
Increased Concentration of Anti-Egg Albumin Antibodies in Cerebrospinal Fluid and Serum of Patients with Alzheimer’s Disease—Discussion on Human Serpins’ Similarity and Probable Involvement in the Disease Mechanism

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Classification of AD Severity
2.2.2. Measurement of Anti-Egg Albumin, Anti-Casein, and Anti-Neu5Gc Antibodies [3]
2.2.3. Statistical Analysis
2.2.4. Protein Similarity Search
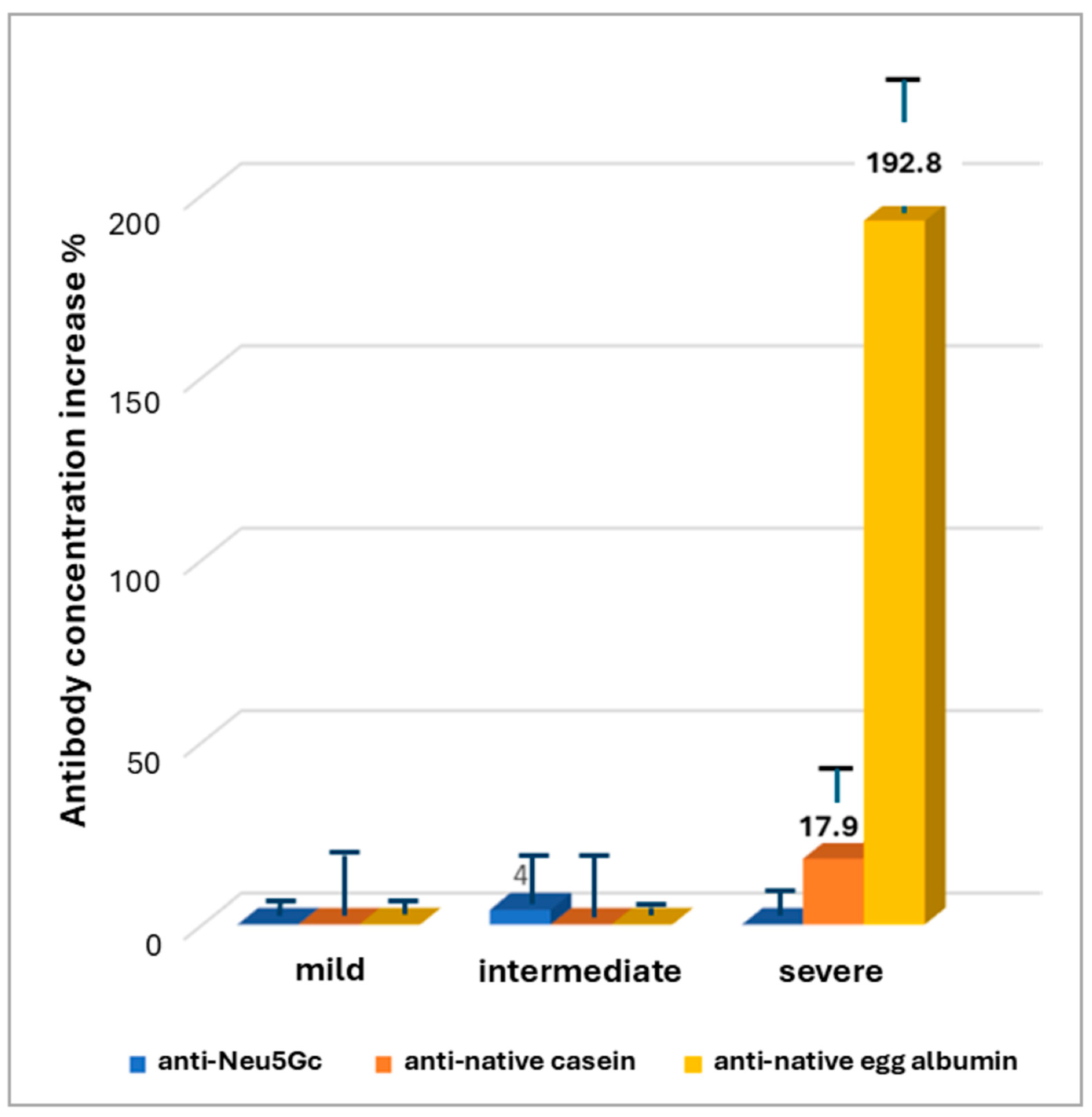
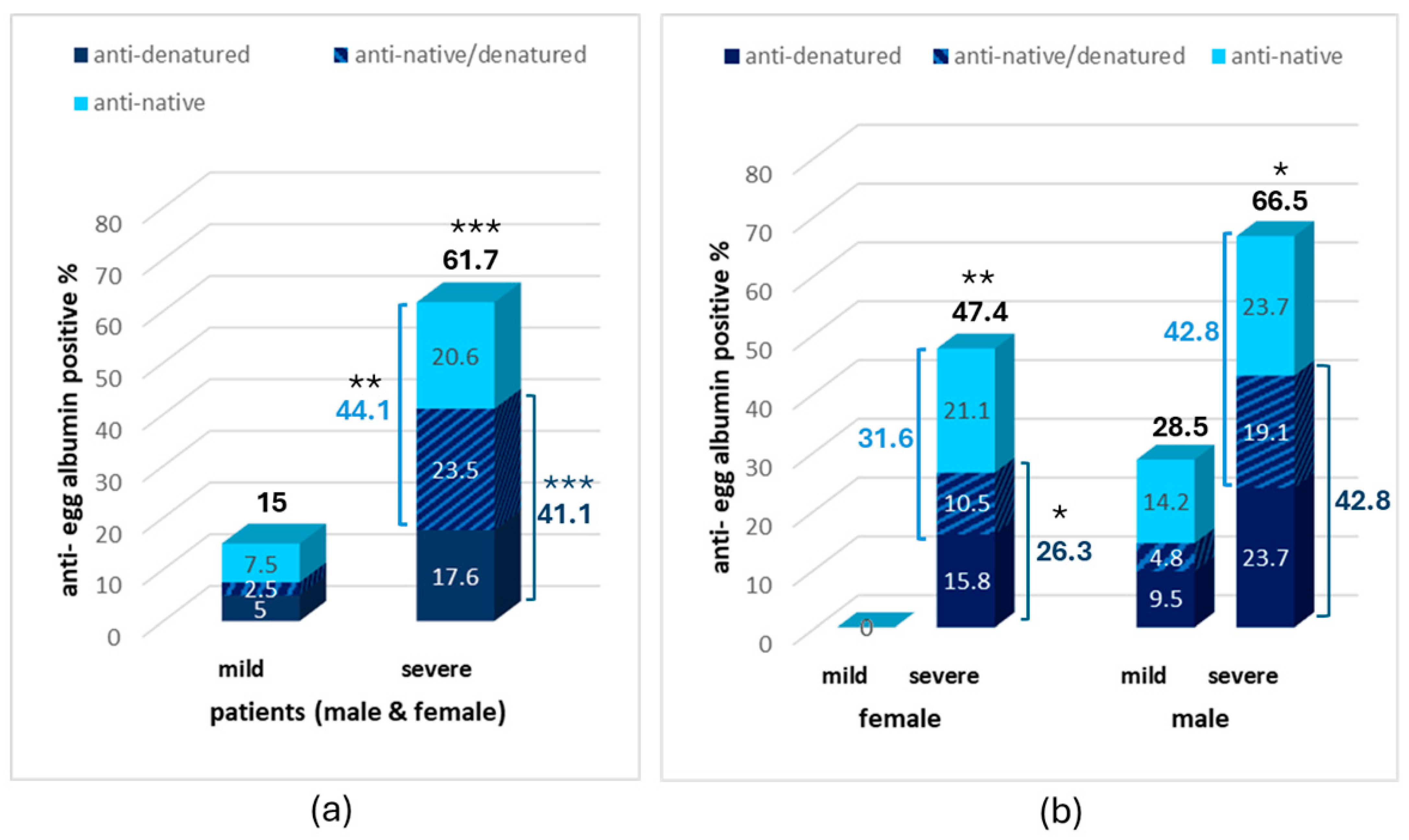
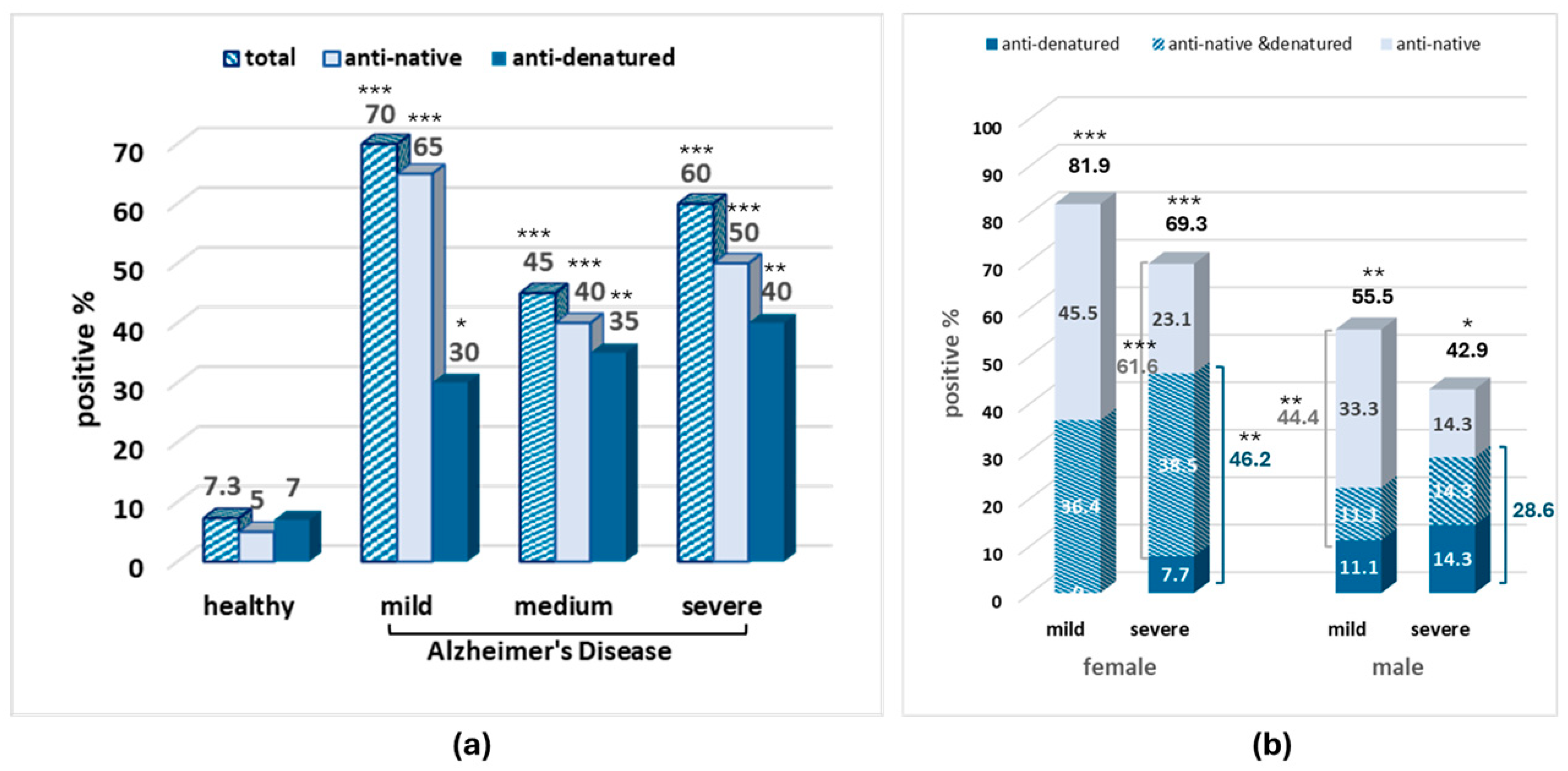
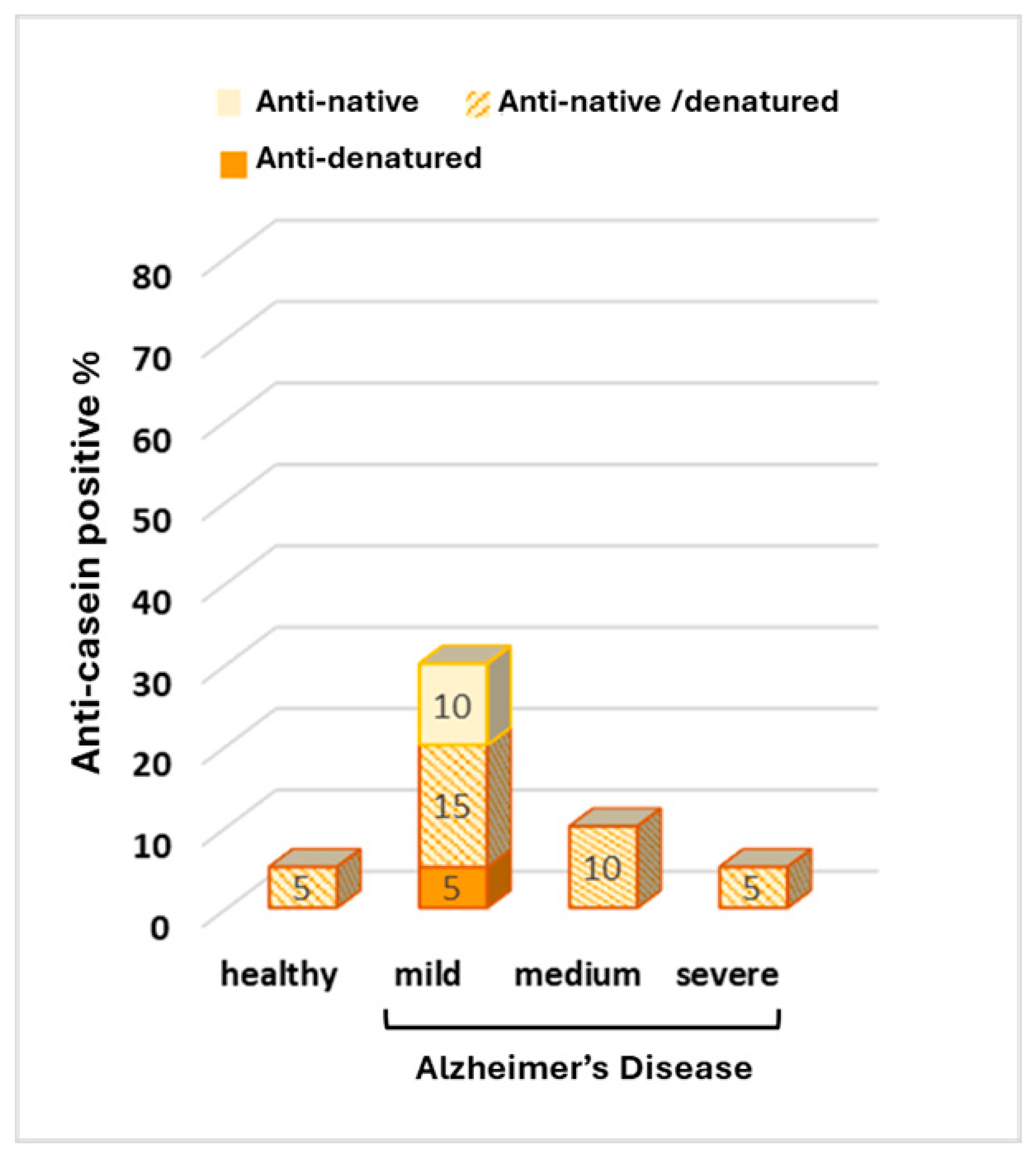
3. Results
4. Discussion
4.1. Main Observations
4.2. Homology with Human Proteins
4.3. Intestinal Permeability and Inflammation
4.4. Blood–Brain Barrier
4.5. CNS
4.6. Aspects on Prevention and Therapeutic Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s Disease |
| BBB BLAST BLASTp | Blood–Brain Barrier Basic Local Alignment Search Tool Basic Local Alignment Search Tool for proteins |
| MBP | Myelin Basic Protein |
| CSF | Cerebrospinal Fluid |
| Neu5Gc | Glycolylneuraminic Acid |
| I.H.U. | International Hellenic University |
| A.U.TH. | Aristotle University of Thessaloniki |
| MMSE | Mini-Mental State Examination |
| AGT | Angiotensin |
| ERK3 | Extracellular Signal-Regulated Kinase 3 |
| MAPK | Mitogen-Activated Protein Kinase |
| TRK | Tyrosine Receptor Kinase |
| NAD(P)H | Nicotinamide Adenine Dinucleotide Phosphate |
| NF-KappaB | Nuclear Factor-Kappa B |
| ER | Endoplasmic Reticulum |
| GZMH | Granzyme H |
| SCCA | Squamous Cell Carcinoma Antigen |
| HMSD | Histocompatibility Minor Serpin Domain Containing |
| HMW | High Molecular Weight |
| PIP4K2C | Phosphatidylinositol 5-Phosphate 4-Kinase Type-2 Gamma |
| RENBP | Renin-Binding Protein |
| PAI-1 | Plasminogen Activator Inhibitor 1 |
| PLAT | Tissue-Type Plasminogen Activator |
| PLAU | Urokinase-Type Plasminogen Activator |
| SEC | Serpin-Enzyme Complex |
| CNS | Central Nervous System |
| Aβ | Amyloid-Beta |
| AβPP | Amyloid-Beta Precursor Protein |
| AGTR | Angiotensin Receptor |
| MD | Mediterranean Diet |
| BSA | Bovine Serum Albumin |
| SDS | Sodium Dodecylsulfate |
| MW | Molecular Weight |
References
- Uzunismail, H.; Cengiz, M.; Uzun, H.; Ozbakir, F.; Goksel, S.; Demirdag, F.; Can, G.; Balci, H. The effects of provocation by foods with raised IgG antibodies and additives on the course of Crohn’s disease: A pilot study. Turk. J. Gastroenterol. 2012, 23, 19–27. [Google Scholar] [CrossRef]
- Cappelletti, M.; Tognon, E.; Vona, L.; Basello, K.; Costanzi, A.; Speciani, M.C.; Speciani, A.F. Food-specific serum IgG and symptom reduction with a personalized, unrestricted-calorie diet of six weeks in Irritable Bowel Syndrome (IBS). Nutr. Metab. 2020, 17, 101. [Google Scholar] [CrossRef]
- Eleftheriou, P.; Kynigopoulos, S.; Giovou, A.; Mazmanidi, A.; Yovos, J.; Skepastianos, P.; Vagdatli, E.; Petrou, C.; Papara, D.; Efterpiou, M. Prevalence of anti-Neu5Gc antibodies in patients with hypothyroidism. Biomed. Res. Int. 2014, 2014, 963230. [Google Scholar] [CrossRef]
- Padler-Karavani, V.; Hurtado-Ziola, N.; Pu, M.; Yu, H.; Huang, S.; Muthana, S.; Chokhawala, H.A.; Cao, H.; Secrest, P.; Friedmann-Morvinski, D.; et al. Human xeno-autoantibodies against a non-human sialic acid serve as novel serum biomarkers and immunotherapeutics in cancer. Cancer Res. 2011, 71, 3352–3363. [Google Scholar] [CrossRef]
- Geiselman, J.F. The clinical use of IgG food sensitivity testing with migraine headache patients: A literature review. Curr. Pain Headache Rep. 2019, 23, 79. [Google Scholar] [CrossRef]
- Chundera, R.; Weiera, A.; Maurerb, H.; Luberb, N.; Endersa, M.; Luberc, G.; Heiderd, T.; Spitzere, A.; Tackeb, S.; Becker-Gototf, J.; et al. Antibody cross-reactivity between casein and myelin-associated glycoprotein results in central nervous system demyelination. Proc. Natl. Acad. Sci. USA 2022, 119, e2117034119. [Google Scholar] [CrossRef]
- Hadjivassiliou, M.; Sanders, D.S.; Grünewald, R.A.; Woodroofe, N.; Boscolo, S.; Aeschlimann, D. Gluten sensitivity: From gut to brain. Lancet Neurol. 2010, 9, 318–330. [Google Scholar] [CrossRef]
- Versino, M.; Biagi, F.; Bianchi, P.I.; Zardini, E.; Colnaghi, S.; Moglia, A.; Corazza, G.R.; Franciotta, D. Gluten sensitivity and the CNS: Diagnosis and treatment. Lancet Neurol. 2010, 9, 653. [Google Scholar] [CrossRef]
- Karakula-Juchnowicz, H.; Galecka, M.; Rog, J.; Bartnicka, A.; Lukaszewicz, Z.; Krukow, P.; Morylowska-Topolska, J.; Skonieczna-Zydecka, K.; Krajka, T.; Jonak, K.; et al. The food-specific serum IgG reactivity in major depressive disorder patients, irritable bowel syndrome patients and healthy controls. Nutrients 2018, 10, 548. [Google Scholar] [CrossRef]
- Severance, E.G.; Dupont, D.; Dickerson, F.B.; Stallings, C.R.; Origoni, A.E.; Krivogorsky, B.; Yang, S.; Haasnoot, W.; Yolken, R.H. Immune activation by casein dietary antigens in bipolar disorder. Bipolar Disord. 2010, 12, 834–842. [Google Scholar] [CrossRef]
- Karakula-Juchnowicz, H.; Szachta, P.; Opolska, A.; Morylowska-Topolska, J.; Galecka, M.; Juchnowicz, D.; Krukow, P.; Lasik, Z. The role of IgG hypersensitivity in the pathogenesis and therapy of depressive disorders. Nutr. Neurosci. 2017, 20, 110–118. [Google Scholar] [CrossRef]
- Vojdani, A. Reaction of food-specific antibodies with different tissue antigens. Int. J. Food Sci. Technol. 2020, 55, 1800–1815. [Google Scholar] [CrossRef]
- Adams, D.H.; Eksteen, B. Aberrant homing of mucosal T cells and extra-intestinal manifestations of inflammatory bowel disease. Nat. Rev. Immunol. 2006, 6, 244–251. [Google Scholar] [CrossRef]
- Molberg, Ø.; Sollid, L.M. A gut feeling for joint inflammation—Using coeliac disease to understand rheumatoid arthritis. Trends Immunol. 2006, 27, 188–194. [Google Scholar] [CrossRef]
- Vojdani, A. A potential link between environmental triggers and autoimmunity. Autoimmune Dis. 2014, 2024, 437231. [Google Scholar] [CrossRef]
- De Magistris, L.; Picardi, A.; Siniscalco, D.; Riccio, M.P.; Sapone, A.; Cariello, R.; Abbadessa, S.; Medici, N.; Lammers, K.M.; Schiraldi, C.; et al. Antibodies against Food Antigens in Patients with Autistic Spectrum Disorders. BioMed Res. Int. 2013, 2013, 729349. [Google Scholar] [CrossRef]
- Rowland, L.M.; Demyanovich, H.K.; Wijtenburg, S.A.; Eaton, W.W.; Rodriguez, K.; Gaston, F.; Cihakova, D.; Talor, M.V.; Liu, F.; McMahon, R.R.; et al. Antigliadin Antibodies (AgA IgG) Are Related to Neurochemistry in Schizophrenia. Front. Psychiatry 2017, 8, 104. [Google Scholar] [CrossRef]
- Bentz, S.; Hausmann, M.; Piberger, H.; Kellermeier, S.; Paul, S.; Held, L.; Falk, W.; Obermeier, F.; Fried, M.; Schölmerich, J.; et al. Clinical relevance of IgG antibodies against food antigens in Crohn’s disease: A double-blind cross-over diet intervention study. Digestion 2010, 81, 252–264. [Google Scholar] [CrossRef]
- Atkinson, W.; Sheldon, T.A.; Shaath, N.; Whorwell, P.J. Food elimination based on IgG anti-bodies in irritable bowel syndrome: A randomised controlled trial. Gut 2004, 53, 1459–1464. [Google Scholar] [CrossRef]
- Mitchell, N.; Hewitt, C.E.; Jayakody, S.; Islam, M.; Adamson, J.; Watt, I.; Torgerson, D.J. Randomised con-trolled trial of food elimination diet based on IgG antibodies for the prevention of migraine like headaches. Nutr. J. 2011, 10, 85. [Google Scholar] [CrossRef]
- Xie, Y.; Zhou, G.; Xu, Y.; He, B.; Wang, Y.; Ma, R.; Chang, Y.; He, D.; Xu, C.; Xiao, Z. Effects of diet based on IgG elimination combined with probiotics on migraine plus irritable bowel syndrome. Pain Res. Manag. 2019, 2019, 7890461. [Google Scholar] [CrossRef]
- Jian, L.; Anqi, H.; Gang, L.; Litian, W.; Yanyan, X.; Mengdi, W.; Tong, L. Food exclusion based on IgG antibodies alleviates symptoms in ulcerative colitis: A prospective study. Inflamm. Bowel Dis. 2018, 24, 1918–1925. [Google Scholar] [CrossRef]
- Gunasekeera, V.; Mendall, M.A.; Chan, D.; Kumar, D. Treatment of Crohn’s disease with an IgG4-guided exclusion diet: A randomized controlled trial. Dig. Dis. Sci. 2016, 61, 1148–1157. [Google Scholar] [CrossRef]
- Ullah, R.; Park, T.J.; Huang, X.; Kim, M.O. Abnormal amyloid beta metabolism in systemic abnormalities and Alzheimer’s pathology: Insights and therapeutic approaches from periphery. Ageing Res. Rev. 2021, 71, 101451. [Google Scholar] [CrossRef]
- De Bruijn, R.F.A.G.; Ikram, M.A. Cardiovascular risk factors and future risk of Alzheimer’s disease. BMC Med. 2014, 12, 130. [Google Scholar] [CrossRef]
- Biessels, G.J.; Staekenborg, S.; Brunner, E.; Brayne, C.; Scheltens, P. Risk of dementia in diabetes mellitus: A systematic review. Lancet Neurol. 2006, 5, 64–74. [Google Scholar] [CrossRef]
- Uranga, R.M.; Keller, J.N. Diet and age interactions with regards to cholesterol regulation and brain pathogenesis. Curr. Gerontol. Geriatr. Res. 2010, 2010, 219683. [Google Scholar] [CrossRef]
- Fann, J.R.; Ribe, A.R.; Pedersen, H.S.; Fenger-Grøn, M.; Christensen, J.; Benros, M.E.; Vestergaard, M. Long-term risk of dementia among people with traumatic brain injury in Denmark: A population-based observational cohort study. Lancet Psychiatry 2018, 5, 424–431. [Google Scholar] [CrossRef]
- Chen, H.; Kwong, J.C.; Copes, R.; Tu, K.; Villeneuve, P.J.; van Donkelaar, A.; Hystad, P.; Martin, R.V.; Murray, B.J.; Jessiman, B.; et al. Living near major roads and the incidence of dementia, Parkinson’s disease, and multiple sclerosis: A population-based cohort study. Lancet 2017, 389, 718–726. [Google Scholar] [CrossRef]
- Butterfield, D.A.; Di Domenico, F.; Barone, E. Elevated risk of type 2 diabetes for development of Alzheimer disease: A key role for oxidative stress in brain. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 1693–1706. [Google Scholar] [CrossRef]
- Engelhart, M.J.; Geerlings, M.I.; Meijer, J.; Kiliaan, A.; Ruitenberg, A.; van Swieten, J.C.; Stijnen, T.; Hofman, A.; Witteman, J.C.M.; Breteler, M.M.B. Inflammatory proteins in plasma and the risk of dementia. Arch. Neurol. 2004, 61, 668–672. [Google Scholar] [CrossRef]
- Akbaraly, T.N.; Singh-Manoux, A.; Dugravot, A.; Brunner, E.J.; Kivimäki, M.; Sabia, S. Association of midlife diet with subsequent risk for dementia. J. Am. Med. Assoc. 2019, 321, 957–968. [Google Scholar] [CrossRef]
- Huang, S.Y.; Yang, Y.X.; Kuo, K.; Li, H.Q.; Shen, X.N.; Chen, S.D.; Cui, M.; Tan, L.; Dong, Q.; Yu, J.T. Herpesvirus infections and Alzheimer’s disease: A Mendelian randomization study. Alzheimer’s Res. Ther. 2021, 13, 158. [Google Scholar] [CrossRef]
- Yang, H.Y.; Chien, W.C.; Chung, C.H.; Su, R.Y.; Lai, C.Y.; Yang, C.C.; Tzeng, N.S. Risk of dementia in patients with toxoplasmosis: A nationwide, population based cohort study in Taiwan. Parasites Vectors 2021, 14, 435. [Google Scholar] [CrossRef]
- Wu, S.; Liu, X.; Jiang, R.; Yan, X.; Ling, Z. Roles and Mechanisms of Gut Microbiota in Patients With Alzheimer’s Disease. Front. Aging Neurosci. 2021, 13, 650047. [Google Scholar] [CrossRef]
- Kilshaw, P.J.; McEwan, F.J.; Baker, K.C.; Cant, A.J. Studies on the specificity of antibodies to ovalbumin in normal human serum: Technical considerations in the use of ELISA methods. Clin. Exp. Immunol. 1986, 66, 481–489. [Google Scholar]
- Yang, W.; Jiang, Y.; Guo, Q.; Tian, Z.; Cheng, Z. Aberrant N-glycolylneuraminic acid in breast MCF-7 cancer cells and cancer stem cells. Front. Mol. Biosci. 2022, 9, 1047672. [Google Scholar] [CrossRef]
- Photchanachai, S.; Mehta, A.; Kitabatake, N. Heating of an Ovalbumin Solution at Neutral pH and High Temperature. Biosci. Biotechnol. Biochem. 2002, 66, 1635–1640. [Google Scholar] [CrossRef]
- Conover, W.J. Practical Nonparametric Statistics, 3rd ed.; John Wiley & Sons: New York, NY, USA, 1999. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality. Biometrika 1999, 52, 591–599. [Google Scholar] [CrossRef]
- Lentner, C.G. Scientific Tablesed, 8th ed.; Ciba-Geigy: Basel, Switzerland, 1982; Volume 2. [Google Scholar]
- Rosner, B. Fundamentals of Biostatistics, 6th ed.; Tomson Brooks/Cole: Pacific Grove, CA, USA, 2006. [Google Scholar]
- Kim, H.-Y. Statistical notes for clinical researchers: Chi-squared test and Fisher’s exact test. Restor. Dent. Endod. 2017, 42, 152–155. [Google Scholar] [CrossRef]
- Kheirkhah, R.; DeMarshall, C.; Sieber, F.; Oh, E.; Nagele, R.G. The origin and nature of the complex autoantibody profile in cerebrospinal fluid. Brain Behav. Immun. Health 2020, 2, 100032. [Google Scholar] [CrossRef]
- Aiello, A.; Farzaneh, F.; Candore, G.; Caruso, C.; Davinelli, S.; Gambino, C.M.; Ligotti, M.E.; Zareian, N.; Accardi, G. Immunosenescence and Its Hallmarks: How to Oppose Aging Strategically? A Review of Potential Options for Therapeutic Intervention. Front. Immunol. 2019, 10, 2247. [Google Scholar] [CrossRef]
- Tushar, K.; Ganesh, D.; Ganesh, B.P. Interlink between the gut microbiota and inflammation in the context of oxidative stress in Alzheimer’s disease progression. Gut Microbes 2023, 15, 2206504. [Google Scholar] [CrossRef]
- Honarpisheh, P.; Reynolds, C.R.; Blasco Conesa, M.P.; Moruno Manchon, J.F.; Putluri, N.; Bhattacharjee, M.B.; Urayama, A.; McCullough, L.D.; Ganesh, B.P. Ganesh Dysregulated Gut Homeostasis Observed Prior to the Accumulation of the Brain Amyloid in Tg2576 Mice. Int. J. Mol. Sci. 2020, 21, 1711. [Google Scholar] [CrossRef]
- Seidler, N.W.; Yeargans, G.S. Albumin-bound polyacrolein: Implications for Alzheimer’s disease. Biochem. Biophys. Res. Commun. 2004, 320, 213–217. [Google Scholar] [CrossRef]
- Tsutsui, Y.; Kuri, B.; Sengupta, T.; Wintrode, P.L. The Structural Basis of Serpin Polymerization Studied by Hydrogen/Deuterium Exchange and Mass Spectrometry. J. Biol. Chem. 2008, 283, 30804–30811. [Google Scholar] [CrossRef]
- Lambrecht, M.A.; Rombouts, I.; De Ketelaere, B.; Delcour, J.A. Prediction of heat-induced polymerization of different globular food proteins in mixtures with wheat gluten. Food Chem. 2017, 221, 1158–1167. [Google Scholar] [CrossRef]
- Abelein, a.; Kaspersen, J.D.; Nielsen, S.B.; Vestergaard Jensen, G.; Christiansen, G.; Pedersen, J.S.; Danielsson, J.; Otzen, D.E.; Graslund, A. Formation of Dynamic Soluble Surfactant-induced Amyloid Peptide Aggregation Intermediates. J. Biol. Chem. 2013, 288, 23518–23528. [Google Scholar] [CrossRef]
- Nybo, M.; Andersen, K.; Sorensen, G.L.; Lolk, A.; Kragh-Sorensen, P.; Holmskov, U. Serum surfactant protein D is correlated to development of dementia and augmented mortality. Clin. Immunol. 2007, 123, 333–337. [Google Scholar] [CrossRef]
- Kannan, S.; Ashokkumar, K.; Krishnamoorthy, G.; Dhasayan, A.; Marudhamuthu, M. Monitoring surfactant mediated defence of gastrointestinal Proteus mirabilis DMTMMK1against pathogenic consortia of Vibrio cholerae. RSC Adv. 2017, 7, 20969–20980. [Google Scholar] [CrossRef]
- Jiang, J.; Zang, J.; Qing, M.; Ma, Y.; Yang, X.; Chi, Y.; Chi, Y. Regulating the thermal properties of egg white by adding surfactants. J. Food Eng. 2024, 362, 111759. [Google Scholar] [CrossRef]
- Cremers, C.M.; Jakob, U. Oxidant Sensing by Reversible Disulfide Bond. J. Biol. Chem. 2013, 288, 26489–26496. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Bistritz, L.; Meddings, J.B. Recent advances in clinical practice. Alterations in intestinal permeability. Gut 2006, 55, 1512–1520. [Google Scholar] [CrossRef]
- Heston, M.B.; Hanslik, K.L.; Zarbock, K.R.; Harding, S.J.; Davenport-Sis, N.J.; Kerby, R.L.; Chin, N.; Sun, Y.; Hoeft, A.; Deming, Y.; et al. Gut inflammation associated with age and Alzheimer’s disease pathology: A human cohort study. Sci. Rep. 2023, 13, 18924. [Google Scholar] [CrossRef]
- Li, Z.; Zhu, H.; Zhang, L.; Qin, C. The intestinal microbiome and Alzheimer’s disease: A review. Animal Models. Exp. Med. 2018, 1, 180–188. [Google Scholar] [CrossRef]
- Pellegrini, C.; Daniele, S.; Antonioli, L.; Benvenuti, L.; D’Antongiovanni, V.; Piccarducci, R.; Pietrobono, D.; Citi, V.; Piragine, E.; Flori, L.; et al. Prodromal Intestinal Events in Alzheimer’s Disease (AD): Colonic Dysmotility and Inflammation Are Associated with Enteric AD-Related Protein Deposition. Int. J. Mol. Sci. 2020, 21, 3523. [Google Scholar] [CrossRef]
- Yoon, K.; Kim, N. Roles of Sex Hormones and Gender in the Gut Microbiota. J. Neurogastroenterol. Motil. 2021, 27, 314–325. [Google Scholar] [CrossRef]
- Kim, Y.S.; Unno, T.; Kim, B.-Y.; Park, M.-S. Sex Differences in Gut Microbiota. World J. Men’s Health 2020, 38, 48–60. [Google Scholar] [CrossRef]
- Towera, J.; Pomattod, L.C.D.; Daviesa, K.J.A. Sex differences in the response to oxidative and proteolytic stress. Redox Biol. 2020, 31, 101488. [Google Scholar] [CrossRef]
- Kander, M.C.; Cui, Y.; Liu, Z. Gender difference in oxidative stress: A new look at the mechanisms for cardiovascular diseases. J. Cell. Mol. Med. 2017, 21, 1024–1032. [Google Scholar] [CrossRef]
- Tiberi, J.; Cesarini, V.; Stefanelli, R.; Canterini, S.; Fiorenza, M.T.; La Rosa, P. Sex differences in antioxidant defence and the regulation of redox homeostasis in physiology and pathology. Mech. Ageing Dev. 2023, 211, 111802. [Google Scholar] [CrossRef]
- Castellazzi, M.; Morotti, A.; Tamborino, C.; Alessi, F.; Pilotto, S.; Baldi, E.; Caniatti, L.M.; Trentini, A.; Casetta, I.; Granieri, E.; et al. Increased age and male sex are independently associated with higher frequency of blood–cerebrospinal fluid barrier dysfunction using the albumin quotient. Fluids Barriers CNS 2020, 17, 14. [Google Scholar] [CrossRef]
- Vojdani, A.; O’Bryan, T.; Green, J.A.; Mccandless, J.; Woeller, K.N.; Vojdani, E.; Nourian, A.A.; Cooper, E.L. Immune response to dietary proteins, gliadin and cerebellar peptides in children with autism. Nutr. Neurosci. 2004, 7, 151–161. [Google Scholar] [CrossRef]
- Cie’ sli’ nska, A.; Fiedorowicz, E.; Rozmus, D.; Sienkiewicz-Szłapka, E.; Jarmołowska, B.; Kami’ nski, S. Does a Little Difference Make a Big Difference? Bovine-Casein A1 and A2Variants and Human Health—An Update. Int. J. Mol. Sci. 2022, 23, 15637. [Google Scholar] [CrossRef]
- Maggonage, M.H.U.; Manjula, P.; Ahn, D.U.; Nalaka, E.D.; Abeyrathne, S. Ovalbumin: A potential functional protein. Food Sci. Preserv. 2024, 31, 346–359. [Google Scholar] [CrossRef]
- Zattoni, M.; Mearelli, M.; Vanni, S.; Colini Baldeschi, A.; Tran, T.H.; Ferracin, C.; Catania, M.; Moda, F.; Di Fede, G.; Giaccone, G.; et al. Serpin Signatures in Prion and Alzheimer’s Diseases. Mol. Neurobiol. 2022, 59, 3778–3799. [Google Scholar] [CrossRef]
- Wosik, K.; Cayrol, R.; Dodelet-Devillers, A.; Berthelet, F.; Bernard, M.; Moumdjian, R.; Bouthillier, A.; Reudelhuber, T.L.; Prat, A. Angiotensin II Controls Occludin Function and Is Required for Blood–Brain Barrier Maintenance: Relevance to Multiple Sclerosis. J. Neurosci. 2007, 27, 9032–9042. [Google Scholar] [CrossRef]
- Mogi, M.; Iwanami, J.; Horiuchi, M. Roles of Brain Angiotensin II in Cognitive Function and Dementia. Int. J. Hypertens. 2012, 2012, 169649. [Google Scholar] [CrossRef]
- Royea, J.; Hamel, E. Brain angiotensin II and angiotensin IV receptors as potential Alzheimer’s disease therapeutic targets. GeroScience 2020, 42, 1237–1256. [Google Scholar] [CrossRef]
- Gebre, A.K.; Altaye, B.M.; Atey, T.M.; Tuem, K.B.; Berhe, D.F. Targeting Renin–Angiotensin System Against Alzheimer’s Disease. Front. Pharmacol. 2018, 9, 440. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, S.; Choi, Y.; Nielsen, T.B.; Yan, J.; Lu, A.; Ruan, J.; Lee, H.-R.; Wu, H.; Spellberg, B.; et al. 8SERPINB1-mediated checkpoint of inflammatory caspase activation. Nat. Immunol. 2019, 20, 276–287. [Google Scholar] [CrossRef]
- Wernersson, S.; Pejler, G. Mast cell secretory granules: Armed for battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- Meyer-Hoffert, U.; Wiedow, O. Neutrophil serine proteases: Mediators of innate immune responses. Curr. Opin. Hematol. 2011, 18, 19–24. [Google Scholar] [CrossRef]
- Dabek, M.; Ferrier, L.; Annahazi, A.; Bézirard, V.; Polizzi, A.; Cartier, C.; Leveque, M.; Roka, R.; Wittmann, T.; Theodorou, V.; et al. Intracolonic infusion of fecal supernatants from ulcerative colitis patients triggers altered permeability and inflammation in mice: Role of cathepsin G and protease-activated receptor-4. Inflamm. Bowel Dis. 2011, 17, 1409–1414. [Google Scholar] [CrossRef]
- Scudamore, C.L.; Jepson, M.A.; Hirst, B.H.; Miller, H.R. The rat mucosal mast cell chymase RMCP-II alters epithelial cell monolayer permeability in association with altered distribution of the tight junction proteins ZO-1 and occludin. Eur. J. Cell Biol. 1998, 75, 321–330. [Google Scholar] [CrossRef]
- Mellet, P.; Michels, B.; Bieth, J.G. Heat-induced Conversion of Ovalbumin into a Proteinase Inhibitor. J. Biol. Chem. 1996, 271, 30311–30314. [Google Scholar] [CrossRef]
- Joslin, G.; Griffin, G.L.; August, A.M.; Adams, S.; Fallon, R.J.; Senior, R.M.; Perlmutter, D.H. The Serpin-Enzyme Complex (SEC) Receptor Mediates the Neutrophil Chemotactic Effect of a-1 Antitrypsin-Elastase Complexes and Amyloid-beta Peptide. J. Clin. Investig. 1992, 90, 1150–1154. [Google Scholar] [CrossRef]
- Kim, Y.N.; Kim, D.H. Decreased serum angiogenin level in Alzheimer’s disease. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 38, 116–120. [Google Scholar] [CrossRef]
- Makuch-Martins, M.; Vieira-Morais, C.G.; Perego, S.M.; Ruggeri, A.; Ceroni, A.; Michelini, L.C. Angiotensin II, blood–brain barrier permeability, and microglia interplay during the transition from pre-to hypertensive phase in spontaneously hypertensive rats. Front. Physiol. 2024, 15, 1452959. [Google Scholar] [CrossRef]
- D’Acunto, E.; Fra, A.; Visentin, C.; Manno , M.; Ricagno, S.; Galliciotti, G.; Miranda, E. Neuroserpin: Structure, function, physiology and pathology. Cell. Mol. Life Sci. 2021, 78, 6409–6430. [Google Scholar] [CrossRef]
- Venugopal, C.; Demos, C.M.; Jagannatha Rao, K.S.; Pappolla, M.A.; Sambamurti, K. Beta-secretase: Structure, Function, and Evolution. CNS Neurol. Disord. Drug Targets 2008, 7, 278–294. [Google Scholar] [CrossRef]
- Saido, T.; Leissring, M.A. Proteolytic Degradation of Amyloid b-Protein. Cold Spring Harb. Perspect. Med. 2012, 2, a006379. [Google Scholar] [CrossRef]
- Abraham, C.R.; Selkoe, D.J.; Potter, H. Immunochemical identification of the serine protease inhibitor alpha 1-antichymotrypsin in the brain amyloid deposits of Alzheimer’s disease. Cell 1988, 52, 487–501. [Google Scholar] [CrossRef]
- Jucker, M.; Walker, L.C. Propagation and spread of pathogenic protein assemblies in neurodegenerative diseases. Nat. Neurosci. 2018, 21, 1341–1349. [Google Scholar] [CrossRef]
- Licastro, F.; Campbell, I.L.; Kincaid, C.; Veinbergs, I.; Van Uden, E.; Rockenstein, E.; Mallory, M.; Gilbert, J.R.; Masliah, E. A Role for apoE in Regulating the Levels of a-1-Antichymotrypsin in the Aging Mouse Brain and in Alzheimer’s Disease. Am. J. Pathol. 1999, 155, 869–875. [Google Scholar] [CrossRef]
- Harris, F.M.; Brecht, W.J.; Xu, Q.; Tesseur, I.; Kekonius, L.; Wyss-Coray, T.; Fish, J.D.; Masliah, E.; Hopkins, P.C.; Scearce-Levie, K.; et al. Carboxyl-terminal-truncated apolipoprotein E4 causes Alzheimer’s disease-like neurodegeneration and behavioral deficits in transgenic mice. Proc. Natl. Acad. Sci. USA 2003, 100, 10967. [Google Scholar] [CrossRef]
- Koo, E.H.; Abraham, C.R.; Potter, H.; Cork, L.C.; Price, D.L. Developmental expression of alpha 1-antichymotrypsin in brain may be related to astrogliosis. Neurobiol. Aging 1991, 12, 495–501. [Google Scholar] [CrossRef]
- Ishigami, S.; Sandkvist, M.; Tsui, F.; Moore, E.; Coleman, T.A.; Lawrence, D.A. Identification of a novel targeting sequence for regulated secretion in the serine protease inhibitor neuroserpin. Biochem. J. 2007, 402, 25–34. [Google Scholar] [CrossRef]
- Davis, R.L.; Shrimpton, A.E.; Holohan, P.D.; Bradshaw, C.; Feiglin, D.; Collins, G.H.; Sonderegger, P.; Kinter, J.; Becker, L.M.; Lacbawan, F.; et al. Familial dementia caused by polymerization of mutant neuroserpin. Nature 1999, 401, 376–379. [Google Scholar] [CrossRef]
- Kinghorn, K.J.; Crowther, D.C.; Sharp, L.K.; Nerelius, C.; Davis, R.L.; Chang, H.T.; Green, C.; Gubb, D.C.; Johansson, J.; Lomas, D.A. Neuroserpin binds Aβ and is a neuroprotective component of amyloid plaques in Alzheimer disease. J. Biol. Chem. 2006, 281, 29268–29277. [Google Scholar] [CrossRef]
- Barker, R.; Kehoe, P.G.; Love, S. Activators and inhibitors of the plasminogen system in Alzheimer’s disease. J. Cell. Mol. Med. 2012, 16, 865–876. [Google Scholar] [CrossRef]
- Tucker, H.M.; Kihiko, M.; Caldwell, J.N.; Wright, S.; Kawarabayashi, T.; Price, D.; Walker, D.; Scheff, S.; McGillis, J.P.; Rydel, R.E.; et al. The plasmin system is induced by and degrades amyloidbeta aggregates. J. Neurosci. 2000, 20, 3937–3946. [Google Scholar] [CrossRef]
- Godinez, A.; Rajput, R.; Chitranshi, N.; Gupta, V.; Basavarajappa, D.; Sharma, S.; You, Y.; Pushpitha, K.; Dhiman, K.; Mirzaei, M.; et al. Neuroserpin, a crucial regulator for axogenesis, synaptic modelling and cell–cell interactions in the pathophysiology of neurological disease. Cell. Mol. Life Sci. 2022, 79, 172. [Google Scholar] [CrossRef]
- Fabbro, S.; Seeds, N.W. Plasminogen activator activity is inhibited while neuroserpin is up-regulated in the Alzheimer disease brain. J. Neurochem. 2009, 109, 303–315. [Google Scholar] [CrossRef]
- Lee, T.W.; Tsang, V.W.; Birch, N.P. Physiological and pathological roles of tissue plasminogen activator and its inhibitor neuroserpin in the nervous system. Front. Cell. Neurosci. 2015, 9, 396. [Google Scholar] [CrossRef]
- Costa, M.R. Switch of innate to adaptative immune responses in the brain of patients with Alzheimer’s disease correlates with tauopathy progression. NPJ Aging 2024, 10, 19. [Google Scholar] [CrossRef]
- Becerra, S.P.; Sagasti, A.; Spinella, P.; Notario, V. Pigment epithelium-derived factor behaves like a noninhibitory serpin. Neurotrophic activity does not require the serpin reactive loop. J. Biol. Chem. 1995, 270, 25992–25999. [Google Scholar] [CrossRef]
- Ribeiro, V.T.; De Souza, L.C.; Simões E Silva, A.C. Renin-Angiotensin System and Alzheimer’s Disease Pathophysiology: From the Potential Interactions to Therapeutic Perspectives. Protein Pept. Lett. 2020, 27, 484–511. [Google Scholar] [CrossRef]
- Pan, Y.; Wallace, T.C.; Karosas, T.; Bennett, D.A.; Agarwal, P.; Chung, M. Association of Egg Intake With Alzheimer’s Dementia Risk in Older Adults: The Rush Memory and Aging Project. J. Nutr. 2024, 154, 2236–2243. [Google Scholar] [CrossRef]
- Margara-Escudero, H.J.; Zamora-Ros, R.; de Villasante, I.; Crous-Bou, M.; Chirlaque, M.-D.; Amiano, P.; Mar, J.; Barricarte, A.; Ardanaz, E.; Huerta, J.M. Association Between Egg Consumption and Dementia Risk in the EPIC-Spain Dementia Cohort. Front. Nutr. 2022, 9, 827307. [Google Scholar] [CrossRef]
- Igbinigie, P.O.; Chen, R.; Tang, J.; Dregan, A.; Yin, J.; Acharya, D.; Nadim, R.; Chen, A.; Bai, Z.; Amirabdollahian, F. Association between Egg Consumption and Dementia in Chinese Adults. Nutrients 2024, 16, 3340. [Google Scholar] [CrossRef]
- Ylilauri, M.P.T.; Voutilainen, S.; Leonnroos, E.; Mursu, J.; Virtanen, H.E.K.; Koskinen, T.T.; Salonen, J.T.; Tuomainen, T.-P.; Virtanen, J.K. Association of dietary cholesterol and egg intakes with the risk of incident dementia or Alzheimer disease: The Kuopio Ischaemic Heart Disease Risk Factor Study. Am. J. Clin. Nutr. 2017, 105, 476–484. [Google Scholar] [CrossRef]
- Kritz-Silverstein, D.; Bettencourt, R. Egg Consumption and 4-Year Change in Cognitive Function in Older Men andWomen: The Rancho Bernardo Study. Nutrients 2024, 16, 2765. [Google Scholar] [CrossRef]
- Gao, R.; Yang, Z.; Yan, W.; Du, W.; Zhou, Y.; Zhu, F. Protein intake from different sources and cognitive decline over 9 years in community-dwelling older adults. Front. Public Health 2022, 10, 1016016. [Google Scholar] [CrossRef]
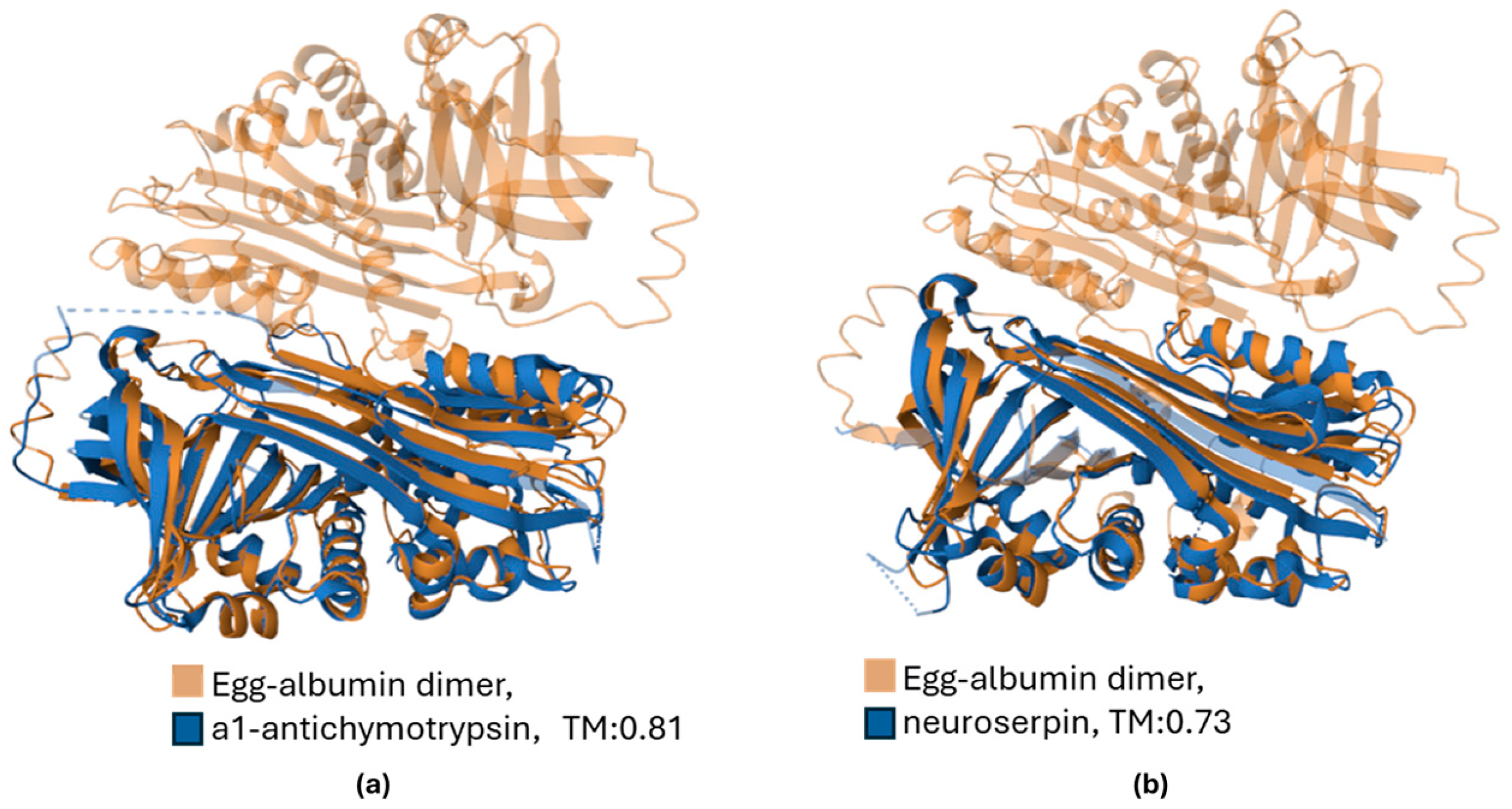
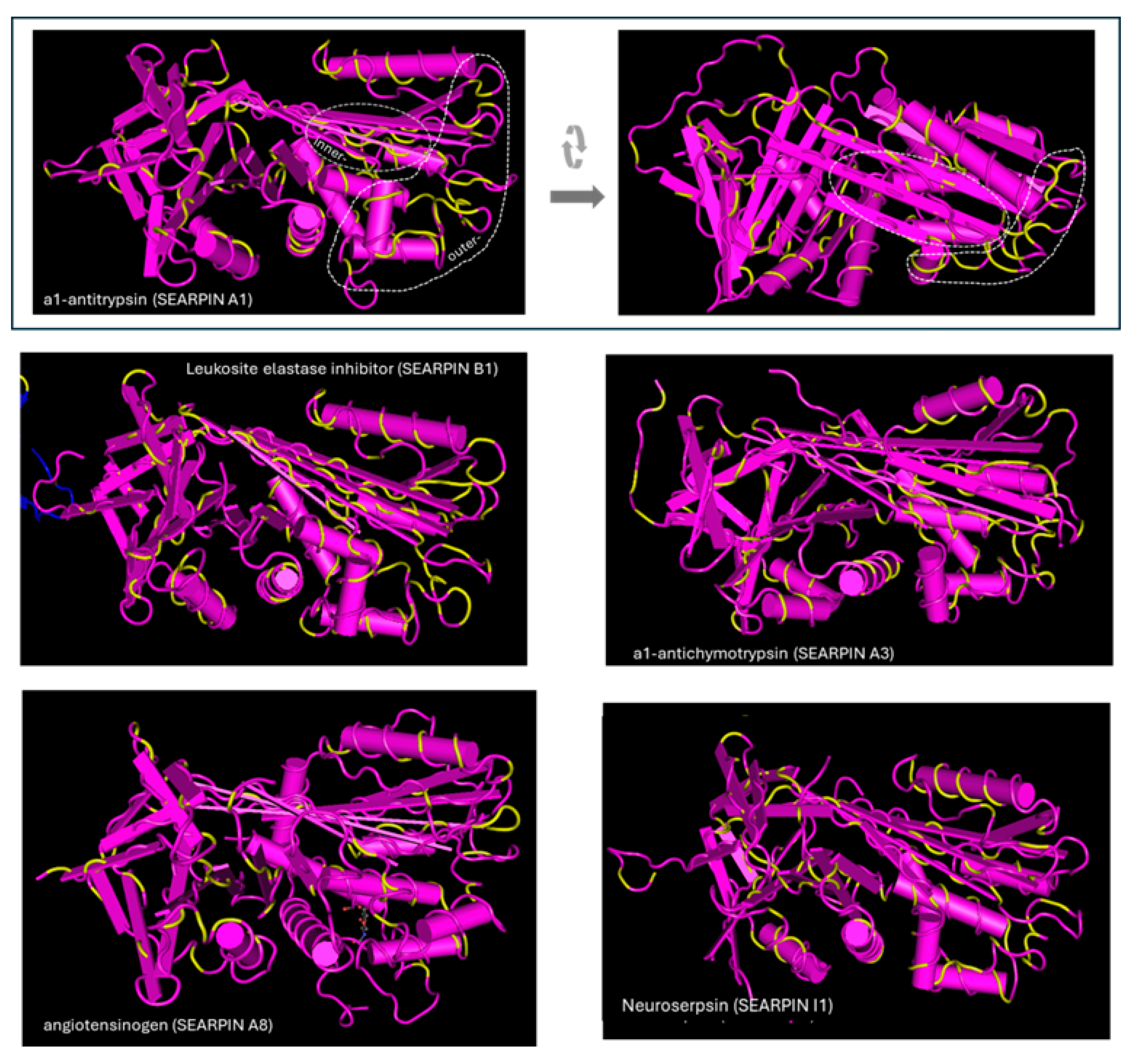

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BLAST Ovalbumin—Homo Sapiens | |||
|---|---|---|---|
| Protein | Description * | Identity | e-Value |
| Serpins | |||
| A1 | Serine protease inhibitor (serpin) A1: α1-antitrypsin, trypsin and chymotrypsin inhibitor, ER and extracellular, anti-coagulant, lung protection | 30.0% | 1.2 × 10−52 |
| A2 | α1-antitrypsin-like (intracellular glycoprotein), inhibition of cathepsin and mast cell chymase (angiotensin I to II converting enzymes) | 28.1% | 1.8 × 10−42 |
| A3 | α1-antichymotrypsin (cell-growth inhibiting gene), intra-/extracellular, inhibition of cathepsin and mast cell chymase (angiotensin I to II converting enzymes), inflammation, lipid metabolism, extracellular matrix remodeling, Alzheimer’s Disease. | 30.0% | 3.6 × 10−52 |
| A4 | Kallistatin (Kallikrein inhibitor, extracellular glycoprotein, expressed in leukocytes, liver, esophagus, and brain) | 27.9% | 5.5 × 10−38 |
| A5 | Serum, urine and seminal serpin, heparin-dependent, anti-coagulant | 29.2% | 5.7 × 10−47 |
| A6 | Extracellular serpin, steroid binding | 26.9% | 2.6 × 10−40 |
| A7 | Serpin, thyroid hormone serum transporter | 25.3% | 4.3 × 10−36 |
| A8 | Angiotensin (AGT). Regulation of blood pressure and electrolyte homeostasis. AGT-2: Vasoconstriction (AGT2), cardiac contractility, and heart rate regulation through its action on the sympathetic nervous system. AGT-3: Aldosterone synthesis and release. AGT 1-7: Vasodilator, antidiuretic effects, antithrombotic through MAS1 (mitochondrial assembly 1)-mediated release of nitric oxide from platelets. Sodium channel regulation, kidney development, vascular remodeling, LDL (Low-Density Lipoprotein) remodeling, associative learning. Involvement in long-term neuronal synaptic plasticity, regulation of transmission of nerve impulse, regulation of norepinephrine secretion, response to estradiol, regulation of protein import into nucleus, nitric oxide-mediated signal transduction, phospholipase C-activating G protein-coupled receptor signaling pathway, RK1 (Extracellular Signal-Regulated Kinase 1) and ERK2 (Extracellular Signal-Regulated Kinase 2) cascade, stress-activated MAPK (mitogen-activated protein kinase) cascade, G protein-coupled receptor signaling Negative regulation of angiogenesis, cell growth, tissue remodeling, gene expression, MAP kinase activity, neurotrophin TRK (Tropomyosin Receptor Kinase) receptor signaling Positive regulation of activation of Janus kinase activity, CoA-transferase activity, NAD(P)H oxidase activity, peptidyl-tyrosine phosphorylation, phosphatidylinositol 3-kinase signaling, protein tyrosine kinase activity, superoxide anion generation, reactive oxygen species metabolic process fibroblast proliferation, endothelial cell migration, positive regulation of vascular-associated smooth muscle cell migration, epidermal growth factor receptor signaling, extrinsic apoptotic signaling pathway, inflammatory response, cytokine production, macrophage derived foam cell differentiation, insulin receptor signaling, membrane hyperpolarization, cytosolic calcium ion concentration, extracellular matrix constituent secretion, L-arginine/L-lysine import across plasma membrane, NF-kappaB (Nuclear factor kappa-light-chain-enhancer of activated B cells) transcription factor activity, nitric oxide biosynthetic process | 21.0% | 4.8 × 10−9 |
| A9 | Extracellular | 29.5% | 2.9 × 10−47 |
| A10 | Serine protease inhibitor, endoplasmic reticulum and extracellular, anticoagulant, heparin-activated, liver regeneration | 25.7% | 2.7 × 10−37 |
| A11 | Serine Protease Inhibitor, extracellular | 27.7% | 1.8 × 10−36 |
| B1 | Extra-/intracellular, neutrophil serine protease inhibitor, cell protection during inflammation, regulates activity of elastase, cathepsin G, proteinase 3, chymase, chymotrypsin, kallikrein 3, caspase 1,4,5, Granzyme H (GZMH), promotes the proliferation of beta cells | 38.8% | 3.9 × 10−87 |
| B2 | Negative regulation apoptosis, fibrinolysis, extra-/intracellular, plasma membrane | 36.9% | 8.8 × 10−83 |
| B3 | SCCA-1 (Squamous Cell Carcinoma Antigen 1), intra-/extracellular, Serine/Cysteine Protease Inhibitor, autocrine/paracrine signal, positive regulation of cell migration and proliferation, anti-apoptotic, modulate the host immune response against tumor cells | 41.4% | 3.8 × 10−104 |
| B4 | SCCA-2, modulate the host immune response against tumor cells. Intra-/extracellular | 40.9% | 1.3 × 10−99 |
| B5 | Tumor suppressor | 30.4% | 6.8 × 10−68 |
| B6 | Probable involvement in the regulation of serine proteinases present in the brain or extravasated from the blood. Inhibitor of cathepsin G, kallikrein-8, and thrombin. Related to hearing loss. | 35.5% | 2.0 × 10−83 |
| B7 | Might function as an inhibitor of Lys-specific proteases. Might influence the maturation of megakaryocytes via its action as a serpin. Intra-/extracellular, mesangial cells, epidermis, positive regulation of collagen biosynthesis, platelet-derived growth factor, transforming growth factor β1, mesangial cell proliferation. | 34.7% | 8.0 × 10−81 |
| B8 | Important role in epithelial desmosome-mediated cell–cell adhesion. Intra-/extracellular. Cytoplasmic antiproteinase 2. | 33.7% | 2.9 × 10−76 |
| B10 | May play a role in the regulation of protease activities during hematopoiesis and apoptosis induced by TNF. May regulate protease activities in the cytoplasm and in the nucleus. | 40.3% | 8.9 × 10−99 |
| B11 | Probable loss of serine protease inhibitory action due to mutation. | 38.8% | 2.4 × 10−100 |
| B12 | Inhibits trypsin and plasmin, but not thrombin, coagulation factor Xa, or urokinase-type plasminogen activator. May play a role in cell differentiation. Intra-/ extracellular. | 37.3% | 1.3 × 10−88 |
| B13 | May play a role in the proliferation or differentiation of keratinocytes. Serine and cysteine protease inhibitor. Intra-/extracellular. | 39.2% | 2.8 × 10−90 |
| C1 | Antithrombin III. Most important serine protease inhibitor in plasma that regulates the blood coagulation cascade. Inhibits thrombin, IXa, Xa, Xia | 32.5% | 9.7 × 10−57 |
| D1 | Thrombin inhibitor activated by the glycosaminoglycans, heparin, or dermatan sulfate. In the presence of the latter, HC-II becomes the predominant thrombin inhibitor in place of antithrombin III (AT-III). Also inhibits chymotrypsin, but in a glycosaminoglycan-independent manner. Peptides at the N-terminal of HC-II have chemotactic activity for both monocytes and neutrophils | 29.6% | 1.8 × 10−49 |
| E1 | PAI-1, inhibitor of tissue-type plasminogen activator (PLAT) and urokinase-type plasminogen activator (PLAU). Cell migration/adhesion. Role in lugs, keratinocyte migration, odontogenesis. | 27.5% | 1.2 × 10−43 |
| E2 | Glia-derived nexin | 30.6% | 1.4 × 10−46 |
| E3 | Extracellular serine protease inhibitor | 25.6% | 2.1 × 10−31 |
| F1 | Pigment epithelium-derived factor. Neurotrophic protein; induces extensive neuronal differentiation in retinoblastoma cells. Potent inhibitor of angiogenesis. Probably no serine protease inhibitory action. Intra-/extracellular. Aging, short-term memory, cellular response to glycose, retinoic acid, dexamethasone, cobalt ion, arsenic containing substances, acidic pH, peptide. Positive regulation of neurogenesis, neuron projection development. Negative regulation of neuron death and inflammation, ovulation, prostate gland, and kidney development. | 22.6% | 6.8 × 10−24 |
| F2 | a2-antiplasmin, extracellular | 26.0% | 1.4 × 10−31 |
| G1 | Plasma protease C1 inhibitor. Activation of the C1 complex. Regulates complement activation, blood coagulation, fibrinolysis, and the generation of kinins. Inhibits FXIIa, chymotrypsin, and kallikrein. Role in innate immune response and aging. | 24.6% | 2.1 × 10−21 |
| H1 | Collagen biosynthesis, protein maturation. | 26.7% | 1.2 × 10−41 |
| I1 | Neuroserpin. Serine protease inhibitor that inhibits plasminogen activators and plasmin but not thrombin. May be involved in the formation or reorganization of synaptic connections as well as in synaptic plasticity in the adult nervous system. May protect neurons from cell damage by tissue-type plasminogen activator (probable) | 29.2% | 1.5 × 10−61 |
| I2 | Myoepithelium-derived serine protease inhibitor. Pancpin. Cell adhesion. Extracellular. | 32.1% | 8.5 × 10−63 |
| Other proteins | |||
| HMSD | Histocompatibility Minor Serpin Domain Containing (HMSD) protein: serpin-like protein, probable serine protease inhibitor. Activation of immune response. Extracellular. | 25.8% | 1.2 × 10−10 |
| PIP4K2C | Phosphatidylinositol 5-phosphate 4-kinase type-2 gamma. Phosphatidylinositol phosphate biosynthetic process, negative regulation of insulin receptor signaling, positive regulation of autophagosome assembly. | 26.2% | 2.7 |
| RENBP | Renin-binding protein (RENBP): N-acylglucosamine 2-epimerase. Catalyzes the interconversion of N-acetylglucosamine to N-acetylmannosamine. Binds to renin forming a protein complex called high-molecular-weight (HMW) renin and inhibits renin activity. Involved in the N-glycolylneuraminic acid (Neu5Gc) degradation pathway. | 29.7% | 5.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amanatidou, D.; Tsolaki, M.; Fouskas, V.; Gavriilidis, I.; Myriouni, M.; Anastasiou, A.; Papageorgiou, E.; Porfyriadou, D.; Parcharidi, Z.; Papasavva, E.; et al. Increased Concentration of Anti-Egg Albumin Antibodies in Cerebrospinal Fluid and Serum of Patients with Alzheimer’s Disease—Discussion on Human Serpins’ Similarity and Probable Involvement in the Disease Mechanism. Biomolecules 2025, 15, 1085. https://doi.org/10.3390/biom15081085
Amanatidou D, Tsolaki M, Fouskas V, Gavriilidis I, Myriouni M, Anastasiou A, Papageorgiou E, Porfyriadou D, Parcharidi Z, Papasavva E, et al. Increased Concentration of Anti-Egg Albumin Antibodies in Cerebrospinal Fluid and Serum of Patients with Alzheimer’s Disease—Discussion on Human Serpins’ Similarity and Probable Involvement in the Disease Mechanism. Biomolecules. 2025; 15(8):1085. https://doi.org/10.3390/biom15081085
Chicago/Turabian StyleAmanatidou, Dionysia, Magdalini Tsolaki, Vasileios Fouskas, Ioannis Gavriilidis, Maria Myriouni, Anna Anastasiou, Efthimia Papageorgiou, Diona Porfyriadou, Zoi Parcharidi, Eleftheria Papasavva, and et al. 2025. "Increased Concentration of Anti-Egg Albumin Antibodies in Cerebrospinal Fluid and Serum of Patients with Alzheimer’s Disease—Discussion on Human Serpins’ Similarity and Probable Involvement in the Disease Mechanism" Biomolecules 15, no. 8: 1085. https://doi.org/10.3390/biom15081085
APA StyleAmanatidou, D., Tsolaki, M., Fouskas, V., Gavriilidis, I., Myriouni, M., Anastasiou, A., Papageorgiou, E., Porfyriadou, D., Parcharidi, Z., Papasavva, E., Fili, M., & Eleftheriou, P. (2025). Increased Concentration of Anti-Egg Albumin Antibodies in Cerebrospinal Fluid and Serum of Patients with Alzheimer’s Disease—Discussion on Human Serpins’ Similarity and Probable Involvement in the Disease Mechanism. Biomolecules, 15(8), 1085. https://doi.org/10.3390/biom15081085