog 2 gene (EZH2) catalyzes the trimethylation of histone H3K27 and dimethylates H3K9, and its overexpression correlates with promoter hypermethylation and tumor aggressiveness. HDAC1 overexpression is associated with PCas containing TMPRSS2–ERG fusion. Lysine-specific demethylase 1 (LSD1) removes mono- or dimethyl groups from H3K4, acting as a transcriptional corepressor in PCa. Histone acetylation regulates androgen receptor regulation, and its activity is downregulated by HDAC1, HDAC2, HDAC3, and siRT1 [34]. The role of miRNAs, including miR-221/miR-222, miR-616, miR-125b, miR-488, miR-146a, miR-331-3p, miR-21, miR-34c, miR-34a, miR-15a, miR-16-1, miR-125b, miR-145, let-7c, and others, has also been discussed in the literature [34,35].
3. Immunotherapy for PCa
Immunotherapy is an emerging strategy for the treatment of advanced PCas that are resistant to other treatments, such as hormone therapy and chemotherapy. PCa showing a high tumor mutational burden, high programmed cell death protein 1 (PD-L1) tumor expression, cyclin-dependent kinase 12 (CDK12) mutations, mismatch repair-deficient (dMMR), a high microsatellite instability, homologous recombination deficiency, and (BRCA2 and ATM) POLE/POLD1 mutations, as well as patients in a good status of health, provide ideal candidates for immunotherapy because these changes increase response to immunotherapy by increasing the tumor mutation burden and neoantigen expression [36,37,38]. Immunotherapy uses patients’ own immune system to fight cancer cells while sparing healthy cells. Immunotherapy is effective because of its ability to recognize and eliminate newly developing cancer cells. Immunotherapy may also augment the therapeutic efficacy of other treatments when used in combination. The two main types of immunotherapies used for PCa are cancer vaccines targeting tumor-specific antigens and using monoclonal antibodies. Cancer vaccines stimulate the immune system to recognize and attack PCa cells and sipuleucel-T (Provenge, a dendritic cell vaccine pulsed with a chimeric protein expressing GM-CSF and PAP as a tumor-associated antigen) is the most common vaccine used for PCa. Immune checkpoint inhibitors (ICI) block proteins in the cancer cells that help them evade the immune system and pembrolizumab (Keytruda) and nivolumab (Opdivo) are the most common ICIs used for PCa. It should be noted that immunotherapy is not a cure for PCa and may not be effective for all patients [39,40].
Antibody–drug conjugates, artificial bi-specific T cell-engaging antibodies, and immune checkpoint inhibitors are approaches to use monoclonal antibodies, while chimeric antigen receptor (CAR)-T cell therapy is a strategy for the adoptive transfer of T cells; both are part of immunotherapy [39] (preclinical and phase I clinical trial). Immune checkpoint inhibitors are monoclonal antibodies that inhibit immune checkpoint receptors to prevent the inactivation of T-cell function, and these antibodies target receptors, including cytotoxic T lymphocyte-associated protein 4 (CTLA-4), programmed death 1 (PD-1), and programmed death ligand 1 (PD-L1) [39].
3.1. Cytotoxic T Lymphocyte-Associated Protein 4
CTLA-4, a transmembrane protein expressed on T lymphocytes, competitively binds to ligands CD80 (B7.1) and CD86 (B7.2) on antigen-presenting cells (APCs), and prevents T-cells from killing other cells, including cancer cells [41]. Ipilimumab, a CTLA-4 inhibitor and the first Food and Drug Administration (FDA)-approved immune checkpoint inhibitor (ICI), blocks CTLA-4, switches off the inhibitor mechanism, and potentiates T-cell effects. CTLA-4 monotherapy significantly increases the ratio of regulator effector T lymphocytes in the tumor microenvironment [42]. However, monotherapy was not found to be associated with increased overall survival and failed to achieve significant benefits [43]. Ipilimumab showed promising results of a significant decline in prostate-specific antigen (PSA) levels when given in combination with granulocyte macrophage colony-stimulating factor (GM-CSF) and anti-tumor activity in phase I and phase I/II clinical trials, respectively, in metastatic castration-resistant PCa (mCRPC). A phase III clinical trial showed significantly increased overall survival (OS) in mCRPC but no significant change in OS was reported in asymptomatic or minimally symptomatic chemotherapy-naïve PCa with Ipilimumab monotherapy [38,39]. A study by Witt et al., using a mouse model of PCa, reported that signal transducer and activator of transcription 3 (STAT3) inhibition by GPB730 enhances the antitumor efficacy and activity of anti-CTLA-4, along with decreasing intratumoral Treg frequency, supporting the combination of STAT3 inhibition with anti-CTLA-4 therapy [44].
3.2. Programmed Death 1
PD-1 expressed on activated T cells, B-cells, and natural killer (NK) cells, when bound with PD-L1 (expressed on both normal and malignant cells), inhibits T cell activation (checkpoint for T lymphocyte inhibition) and converts naïve T cells into regulatory (Tregs) cells. Tumor cells expressing PD-L1 evade T cell antitumor response involving anergy or apoptosis of the effector T cells because the binding of PD-1 with PD-L1 results in apoptosis inhibition, T-lymphocyte tolerance, and increased tumor cell survival [45]. PD-L1 expression in PCa correlates with shorter clinical failure-free survival, proliferation (Ki-67), a lower Gleason score, and reduced prognostics for biochemical recurrence, but not with outcomes. Further, the expression profile in various studies also varied; these findings may be due to a lack of uniform criterion for determining PD-L1 positivity [38].
Nivolumab and pembrolizumab are ICIs for PD-1 inhibition, while atezolizumab, avelumab, and durvalumab are ICIs for PD-L1 inhibition [41]. Nivolumab or pembrolizumab have shown partial response in prostate tumors resistant to enzalutamide and with microsatellite instability (MSI). Patients with DNA mismatch repair mechanism (MRM) mutations may benefit from pembrolizumab [39]. Pembrolizumab, as a monotherapy or in combination (KEYNOTE-028 and KEYNOTE-199), induces durable objective responses, with minimal adverse events, and leads to anti-tumor activity with durable clinical responses and significant OS in patients with advanced PD-L1-positive mCRPC; therefore, it has been suggested as a form of immunotherapy. A high tumor mutational burden (TMB), PD-L1 expression, and high microsatellite instability (MSI) are the biomarkers for Pembrolizumab therapy [38,46,47,48]. Atezolizumab is a PD-L1 antibody, and its monotherapy has shown safety and clinical activity in terms of disease control and 12-month OS in mCRPC patients in a phase Ia, open-label, dose-escalation and dose-expansion study [49]. However, combination therapies are needed. The combination therapies of PD-L1 blockade with androgen inhibitors including The IMbassador 250 trial (atezolizumab with enzalutamide or enzalutamide alone) showed no overall significant improvement in OS [50] in KEYNOTE 365, COHORT C (enzalutamide in combination with pembrolizumab), showing a PSA response rate of 22% and ORR of 12% but not reaching the median duration of response [51], and in KEYNOTE 199 (enzalutamide plus pembrolizumab combination), showing a 12% ORR and 70–75% OS but not reaching median OS [52]. This suggests the need for more large-scale clinical trials with combination therapies.
3.3. Anti-Tumor Vaccines
The presence of tumor-associated antigens (TAA) in PCa makes it suitable for anti-tumor vaccines that elicit an adaptive immune response via antigen presentation [41]. Cell-based vaccines (dendritic cell or tumor cell), vector-based vaccines, DNA/mRNA-Based vaccines, and antigen- or peptide-based vaccines are common vaccines for PCa (Table 1). Vector-based vaccines include vectors derived from oncolytic viruses or bacterial pathogens inducing a specific immune response. DNA- and RNA-based vaccines consisting of plasmid DNA and mRNA, respectively, which elicit an immune response via both the major histocompatibility complex (MHC) class I and MHC class II pathways. The high potency, low manufacturing cost, developmental feasibility, and acceptable safety profile of DNA- and RNA-based vaccines make them an alternative to conventional vaccines [39,53].

Table 1.
Anti-tumor vaccines for PCa (prepared based on the description in [39]).
Antigen or peptide-based vaccines leverage short chains of amino acids (peptides) mimicking or representing specific parts of the antigens (epitopes) that best stimulate the immune system, and thus offer a targeted approach for inducing protective or therapeutic responses against cancer. The peptides presented to the body’s immune system lead to the production of antibodies and the activation of T cells. These activated T cells and antibodies then recognize and destroy the target cells carrying the same antigen. High purity, a minimal risk of side effects, potential autoimmunity, the inclusion of multiple antigens or epitopes in a single vaccine, and the ability to tailor vaccines to specific tumor types or individual patient characteristics are advantages of antigen or peptide-based vaccines [54,55]. Using these vaccines constitutes an attractive immunotherapeutic approach called personalized peptide vaccination (PPV) [56]. PPV plus chemotherapy, PPV plus glucocorticoids, HER2/neu peptides, and the AE37 vaccine are the most promising immunotherapeutic platforms involving PPV, as discussed by Adamaki et al. [39].
4. Combination Therapies
In mCRPC, combination therapies have shown promising results (Table 2), and various clinical trials are ongoing (Tables 2 and 3 of [57]) to evaluate their efficacy and safety.
Table 2.
Combination therapies for metastatic castration-resistant PCa (mCRPC).
5. Therapies for Neuroendocrine Prostate Cancer and Castration-Resistant Prostate Cancer
Table 3 lists other therapies for neuroendocrine prostate cancer (NEPC) and castration-resistant prostate cancer (CRPC, in addition to Table 1 and Table 2). NEPC, a rare tumor, is a highly aggressive variant of CRPC. When adenocarcinoma is treated with hormonal therapies , it can transform to NEPC, with a poor prognosis. Currently, NEPC lacks specific treatment options, and the available treatments are ineffective. Potential novel targets for NEPC are zeste homolog 2 (EZH2) inhibitors, lysine-specific demethylase 1 (LSD1) inhibitors, poly-ADP-ribose polymerase (PARP) inhibitors, aurora kinase A inhibitors, and delta-like ligand 3 (DLL3)-targeted therapies [74]. A recent study suggested Lewis Y antigen as a novel target for CAR-T cell therapy in patients with NEPC [75].
Table 3.
Immunotherapy of neuroendocrine and castration-resistant prostate cancer. PFS—progression-free survival; OS—overall survival; DLL3—delta-like ligand 3; mNEPC—metastatic neuroendocrine prostate cancer; CRPC—castration-resistant prostate cancer; NK cells—natural killer cells.
Other therapies for neuroendocrine prostate cancer targeting SSTR include somatostatin receptor; DLL-3, delta-like ligand 3; GPC3, glypican-3; GRPR, gastrin-releasing peptide receptor; B7-H3; CXCR2, CXC chemokine receptor 2; NCAM1, neural cell adhesion molecule 1; CEACAM5, carcinoembryonic antigen-related cell adhesion molecule 5; and HEPACAM2, HEPACAM Family Member 2. These have been summarized elsewhere [80]. Additionally, various clinical trials, including NCT05652686 (PT217), NCT04702737 (AMG757), NCT03480646 (CPI-1205), NCT02875548 (Tazemetostat), NCT05293496 (Vobramitamab and Duocarmazine), NCT04926181 (Cetrelimab, JNJ-63723283), NCT04592237 (Cetrelimab, JNJ-63723283), and NCT03866382 (Cabozantinib), are exploring the role of various agents in neuroendocrine prostate cancer [81]. For NEPCs, ICIs alone have achieved limited success in NEPC; they are being explored in combination with other therapies. For example, a combination therapy with ADT and immune checkpoint inhibitors has been investigated. Immunotherapeutic approaches for NEPC are still in their early stages, but promising preclinical and clinical data suggest that targeted therapies, including DLL3-targeted ADCs and bispecific antibodies, have the potential to treat this aggressive form of prostate cancer. Further research is needed to optimize immunotherapy strategies and identify the best combination therapies for NEPC patients [81,82].
6. Limitations of Immunotherapy
The immunologically cold behavior, strong immunosuppressive tumor microenvironment, and low tumor mutation burden observed in PCa limit the use of immunotherapy [83]. As discussed above, TAMs, fibroblasts within the tumor stroma, Trges, and MDSCs are present in TME, and molecules contributing to immunosuppression and the increased expression and activation of TGF-β, IL-10, IL-6, IL-8, prostaglandin E2, VEGF, PD-1, and PD-L1 are secreted by these cells. These factors further contribute to resistance to immunotherapy [17]. The poor T cell infiltration (reason for poor response to immunotherapy) in PCa [84,85] is due to a hypoxic environment [86] causing an acidic pH [87], the depletion of essential nutrients, the increased expression of adenosine and PD-L1 [88], and the increased expression of immunosuppressive transforming growth factor (TFG)-β. Primary PCas are characterized by PD-1-expressing CD8+ T cell infiltration, but this decreases in mCRPC, and thus a poor immune response is shown for ICIs targeting PD-1/PD-L1. However, mCRPC that expresses PD-L1 and develops resistance to the anti-androgen enzalutamide responds to ICIs inhibiting PD-1/PD-L1 [38,39]. T cells in the tumor cells (CD4+ FOXP3+ CD25+ T cells and CD8+ FOXP3+ CD25+ T cells) are immunosuppressive due to their increased secretion of inhibitory cytokines and inhibition of naïve T cell proliferation. The decreased expression of major histocompatibility complex (MHC) class I, low tumor mutation burden, low PD-1 expression, chronic activation of the interferon-1 (IFN-1), and PTEN loss adversely affect the tumor microenvironment and immunotherapy outcomes [38,41,83]. The exhaustion or dysfunction of infiltrating T cells, rendering them unable to effectively target and kill cancer cells, also contributes to their nonresponsiveness to ICIs [89]. The presence of myeloid-derived suppressor cells (MDSC) in the tumor microenvironment suppresses cytotoxic T-cell infiltration and restricts the anti-tumor effects of immune checkpoint inhibitors [43].
Tumor heterogeneity adds more complexity to the treatment of PCa because tumors expressing PD-L1 and not responding to immunotherapy are due to tumor heterogeneity. PD-L1 expression also varies with tumor progression and its expression is modulated by chemotherapy and radiation therapy [38]. Further, the presence of cancer-associated fibroblasts (CAFs) regulating tumor microenvironment modifies the response to immune checkpoint inhibitors and contributes to chemoresistance [90,91]. A tumor-immune microenvironment with a low number of tumor-infiltrating lymphocytes (TILs) and a high number of tumor-associated macrophages (TAMs) in PCa is unique and resistant to ICIs [92]. Racial difference has also been suggested as a factor for the different outcomes obtained in the treatment of PCa due to differential immune responses [40]. A lower TMB associated with fewer neoantigens hindering T-cell priming [93,94], an impaired T-cell priming and activation due to defects in tumor antigen presentation [94], the inhibitory effect of androgen deprivation therapy on the immune system [95], and the lack of effective biomarkers potentially reduce the effectiveness of ICI therapy. The use of a combination of ICIs is a current area of research aiming to overcome these limitations (Table 1 of [39] and Table 2 of [38]).
7. Future Prospectives
The inconsistent and ineffective results of CTLA-4 and PD-1/PD-L1 monotherapy and the ongoing trials of combination therapies suggest the need for further research and novel strategies for the treatment of mCRPA. As discussed above, a “cold” tumor type, limited ICI efficacy, the tumor microenvironment, the presence of CAFs, MDSC, and TAMs, and the lack of specific ICI therapy approval from the FDA are current challenges in the treatment of mCRPC. Combination immune checkpoint and adenosine axis blockade, Bispecific T cell engager and immune checkpoint blockade, Lu-PSMA-617 and immune checkpoint blockade, adoptive T-cell therapy, and immune checkpoint blockade [96,97,98] have been suggested as potential therapies to improve clinical outcome [38] (Figure 5). Research and clinical trials related to other strategies, such as fecal microbiota transplant (to improve gut microbiota), VEGF receptor-inhibitors (to decrease neoangiogenesis, thereby decreasing tumor growth), TPST-1120 (a peroxisome proliferator-activated receptor alpha (PPARα) antagonist that reduces fatty acid oxidation and shifts tumor cells to a more glycolytic metabolism), valemetostat-EZH1/2 dual inhibitor (inhibits the polycomb repressive complex 2 (PRC2), regulating gene expression through histone methylation), DF6002 (a monovalent IL-12 immunoglobulin Fc fusion protein that is immunostimulant, immunomodulatory, and antineoplastic), and M7824 (a fusion protein with two extracellular domains of TGF-RII and a PD-L1 monoclonal antibody (targets PD-1 and TGF-β)) are ongoing to improve clinical outcome in resistant PCa (Figure 5).
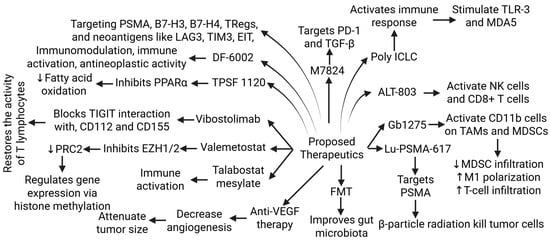
Figure 5.
Proposed newer therapies for metastatic chemoresistant prostate cancer (mCRPC) undergoing pre-clinical or clinical trials. Tumor-associated macrophages (TAMs), myeloid-derived stem cells (MDSCs), toll-like receptor (TLR)-3, peroxisome proliferator-activated receptor alpha (PPARα), polycomb repressive complex 2 (PRC2), T cell immunoreceptor with Ig and ITIM domains (TIGIT), enhancer of zeste homolog (EZH) 1, prostate-specific membrane antigen (PSMA), lymphocyte activation gene 3 (LAG3), T-regulatory cells (Tregs), T-cell immunoglobulin and mucin-domain containing-3 (TIM-3), epitope-based immunotherapy (EIT), Programmed Cell Death 1 (PD-1), transforming growth factor (TGF)-β, melanoma differentiation-associated protein (MDA) 5, natural killer (NK) cells, fecal microbial transplantation (FMT), and vascular endothelial growth factor (VEGF). ”↑”—increased expression; “↓”—decreased expression.
Poly ICLC, a synthetic double-stranded RNA complex activating innate immune responses, primarily through the stimulation of TLR3 and MDA5 receptors; ALT-803, a recombinant IL15 complex that enhances anti-tumor immune responses by directly activating NK cells and CD8+ T cells; GB1275,—a CD11b modulator that binds to and activates CD11b on TAMs and MDSCs in TME, which leads to reduced MDSC infiltration, the repolarization of M2 TAMs to an M1 phenotype, and increased tumor infiltration of cytotoxic T cells; vibostolimab, a monoclonal antibody that binds to the T-cell immunoreceptor and blocks its interaction with its ligands (targets and blocks the TIGIT receptor, an immune checkpoint protein); and Talabostat Mesylate, a small molecule inhibitor of dipeptidyl peptidases (acts through dipeptidyl peptidase inhibition and immune system activation) are under clinical trials to potentiate the treatment of mCRPC (as discussed in Table 3 of [38]) (Figure 5).
Adenosine has immunosuppressive effects, and the current interest is in blocking the adenosine-generating enzymes or antagonize adenosine receptors for immunomodulation [99]. Bispecific T-cell engagers are a type of antibody that helps the body’s immune system destroy cancer cells by simultaneously binding to tumor antigens and T cells, forming a bridge between tumor cells and cytotoxic T cells [96]. Recently, Grover et al. reported on the promising role of F77-expressing lentiviral CAR T cells with CD28 and CD137 (4-1BB) costimulatory signals for adoptive T cell therapy of PCa [98]. Another study by Aggarwal et al. reported on the safety of, and encouraging results obtained using, a single priming dose of 177Lu-PSMA-617 followed by a maintenance dose of pembrolizumab in mCRPC [97]. Another phase II study by Sandhu et al. is investigating the effects of radionuclide 177Lu-PSMA-617 therapy versus 177Lu-PSMA-617 in combination with ipilimumab and nivolumab [100]. Additionally, the B7 superfamily molecule B7-H3 (CD276 or PD-L3), which is highly expressed in PCa, has also been suggested as a novel target in PCa. B7-H3 plays an immunomodulatory role and a negative regulatory role in immune response. A lower CD3+ T cell density and higher Treg density are associated with a higher B7-H3 expression, which is also correlated with androgen receptors and signaling [57]. These findings suggest B7-H3 could be a potential target for mCRPC treatment.
Further, targeting neoantigens, antiangiogenics to attenuate angiogenesis, T-cell engagers (bispecific monoclonal antibodies), and adenosine pathway inhibitors may be beneficial in turning the cold immune environment into a responsive hot environment [17]. Targeting the EIT-mediated immunosuppressive environment involving B-7-H3, B7-H4, TIM3, and LAG3, in addition to PD-1/PD-L1 and CTLA-4, may be beneficial in designing novel therapeutic strategies for PCa [18] (Figure 5). Further, targeting Tregs may lead to promising therapeutic strategies. Such strategies include the depletion of Tregs, targeting immune checkpoint inhibitors related to Tregs, tumor necrosis factor receptor engagement, and targeting Tregs-derived cytokines [19]. Targeting Tregs is also important because Tregs may be a causative factor in the failure of various treatment strategies, such as the active vaccination of cancer patients, due to the stimulation of Tregs-specific antigens.
Using multi-omics (transcriptomics, proteomics, and metabolomics) in combination with bioinformatics and machine learning may be another avenue to identify novel therapeutic targets and the cause for PCa’s resistance to treatment, which could be due to a change in gene expression. Huang et al. [101] recently reported on the C1 and C2 subtypes of PCa, with C1 showing significantly increased expression of CGA, HSD17B12, BIRC5, CENPA, and MMP11, as well as being associated with shorter progression-free survival. The study also concluded that BIRC5, CENPA, and MMP11 are significant therapeutic targets in PCa.
Taken together, the results of this study may be beneficial in treating mCRPC and advanced PCa. Targeting specific targets and neoantigens seems to be a promising approach because the presence of neoantigens contributes to resistance to therapy.
8. Completed and Ongoing Clinical Trials for PCa Therapy
Table 4 summarizes the clinical trials for mCRPC. Clinical trials including NCT00861614 (ipilimumab + radiotherapy (RT) vs. placebo + RT), NCT01057810 (ipilimumab vs. placebo), NCT03834493 (pembrolizumab + enzalutamide vs. placebo + enzalutamide), NCT03834519 (pembrolizumab + olaparib vs. next-generation hormonal agent monotherapy), NCT03834506 (pembrolizumab + docetaxel vs. docetaxel), NCT03016312 (atezolizumab + enzalutamide vs. placebo + enzalutamide), and NCT04100018 (nivolumab + docetaxel vs. placebo + docetaxel) were conducted for mCRPC but the outcome showed unimproved survival [102]. Further, B7-H3 is an attractive target in B7-H3-positive Pca, and various clinical trials are testing agents targeting it. NCT01391143 (Phase 1, completed) and NCT02923180 (Phase 2, active) tested Enoblituzumab and found a significant activation of immune response in their preliminary results. NCT03729596 (Phase 1, completed) and NCT05555117 (Phase 2, active) are being used to investigate humanized mAb vobramitamab loaded with duocarmazine targeting B7-H3, and the preliminary results showed a tolerable safety profile with a decrease in tumor lesion size in mCRPC patients. NCT04145622 is testing anti-B7-H3 ifinatamab deruxtecan (ADC DS-7300a), and NCT05914116 is investigating DB-1311, a humanized anti-B7-H3 mAb linked to a cleavable DNA topoisomerase I inhibitor for their anti-tumor activity [103]. Recently, Li et al. tested a second-generation CAR specifically targeting B7-H3 and CD28 in PCa that was positive for B7-H3 and found that they efficiently controlled the growth of the tumor and increased the release of TNF-α and IFN-γ in vitro [104].
Table 4.
Clinical trials testing the safety and efficacy of various immunotherapeutic agents for prostate cancer.
9. Conclusions
The presence of an immunosuppressive tumor microenvironment, a tumor-immune microenvironment, a low tumor mutation burden, higher expression of PD-1 and low expression of PD-L1, the presence of MDSCs, tumor heterogeneity, and racial differences make prostate tumors resistant to ICIs. Although monotherapy ICI trials in mCRPC have not been very promising, combination therapies have shown some promising results. Still, there is an unmet need for a better therapeutic approach. To design novel personalized therapeutics for mCRPC, understanding the molecular mechanisms and the mechanisms contributing to resistance to ICIs in mCRPC is important. Utilizing other novel targets in the signaling pathway of PCa, along with ICIs, may be more synergistically effective. Theranostics (combining therapy with testing) is an emerging field for PCa treatment, with fewer tests. Further, PSMA therapy, a promising treatment for advanced PCa, is a type of radionuclide therapy that uses a radioactive isotope (lutetium-177) attached to a molecule that specifically binds to the prostate-specific membrane antigen (PSMA) to deliver radioactive particles directly to cancer cells. Future research activities to investigate reliable biomarkers of response to immunotherapy and the application of theranostics technology in clinical practice are warranted for an improved clinical outcome.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| AR | Androgen receptor |
| CDK12 | Cyclin-dependent kinase 12 |
| CAR-T cells | Chimeric antigen receptor -T cell |
| CAFs | Cancer-associated fibroblasts |
| HDAC | Histone deacetylases |
| HMTs | Histone methyltransferase |
| HDMs | Histone demethylase |
| ICIs | Immune checkpoint inhibitors |
| mTOR | Mammalian target of rapamycin |
| mCRPC | Metastatic castration-resistant prostate cancer |
| MHC | Major histocompatibility complex |
| MDSCs | Myeloid-derived suppressor cells |
| PI3K | Phosphoinositide 3-kinase |
| PTEN | Phosphatase and tensin homolog |
| PD-1 | Programmed cell death protein 1 |
| PD-L1 | Programmed cell death ligand 1 |
| PSA | Prostate specific antigen |
| STAT3 | Signal transducer and activator of transcription 3 |
| TMB | Tumor mutational burden |
| TAA | Tumor-associated antigens |
| TILs | Tumor-infiltrating lymphocytes |
| TAMs | Tumor-associated macrophages |
References
- Lowrance, W.; Dreicer, R.; Jarrard, D.F.; Scarpato, K.R.; Kim, S.K.; Kirkby, E.; Buckley, D.I.; Griffin, J.C.; Cookson, M.S. Updates to Advanced Prostate Cancer: AUA/SUO Guideline (2023). J. Urol. 2023, 209, 1082–1090. [Google Scholar] [CrossRef]
- Gandaglia, G.; Leni, R.; Bray, F.; Fleshner, N.; Freedland, S.J.; Kibel, A.; Stattin, P.; Van Poppel, H.; La Vecchia, C. Epidemiology and Prevention of Prostate Cancer. Eur. Urol. Oncol. 2021, 4, 877–892. [Google Scholar] [CrossRef]
- Rawla, P. Epidemiology of Prostate Cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef]
- Gann, P.H. Risk factors for prostate cancer. Rev. Urol. 2002, 4 (Suppl. 5), S3–S10. [Google Scholar]
- Cicione, A.; Brassetti, A.; Lombardo, R.; Franco, A.; Turchi, B.; D’Annunzio, S.; Nacchia, A.; Tubaro, A.; Simone, G.; De Nunzio, C. Metabolic Syndrome and Physical Inactivity May Be Shared Etiological Agents of Prostate Cancer and Coronary Heart Diseases. Cancers 2022, 14, 936. [Google Scholar] [CrossRef]
- Dunn, M.W.; Kazer, M.W. Prostate cancer overview. Semin. Oncol. Nurs. 2011, 27, 241–250. [Google Scholar] [CrossRef]
- Litwin, M.S.; Tan, H.-J. The diagnosis and treatment of prostate cancer: A review. Jama 2017, 317, 2532–2542. [Google Scholar] [CrossRef]
- Board, P.A.T.E. Prostate cancer treatment (PDQ®). In PDQ Cancer Information Summaries [Internet]; National Cancer Institute (US): Bethesda, MD, USA, 2024. [Google Scholar]
- Wang, F.; Li, Z.; Feng, X.; Yang, D.; Lin, M. Advances in PSMA-targeted therapy for prostate cancer. Prostate Cancer Prostatic Dis. 2022, 25, 11–26. [Google Scholar] [CrossRef]
- Cicione, A.; Nacchia, A.; Guercio, A.; Gravina, C.; Franco, A.; Grimaldi, M.C.; Tema, G.; Lombardo, R.; Tubaro, A.; De Nunzio, C. Cardiovascular adverse events-related to GnRH agonists and GnRH antagonists: Analysis of real-life data from Eudra-Vigilance and Food and Drug Administration databases entries. Prostate Cancer Prostatic Dis. 2023, 26, 765–771. [Google Scholar] [CrossRef]
- Petrylak, D.P.; Ratta, R.; Matsubara, N.; Korbenfeld, E.; Gafanov, R.; Mourey, L.; Todenhofer, T.; Gurney, H.; Kramer, G.; Bergman, A.M.; et al. Pembrolizumab Plus Docetaxel Versus Docetaxel for Previously Treated Metastatic Castration-Resistant Prostate Cancer: The Randomized, Double-Blind, Phase III KEYNOTE-921 Trial. J. Clin. Oncol. 2025, 43, JCO2401283. [Google Scholar] [CrossRef]
- Onishi, K.; Nakai, Y.; Tachibana, A.; Nishimura, N.; Maesaka, F.; Tomizawa, M.; Morizawa, Y.; Hori, S.; Gotoh, D.; Miyake, M.; et al. Testosterone Recovery and Quality of Life of Japanese Patients After Short-Term Neoadjuvant Androgen Deprivation Therapy With Low-Dose-Rate Brachytherapy for Prostate Cancer. Int. J. Urol. 2025. [Google Scholar] [CrossRef]
- Strasner, A.; Karin, M. Immune Infiltration and Prostate Cancer. Front. Oncol. 2015, 5, 128. [Google Scholar] [CrossRef]
- Palano, M.T.; Gallazzi, M.; Cucchiara, M.; Deho, F.; Capogrosso, P.; Bruno, A.; Mortara, L. The tumor innate immune microenvironment in prostate cancer: An overview of soluble factors and cellular effectors. Explor. Target. Anti-Tumor Ther. 2022, 3, 694–718. [Google Scholar] [CrossRef]
- Harder, J.L.; Linden, P.; Jahn, L.; Aslan, M.; Schmucker, V. [Cross-regional telemedicine services as a supplement to rural primary care: A mixed-methods analysis]. Z. Evidenz Fortbild. Qual. Gesundheitswesen 2022, 169, 67–74. [Google Scholar] [CrossRef]
- Kaminski, A.; Hahne, J.C.; Haddouti, E.M.; Florin, A.; Wellmann, A.; Wernert, N. Tumour-stroma interactions between metastatic prostate cancer cells and fibroblasts. Int. J. Mol. Med. 2006, 18, 941–950. [Google Scholar] [CrossRef]
- Stultz, J.; Fong, L. How to turn up the heat on the cold immune microenvironment of metastatic prostate cancer. Prostate Cancer Prostatic Dis. 2021, 24, 697–717. [Google Scholar] [CrossRef]
- Lin, D.; Wang, X.; Choi, S.Y.C.; Ci, X.; Dong, X.; Wang, Y. Immune phenotypes of prostate cancer cells: Evidence of epithelial immune cell-like transition? Asian J. Urol. 2016, 3, 195–202. [Google Scholar] [CrossRef]
- Karpisheh, V.; Mousavi, S.M.; Naghavi Sheykholeslami, P.; Fathi, M.; Mohammadpour Saray, M.; Aghebati-Maleki, L.; Jafari, R.; Majidi Zolbanin, N.; Jadidi-Niaragh, F. The role of regulatory T cells in the pathogenesis and treatment of prostate cancer. Life Sci. 2021, 284, 119132. [Google Scholar] [CrossRef]
- Testa, U.; Castelli, G.; Pelosi, E. Cellular and Molecular Mechanisms Underlying Prostate Cancer Development: Therapeutic Implications. Medicines 2019, 6, 82. [Google Scholar] [CrossRef]
- Maekawa, S.; Takata, R.; Obara, W. Molecular Mechanisms of Prostate Cancer Development in the Precision Medicine Era: A Comprehensive Review. Cancers 2024, 16, 523. [Google Scholar] [CrossRef]
- Fontana, F.; Anselmi, M.; Limonta, P. Molecular mechanisms and genetic alterations in prostate cancer: From diagnosis to targeted therapy. Cancer Lett. 2022, 534, 215619. [Google Scholar] [CrossRef]
- Shtivelman, E.; Beer, T.M.; Evans, C.P. Molecular pathways and targets in prostate cancer. Oncotarget 2014, 5, 7217–7259. [Google Scholar] [CrossRef]
- Perdomo, H.A.G.; Zapata-Copete, J.A.; Sanchez, A. Molecular alterations associated with prostate cancer. Cent. Eur. J. Urol. 2018, 71, 168–176. [Google Scholar] [CrossRef]
- Paltoglou, S.; Das, R.; Townley, S.L.; Hickey, T.E.; Tarulli, G.A.; Coutinho, I.; Fernandes, R.; Hanson, A.R.; Denis, I.; Carroll, J.S.; et al. Novel Androgen Receptor Coregulator GRHL2 Exerts Both Oncogenic and Antimetastatic Functions in Prostate Cancer. Cancer Res. 2017, 77, 3417–3430. [Google Scholar] [CrossRef]
- Ying, Y.; Wu, Y.; Zhang, F.; Tang, Y.; Yi, J.; Ma, X.; Li, J.; Chen, D.; Wang, X.; Liu, X.; et al. Co-transcriptional R-loops-mediated epigenetic regulation drives growth retardation and docetaxel chemosensitivity enhancement in advanced prostate cancer. Mol. Cancer 2024, 23, 79. [Google Scholar] [CrossRef]
- Cai, C.; Chen, S.; Ng, P.; Bubley, G.J.; Nelson, P.S.; Mostaghel, E.A.; Marck, B.; Matsumoto, A.M.; Simon, N.I.; Wang, H.; et al. Intratumoral de novo steroid synthesis activates androgen receptor in castration-resistant prostate cancer and is upregulated by treatment with CYP17A1 inhibitors. Cancer Res. 2011, 71, 6503–6513. [Google Scholar] [CrossRef]
- Monge, A.; Jagla, M.; Lapouge, G.; Sasorith, S.; Cruchant, M.; Wurtz, J.M.; Jacqmin, D.; Bergerat, J.P.; Ceraline, J. Unfaithfulness and promiscuity of a mutant androgen receptor in a hormone-refractory prostate cancer. Cell. Mol. Life Sci. 2006, 63, 487–497. [Google Scholar] [CrossRef]
- Tan, M.H.; Li, J.; Xu, H.E.; Melcher, K.; Yong, E.L. Androgen receptor: Structure, role in prostate cancer and drug discovery. Acta Pharmacol. Sin. 2015, 36, 3–23. [Google Scholar] [CrossRef]
- Shiota, M.; Akamatsu, S.; Tsukahara, S.; Nagakawa, S.; Matsumoto, T.; Eto, M. Androgen receptor mutations for precision medicine in prostate cancer. Endocr. Relat. Cancer 2022, 29, R143–R155. [Google Scholar] [CrossRef]
- Schulz, W.A.; Burchardt, M.; Cronauer, M.V. Molecular biology of prostate cancer. Mol. Hum. Reprod. 2003, 9, 437–448. [Google Scholar] [CrossRef][Green Version]
- Fordyce, C.A.; Heaphy, C.M.; Joste, N.E.; Smith, A.Y.; Hunt, W.C.; Griffith, J.K. Association between cancer-free survival and telomere DNA content in prostate tumors. J. Urol. 2005, 173, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Heaphy, C.M.; Yoon, G.S.; Peskoe, S.B.; Joshu, C.E.; Lee, T.K.; Giovannucci, E.; Mucci, L.A.; Kenfield, S.A.; Stampfer, M.J.; Hicks, J.L.; et al. Prostate cancer cell telomere length variability and stromal cell telomere length as prognostic markers for metastasis and death. Cancer Discov. 2013, 3, 1130–1141. [Google Scholar] [CrossRef] [PubMed]
- Jeronimo, C.; Bastian, P.J.; Bjartell, A.; Carbone, G.M.; Catto, J.W.; Clark, S.J.; Henrique, R.; Nelson, W.G.; Shariat, S.F. Epigenetics in prostate cancer: Biologic and clinical relevance. Eur. Urol. 2011, 60, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Ragavi, R.; Muthukumaran, P.; Nandagopal, S.; Ahirwar, D.K.; Tomo, S.; Misra, S.; Guerriero, G.; Shukla, K.K. Epigenetics regulation of prostate cancer: Biomarker and therapeutic potential. Urol. Oncol. 2023, 41, 340–353. [Google Scholar] [CrossRef]
- Bilusic, M.; Madan, R.A.; Gulley, J.L. Immunotherapy of Prostate Cancer: Facts and Hopes. Clin. Cancer Res. 2017, 23, 6764–6770. [Google Scholar] [CrossRef]
- Markowski, M.C.; Shenderov, E.; Eisenberger, M.A.; Kachhap, S.; Pardoll, D.M.; Denmeade, S.R.; Antonarakis, E.S. Extreme responses to immune checkpoint blockade following bipolar androgen therapy and enzalutamide in patients with metastatic castration resistant prostate cancer. Prostate 2020, 80, 407–411. [Google Scholar] [CrossRef]
- Venkatachalam, S.; McFarland, T.R.; Agarwal, N.; Swami, U. Immune Checkpoint Inhibitors in Prostate Cancer. Cancers 2021, 13, 2187. [Google Scholar] [CrossRef]
- Adamaki, M.; Zoumpourlis, V. Immunotherapy as a Precision Medicine Tool for the Treatment of Prostate Cancer. Cancers 2021, 13, 173. [Google Scholar] [CrossRef]
- Wang, I.; Song, L.; Wang, B.Y.; Rezazadeh Kalebasty, A.; Uchio, E.; Zi, X. Prostate cancer immunotherapy: A review of recent advancements with novel treatment methods and efficacy. Am. J. Clin. Exp. Urol. 2022, 10, 210–233. [Google Scholar]
- Patel, D.; McKay, R.; Parsons, J.K. Immunotherapy for Localized Prostate Cancer: The Next Frontier? Urol. Clin. N. Am. 2020, 47, 443–456. [Google Scholar] [CrossRef]
- Fellner, C. Ipilimumab (yervoy) prolongs survival in advanced melanoma: Serious side effects and a hefty price tag may limit its use. Pharm. Ther. 2012, 37, 503–530. [Google Scholar]
- Jeong, S.H.; Kwak, C. Immunotherapy for prostate cancer: Requirements for a successful regime transfer. Investig. Clin. Urol. 2022, 63, 3–13. [Google Scholar] [CrossRef]
- Witt, K.; Evans-Axelsson, S.; Lundqvist, A.; Johansson, M.; Bjartell, A.; Hellsten, R. Inhibition of STAT3 augments antitumor efficacy of anti-CTLA-4 treatment against prostate cancer. Cancer Immunol. Immunother. 2021, 70, 3155–3166. [Google Scholar] [CrossRef]
- Francisco, L.M.; Salinas, V.H.; Brown, K.E.; Vanguri, V.K.; Freeman, G.J.; Kuchroo, V.K.; Sharpe, A.H. PD-L1 regulates the development, maintenance, and function of induced regulatory T cells. J. Exp. Med. 2009, 206, 3015–3029. [Google Scholar] [CrossRef]
- Hansen, A.R.; Massard, C.; Ott, P.A.; Haas, N.B.; Lopez, J.S.; Ejadi, S.; Wallmark, J.M.; Keam, B.; Delord, J.P.; Aggarwal, R.; et al. Pembrolizumab for advanced prostate adenocarcinoma: Findings of the KEYNOTE-028 study. Ann. Oncol. 2018, 29, 1807–1813. [Google Scholar] [CrossRef]
- Antonarakis, E.S.; Piulats, J.M.; Gross-Goupil, M.; Goh, J.; Ojamaa, K.; Hoimes, C.J.; Vaishampayan, U.; Berger, R.; Sezer, A.; Alanko, T.; et al. Pembrolizumab for Treatment-Refractory Metastatic Castration-Resistant Prostate Cancer: Multicohort, Open-Label Phase II KEYNOTE-199 Study. J. Clin. Oncol. 2020, 38, 395–405. [Google Scholar] [CrossRef]
- Marcus, L.; Lemery, S.J.; Keegan, P.; Pazdur, R. FDA Approval Summary: Pembrolizumab for the Treatment of Microsatellite Instability-High Solid Tumors. Clin. Cancer Res. 2019, 25, 3753–3758. [Google Scholar] [CrossRef]
- Petrylak, D.P.; Loriot, Y.; Shaffer, D.R.; Braiteh, F.; Powderly, J.; Harshman, L.C.; Conkling, P.; Delord, J.P.; Gordon, M.; Kim, J.W.; et al. Safety and Clinical Activity of Atezolizumab in Patients with Metastatic Castration-Resistant Prostate Cancer: A Phase I Study. Clin. Cancer Res. 2021, 27, 3360–3369. [Google Scholar] [CrossRef]
- Sweeney, C.J.; Gillessen, S.; Rathkopf, D.; Matsubara, N.; Drake, C.; Fizazi, K.; Piulats, J.M.; Wysocki, P.J.; Buchschacher, G.L., Jr.; Doss, J. IMbassador250: A phase III trial comparing atezolizumab with enzalutamide vs enzalutamide alone in patients with metastatic castration-resistant prostate cancer (mCRPC). Cancer Res. 2020, 80, TPS5090. [Google Scholar] [CrossRef]
- Yu*, E.Y.; Fong, P.; Piulats, J.M.; Appleman, L.; Conter, H.; Feyerabend, S.; Shore, N.; Gravis, G.; Laguerre, B.; Gurney, H. PD16-12 pembrolizumab plus enzalutamide in abiraterone-pretreated patients with metastatic castration-resistant prostate cancer: Updated results from KEYNOTE-365 cohort C. J. Urol. 2020, 203, e368. [Google Scholar] [CrossRef]
- Hoimes, C.J.; Graff, J.N.; Tagawa, S.T.; Hwang, C.; Kilari, D.; Ten Tije, A.; Omlin, A.; McDermott, R.S.; Vaishampayan, U.N.; Elliott, T. KEYNOTE-199 cohorts (C) 4 and 5: Phase II study of pembrolizumab (pembro) plus enzalutamide (enza) for enza-resistant metastatic castration-resistant prostate cancer (mCRPC). J. Clin. Oncol. 2020, 38, 5543. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef]
- Buonaguro, L.; Tagliamonte, M. Peptide-based vaccine for cancer therapies. Front. Immunol. 2023, 14, 1210044. [Google Scholar] [CrossRef]
- Stephens, A.J.; Burgess-Brown, N.A.; Jiang, S. Beyond Just Peptide Antigens: The Complex World of Peptide-Based Cancer Vaccines. Front. Immunol. 2021, 12, 696791. [Google Scholar] [CrossRef]
- Sasada, T.; Noguchi, M.; Yamada, A.; Itoh, K. Personalized peptide vaccination: A novel immunotherapeutic approach for advanced cancer. Hum. Vaccines Immunother. 2012, 8, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Lanka, S.M.; Zorko, N.A.; Antonarakis, E.S.; Barata, P.C. Metastatic Castration-Resistant Prostate Cancer, Immune Checkpoint Inhibitors, and Beyond. Curr. Oncol. 2023, 30, 4246–4256. [Google Scholar] [CrossRef] [PubMed]
- Karzai, F.; VanderWeele, D.; Madan, R.A.; Owens, H.; Cordes, L.M.; Hankin, A.; Couvillon, A.; Nichols, E.; Bilusic, M.; Beshiri, M.L.; et al. Activity of durvalumab plus olaparib in metastatic castration-resistant prostate cancer in men with and without DNA damage repair mutations. J. Immunother. Cancer 2018, 6, 141. [Google Scholar] [CrossRef]
- Yu, E.Y.; Piulats, J.M.; Gravis, G.; Laguerre, B.; Arranz Arija, J.A.; Oudard, S.; Fong, P.C.; Kolinsky, M.P.; Augustin, M.; Feyerabend, S. KEYNOTE-365 cohort A updated results: Pembrolizumab (pembro) plus olaparib in docetaxel-pretreated patients (pts) with metastatic castration-resistant prostate cancer (mCRPC). J. Clin. Oncol. 2020, 38, 100. [Google Scholar] [CrossRef]
- Kolinsky, M.P.; Gravis, G.; Mourey, L.; Piulats, J.M.; Sridhar, S.S.; Romano, E.; Berry, W.R.; Gurney, H.; Retz, M.; Appleman, L.J. KEYNOTE-365 cohort B updated results: Pembrolizumab (pembro) plus docetaxel and prednisone in abiraterone (abi) or enzalutamide (enza)-pretreated patients (pts) with metastatic castrate-resistant prostate cancer (mCRPC). J. Clin. Oncol. 2020, 38, 103. [Google Scholar] [CrossRef]
- Morris, M.J.; Fong, L.; Petrylak, D.P.; Sartor, A.O.; Higano, C.S.; Pagliaro, L.C.; Alva, A.S.; Appleman, L.J.; Tan, W.; Vaishampayan, U.N. Safety and clinical activity of atezolizumab (atezo)+ radium-223 dichloride (r-223) in 2L metastatic castration-resistant prostate cancer (mCRPC): Results from a phase Ib clinical trial. J. Clin. Oncol. 2020, 38, 5565. [Google Scholar] [CrossRef]
- Slovin, S.F.; Higano, C.S.; Hamid, O.; Tejwani, S.; Harzstark, A.; Alumkal, J.J.; Scher, H.I.; Chin, K.; Gagnier, P.; McHenry, M.B.; et al. Ipilimumab alone or in combination with radiotherapy in metastatic castration-resistant prostate cancer: Results from an open-label, multicenter phase I/II study. Ann. Oncol. 2013, 24, 1813–1821. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.D.; Drake, C.G.; Scher, H.I.; Fizazi, K.; Bossi, A.; van den Eertwegh, A.J.; Krainer, M.; Houede, N.; Santos, R.; Mahammedi, H.; et al. Ipilimumab versus placebo after radiotherapy in patients with metastatic castration-resistant prostate cancer that had progressed after docetaxel chemotherapy (CA184-043): A multicentre, randomised, double-blind, phase 3 trial. Lancet Oncol. 2014, 15, 700–712. [Google Scholar] [CrossRef] [PubMed]
- Fizazi, K.; Drake, C.G.; Beer, T.M.; Kwon, E.D.; Scher, H.I.; Gerritsen, W.R.; Bossi, A.; den Eertwegh, A.; Krainer, M.; Houede, N.; et al. Final Analysis of the Ipilimumab Versus Placebo Following Radiotherapy Phase III Trial in Postdocetaxel Metastatic Castration-resistant Prostate Cancer Identifies an Excess of Long-term Survivors. Eur. Urol. 2020, 78, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.E.; Hurley, P.J.; Tran, P.T.; Rowe, S.P.; Benzon, B.; Neal, T.O.; Chapman, C.; Harb, R.; Milman, Y.; Trock, B.J.; et al. A pilot trial of pembrolizumab plus prostatic cryotherapy for men with newly diagnosed oligometastatic hormone-sensitive prostate cancer. Prostate Cancer Prostatic Dis. 2020, 23, 184–193. [Google Scholar] [CrossRef]
- Rosser, C.J.; Hirasawa, Y.; Acoba, J.D.; Tamura, D.J.; Pal, S.K.; Huang, J.; Scholz, M.C.; Dorff, T.B. Phase Ib study assessing different sequencing regimens of atezolizumab (anti-PD-L1) and sipuleucel-T (SipT) in patients who have asymptomatic or minimally symptomatic metastatic castrate resistant prostate cancer. J. Clin. Oncol. 2020, 38, e17564. [Google Scholar] [CrossRef]
- Tuthill, M.; Cappuccini, F.; Carter, L.; Pollock, E.; Poulton, I.; Verrill, C.; Evans, T.; Gillessen, S.; Attard, G.; Protheroe, A. 682P Results from ADVANCE: A phase I/II open-label non-randomised safety and efficacy study of the viral vectored ChAdOx1-MVA 5T4 (VTP-800) vaccine in combination with PD-1 checkpoint blockade in metastatic prostate cancer. Ann. Oncol. 2020, 31, S543. [Google Scholar] [CrossRef]
- Madan, R.A.; Mohebtash, M.; Arlen, P.M.; Vergati, M.; Rauckhorst, M.; Steinberg, S.M.; Tsang, K.Y.; Poole, D.J.; Parnes, H.L.; Wright, J.J.; et al. Ipilimumab and a poxviral vaccine targeting prostate-specific antigen in metastatic castration-resistant prostate cancer: A phase 1 dose-escalation trial. Lancet Oncol. 2012, 13, 501–508. [Google Scholar] [CrossRef]
- Boudadi, K.; Suzman, D.L.; Anagnostou, V.; Fu, W.; Luber, B.; Wang, H.; Niknafs, N.; White, J.R.; Silberstein, J.L.; Sullivan, R.; et al. Ipilimumab plus nivolumab and DNA-repair defects in AR-V7-expressing metastatic prostate cancer. Oncotarget 2018, 9, 28561–28571. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Pachynski, R.K.; Narayan, V.; Flechon, A.; Gravis, G.; Galsky, M.D.; Mahammedi, H.; Patnaik, A.; Subudhi, S.K.; Ciprotti, M.; et al. Nivolumab Plus Ipilimumab for Metastatic Castration-Resistant Prostate Cancer: Preliminary Analysis of Patients in the CheckMate 650 Trial. Cancer Cell 2020, 38, 489–499.e3. [Google Scholar] [CrossRef]
- Hotte, S.; Winquist, E.; Chi, K.; Ellard, S.; Sridhar, S.; Emmenegger, U.; Salim, M.; Iqbal, N.; Canil, C.; Kollmannsberger, C. CCTG IND 232: A phase II study of durvalumab with or without tremelimumab in patients with metastatic castration resistant prostate cancer (mCRPC). Ann. Oncol. 2019, 30, v885. [Google Scholar] [CrossRef]
- Subudhi, S.K.; Siddiqui, B.A.; Aparicio, A.M.; Yadav, S.S.; Basu, S.; Chen, H.; Jindal, S.; Tidwell, R.S.S.; Varma, A.; Logothetis, C.J.; et al. Combined CTLA-4 and PD-L1 blockade in patients with chemotherapy-naive metastatic castration-resistant prostate cancer is associated with increased myeloid and neutrophil immune subsets in the bone microenvironment. J. Immunother. Cancer 2021, 9, e002919. [Google Scholar] [CrossRef]
- Agarwal, N.; Loriot, Y.; McGregor, B.A.; Dreicer, R.; Dorff, T.B.; Maughan, B.L.; Kelly, W.K.; Pagliaro, L.C.; Srinivas, S.; Squillante, C.M. Cabozantinib in combination with atezolizumab in patients with metastatic castration-resistant prostate cancer: Results of cohort 6 of the COSMIC-021 study. J. Clin. Oncol. 2020, 38, 5564. [Google Scholar] [CrossRef]
- Fei, X.; Xue, J.W.; Wu, J.Z.; Yang, C.Y.; Wang, K.J.; Ma, Q. Promising therapy for neuroendocrine prostate cancer: Current status and future directions. Ther. Adv. Med. Oncol. 2024, 16, 17588359241269676. [Google Scholar] [CrossRef] [PubMed]
- Kulakowska, W.; Taylor, R.; Ranasinghe, W.; Darcy, P.K.; Trapani, J.; Risbridger, G. Lewis Y antigen as a novel target for CAR (chimeric antigen receptor) T-cell therapy in patients with neuroendocrine prostate cancer. J. Clin. Oncol. 2025, 43, 217. [Google Scholar] [CrossRef]
- Wee, C.E.; Costello, B.A.; Orme, J.J.; Quevedo, J.F.; Pagliaro, L.C. Chemotherapy with atezolizumab for small cell or neuroendocrine carcinoma of the prostate: A single institution experience. Prostate 2021, 81, 938–943. [Google Scholar] [CrossRef]
- Salhab, M.; Donahue, M.; Walsh, W. Pembrolizumab for platinum refractory small cell carcinoma of the prostate: Case report. Hematol. Med. Oncol. 2018, 3, 1–3. [Google Scholar]
- Puca, L.; Gavyert, K.; Sailer, V.; Conteduca, V.; Dardenne, E.; Sigouros, M.; Isse, K.; Kearney, M.; Vosoughi, A.; Fernandez, L.; et al. Delta-like protein 3 expression and therapeutic targeting in neuroendocrine prostate cancer. Sci. Transl. Med. 2019, 11, eaav0891. [Google Scholar] [CrossRef]
- Haque Chowdhury, H.; Hawlina, S.; Gabrijel, M.; Trkov Bobnar, S.; Kreft, M.; Lenart, G.; Cukjati, M.; Kopitar, A.N.; Kejzar, N.; Ihan, A.; et al. Survival of castration-resistant prostate cancer patients treated with dendritic-tumor cell hybridomas is negatively correlated with changes in peripheral blood CD56(bright) CD16(-) natural killer cells. Clin. Transl. Med. 2021, 11, e505. [Google Scholar] [CrossRef] [PubMed]
- Bilgin, G.B.; Lucien-Matteoni, F.; Chaudhuri, A.A.; Orme, J.J.; Childs, D.S.; Rincon, M.M.; Li, G.G.; Chauhan, P.S.; Lee, S.; Gupta, S. Current and future directions in theranostics for neuroendocrine prostate cancer. Cancer Treat. Rev. 2025, 136, 102941. [Google Scholar] [CrossRef]
- Bernal, A.; Bechler, A.J.; Mohan, K.; Rizzino, A.; Mathew, G. The Current Therapeutic Landscape for Metastatic Prostate Cancer. Pharmaceuticals 2024, 17, 351. [Google Scholar] [CrossRef]
- Chen, J.; Ma, N.; Chen, B.; Huang, Y.; Li, J.; Li, J.; Chen, Z.; Wang, P.; Ran, B.; Yang, J. Synergistic effects of immunotherapy and adjunctive therapies in prostate cancer management. Crit. Rev. Oncol./Hematol. 2024, 207, 104604. [Google Scholar] [CrossRef] [PubMed]
- Berger, M.F.; Lawrence, M.S.; Demichelis, F.; Drier, Y.; Cibulskis, K.; Sivachenko, A.Y.; Sboner, A.; Esgueva, R.; Pflueger, D.; Sougnez, C.; et al. The genomic complexity of primary human prostate cancer. Nature 2011, 470, 214–220. [Google Scholar] [CrossRef]
- Hua, L.; Xia, H.; Zheng, W. The Association between Immune Infiltration and Clinical Phenotypes and Prognosis of Prostate Cancer. Iran. J. Biotechnol. 2020, 18, e2538. [Google Scholar] [CrossRef]
- Niu, W.; Zhang, T.; Ma, L. Correlation analysis between immune-related genes and cell infiltration revealed prostate cancer immunotherapy biomarkers linked to T cells gamma delta. Sci. Rep. 2023, 13, 2459. [Google Scholar] [CrossRef]
- Jayaprakash, P.; Ai, M.; Liu, A.; Budhani, P.; Bartkowiak, T.; Sheng, J.; Ager, C.; Nicholas, C.; Jaiswal, A.R.; Sun, Y.; et al. Targeted hypoxia reduction restores T cell infiltration and sensitizes prostate cancer to immunotherapy. J. Clin. Investig. 2018, 128, 5137–5149. [Google Scholar] [CrossRef] [PubMed]
- El-Kenawi, A.; Gatenbee, C.; Robertson-Tessi, M.; Bravo, R.; Dhillon, J.; Balagurunathan, Y.; Berglund, A.; Vishvakarma, N.; Ibrahim-Hashim, A.; Choi, J.; et al. Acidity promotes tumour progression by altering macrophage phenotype in prostate cancer. Br. J. Cancer 2019, 121, 556–566. [Google Scholar] [CrossRef]
- Bishop, J.L.; Sio, A.; Angeles, A.; Roberts, M.E.; Azad, A.A.; Chi, K.N.; Zoubeidi, A. PD-L1 is highly expressed in Enzalutamide resistant prostate cancer. Oncotarget 2015, 6, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Ni, Y.; Liang, X.; Lin, Y.; An, B.; He, X.; Zhao, X. Mechanisms of tumor resistance to immune checkpoint blockade and combination strategies to overcome resistance. Front. Immunol. 2022, 13, 915094. [Google Scholar] [CrossRef]
- Liu, T.; Han, C.; Wang, S.; Fang, P.; Ma, Z.; Xu, L.; Yin, R. Cancer-associated fibroblasts: An emerging target of anti-cancer immunotherapy. J. Hematol. Oncol. 2019, 12, 86. [Google Scholar] [CrossRef]
- Cheteh, E.H.; Augsten, M.; Rundqvist, H.; Bianchi, J.; Sarne, V.; Egevad, L.; Bykov, V.J.; Ostman, A.; Wiman, K.G. Human cancer-associated fibroblasts enhance glutathione levels and antagonize drug-induced prostate cancer cell death. Cell Death Dis. 2017, 8, e2848. [Google Scholar] [CrossRef]
- Wang, T.Y.; Wang, L.; Alam, S.K.; Hoeppner, L.H.; Yang, R. ScanNeo: Identifying indel-derived neoantigens using RNA-Seq data. Bioinformatics 2019, 35, 4159–4161. [Google Scholar] [CrossRef] [PubMed]
- Maselli, F.M.; Giuliani, F.; Laface, C.; Perrone, M.; Melaccio, A.; De Santis, P.; Santoro, A.N.; Guarini, C.; Iaia, M.L.; Fedele, P. Immunotherapy in Prostate Cancer: State of Art and New Therapeutic Perspectives. Curr. Oncol. 2023, 30, 5769–5794. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Qian, X.; Liu, J.; Xue, F.; Luo, J.; Yao, G.; Yan, J.; Liu, X.; Xiao, B.; Li, J. Radiotherapy plus immune checkpoint inhibitor in prostate cancer. Front. Oncol. 2023, 13, 1210673. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Lee, K.; Kim, I.Y. Current update on prostate cancer immunotherapy. Korean J. Urol. Oncol. 2023, 21, 14–22. [Google Scholar] [CrossRef]
- Lampe, H.; Tam, L.; Hansen, A.R. Bi-specific T-cell engagers (BiTEs) in prostate cancer and strategies to enhance development: Hope for a BiTE-r future. Front. Pharmacol. 2024, 15, 1399802. [Google Scholar] [CrossRef]
- Aggarwal, R.; Starzinski, S.; de Kouchkovsky, I.; Koshkin, V.; Bose, R.; Chou, J.; Desai, A.; Kwon, D.; Kaushal, S.; Trihy, L.; et al. Single-dose (177)Lu-PSMA-617 followed by maintenance pembrolizumab in patients with metastatic castration-resistant prostate cancer: An open-label, dose-expansion, phase 1 trial. Lancet Oncol. 2023, 24, 1266–1276. [Google Scholar] [CrossRef]
- Grover, P.; Nunez-Cruz, S.; Leferovich, J.; Wentz, T.; Bagchi, A.; Milone, M.C.; Greene, M.I. F77 antigen is a promising target for adoptive T cell therapy of prostate cancer. Biochem. Biophys. Res. Commun. 2023, 680, 51–60. [Google Scholar] [CrossRef]
- Wei, Q.; Costanzi, S.; Balasubramanian, R.; Gao, Z.G.; Jacobson, K.A. A2B adenosine receptor blockade inhibits growth of prostate cancer cells. Purinergic Signal 2013, 9, 271–280. [Google Scholar] [CrossRef]
- Sandhu, S.; Subramaniam, S.; Hofman, M.S.; Stockler, M.R.; Martin, A.J.; Pokorski, I.; Goh, J.C.; Pattison, D.A.; Dhiantravan, N.; Gedye, C. Evolution: Phase II study of radionuclide 177Lu-PSMA-617 therapy versus 177Lu-PSMA-617 in combination with ipilimumab and nivolumab for men with metastatic castration-resistant prostate cancer (mCRPC; ANZUP 2001). J. Clin. Oncol. 2023, 41, TPS271. [Google Scholar] [CrossRef]
- Huang, L.; Xie, Y.; Jiang, S.; Dai, T.; Xu, Z.; Shan, H. Insights into immune microenvironment and therapeutic targeting in androgen-associated prostate cancer subtypes. Sci. Rep. 2024, 14, 18036. [Google Scholar] [CrossRef]
- Meng, L.; Yang, Y.; Mortazavi, A.; Zhang, J. Emerging Immunotherapy Approaches for Treating Prostate Cancer. Int. J. Mol. Sci. 2023, 24, 14347. [Google Scholar] [CrossRef] [PubMed]
- Pulido, R.; Lopez, J.I.; Nunes-Xavier, C.E. B7-H3: A robust target for immunotherapy in prostate cancer. Trends Cancer 2024, 10, 584–587. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, M.; Wang, M.; Wang, H.; Wu, H.; Mao, L.; Zhang, M.; Li, H.; Zheng, J.; Ma, P.; et al. B7-H3 specific CAR-T cells exhibit potent activity against prostate cancer. Cell Death Discov. 2023, 9, 147. [Google Scholar] [CrossRef] [PubMed]
- Yap, T.A.; Winter, J.N.; Giulino-Roth, L.; Longley, J.; Lopez, J.; Michot, J.M.; Leonard, J.P.; Ribrag, V.; McCabe, M.T.; Creasy, C.L.; et al. Phase I Study of the Novel Enhancer of Zeste Homolog 2 (EZH2) Inhibitor GSK2816126 in Patients with Advanced Hematologic and Solid Tumors. Clin. Cancer Res. 2019, 25, 7331–7339. [Google Scholar] [CrossRef]
- Deng, K.; Zou, Y.; Zou, C.; Wang, H.; Xiang, Y.; Yang, X.; Yang, S.; Cui, C.; Yang, G.; Huang, J. Study on pharmacokinetic interactions between SHR2554 and itraconazole in healthy subjects: A single-center, open-label phase I trial. Cancer Med. 2023, 12, 1431–1440. [Google Scholar] [CrossRef]
- Choudhury, A.D.; Kwak, L.; Cheung, A.; Allaire, K.M.; Marquez, J.; Yang, D.D.; Tripathi, A.; Kilar, J.M.; Flynn, M.; Maynard, B.; et al. Randomized Phase II Study Evaluating the Addition of Pembrolizumab to Radium-223 in Metastatic Castration-resistant Prostate Cancer. Cancer Immunol. Res. 2024, 12, 704–718. [Google Scholar] [CrossRef]
- Agarwal, N.; Azad, A.; Carles, J.; Matsubara, N.; Oudard, S.; Saad, F.; Merseburger, A.S.; Soares, A.; McGregor, B.A.; Zurawski, B. CONTACT-02: Phase 3 study of cabozantinib (C) plus atezolizumab (A) vs second novel hormonal therapy (NHT) in patients (pts) with metastatic castration-resistant prostate cancer (mCRPC). J. Clin. Oncol. 2024, 42, 18. [Google Scholar] [CrossRef]
- Fan, Z.; Oh, D.Y.; Wong, A.; Shinohara, K.; Nguyen, H.; Hwang, C.; Chang, H.; Starzinski, A.; Li, T.; De Leon, C. 862 TLR9 agonism promotes cytotoxic T cell persistence and myeloid remodeling when combined with radiation therapy and PD-1 blockade in oligometastatic prostate cancer patients. J. ImmunoTher. Cancer 2023, 11. [Google Scholar] [CrossRef]
- Linch, M.D.; Leone, G.; Wong, Y.N.S.; Jones, R.J.; Sankey, P.; Josephs, D.H.; Crabb, S.J.; Harris, L.; Tasnim, A.; Rashid, M. Nivolumab and ipilimumab for metastatic prostate cancer with an immunogenic signature: The NEPTUNES multi-centre two-cohort, biomarker-selected phase 2 trial. J. Clin. Oncol. 2024, 42, 5013. [Google Scholar] [CrossRef]
- Sharma, P.; Krainer, M.; Saad, F.; Castellano, D.; Bedke, J.; Kwiatkowski, M.; Patnaik, A.; Procopio, G.; Wiechno, P.; Kochuparambil, S.T. Nivolumab plus ipilimumab for the treatment of post-chemotherapy metastatic castration-resistant prostate cancer (mCRPC): Additional results from the randomized phase 2 CheckMate 650 trial. J. Clin. Oncol. 2023, 41, 22. [Google Scholar] [CrossRef]
- Tsimberidou, A.M.; Drakaki, A.; Khalil, D.N.; Okrah, K.; Schachter, M.; Gautam, S.; Da Silva, D.M.; Alayli, F.A.; Spencer, C.N.; Foo, W.C. Evaluating nivolumab (nivo) plus ipilimumab (ipi) in patients with metastatic castration-resistant prostate cancer (mCRPC): Clinical and translational results from the AMADEUS prostate expansion cohort. In Proceedings of the American Association for Cancer Research Annual Meeting 2024, San Diego, CA, USA, 5–10 April 2024. [Google Scholar]
- Kelly, W.; Danila, D.; Lin, C.; Lee, J.; Matsubara, N.; Ward, P.; Armstrong, A.; Pook, D.; Kim, M.; Dorff, T. 1765O Interim results from a phase I study of AMG 509 (xaluritamig), a STEAP1 x CD3 XmAb 2+1 immune therapy, in patients with metastatic castration-resistant prostate cancer (mCRPC). Ann. Oncol. 2023, 34, S953–S954. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).