
Dynamics of Fatty Acid Composition in Lipids and Their Distinct Roles in Cardiometabolic Health


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Role of FAs in Maintaining Membrane Structure and Integrity
2.1. FAs in Lipid Structures of Cell Membranes
2.2. FAs Influence Membrane Fluidity, Curvature, and Permeability
2.3. Impacts of FA-Induced Changes in Membrane Properties on CMD Risks
3. Roles of FAs in Cell Signaling
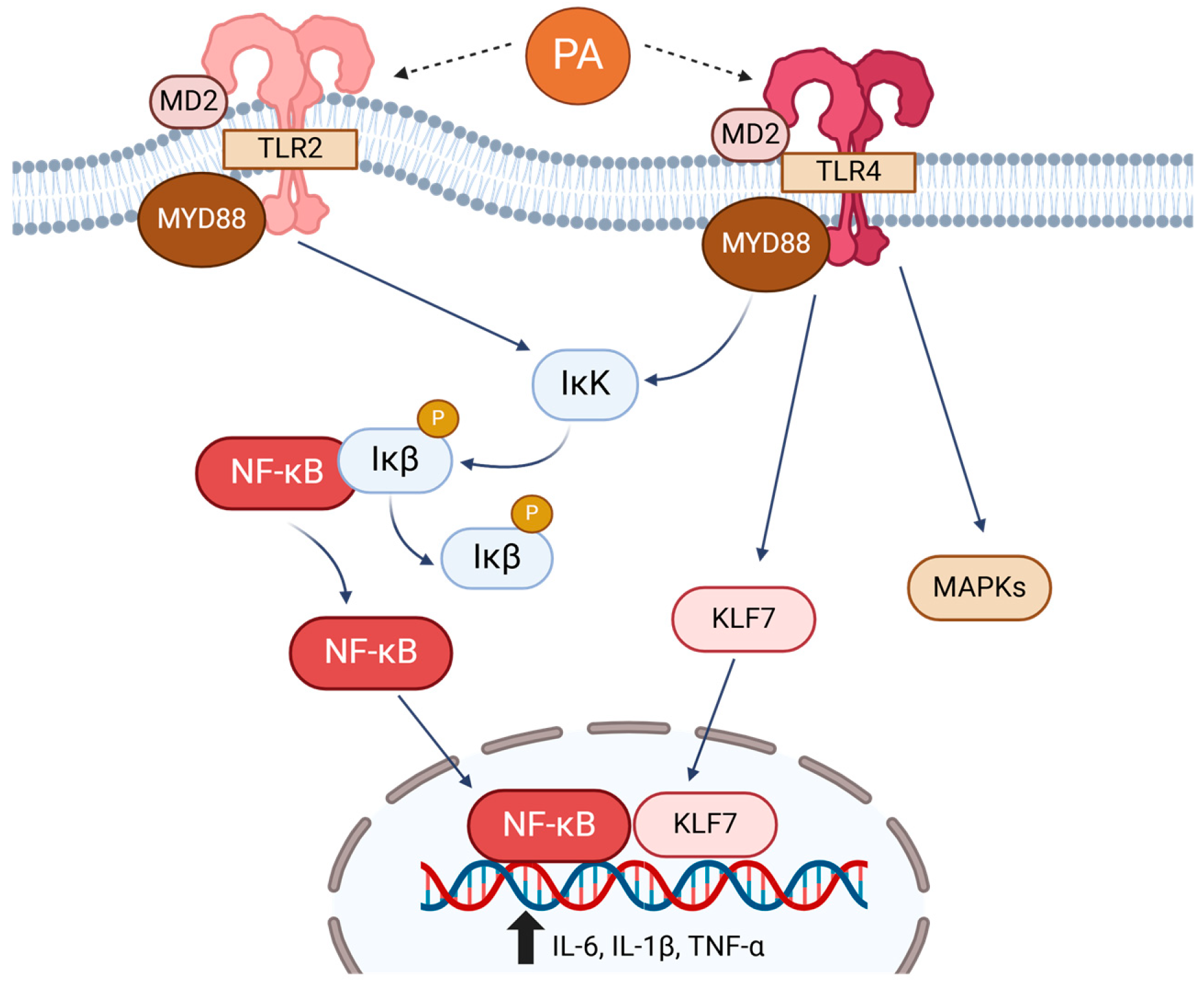
3.1. Toll-like Receptors (TLRs)
3.2. Free Fatty Acid Receptors (FFARs)
3.3. Transient Receptor Potential Channel of the Vanilloid Type (TRPV)
4. FAs as Precursors for Signaling Molecules
4.1. PA Increases Synthesis of Ceramides and DAG
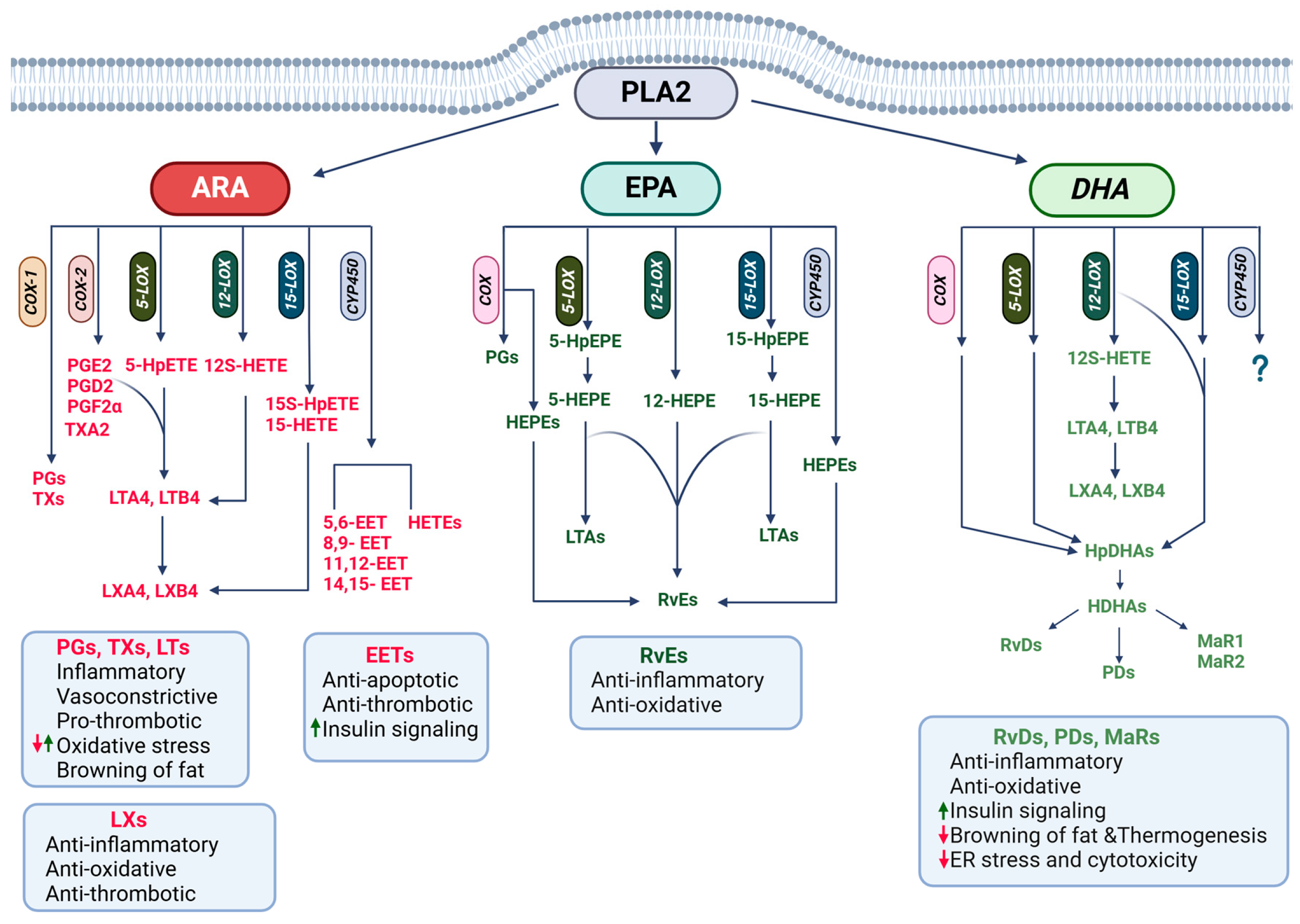
4.2. PUFAs as Precursors for Lipid Mediators
4.2.1. Roles of ARA Derivatives in CMDs
4.2.2. Roles of n-3 PUFA Derivatives in CMDs
4.3. Other Lipid Mediators
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALA | Alpha-linolenic acid |
| AMPK | AMP-activated protein kinase |
| ARA | Arachidonic acid |
| CMDs | Cardiometabolic diseases |
| COX | Cyclooxygenase |
| CVDs | Cardiovascular diseases |
| CYP450 | Cytochrome P450 |
| DAG | Diacylglycerol |
| DHA | Docosahexaenoic acid |
| DNL | De novo lipogenesis |
| DPA | Docosapentaenoic acid |
| EETs | Epoxyeicosatrienoic acids |
| EPA | Eicosapentaenoic acid |
| FAs | Fatty acids |
| FAHFAs | Fatty acid esters of hydroxy fatty acids |
| FFARs | Free fatty acid receptors |
| GPCR | G-protein-coupled receptor |
| GIP | Gastric inhibitory polypeptide |
| GLP-1 | Glucagon-like protein 1 |
| HDHAs | Hydroxydocosahexaenoic acids |
| HEPEs | Hydroxyeicosapentaenoic acids |
| HpDHAs | Hydroxyperoxydocosahexaenoic acids |
| HpETEs | Hydroperoxyeicosatetraenoic acids |
| IL | Interleukin |
| IR | Insulin resistance |
| JNK | c-Jun N-terminal kinase |
| KLF | Kruppel-like factor |
| LA | Linoleic acid |
| LCFAs | Long-chain fatty acids |
| LOX | Lipoxygenase |
| LTs | Leukotrienes |
| LXs | Lipoxins |
| MAPKs | Mitogen-activated protein kinases |
| MaRs | Maresins |
| MASLD | Metabolic dysfunction-associated steatotic liver disease |
| MUFAs | Monounsaturated fatty acids |
| NF-κB | Nuclear factor-kappa B |
| NRF2 | Nuclear factor erythroid 2–related factor 2 |
| OA | Oleic acid |
| PA | Palmitic acid |
| PAHSAs | Palmitic acid-hydroxy stearic acid |
| PC | Phosphatidyl-choline |
| PDs | Protectins |
| PGs | Prostaglandin |
| PKA | Protein kinase A |
| PLs | Phospholipids |
| PLA | Phospholipase A |
| PPARγ | Peroxisome proliferator-activated receptor-gamma |
| PUFAs | Poly-unsaturated fatty acids |
| Rvs | Resolvins |
| ROS | Reactive oxidative species |
| SCFAs | Short-chain fatty acids |
| SFAs | Saturated fatty acids |
| SPMs | Specialized proresolving mediators |
| STAT | Signal transducer and activator of transcription |
| TAG | Triacylglycerol |
| TFAs | Trans fatty acids |
| TLRs | Toll-like receptors |
| TNF-α | Tumor necrosis factor-alpha |
| TRPV | Transient receptor potential, of the vanilloid type |
| TXs | Thromboxanes |
References
- Shi, S.; Huang, H.; Huang, Y.; Zhong, V.W.; Feng, N. Lifestyle Behaviors and Cardiometabolic Diseases by Race and Ethnicity and Social Risk Factors Among US Young Adults, 2011 to 2018. J. Am. Heart Assoc. 2023, 12, e028926. [Google Scholar] [CrossRef]
- Gaggini, M.; Vassalle, C. Lipids Metabolism and Cardiometabolic Diseases. Int. J. Mol. Sci. 2023, 24, 17460. [Google Scholar] [CrossRef]
- Miranda, J.J.; Barrientos-Gutiérrez, T.; Corvalan, C.; Hyder, A.A.; Lazo-Porras, M.; Oni, T.; Wells, J.C.K. Understanding the Rise of Cardiometabolic Diseases in Low- and Middle-Income Countries. Nat. Med. 2019, 25, 1667–1679. [Google Scholar] [CrossRef]
- Domínguez, R.; Pateiro, M.; Purriños, L.; Munekata, P.E.S.; Echegaray, N.; Lorenzo, J.M. In Food Lipids; Lorenzo, J.M., Munekata, P.E.S., Pateiro, M., Barba, F.J., Domínguez, R., Eds.; Chapter 1-Introduction and Classification of Lipids. Academic Press: Cambridge, MA, USA, 2022; pp. 1–16. ISBN 978-0-12-823371-9. [Google Scholar]
- Fahy, E.; Cotter, D.; Sud, M.; Subramaniam, S. Lipid Classification, Structures and Tools. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2011, 1811, 637–647. [Google Scholar] [CrossRef]
- LIPID MAPS. Available online: https://www.lipidmaps.org/databases/lmsd/classification_updates (accessed on 7 February 2025).
- Sztalryd, C.; Brasaemle, D.L. The perilipin family of lipid droplet proteins: Gatekeepers of intracellular lipolysis. Biochim Biophys Acta Mol Cell Biol Lipids 2017, 1862, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane Lipids: Where They Are and How They Behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Ammendolia, D.A.; Bement, W.M.; Brumell, J.H. Plasma Membrane Integrity: Implications for Health and Disease. BMC Biol. 2021, 19, 71. [Google Scholar] [CrossRef]
- Hernandez-Saavedra, D.; Stanford, K.I. In Cardiovascular Signaling in Health and Disease; Parinandi, N.L., Hund, T.J., Eds.; Lipid Mediators in Cardiovascular Physiology and Disease. Springer: Cham, Switzerland, 2022; ISBN 978-3-031-08308-2. [Google Scholar]
- Nunez, E.A. Fatty Acids Involved in Signal Cross-Talk between Cell Membrane and Nucleus. Prostaglandins Leukot. Essent. Fatty Acids 1997, 57, 429–434. [Google Scholar] [CrossRef]
- Coskun, Ü.; Simons, K. Cell Membranes: The Lipid Perspective. Structure 2011, 19, 1543–1548. [Google Scholar] [CrossRef]
- Choi, R.H.; Tatum, S.M.; Symons, J.D.; Summers, S.A.; Holland, W.L. Ceramides and Other Sphingolipids as Drivers of Cardiovascular Disease. Nat. Rev. Cardiol. 2021, 18, 701–711. [Google Scholar] [CrossRef]
- Casares, D.; Escribá, P.V.; Rosselló, C.A. Membrane Lipid Composition: Effect on Membrane and Organelle Structure, Function and Compartmentalization and Therapeutic Avenues. Int. J. Mol. Sci. 2019, 20, 2167. [Google Scholar] [CrossRef]
- Asokapandian, S.; Sreelakshmi, S.; Rajamanickam, G. In Food Biopolymers: Structural, Functional and Nutraceutical Properties; Gani, A., Ashwar, B.A., Eds.; Lipids and Oils: An Overview. Springer International Publishing: Cham, Switzerland, 2021; pp. 389–411. ISBN 978-3-030-27061-2. [Google Scholar]
- De Carvalho, C.C.C.R.; Caramujo, M.J. The Various Roles of Fatty Acids. Molecules 2018, 23, 2583. [Google Scholar] [CrossRef]
- Guimarães, R.C.; Gonçalves, T.T.; Leiria, L.O. Exploiting Oxidized Lipids and the Lipid-Binding GPCRs against Cardiometabolic Diseases. Br. J. Pharmacol. 2021, 178, 531–549. [Google Scholar] [CrossRef]
- Barbosa, A.R.; Pais, S.; Marreiros, A.; Correia, M. Impact of a Mediterranean-Inspired Diet on Cardiovascular Disease Risk Factors: A Randomized Clinical Trial. Nutrients 2024, 16, 2443. [Google Scholar] [CrossRef]
- Wang, D.D.; Nguyen, L.H.; Li, Y.; Yan, Y.; Ma, W.; Rinott, E.; Ivey, K.L.; Shai, I.; Willett, W.C.; Hu, F.B.; et al. The Gut Microbiome Modulates the Protective Association between a Mediterranean Diet and Cardiometabolic Disease Risk. Nat. Med. 2021, 27, 333–343. [Google Scholar] [CrossRef]
- Khalili, L.; Valdes-Ramos, R.; Harbige, L.S. Effect of N-3 (Omega-3) Polyunsaturated Fatty Acid Supplementation on Metabolic and Inflammatory Biomarkers and Body Weight in Patients with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis of RCTs. Metabolites 2021, 11, 742. [Google Scholar] [CrossRef]
- O’Mahoney, L.L.; Matu, J.; Price, O.J.; Birch, K.M.; Ajjan, R.A.; Farrar, D.; Tapp, R.; West, D.J.; Deighton, K.; Campbell, M.D. Omega-3 Polyunsaturated Fatty Acids Favourably Modulate Cardiometabolic Biomarkers in Type 2 Diabetes: A Meta-Analysis and Meta-Regression of Randomized Controlled Trials. Cardiovasc. Diabetol. 2018, 17, 98. [Google Scholar] [CrossRef]
- Hariri, M.; Ghiasvand, R.; Shiranian, A.; Askari, G.; Iraj, B.; Salehi-Abargouei, A. Does Omega-3 Fatty Acids Supplementation Affect Circulating Leptin Levels? A Systematic Review and Meta-Analysis on Randomized Controlled Clinical Trials. Clin. Endocrinol. 2015, 82, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Heshmati, J. Effect of Omega-3 Fatty Acid Supplementation on Gene Expression of Inflammation, Oxidative Stress and Cardiometabolic Parameters: Systematic Review and Meta-Analysis. J. Funct. Foods 2021, 85, 104619. [Google Scholar] [CrossRef]
- Khan, S.U.; Lone, A.N.; Khan, M.S.; Virani, S.S.; Blumenthal, R.S.; Nasir, K.; Miller, M.; Michos, E.D.; Ballantyne, C.M.; Boden, W.E.; et al. Effect of Omega-3 Fatty Acids on Cardiovascular Outcomes: A Systematic Review and Meta-Analysis. EClinicalMedicine 2021, 38, 100997. [Google Scholar] [CrossRef]
- Bae, J.H.; Lim, H.; Lim, S. The Potential Cardiometabolic Effects of Long-Chain ω-3 Polyunsaturated Fatty Acids: Recent Updates and Controversies. Adv. Nutr. 2023, 14, 612–628. [Google Scholar] [CrossRef]
- Moore, E.; Patanwala, I.; Jafari, A.; Davies, I.G.; Kirwan, R.P.; Newson, L.; Mazidi, M.; Lane, K.E. A Systematic Review and Meta-Analysis of Randomized Controlled Trials to Evaluate Plant-Based Omega-3 Polyunsaturated Fatty Acids in Nonalcoholic Fatty Liver Disease Patient Biomarkers and Parameters. Nutr. Rev. 2023, 82, 143–165. [Google Scholar] [CrossRef]
- Zhou, L.; Sun, D.; Bai, H. Efficacy of Fish Oil Supplementation on Metabolic Dysfunction-Associated Steatotic Liver Disease: A Meta-Analysis. Front. Nutr. 2025, 12, 1524830. [Google Scholar] [CrossRef]
- Hashimoto, M.; Hossain, M.S.; Yamasaki, H.; Yazawa, K.; Masumura, S. Effects of Eicosapentaenoic Acid and Docosahexaenoic Acid on Plasma Membrane Fluidity of Aortic Endothelial Cells. Lipids 1999, 34, 1297–1304. [Google Scholar] [CrossRef]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors in Health and Disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef]
- Abbott, S.K.; Else, P.L.; Atkins, T.A.; Hulbert, A.J. Fatty Acid Composition of Membrane Bilayers: Importance of Diet Polyunsaturated Fat Balance. Biochim. Biophys. Acta BBA Biomembr. 2012, 1818, 1309–1317. [Google Scholar] [CrossRef]
- Cucchi, D.; Camacho-Muñoz, D.; Certo, M.; Pucino, V.; Nicolaou, A.; Mauro, C. Fatty Acids—From Energy Substrates to Key Regulators of Cell Survival, Proliferation and Effector Function. Cell Stress 2019, 4, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.M.; Lodhi, I.J. Structural and Functional Roles of Ether Lipids. Protein Cell 2018, 9, 196–206. [Google Scholar] [CrossRef]
- Brown, H.A.; Marnett, L.J. Introduction to Lipid Biochemistry, Metabolism, and Signaling. Chem. Rev. 2011, 111, 5817–5820. [Google Scholar] [CrossRef] [PubMed]
- Maulucci, G.; Cohen, O.; Daniel, B.; Sansone, A.; Petropoulou, P.I.; Filou, S.; Spyridonidis, A.; Pani, G.; De Spirito, M.; Chatgilialoglu, C.; et al. Fatty Acid-Related Modulations of Membrane Fluidity in Cells: Detection and Implications. Free Radic. Res. 2016, 50, S40–S50. [Google Scholar] [CrossRef]
- Hussein, J.S. Cell Membrane Fatty Acids and Health. Int. J. Pharm. Pharm. Sci. 2013, 5, 38–46. [Google Scholar]
- Arouri, A.; Mouritsen, O.G. Membrane-Perturbing Effect of Fatty Acids and Lysolipids. Prog. Lipid Res. 2013, 52, 130–140. [Google Scholar] [CrossRef]
- Jespersen, H.; Andersen, J.H.; Ditzel, H.J.; Mouritsen, O.G. Lipids, Curvature Stress, and the Action of Lipid Prodrugs: Free Fatty Acids and Lysolipid Enhancement of Drug Transport across Liposomal Membranes. Biochimie 2012, 94, 2–10. [Google Scholar] [CrossRef]
- Díaz, M.; Pereda de Pablo, D.; Valdés-Baizabal, C.; Santos, G.; Marin, R. Molecular and Biophysical Features of Hippocampal “Lipid Rafts Aging” Are Modified by Dietary n-3 Long-Chain Polyunsaturated Fatty Acids. Aging Cell 2023, 22, e13867. [Google Scholar] [CrossRef]
- Das, U.N. Essential Fatty Acids—A Review. Curr. Pharm. Biotechnol. 2006, 7, 467–482. [Google Scholar] [CrossRef]
- Baccouch, R.; Shi, Y.; Vernay, E.; Mathelié-Guinlet, M.; Taib-Maamar, N.; Villette, S.; Feuillie, C.; Rascol, E.; Nuss, P.; Lecomte, S.; et al. The Impact of Lipid Polyunsaturation on the Physical and Mechanical Properties of Lipid Membranes. Biochim. Biophys. Acta BBA Biomembr. 2023, 1865, 184084. [Google Scholar] [CrossRef]
- Sherratt, S.C.R.; Libby, P.; Budoff, M.J.; Bhatt, D.L.; Mason, R.P. Role of Omega-3 Fatty Acids in Cardiovascular Disease: The Debate Continues. Curr. Atheroscler. Rep. 2023, 25, 1–17. [Google Scholar] [CrossRef]
- Sherratt, S.C.R.; Juliano, R.A.; Copland, C.; Bhatt, D.L.; Libby, P.; Mason, R.P. EPA and DHA Containing Phospholipids Have Contrasting Effects on Membrane Structure. J. Lipid Res. 2021, 62, 100106. [Google Scholar] [CrossRef] [PubMed]
- Arouri, A.; Lauritsen, K.E.; Nielsen, H.L.; Mouritsen, O.G. Effect of Fatty Acids on the Permeability Barrier of Model and Biological Membranes. Chem. Phys. Lipids 2016, 200, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wiedmann, T.S.; Kandimalla, K.K. Modulating Insulin Signaling and Trafficking at the Blood-Brain Barrier Endothelium Using Lipid Based Nanoemulsions. Int. J. Pharm. 2022, 622, 121823. [Google Scholar] [CrossRef] [PubMed]
- Loehfelm, A.; Elder, M.K.; Boucsein, A.; Jones, P.P.; Williams, J.M.; Tups, A. Docosahexaenoic Acid Prevents Palmitate-induced Insulin-dependent Impairments of Neuronal Health. FASEB J. 2020, 34, 4635–4652. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, H.; Petkevicius, K.; Bartesaghi, S.; Ahnmark, A.; Ruiz, M.; Nilsson, R.; Löfgren, L.; Glover, M.S.; Andréasson, A.-C.; Andersson, L.; et al. Elevated Adipocyte Membrane Phospholipid Saturation Does Not Compromise Insulin Signaling. Diabetes 2022, 72, 1350–1363. [Google Scholar] [CrossRef] [PubMed]
- Pennington, E.R.; Virk, R.; Bridges, M.D.; Bathon, B.E.; Beatty, N.; Gray, R.S.; Kelley, P.; Wassall, S.R.; Manke, J.; Armstrong, M.; et al. Docosahexaenoic Acid Controls Pulmonary Macrophage Lipid Raft Size and Inflammation. J. Nutr. 2024, 154, 1945–1958. [Google Scholar] [CrossRef]
- Hashimoto, M.; Hossain, S.; Shido, O. Docosahexaenoic Acid but Not Eicosapentaenoic Acid Withstands Dietary Cholesterol-Induced Decreases in Platelet Membrane Fluidity. Mol. Cell. Biochem. 2006, 293, 1–8. [Google Scholar] [CrossRef]
- Tzeng, H.-T.; Chyuan, I.-T.; Chen, W.-Y. Shaping of Innate Immune Response by Fatty Acid Metabolite Palmitate. Cells 2019, 8, 1633. [Google Scholar] [CrossRef]
- Serasanambati, M.; Chilakapati, S.R. Function of Nuclear Factor Kappa B (NF-kB) in Human Diseases-A Review. S. Indian J. Biol. Sci. 2016, 2, 368. [Google Scholar] [CrossRef]
- Seliga, A.K.; Zabłocki, K.; Bandorowicz-Pikuła, J. Palmitate Stimulates Expression of the von Willebrand Factor and Modulates Toll-like Receptors Level and Activity in Human Umbilical Vein Endothelial Cells (HUVECs). Int. J. Mol. Sci. 2024, 25, 254. [Google Scholar] [CrossRef]
- Huang, S.; Rutkowsky, J.M.; Snodgrass, R.G.; Ono-Moore, K.D.; Schneider, D.A.; Newman, J.W.; Adams, S.H.; Hwang, D.H. Saturated Fatty Acids Activate TLR-Mediated Proinflammatory Signaling Pathways. J. Lipid Res. 2012, 53, 2002–2013. [Google Scholar] [CrossRef]
- Nicholas, D.A.; Zhang, K.; Hung, C.; Glasgow, S.; Aruni, A.W.; Unternaehrer, J.; Payne, K.J.; Langridge, W.H.R.; Leon, M.D. Palmitic Acid Is a Toll-like Receptor 4 Ligand That Induces Human Dendritic Cell Secretion of IL-1β. PLoS ONE 2017, 12, e0176793. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, Y.; Fang, Q.; Zhong, P.; Li, W.; Wang, L.; Fu, W.; Zhang, Y.; Xu, Z.; Li, X.; et al. Saturated Palmitic Acid Induces Myocardial Inflammatory Injuries through Direct Binding to TLR4 Accessory Protein MD2. Nat. Commun. 2017, 8, 13997. [Google Scholar] [CrossRef]
- Dong, Z.; Zhuang, Q.; Ning, M.; Wu, S.; Lu, L.; Wan, X. Palmitic Acid Stimulates NLRP3 Inflammasome Activation through TLR4-NF-κB Signal Pathway in Hepatic Stellate Cells. Ann. Transl. Med. 2020, 8, 168. [Google Scholar] [CrossRef] [PubMed]
- Snodgrass, R.G.; Huang, S.; Choi, I.-W.; Rutledge, J.C.; Hwang, D.H. Inflammasome-Mediated Secretion of IL-1β in Human Monocytes through TLR2 Activation; Modulation by Dietary Fatty Acids. J. Immunol. 2013, 191, 4337–4347. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, C.; Wu, J.; Ha, X.; Deng, Y.; Zhang, X.; Wang, J.; Chen, K.; Feng, J.; Zhu, J.; et al. The Effect and Mechanism of KLF7 in the TLR4/NF-κB/IL-6 Inflammatory Signal Pathway of Adipocytes. Mediat. Inflamm. 2018, 2018, 1756494. [Google Scholar] [CrossRef]
- Zhang, Y.; Lei, C.-Q.; Hu, Y.-H.; Xia, T.; Li, M.; Zhong, B.; Shu, H.-B. Krüppel-like Factor 6 Is a Co-Activator of NF-κB That Mediates P65-Dependent Transcription of Selected Downstream Genes. J. Biol. Chem. 2014, 289, 12876–12885. [Google Scholar] [CrossRef]
- Okla, M.; Wang, W.; Kang, I.; Pashaj, A.; Carr, T.; Chung, S. Activation of Toll-like Receptor 4 (TLR4) Attenuates Adaptive Thermogenesis via Endoplasmic Reticulum Stress. J. Biol. Chem. 2015, 290, 26476–26490. [Google Scholar] [CrossRef]
- Kim, J.; Jang, H.-J.; Hwang, D.H. Toll-like Receptor 4-Induced Endoplasmic Reticulum Stress Contributes to Impairment of Vasodilator Action of Insulin. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E767–E776. [Google Scholar] [CrossRef]
- Zhang, Y.; Xia, G.; Zhang, Y.; Liu, J.; Liu, X.; Li, W.; Lv, Y.; Wei, S.; Liu, J.; Quan, J. Palmitate Induces VSMC Apoptosis via Toll like Receptor (TLR)4/ROS/P53 Pathway. Atherosclerosis 2017, 263, 74–81. [Google Scholar] [CrossRef]
- Lancaster, G.I.; Langley, K.G.; Berglund, N.A.; Kammoun, H.L.; Reibe, S.; Estevez, E.; Weir, J.; Mellett, N.A.; Pernes, G.; Conway, J.R.W.; et al. Evidence That TLR4 Is Not a Receptor for Saturated Fatty Acids but Mediates Lipid-Induced Inflammation by Reprogramming Macrophage Metabolism. Cell Metab. 2018, 27, 1096–1110.e5. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Ye, J.; Gao, Z.; Youn, H.S.; Lee, W.H.; Zhao, L.; Sizemore, N.; Hwang, D.H. Reciprocal Modulation of Toll-like Receptor-4 Signaling Pathways Involving MyD88 and Phosphatidylinositol 3-Kinase/AKT by Saturated and Polyunsaturated Fatty Acids. J. Biol. Chem. 2003, 278, 37041–37051. [Google Scholar] [CrossRef] [PubMed]
- Nakakuki, M.; Kawano, H.; Notsu, T.; Imada, K. Eicosapentaenoic Acid Suppresses Palmitate-Induced Cytokine Production by Modulating Long-Chain Acyl-CoA Synthetase 1 Expression in Human THP-1 Macrophages. Atherosclerosis 2013, 227, 289–296. [Google Scholar] [CrossRef]
- Hirasawa, A.; Tsumaya, K.; Awaji, T.; Katsuma, S.; Adachi, T.; Yamada, M.; Sugimoto, Y.; Miyazaki, S.; Tsujimoto, G. Free Fatty Acids Regulate Gut Incretin Glucagon-like Peptide-1 Secretion through GPR120. Nat. Med. 2005, 11, 90–94. [Google Scholar] [CrossRef]
- Briscoe, C.P.; Peat, A.J.; McKeown, S.C.; Corbett, D.F.; Goetz, A.S.; Littleton, T.R.; McCoy, D.C.; Kenakin, T.P.; Andrews, J.L.; Ammala, C.; et al. Pharmacological Regulation of Insulin Secretion in MIN6 Cells through the Fatty Acid Receptor GPR40: Identification of Agonist and Antagonist Small Molecules. Br. J. Pharmacol. 2006, 148, 619–628. [Google Scholar] [CrossRef]
- Campbell, A.P.; Smrcka, A.V. Targeting G Protein-Coupled Receptor Signalling by Blocking G Proteins. Nat. Rev. Drug Discov. 2018, 17, 789–803. [Google Scholar] [CrossRef] [PubMed]
- Edfalk, S.; Steneberg, P.; Edlund, H. Gpr40 Is Expressed in Enteroendocrine Cells and Mediates Free Fatty Acid Stimulation of Incretin Secretion. Diabetes 2008, 57, 2280–2287. [Google Scholar] [CrossRef]
- Liou, A.P.; Lu, X.; Sei, Y.; Zhao, X.; Pechhold, S.; Carrero, R.J.; Raybould, H.E.; Wank, S. The G-Protein–Coupled Receptor GPR40 Directly Mediates Long-Chain Fatty Acid–Induced Secretion of Cholecystokinin. Gastroenterology 2011, 140, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Steneberg, P.; Rubins, N.; Bartoov-Shifman, R.; Walker, M.D.; Edlund, H. The FFA Receptor GPR40 Links Hyperinsulinemia, Hepatic Steatosis, and Impaired Glucose Homeostasis in Mouse. Cell Metab. 2005, 1, 245–258. [Google Scholar] [CrossRef]
- Kristinsson, H.; Smith, D.M.; Bergsten, P.; Sargsyan, E. FFAR1 Is Involved in Both the Acute and Chronic Effects of Palmitate on Insulin Secretion. Endocrinology 2013, 154, 4078–4088. [Google Scholar] [CrossRef]
- Lan, H.; Hoos, L.M.; Liu, L.; Tetzloff, G.; Hu, W.; Abbondanzo, S.J.; Vassileva, G.; Gustafson, E.L.; Hedrick, J.A.; Davis, H.R. Lack of FFAR1/GPR40 Does Not Protect Mice From High-Fat Diet–Induced Metabolic Disease. Diabetes 2008, 57, 2999–3006. [Google Scholar] [CrossRef]
- Mo, Z.; Tang, C.; Li, H.; Lei, J.; Zhu, L.; Kou, L.; Li, H.; Luo, S.; Li, C.; Chen, W.; et al. Eicosapentaenoic Acid Prevents Inflammation Induced by Acute Cerebral Infarction through Inhibition of NLRP3 Inflammasome Activation. Life Sci. 2020, 242, 117133. [Google Scholar] [CrossRef] [PubMed]
- Poul, E.L.; Loison, C.; Struyf, S.; Springael, J.-Y.; Lannoy, V.; Decobecq, M.-E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Damme, J.V.; et al. Functional Characterization of Human Receptors for Short Chain Fatty Acids and Their Role in Polymorphonuclear Cell Activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-Chain Fatty Acids Stimulate Glucagon-Like Peptide-1 Secretion via the G-Protein–Coupled Receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Nøhr, M.K.; Pedersen, M.H.; Gille, A.; Egerod, K.L.; Engelstoft, M.S.; Husted, A.S.; Sichlau, R.M.; Grunddal, K.V.; Seier Poulsen, S.; Han, S.; et al. GPR41/FFAR3 and GPR43/FFAR2 as Cosensors for Short-Chain Fatty Acids in Enteroendocrine Cells vs FFAR3 in Enteric Neurons and FFAR2 in Enteric Leukocytes. Endocrinology 2013, 154, 3552–3564. [Google Scholar] [CrossRef]
- Tang, C.; Ahmed, K.; Gille, A.; Lu, S.; Gröne, H.-J.; Tunaru, S.; Offermanns, S. Loss of FFA2 and FFA3 Increases Insulin Secretion and Improves Glucose Tolerance in Type 2 Diabetes. Nat. Med. 2015, 21, 173–177. [Google Scholar] [CrossRef]
- Hong, Y.-H.; Nishimura, Y.; Hishikawa, D.; Tsuzuki, H.; Miyahara, H.; Gotoh, C.; Choi, K.-C.; Feng, D.D.; Chen, C.; Lee, H.-G.; et al. Acetate and Propionate Short Chain Fatty Acids Stimulate Adipogenesis via GPCR43. Endocrinology 2005, 146, 5092–5099. [Google Scholar] [CrossRef]
- Ge, H.; Li, X.; Weiszmann, J.; Wang, P.; Baribault, H.; Chen, J.-L.; Tian, H.; Li, Y. Activation of G Protein-Coupled Receptor 43 in Adipocytes Leads to Inhibition of Lipolysis and Suppression of Plasma Free Fatty Acids. Endocrinology 2008, 149, 4519–4526. [Google Scholar] [CrossRef]
- Xiong, Y.; Miyamoto, N.; Shibata, K.; Valasek, M.A.; Motoike, T.; Kedzierski, R.M.; Yanagisawa, M. Short-Chain Fatty Acids Stimulate Leptin Production in Adipocytes through the G Protein-Coupled Receptor GPR41. Proc. Natl. Acad. Sci. USA 2004, 101, 1045–1050. [Google Scholar] [CrossRef]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 Is an Omega-3 Fatty Acid Receptor Mediating Potent Anti-Inflammatory and Insulin Sensitizing Effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef]
- Li, X.; Ballantyne, L.L.; Che, X.; Mewburn, J.D.; Kang, J.X.; Barkley, R.M.; Murphy, R.C.; Yu, Y.; Funk, C.D. Endogenously Generated Omega-3 Fatty Acids Attenuate Vascular Inflammation and Neointimal Hyperplasia by Interaction With Free Fatty Acid Receptor 4 in Mice. J. Am. Heart Assoc. Cardiovasc. Cerebrovasc. Dis. 2015, 4, e001856. [Google Scholar] [CrossRef]
- Kim, J.; Okla, M.; Erickson, A.; Carr, T.; Natarajan, S.K.; Chung, S. Eicosapentaenoic Acid Potentiates Brown Thermogenesis through FFAR4-Dependent Up-Regulation of miR-30b and miR-378. J. Biol. Chem. 2016, 291, 20551–20562. [Google Scholar] [CrossRef]
- Quesada-López, T.; Cereijo, R.; Turatsinze, J.-V.; Planavila, A.; Cairó, M.; Gavaldà-Navarro, A.; Peyrou, M.; Moure, R.; Iglesias, R.; Giralt, M.; et al. The Lipid Sensor GPR120 Promotes Brown Fat Activation and FGF21 Release from Adipocytes. Nat. Commun. 2016, 7, 13479. [Google Scholar] [CrossRef] [PubMed]
- Gharanei, S.; Shabir, K.; Brown, J.E.; Weickert, M.O.; Barber, T.M.; Kyrou, I.; Randeva, H.S. Regulatory microRNAs in Brown, Brite and White Adipose Tissue. Cells 2020, 9, 2489. [Google Scholar] [CrossRef]
- Ichimura, A.; Hirasawa, A.; Poulain-Godefroy, O.; Bonnefond, A.; Hara, T.; Yengo, L.; Kimura, I.; Leloire, A.; Liu, N.; Iida, K.; et al. Dysfunction of Lipid Sensor GPR120 Leads to Obesity in Both Mouse and Human. Nature 2012, 483, 350–354. [Google Scholar] [CrossRef]
- Bjursell, M.; Xu, X.; Admyre, T.; Böttcher, G.; Lundin, S.; Nilsson, R.; Stone, V.M.; Morgan, N.G.; Lam, Y.Y.; Storlien, L.H.; et al. The Beneficial Effects of N-3 Polyunsaturated Fatty Acids on Diet Induced Obesity and Impaired Glucose Control Do Not Require Gpr120. PLoS ONE 2014, 9, e114942. [Google Scholar] [CrossRef]
- Pærregaard, S.I.; Agerholm, M.; Serup, A.K.; Ma, T.; Kiens, B.; Madsen, L.; Kristiansen, K.; Jensen, B.A.H. FFAR4 (GPR120) Signaling Is Not Required for Anti-Inflammatory and Insulin-Sensitizing Effects of Omega-3 Fatty Acids. Mediators Inflamm. 2016, 2016, 1536047. [Google Scholar] [CrossRef]
- Briscoe, C.P.; Tadayyon, M.; Andrews, J.L.; Benson, W.G.; Chambers, J.K.; Eilert, M.M.; Ellis, C.; Elshourbagy, N.A.; Goetz, A.S.; Minnick, D.T.; et al. The Orphan G Protein-Coupled Receptor GPR40 Is Activated by Medium and Long Chain Fatty Acids. J. Biol. Chem. 2003, 278, 11303–11311. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N.R.; Bannon, A.W.; Surapaneni, S.; Hovland, D.N.; Lehto, S.G.; Gore, A.; Juan, T.; Deng, H.; Han, B.; Klionsky, L.; et al. The Vanilloid Receptor TRPV1 Is Tonically Activated In Vivo and Involved in Body Temperature Regulation. J. Neurosci. 2007, 27, 3366–3374. [Google Scholar] [CrossRef]
- Baskaran, P.; Nazminia, K.; Frantz, J.; O’Neal, J.; Thyagarajan, B. Mice Lacking Endogenous TRPV1 Express Reduced Levels of Thermogenic Proteins and Are Susceptible to Diet-Induced Obesity and Metabolic Dysfunction. FEBS Lett. 2021, 595, 1768–1781. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Uchida, K.; Suzuki, Y.; Zhou, Y.; Kim, M.; Takayama, Y.; Takahashi, N.; Goto, T.; Wakabayashi, S.; Kawada, T.; et al. Lack of TRPV2 Impairs Thermogenesis in Mouse Brown Adipose Tissue. EMBO Rep. 2016, 17, 383–399. [Google Scholar] [CrossRef]
- Ye, L.; Kleiner, S.; Wu, J.; Sah, R.; Gupta, R.K.; Banks, A.S.; Cohen, P.; Khandekar, M.J.; Boström, P.; Mepani, R.; et al. TRPV4 Is a Regulator of Adipose Oxidative Metabolism, Inflammation and Energy Homeostasis. Cell 2012, 151, 96–110. [Google Scholar] [CrossRef]
- Matta, J.A.; Miyares, R.L.; Ahern, G.P. TRPV1 Is a Novel Target for Omega-3 Polyunsaturated Fatty Acids. J. Physiol. 2007, 578, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Goto, T.; Yu, R.; Uchida, K.; Tominaga, M.; Kano, Y.; Takahashi, N.; Kawada, T. Fish Oil Intake Induces UCP1 Upregulation in Brown and White Adipose Tissue via the Sympathetic Nervous System. Sci. Rep. 2015, 5, 18013. [Google Scholar] [CrossRef]
- Kim, M.; Furuzono, T.; Yamakuni, K.; Li, Y.; Kim, Y.I.; Takahashi, H.; Ohue-Kitano, R.; Jheng, H.F.; Takahashi, N.; Kano, Y.; et al. 10-oxo-12(Z)-octadecenoic Acid, a Linoleic Acid Metabolite Produced by Gut Lactic Acid Bacteria, Enhances Energy Metabolism by Activation of TRPV1. FASEB J. 2017, 31, 5036–5048. [Google Scholar] [CrossRef]
- Cao, S.; Anishkin, A.; Zinkevich, N.S.; Nishijima, Y.; Korishettar, A.; Wang, Z.; Fang, J.; Wilcox, D.A.; Zhang, D.X. Transient Receptor Potential Vanilloid 4 (TRPV4) Activation by Arachidonic Acid Requires Protein Kinase A–Mediated Phosphorylation. J. Biol. Chem. 2018, 293, 5307–5322. [Google Scholar] [CrossRef]
- Haus, J.M.; Kashyap, S.R.; Kasumov, T.; Zhang, R.; Kelly, K.R.; DeFronzo, R.A.; Kirwan, J.P. Plasma Ceramides Are Elevated in Obese Subjects with Type 2 Diabetes and Correlate with the Severity of Insulin Resistance. Diabetes 2009, 58, 337–343. [Google Scholar] [CrossRef]
- Boon, J.; Hoy, A.J.; Stark, R.; Brown, R.D.; Meex, R.C.; Henstridge, D.C.; Schenk, S.; Meikle, P.J.; Horowitz, J.F.; Kingwell, B.A.; et al. Ceramides Contained in LDL Are Elevated in Type 2 Diabetes and Promote Inflammation and Skeletal Muscle Insulin Resistance. Diabetes 2013, 62, 401–410. [Google Scholar] [CrossRef]
- Huang, Y.; Sulek, K.; Stinson, S.E.; Holm, L.A.; Kim, M.; Trost, K.; Hooshmand, K.; Lund, M.A.V.; Fonvig, C.E.; Juel, H.B.; et al. Lipid Profiling Identifies Modifiable Signatures of Cardiometabolic Risk in Children and Adolescents with Obesity. Nat. Med. 2025, 31, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Eichelmann, F.; Sellem, L.; Wittenbecher, C.; Jäger, S.; Kuxhaus, O.; Prada, M.; Cuadrat, R.; Jackson, K.G.; Lovegrove, J.A.; Schulze, M.B. Deep Lipidomics in Human Plasma: Cardiometabolic Disease Risk and Effect of Dietary Fat Modulation. Circulation 2022, 146, 21–35. [Google Scholar] [CrossRef]
- Pickersgill, L.; Litherland, G.J.; Greenberg, A.S.; Walker, M.; Yeaman, S.J. Key Role for Ceramides in Mediating Insulin Resistance in Human Muscle Cells. J. Biol. Chem. 2007, 282, 12583–12589. [Google Scholar] [CrossRef] [PubMed]
- Teruel, T.; Hernandez, R.; Lorenzo, M. Ceramide Mediates Insulin Resistance by Tumor Necrosis Factor-α in Brown Adipocytes by Maintaining Akt in an Inactive Dephosphorylated State. Diabetes 2001, 50, 2563–2571. [Google Scholar] [CrossRef] [PubMed]
- Walchuk, C.; Wang, Y.; Suh, M. The Impact of EPA and DHA on Ceramide Lipotoxicity in the Metabolic Syndrome. Br. J. Nutr. 2021, 125, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Chavez, J.A.; Summers, S.A. Characterizing the Effects of Saturated Fatty Acids on Insulin Signaling and Ceramide and Diacylglycerol Accumulation in 3T3-L1 Adipocytes and C2C12 Myotubes. Arch. Biochem. Biophys. 2003, 419, 101–109. [Google Scholar] [CrossRef]
- Abildgaard, J.; Henstridge, D.C.; Pedersen, A.T.; Langley, K.G.; Scheele, C.; Pedersen, B.K.; Lindegaard, B. In Vitro Palmitate Treatment of Myotubes from Postmenopausal Women Leads to Ceramide Accumulation, Inflammation and Affected Insulin Signaling. PLoS ONE 2014, 9, e101555. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, D.; Ma, Y.; Gaspar, R.C.; Kahn, M.; Nasiri, A.; Murray, S.; Samuel, V.T.; Shulman, G.I. Ceramide Synthesis Inhibitors Prevent Lipid-Induced Insulin Resistance through the DAG-PKCε-Insulin receptorT1150 Phosphorylation Pathway. Cell Rep. 2024, 43, 114746. [Google Scholar] [CrossRef]
- Sarnyai, F.; Somogyi, A.; Gór-Nagy, Z.; Zámbó, V.; Szelényi, P.; Mátyási, J.; Simon-Szabó, L.; Kereszturi, É.; Tóth, B.; Csala, M. Effect of Cis- and Trans-Monounsaturated Fatty Acids on Palmitate Toxicity and on Palmitate-Induced Accumulation of Ceramides and Diglycerides. Int. J. Mol. Sci. 2020, 21, 2626. [Google Scholar] [CrossRef]
- Kovacevic, I.; Schmidt, P.H.; Kowalski, A.; Helms, B.J.; van de Lest, C.H.A.; Kluttig, A.; Posern, G. ER Stress Inhibition Enhances Formation of Triacylglcerols and Protects Endothelial Cells from Lipotoxicity. Cell Commun. Signal. CCS 2024, 22, 304. [Google Scholar] [CrossRef]
- Guo, M.; Huang, X.; Zhang, J.; Huang, Y.; Tang, Y.; Wen, H.; Xu, Y.; Zhang, S.; Wei, X.; Sun, S.; et al. Palmitic Acid Induces β-Cell Ferroptosis by Activating Ceramide Signaling Pathway. Exp. Cell Res. 2024, 440, 114134. [Google Scholar] [CrossRef]
- Raj, R.R.; Lofquist, S.; Lee, M.-J. Remodeling of Adipose Tissues by Fatty Acids: Mechanistic Update on Browning and Thermogenesis by n-3 Polyunsaturated Fatty Acids. Pharm. Res. 2023, 40, 467–480. [Google Scholar] [CrossRef]
- Murakami, M. Lipid Mediators in Life Science. Exp. Anim. 2011, 60, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.; Rhee, S.H. Receptor-Mediated Signaling Pathways: Potential Targets of Modulation by Dietary Fatty Acids23. Am. J. Clin. Nutr. 1999, 70, 545–556. [Google Scholar] [CrossRef]
- Crosstalk in the Arachidonic Acid Cascade. Available online: https://www.caymanchem.com/news/crosstalk-in-the-arachidonic-acid-cascade (accessed on 19 February 2025).
- Hu, Y.; Li, W.; Cheng, X.; Yang, H.; She, Z.G.; Cai, J.; Li, H.; Zhang, X.J. Emerging Roles and Therapeutic Applications of Arachidonic Acid Pathways in Cardiometabolic Diseases. Circ. Research 2024, 135, 222–260. [Google Scholar] [CrossRef] [PubMed]
- Harayama, T.; Shimizu, T. Roles of Polyunsaturated Fatty Acids, from Mediators to Membranes. J. Lipid Res. 2020, 61, 1150–1160. [Google Scholar] [CrossRef] [PubMed]
- Regulska, M.; Szuster-Głuszczak, M.; Trojan, E.; Leśkiewicz, M.; Basta-Kaim, A. The Emerging Role of the Double-Edged Impact of Arachidonic Acid-Derived Eicosanoids in the Neuroinflammatory Background of Depression. Curr. Neuropharmacol. 2021, 19, 278–293. [Google Scholar] [CrossRef]
- Khan, M.A.H.; Neckar, J.; Manthati, V.; Errabelli, R.; Pavlov, T.S.; Staruschenko, A.; Falck, J.R.; Imig, J.D. An orally active epoxyeicosatrienoic acid analog attenuates kidney injury in hypertensive dahl salt sensitive rat. Hypertension 2013, 62, A29. [Google Scholar] [CrossRef]
- Deng, Y.; Edin, M.L.; Theken, K.N.; Schuck, R.N.; Flake, G.P.; Kannon, M.A.; DeGraff, L.M.; Lih, F.B.; Foley, J.; Bradbury, J.A.; et al. Endothelial CYP Epoxygenase Overexpression and Soluble Epoxide Hydrolase Disruption Attenuate Acute Vascular Inflammatory Responses in Mice. FASEB J. 2011, 25, 703–713. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.; Schmelzer, K.; Lee, T.S.; Fang, X.; Zhu, Y.; Spector , A.A.; Gill, S.; Morisseau , C.; Hammock, B.D.; et al. The Antiinflammatory Effect of Laminar Flow: The Role of PPARγ, Epoxyeicosatrienoic Acids, and Soluble Epoxide Hydrolase. Proc Natl Acad Sci USA 2005, 105, 16747–16752. [Google Scholar] [CrossRef] [PubMed]
- Perepechaeva, M.L.; Grishanova, A.Y. The Role of Arachidonic Acid Metabolizing Cytochromes P450 in the Control of Cardiovascular Functions. Biochem. Mosc. Suppl. Ser. B Biomed. Chem. 2024, 18, 192–213. [Google Scholar] [CrossRef]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism Pathways of Arachidonic Acids: Mechanisms and Potential Therapeutic Targets. Signal Transduct. Target. Ther. 2021, 6, 1–30. [Google Scholar] [CrossRef]
- Tourki, B.; Kain, V.; Pullen, A.B.; Norris, P.C.; Patel, N.; Arora, P.; Leroy, X.; Serhan, C.N.; Halade, G.V. Lack of Resolution Sensor Drives Age-Related Cardiometabolic and Cardiorenal Defects and Impedes Inflammation-Resolution in Heart Failure. Mol. Metab. 2019, 31, 138–149. [Google Scholar] [CrossRef]
- Sousa, T.; Pinho, D.; Recchiuti, A.; Isopi, E.; Verri, W.A. Editorial: Specialized Proresolving Mediators: Benefits within and beyond Inflammation Resolution in Cardiometabolic, Neurological and Psychiatric Disorders. Front. Physiol. 2023, 14, 1176700. [Google Scholar] [CrossRef]
- Soták, M.; Rajan, M.R.; Clark, M.; Harms, M.; Rani, A.; Kraft, J.D.; Tandio, D.; Shen, T.; Borkowski, K.; Fiehn, O.; et al. Lipoxins Reduce Obesity-Induced Adipose Tissue Inflammation in 3D-Cultured Human Adipocytes and Explant Cultures. iScience 2022, 25, 104602. [Google Scholar] [CrossRef] [PubMed]
- Kollareth, D.J.M.; Leroy, V.; Tu, Z.; Woolet-Stockton, M.J.; Kamat, M.; Garrett, T.J.; Atkinson, C.; Cai, G.; Upchurch, G.R.; Sharma, A.K. Lipoxin A 4/FPR2 Signaling Mitigates Ferroptosis of Alveolar Epithelial Cells via NRF2-Dependent Pathway during Lung Ischemia-Reperfusion Injury. bioRxiv 2024. [Google Scholar] [CrossRef]
- Wan, M.; Godson, C.; Guiry, P.J.; Agerberth, B.; Haeggström, J.Z. Leukotriene B4/Antimicrobial Peptide LL-37 Proinflammatory Circuits Are Mediated by BLT1 and FPR2/ALX and Are Counterregulated by Lipoxin A4 and Resolvin E1. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2011, 25, 1697–1705. [Google Scholar] [CrossRef] [PubMed]
- Madsen, L.; Pedersen, L.M.; Lillefosse, H.H.; Fjære, E.; Bronstad, I.; Hao, Q.; Petersen, R.K.; Hallenborg, P.; Ma, T.; De Matteis, R.; et al. UCP1 Induction during Recruitment of Brown Adipocytes in White Adipose Tissue Is Dependent on Cyclooxygenase Activity. PLoS ONE 2010, 5, e11391. [Google Scholar] [CrossRef]
- Gantert, T.; Henkel, F.; Wurmser, C.; Oeckl, J.; Fischer, L.; Haid, M.; Adamski, J.; Esser-von Bieren, J.; Klingenspor, M.; Fromme, T. Fibroblast Growth Factor Induced Ucp1 Expression in Preadipocytes Requires PGE2 Biosynthesis and Glycolytic Flux. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2021, 35, e21572. [Google Scholar] [CrossRef]
- Ghandour, R.A.; Colson, C.; Giroud, M.; Maurer, S.; Rekima, S.; Ailhaud, G.; Klingenspor, M.; Amri, E.-Z.; Pisani, D.F. Impact of Dietary Ω3 Polyunsaturated Fatty Acid Supplementation on Brown and Brite Adipocyte Function. J. Lipid Res. 2018, 59, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Ghandour, R.A.; Giroud, M.; Vegiopoulos, A.; Herzig, S.; Ailhaud, G.; Amri, E.-Z.; Pisani, D.F. IP-Receptor and PPARs Trigger the Conversion of Human White to Brite Adipocyte Induced by Carbaprostacyclin. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2016, 1861, 285–293. [Google Scholar] [CrossRef]
- Félix-Soriano, E.; Sáinz, N.; Gil-Iturbe, E.; Collantes, M.; Fernández-Galilea, M.; Castilla-Madrigal, R.; Ly, L.; Dalli, J.; Moreno-Aliaga, M.J. Changes in Brown Adipose Tissue Lipid Mediator Signatures with Aging, Obesity, and DHA Supplementation in Female Mice. FASEB J. 2021, 35, e21592. [Google Scholar] [CrossRef]
- Elias, I.; Ferré, T.; Vilà, L.; Muñoz, S.; Casellas, A.; Garcia, M.; Molas, M.; Agudo, J.; Roca, C.; Ruberte, J.; et al. ALOX5AP Overexpression in Adipose Tissue Leads to LXA4 Production and Protection Against Diet-Induced Obesity and Insulin Resistance. Diabetes 2016, 65, 2139–2150. [Google Scholar] [CrossRef]
- Barden, A.; Shinde, S.; Beilin, L.J.; Phillips, M.; Adams, L.; Bollmann, S.; Mori, T.A. Adiposity Associates with Lower Plasma Resolvin E1 (Rve1): A Population Study. Int. J. Obes. 2024, 48, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Molaie, M.; Lotfi, R.; Heidari Moghadam, R.; Rezaiemanesh, A.; Karaji, A.G.; Salari, F. Imbalanced Serum Levels of Resolvin E1 (RvE1) and Leukotriene B4 (LTB4) May Contribute to the Pathogenesis of Atherosclerosis. Prostaglandins Other Lipid Mediat. 2023, 169, 106781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yin, Z.; Xu, Y.; Wei, C.; Peng, S.; Zhao, M.; Liu, J.; Xu, S.; Pan, W.; Zheng, Z.; et al. Resolvin E1/ChemR23 Protects Against Hypertension and Vascular Remodeling in Angiotensin II–Induced Hypertensive Mice. Hypertension 2023, 80, 2650–2664. [Google Scholar] [CrossRef] [PubMed]
- Hasturk, H.; Abdallah, R.; Kantarci, A.; Nguyen, D.; Giordano, N.; Hamilton, J.; Van Dyke, T.E. Resolvin E1 Attenuates Atherosclerotic Plaque Formation in Diet and Inflammation Induced Atherogenesis. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1123–1133. [Google Scholar] [CrossRef]
- Dona, M.; Fredman, G.; Schwab, J.M.; Chiang, N.; Arita, M.; Goodarzi, A.; Cheng, G.; von Andrian, U.H.; Serhan, C.N. Resolvin E1, an EPA-Derived Mediator in Whole Blood, Selectively Counterregulates Leukocytes and Platelets. Blood 2008, 112, 848–855. [Google Scholar] [CrossRef]
- Shamoon, L.; Espitia-Corredor, J.A.; Dongil, P.; Menéndez-Ribes, M.; Romero, A.; Valencia, I.; Díaz-Araya, G.; Sánchez-Ferrer, C.F.; Peiró, C. Resolvin E1 Attenuates Doxorubicin-Induced Endothelial Senescence by Modulating NLRP3 Inflammasome Activation. Biochem. Pharmacol. 2022, 201, 115078. [Google Scholar] [CrossRef]
- Espitia-Corredor, J.A.; Shamoon, L.; Olivares-Silva, F.; Rimassa-Taré, C.; Muñoz-Rodríguez, C.; Espinoza-Pérez, C.; Sánchez-Ferrer, C.F.; Peiró, C.; Díaz-Araya, G. Resolvin E1 Attenuates Doxorubicin-Induced Cardiac Fibroblast Senescence: A Key Role for IL-1β. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2022, 1868, 166525. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, M.; Ding, W.; Zhao, M.; Ye, J.; Xu, Y.; Wang, Z.; Ye, D.; Li, D.; Liu, J.; et al. Resolvin E1 Protects against Doxorubicin-Induced Cardiotoxicity by Inhibiting Oxidative Stress, Autophagy and Apoptosis by Targeting AKT/mTOR Signaling. Biochem. Pharmacol. 2020, 180, 114188. [Google Scholar] [CrossRef]
- Zheng, A.; Huang, N.; Bean, D.; Rayapaneni, S.; Deeney, J.; Sagar, M.; Hamilton, J.A. Resolvin E1 Heals Injured Cardiomyocytes: Therapeutic Implications and H-FABP as a Readout for Cardiovascular Disease & Systemic Inflammation. Prostaglandins Leukot. Essent. Fatty Acids 2023, 197, 102586. [Google Scholar] [CrossRef]
- Arita, M.; Ohira, T.; Sun, Y.-P.; Elangovan, S.; Chiang, N.; Serhan, C.N. Resolvin E1 Selectively Interacts with Leukotriene B4 Receptor BLT1 and ChemR23 to Regulate Inflammation1. J. Immunol. 2007, 178, 3912–3917. [Google Scholar] [CrossRef]
- Unno, Y.; Sato, Y.; Fukuda, H.; Ishimura, K.; Ikeda, H.; Watanabe, M.; Tansho-Nagakawa, S.; Ubagai, T.; Shuto, S.; Ono, Y. Resolvin E1, but Not Resolvins E2 and E3, Promotes fMLF-Induced ROS Generation in Human Neutrophils. FEBS Lett. 2018, 592, 2706–2715. [Google Scholar] [CrossRef]
- Neuhofer, A.; Zeyda, M.; Mascher, D.; Itariu, B.K.; Murano, I.; Leitner, L.; Hochbrugger, E.E.; Fraisl, P.; Cinti, S.; Serhan, C.N.; et al. Impaired Local Production of Proresolving Lipid Mediators in Obesity and 17-HDHA as a Potential Treatment for Obesity-Associated Inflammation. Diabetes 2013, 62, 1945–1956. [Google Scholar] [CrossRef]
- Hellmann, J.; Tang, Y.; Kosuri, M.; Bhatnagar, A.; Spite, M. Resolvin D1 Decreases Adipose Tissue Macrophage Accumulation and Improves Insulin Sensitivity in Obese-Diabetic Mice. FASEB J. 2011, 25, 2399–2407. [Google Scholar] [CrossRef]
- Martínez-Fernández, L.; González-Muniesa, P.; Laiglesia, L.M.; Sáinz, N.; Prieto-Hontoria, P.L.; Escoté, X.; Odriozola, L.; Corrales, F.J.; Arbones-Mainar, J.M.; Martínez, J.A.; et al. Maresin 1 Improves Insulin Sensitivity and Attenuates Adipose Tissue Inflammation in Ob/Ob and Diet-Induced Obese Mice. FASEB J. 2017, 31, 2135–2145. [Google Scholar] [CrossRef]
- Titos, E.; Rius, B.; López-Vicario, C.; Alcaraz-Quiles, J.; García-Alonso, V.; Lopategi, A.; Dalli, J.; Lozano, J.J.; Arroyo, V.; Delgado, S.; et al. Signaling and Immunoresolving Actions of Resolvin D1 in Inflamed Human Visceral Adipose Tissue. J. Immunol. 2016, 197, 3360–3370. [Google Scholar] [CrossRef] [PubMed]
- Weylandt, K.H.; Chiu, C.-Y.; Gomolka, B.; Waechter, S.F.; Wiedenmann, B. Omega-3 Fatty Acids and Their Lipid Mediators: Towards an Understanding of Resolvin and Protectin Formation. Prostaglandins Other Lipid Mediat. 2012, 97, 73–82. [Google Scholar] [CrossRef]
- Olivares-Silva, F.; De Gregorio, N.; Espitia-Corredor, J.; Espinoza, C.; Vivar, R.; Silva, D.; Osorio, J.M.; Lavandero, S.; Peiró, C.; Sánchez-Ferrer, C.; et al. Resolvin-D1 Attenuation of Angiotensin II-Induced Cardiac Inflammation in Mice Is Associated with Prevention of Cardiac Remodeling and Hypertension. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2021, 1867, 166241. [Google Scholar] [CrossRef]
- Díaz del Campo, L.S.; García-Redondo, A.B.; Rodríguez, C.; Zaragoza, C.; Duro-Sánchez, S.; Palmas, F.; de Benito-Bueno, A.; Socuéllamos, P.G.; Peraza, D.A.; Rodrigues-Díez, R.; et al. Resolvin D2 Attenuates Cardiovascular Damage in Angiotensin II-Induced Hypertension. Hypertension 2023, 80, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, J.; Zhao, M.; Liu, J.; Ye, J.; Xu, Y.; Wang, Z.; Ye, D.; Li, D.; Wan, J. Resolvin D1 Attenuates Doxorubicin-Induced Cardiotoxicity by Inhibiting Inflammation, Oxidative and Endoplasmic Reticulum Stress. Front. Pharmacol. 2022, 12, 749899. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Corcuera, A.; Zhu, Y.; Ma, F.; Gupta, N.; Asplund, H.; Yuan, F.; Friedman, S.; Sansbury, B.E.; Huang, X.; Cai, B. Therapeutic Activity of Resolvin D1 (RvD1) in Murine MASH. bioRxiv 2024. [Google Scholar] [CrossRef]
- Dawczynski, C.; Dittrich, M.; Neumann, T.; Goetze, K.; Welzel, A.; Oelzner, P.; Völker, S.; Schaible, A.M.; Troisi, F.; Thomas, L.; et al. Docosahexaenoic Acid in the Treatment of Rheumatoid Arthritis: A Double-Blind, Placebo-Controlled, Randomized Cross-over Study with Microalgae vs. Sunflower Oil. Clin. Nutr. 2018, 37, 494–504. [Google Scholar] [CrossRef]
- Leiria, L.O.; Wang, C.-H.; Lynes, M.D.; Yang, K.; Shamsi, F.; Sato, M.; Sugimoto, S.; Chen, E.Y.; Bussberg, V.; Narain, N.R.; et al. 12-Lipoxygenase Regulates Cold Adaptation and Glucose Metabolism by Producing the Omega-3 Lipid 12-HEPE from Brown Fat. Cell Metab. 2019, 30, 768–783.e7. [Google Scholar] [CrossRef]
- Flachs, P.; Rühl, R.; Hensler, M.; Janovska, P.; Zouhar, P.; Kus, V.; Macek Jilkova, Z.; Papp, E.; Kuda, O.; Svobodova, M.; et al. Synergistic Induction of Lipid Catabolism and Anti-Inflammatory Lipids in White Fat of Dietary Obese Mice in Response to Calorie Restriction and n-3 Fatty Acids. Diabetologia 2011, 54, 2626–2638. [Google Scholar] [CrossRef]
- Lynes, M.D.; Leiria, L.O.; Lundh, M.; Bartelt, A.; Shamsi, F.; Huang, T.L.; Takahashi, H.; Hirshman, M.F.; Schlein, C.; Lee, A.; et al. The Cold-Induced Lipokine 12,13-diHOME Promotes Fatty Acid Transport into Brown Adipose Tissue. Nat. Med. 2017, 23, 631–637. [Google Scholar] [CrossRef]
- Lecka-Czernik, B.; Moerman, E.J.; Grant, D.F.; Lehmann, J.M.; Manolagas, S.C.; Jilka, R.L. Divergent Effects of Selective Peroxisome Proliferator-Activated Receptor-Γ2 Ligands on Adipocyte Versus Osteoblast Differentiation. Endocrinology 2002, 143, 2376–2384. [Google Scholar] [CrossRef]
- Wendel, A.A.; Purushotham, A.; Liu, L.-F.; Belury, M.A. Conjugated Linoleic Acid Induces Uncoupling Protein 1 in White Adipose Tissue of Ob/Ob Mice. Lipids 2009, 44, 975–982. [Google Scholar] [CrossRef]
- Saleh-Ghadimi, S.; Kheirouri, S.; Maleki, V.; Jafari-Vayghan, H.; Alizadeh, M. Endocannabinoid System and Cardiometabolic Risk Factors: A Comprehensive Systematic Review Insight into the Mechanistic Effects of Omega-3 Fatty Acids. Life Sci. 2020, 250, 117556. [Google Scholar] [CrossRef]
- Syed, I.; Lee, J.; Moraes-Vieira, P.M.; Donaldson, C.J.; Sontheimer, A.; Aryal, P.; Wellenstein, K.; Kolar, M.J.; Nelson, A.T.; Siegel, D.; et al. Palmitic Acid Hydroxy Stearic Acids Activate GPR40 Which Is Involved in Their Beneficial Effects on Glucose Homeostasis. Cell Metab. 2018, 27, 419–427.e4. [Google Scholar] [CrossRef]
- Hammarstedt, A.; Syed, I.; Vijayakumar, A.; Eliasson, B.; Gogg, S.; Kahn, B.B.; Smith, U. Adipose Tissue Dysfunction Is Associated with Low Levels of the Novel Palmitic Acid Hydroxystearic Acids. Sci. Rep. 2018, 8, 15757. [Google Scholar] [CrossRef] [PubMed]
- Aryal, P.; Syed, I.; Lee, J.; Patel, R.; Nelson, A.T.; Siegel, D.; Saghatelian, A.; Kahn, B.B. Distinct Biological Activities of Isomers from Several Families of Branched Fatty Acid Esters of Hydroxy Fatty Acids (FAHFAs). J. Lipid Res. 2021, 62, 100108. [Google Scholar] [CrossRef] [PubMed]
- Colas, R.A.; Souza, P.R.; Walker, M.E.; Burton, M.; Zasłona, Z.; Curtis, A.M.; Marques, R.M.; Dalli, J. Impaired Production and Diurnal Regulation of Vascular RvDn-3 DPA Increase Systemic Inflammation and Cardiovascular Disease. Circ. Res. 2018, 122, 855–863. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toncan, F.; Raj, R.R.; Lee, M.-J. Dynamics of Fatty Acid Composition in Lipids and Their Distinct Roles in Cardiometabolic Health. Biomolecules 2025, 15, 696. https://doi.org/10.3390/biom15050696
Toncan F, Raj RR, Lee M-J. Dynamics of Fatty Acid Composition in Lipids and Their Distinct Roles in Cardiometabolic Health. Biomolecules. 2025; 15(5):696. https://doi.org/10.3390/biom15050696
Chicago/Turabian StyleToncan, Fiorenzo, Radha Raman Raj, and Mi-Jeong Lee. 2025. "Dynamics of Fatty Acid Composition in Lipids and Their Distinct Roles in Cardiometabolic Health" Biomolecules 15, no. 5: 696. https://doi.org/10.3390/biom15050696
APA StyleToncan, F., Raj, R. R., & Lee, M.-J. (2025). Dynamics of Fatty Acid Composition in Lipids and Their Distinct Roles in Cardiometabolic Health. Biomolecules, 15(5), 696. https://doi.org/10.3390/biom15050696