

Salt-Induced Membrane-Bound Conformation of the NAC Domain of α-Synuclein Leads to Structural Polymorphism of Amyloid Fibrils


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
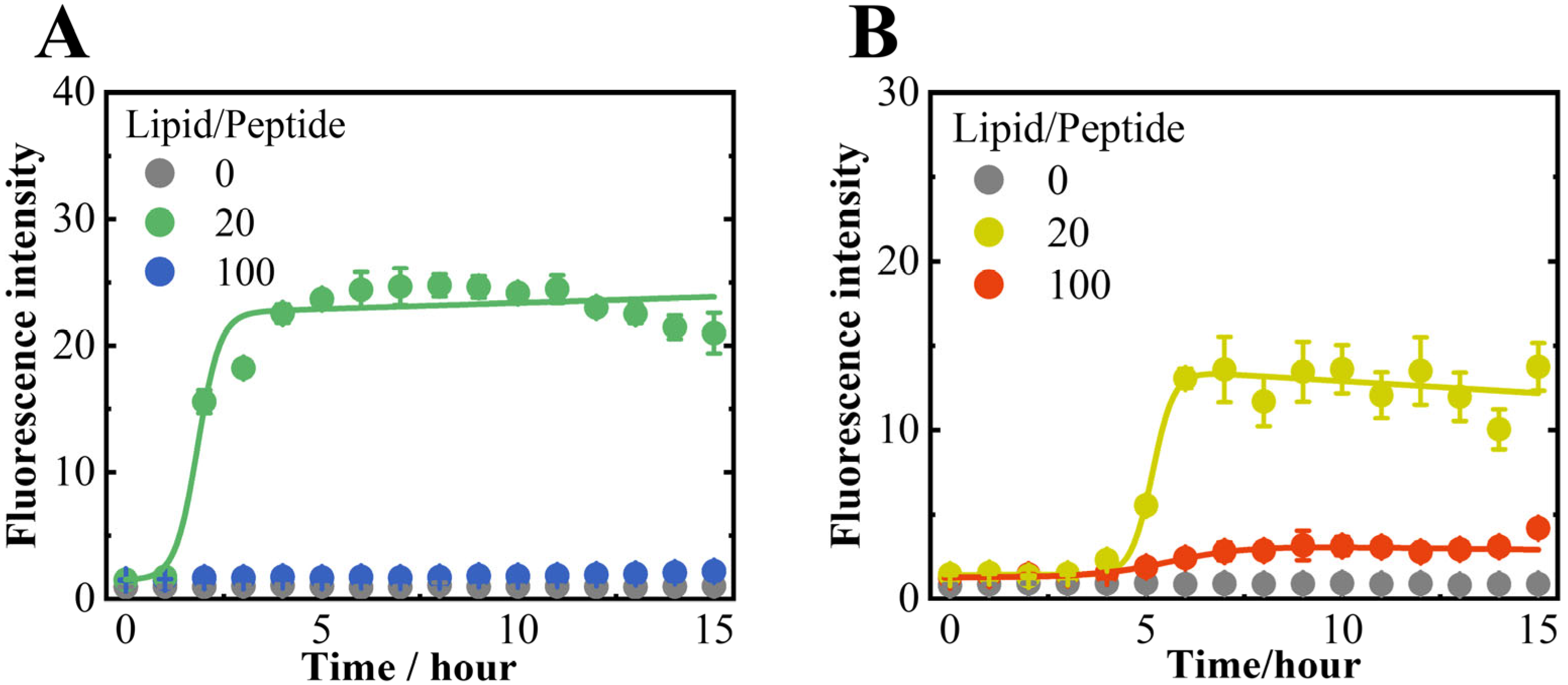
2.3. ThT Fluorescence Assay
2.4. TEM
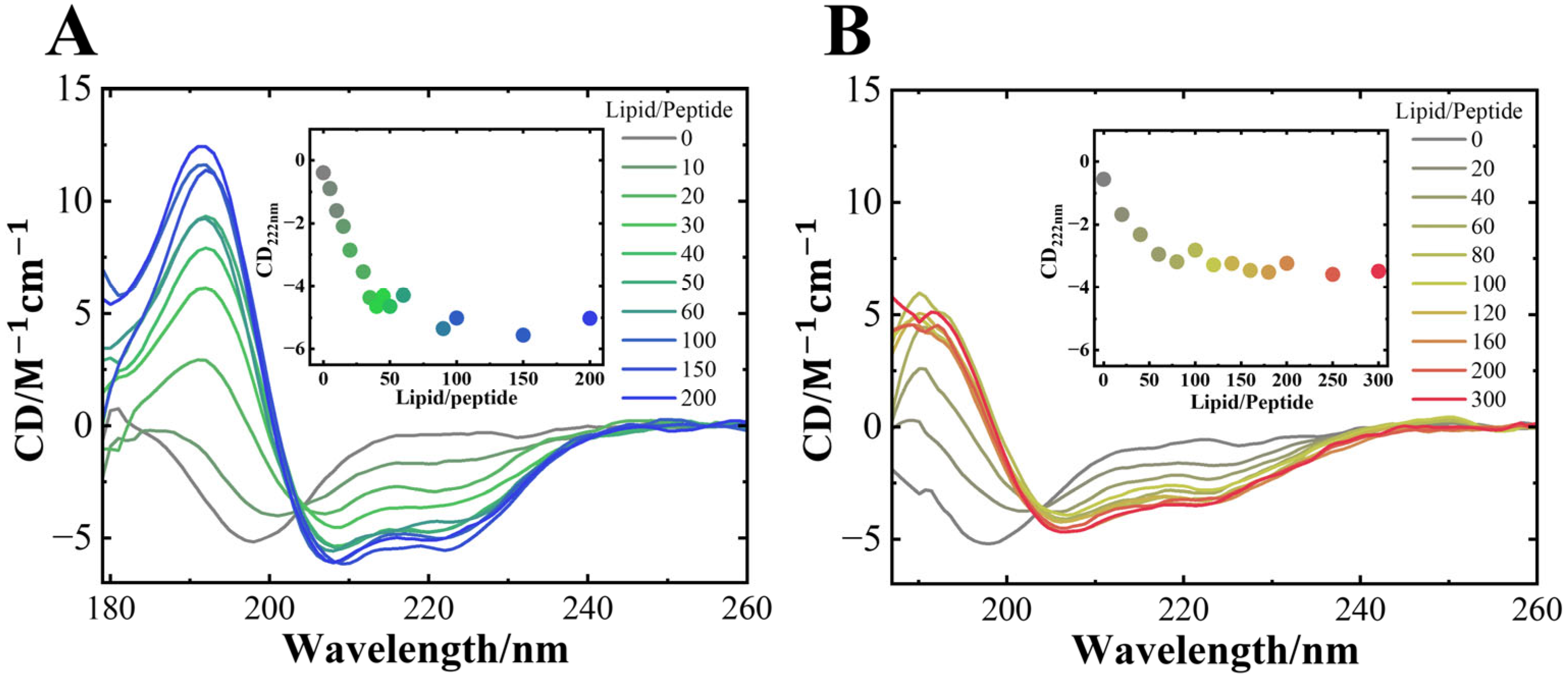
2.5. SRCD Measurement
2.6. Secondary Structure Analysis
2.7. Kinetics Analysis
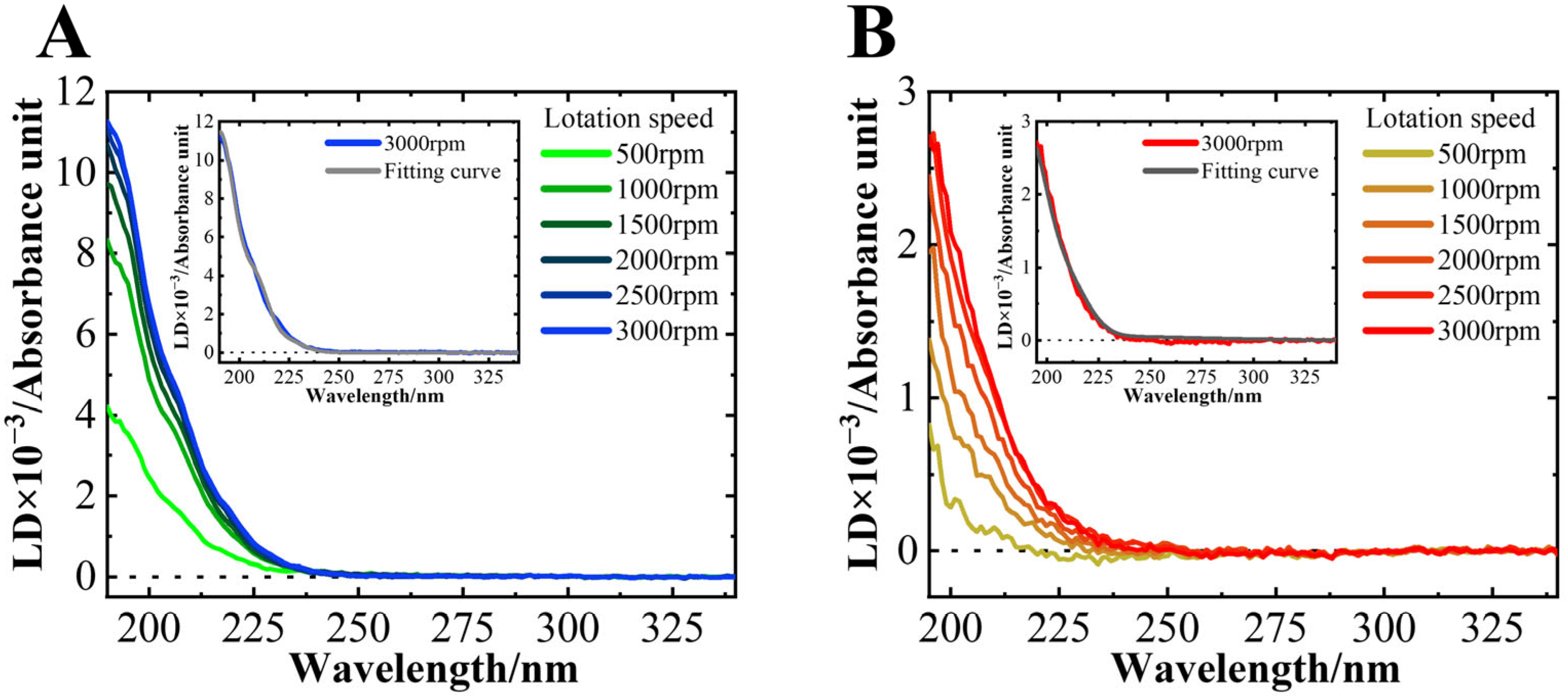
2.8. LD Measurement
2.9. Data Analysis for LD Spectra
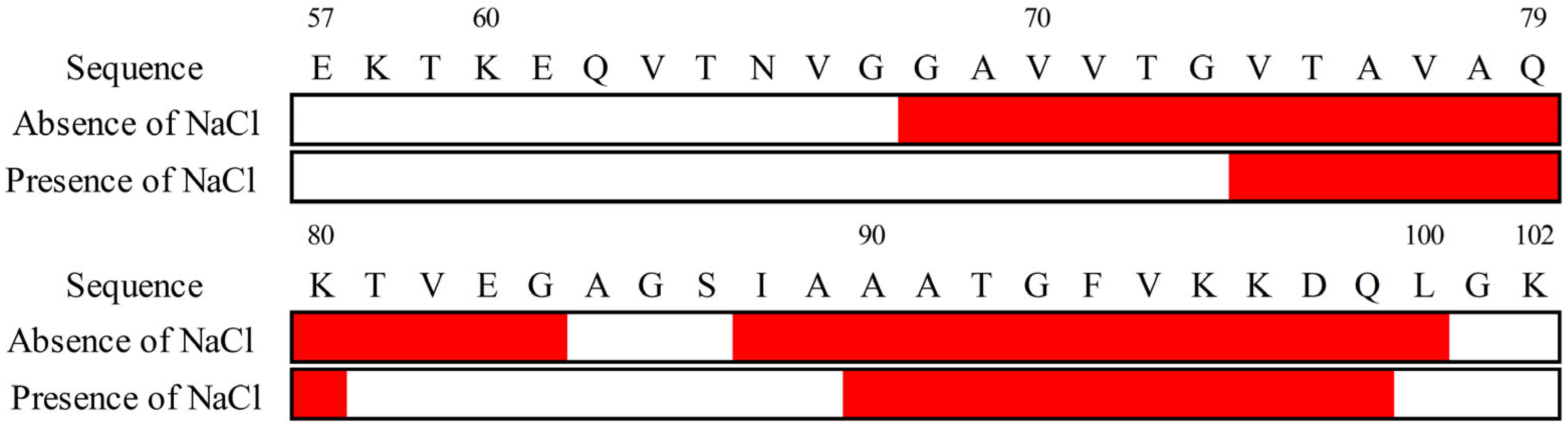
2.10. ATR-FTIR Measurement
2.11. MD Simulation
3. Results and Discussion
3.1. NaCl Induced the Structural Polymorphism in αS57–102 Amyloid Fibrils on Anionic Lipid Membranes
3.2. NaCl Depressed the Amount and Length of Helical Structures of αS57–102 on Membranes
3.3. NaCl Did Not Affect the Average Orientation of the Helical Structure on the Membrane Surface
3.4. NaCl Significantly Exposed the First Helical Regions of αS57–102 to the Solvent
3.5. Structural Polymorphism of αS57–102 Fibrils Occurred Owing to Different Fibrillation Pathways
3.6. Structural Differences in Polymorphisms of αS57–102 Fibrils Were Originated from the Orientations of β-Strands
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, S.; Kim, H.J. Prion-like Mechanism in Amyotrophic Lateral Sclerosis: Are Protein Aggregates the Key? Exp. Neurobiol. 2015, 24, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lansbury, P.T.; Lashuel, H.A. A century-old debate on protein aggregation and neurodegeneration enters the clinic. Nature 2006, 443, 774–779. [Google Scholar] [CrossRef]
- Selkoe, D.J. Folding proteins in fatal ways. Nature 2003, 426, 900–904. [Google Scholar] [CrossRef]
- Uversky, V.N. Neuropathology, biochemistry, and biophysics of α-synuclein aggregation. J. Neurochem. 2007, 103, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Bartels, T.; Ahlstrom, L.S.; Leftin, A.; Kamp, F.; Haass, C.; Brown, M.F.; Beyer, K. The N-Terminus of the Intrinsically Disordered Protein α-Synuclein Triggers Membrane Binding and Helix Folding. Biophys. J. 2010, 99, 2116–2124. [Google Scholar] [CrossRef] [PubMed]
- Lashuel, H.A.; Overk, C.R.; Oueslati, A.; Masliah, E. The many faces of α-synuclein: From structure and toxicity to therapeutic target. Nat. Rev. Neurosci. 2013, 14, 38–48. [Google Scholar] [CrossRef]
- Galvagnion, C.; Brown, J.W.P.; Ouberai, M.M.; Flagmeier, P.; Vendruscolo, M.; Buell, A.K.; Sparr, E.; Dobson, C.M. Chemical properties of lipids strongly affect the kinetics of the membrane-induced aggregation of α-synuclein. Proc. Natl. Acad. Sci. USA 2016, 113, 7065–7070. [Google Scholar] [CrossRef]
- Pfefferkorn, C.M.; Jiang, Z.P.; Lee, J.C. Biophysics of α-synuclein membrane interactions. Biochim. Biophys. Acta Biomembr. 2012, 1818, 162–171. [Google Scholar] [CrossRef]
- O’Leary, E.I.; Lee, J.C. Interplay between α-synuclein amyloid formation and membrane structure. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 483–491. [Google Scholar] [CrossRef]
- Fakhranurova, L.; Marchenkov, V.; Glukhov, A.; Balobanov, V.; Ryabova, N.; Ilyina, N.; Katina, N. Acceleration of carbonic anhydrase amyloid aggregation leads to a decrease in the fibrils toxicity. Biochem. Biophys. Res. Commun. 2024, 741, 151082. [Google Scholar] [CrossRef]
- Madelin, G.; Regatte, R.R. Biomedical applications of sodium MRI in vivo. J. Magn. Reson. Imaging 2013, 38, 511–529. [Google Scholar] [CrossRef]
- Biga, L.M.; Dawson, S.; Harwell, A.; Hopkins, R.; Kaufmann, J.; LeMaster, M.; Matern, P.; Morrison-Graham, K.; Quick, D.; Runyeon, J. Anatomy & Physiology; OpenStax/Oregon State University: Corvallis, OR, USA, 2020. [Google Scholar]
- Bachelard, H.S.; Badargoffer, R.S.; Brooks, K.J.; Dolin, S.J.; Morris, P.G. Measurement of Free Intracellular Calcium in the Brain by 19F-Nuclear Magnetic Resonance Spectroscopy. J. Neurochem. 1988, 51, 1311–1313. [Google Scholar] [CrossRef] [PubMed]
- Imaura, R.; Kawata, Y.; Matsuo, K. Salt-Induced Hydrophobic C-Terminal Region of α-Synuclein Triggers Its Fibrillation under the Mimic Physiologic Condition. Langmuir 2024, 40, 20537–20549. [Google Scholar] [CrossRef] [PubMed]
- Atarod, D.; Mamashli, F.; Ghasemi, A.; Moosavi-Movahedi, F.; Pirhaghi, M.; Nedaei, H.; Muronetz, V.; Haertlé, T.; Tatzelt, J.; Riazi, G.; et al. Bivalent metal ions induce formation of α-synuclein fibril polymorphs with different cytotoxicities. Sci. Rep. 2022, 12, 11898. [Google Scholar] [CrossRef]
- Hoyer, W.; Antony, T.; Cherny, D.; Heim, G.; Jovin, T.M.; Subramaniam, V. Dependence of α-synuclein aggregate morphology on solution conditions. J. Mol. Biol. 2002, 322, 383–393. [Google Scholar] [CrossRef]
- Petkova, A.T.; Leapman, R.D.; Guo, Z.; Yau, W.M.; Mattson, M.P.; Tycko, R. Self-propagating, molecular-level polymorphism in Alzheimer’s beta-amyloid fibrils. Science 2005, 307, 262–265. [Google Scholar] [CrossRef]
- Okuzumi, A.; Hatano, T.; Matsumoto, G.; Nojiri, S.; Ueno, S.I.; Imamichi-Tatano, Y.; Kimura, H.; Kakuta, S.; Kondo, A.; Fukuhara, T.; et al. Propagative α-synuclein seeds as serum biomarkers for synucleinopathies. Nat. Med. 2023, 29, 1448–1455. [Google Scholar] [CrossRef]
- Rawat, A.; Langen, R.; Varkey, J. Membranes as modulators of amyloid protein misfolding and target of toxicity. Biochim. Biophys. Acta Biomembr. 2018, 1860, 1863–1875. [Google Scholar] [CrossRef]
- Ugalde, C.L.; Lawson, V.A.; Finkelstein, D.I.; Hill, A.F. The role of lipids in α-synuclein misfolding and neurotoxicity. J. Biol. Chem. 2019, 294, 9016–9028. [Google Scholar] [CrossRef]
- Mansueto, S.; Fusco, G.; De Simone, A. α-Synuclein and biological membranes: The danger of loving too much. Chem. Commun. 2023, 59, 8769–8778. [Google Scholar] [CrossRef]
- Rodriguez, J.A.; Ivanova, M.I.; Sawaya, M.R.; Cascio, D.; Reyes, F.E.; Shi, D.; Sangwan, S.; Guenther, E.L.; Johnson, L.M.; Zhang, M.; et al. Structure of the toxic core of α-synuclein from invisible crystals. Nature 2015, 525, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Bodles, A.M.; Guthrie, D.J.; Greer, B.; Irvine, G.B. Identification of the region of non-Abeta component (NAC) of Alzheimer’s disease amyloid responsible for its aggregation and toxicity. J. Neurochem. 2001, 78, 384–395. [Google Scholar] [CrossRef]
- El-Agnaf, O.M.A.; Jakes, R.; Curran, M.D.; Middleton, D.; Ingenito, R.; Bianchi, E.; Pessi, A.; Neill, D.; Wallace, A. Aggregates from mutant and wild-type α-synuclein proteins and NAC peptide induce apoptotic cell death in human neuroblastoma cells by formation of β-sheet and amyloid-like filaments. FEBS Lett. 1998, 440, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Guzzo, A.; Delarue, P.; Rojas, A.; Nicolaï, A.; Maisuradze, G.G.; Senet, P. Wild-Type α-Synuclein and Variants Occur in Different Disordered Dimers and Pre-Fibrillar Conformations in Early Stage of Aggregation. Front. Mol. Biosci. 2022, 9, 910104. [Google Scholar] [CrossRef] [PubMed]
- Sot, B.; Rubio-Muñoz, A.; Leal-Quintero, A.; Martínez-Sabando, J.; Marcilla, M.; Roodveldt, C.; Valpuesta, J.M. The chaperonin CCT inhibits assembly of α-synuclein amyloid fibrils by a specific, conformation-dependent interaction. Sci. Rep. 2017, 7, 40859. [Google Scholar] [CrossRef]
- Atsmon-Raz, Y.; Miller, Y. A Proposed Atomic Structure of the Self-Assembly of the Non-Amyloid-β Component of Human α-Synuclein As Derived by Computational Tools. J. Phys. Chem. B 2015, 119, 10005–10015. [Google Scholar] [CrossRef]
- Bisaglia, M.; Trolio, A.; Bellanda, M.; Bergantino, E.; Bubacco, L.; Mammi, S. Structure and topology of the non-amyloid-β component fragment of human α-synuclein bound to micelles: Implications for the aggregation process. Protein. Sci. 2006, 15, 1408–1416. [Google Scholar] [CrossRef]
- Miles, A.J.; Wallace, B.A. Circular dichroism spectroscopy of membrane proteins. Chem. Soc. Rev. 2016, 45, 4859–4872. [Google Scholar] [CrossRef]
- Matsuo, K.; Gekko, K. Circular-Dichroism and Synchrotron-Radiation Circular-Dichroism Spectroscopy as Tools to Monitor Protein Structure in a Lipid Environment. J. Mol. Biol. 2019, 2003, 253–279. [Google Scholar] [CrossRef]
- Matsuo, K.; Kumashiro, M.; Gekko, K. Characterization of the mechanism of interaction between α1-acid glycoprotein and lipid membranes by vacuum-ultraviolet circular-dichroism spectroscopy. Chirality 2020, 32, 594–604. [Google Scholar] [CrossRef]
- Kumashiro, M.; Izumi, Y.; Matsuo, K. Conformation of myelin basic protein bound to phosphatidylinositol membrane characterized by vacuum-ultraviolet circular-dichroism spectroscopy and molecular-dynamics simulations. Proteins Struct. Funct. Bioinf. 2021, 89, 1251–1261. [Google Scholar] [CrossRef]
- Hashimoto, S.; Matsuo, K. Dynamic Observation of the Membrane Interaction Processes of β-Lactoglobulin by Time-Resolved Vacuum-Ultraviolet Circular Dichroism. Anal. Chem. 2024, 96, 10524–10533. [Google Scholar] [CrossRef] [PubMed]
- Kumashiro, M.; Tsuji, R.; Suenaga, S.; Matsuo, K. Formation of β-Strand Oligomers of Antimicrobial Peptide Magainin 2 Contributes to Disruption of Phospholipid Membrane. Membranes 2022, 12, 131. [Google Scholar] [CrossRef] [PubMed]
- Ausili, A.; Sánchez, M.; Gómez-Fernández, J.C. Attenuated total reflectance infrared spectroscopy: A powerful method for the simultaneous study of structure and spatial orientation of lipids and membrane proteins. Biomed. Spectrosc. Imaging 2015, 4, 159–170. [Google Scholar]
- Wallace, B.A. The role of circular dichroism spectroscopy in the era of integrative structural biology. Curr. Opin. Struct. Biol. 2019, 58, 191–196. [Google Scholar] [CrossRef]
- Matsuo, K.; Gekko, K. Construction of a Synchrotron-Radiation Vacuum-Ultraviolet Circular-Dichroism Spectrophotometer and Its Application to the Structural Analysis of Biomolecules. Bull. Chem. Soc. Jpn. 2013, 86, 675–689. [Google Scholar] [CrossRef]
- Szigyártó, I.C.; Deák, R.; Mihály, J.; Rocha, S.; Zsila, F.; Varga, Z.; Beke-Somfai, T. Flow Alignment of Extracellular Vesicles: Structure and Orientation of Membrane-Associated Bio-macromolecules Studied with Polarized Light. Chembiochem 2018, 19, 545–551. [Google Scholar] [CrossRef]
- Bisaglia, M.; Trolio, A.; Tessari, I.; Bubacco, L.; Mammi, S.; Bergantino, E. Cloning, expression, purification, and spectroscopic analysis of the fragment 57-102 of human α-synuclein. Protein Expr. Purif. 2005, 39, 90–96. [Google Scholar] [CrossRef]
- Hope, M.J.; Bally, M.B.; Webb, G.; Cullis, P.R. Production of large unilamellar vesicles by a rapid extrusion procedure. Characterization of size distribution, trapped volume and ability to maintain a membrane potential. Biochim. Biophys. Acta 1985, 812, 55–65. [Google Scholar] [CrossRef]
- Wang, H.; Duo, L.; Hsu, F.; Xue, C.; Lee, Y.K.; Guo, Z. Polymorphic Aβ42 fibrils adopt similar secondary structure but differ in cross-strand side chain stacking interactions within the same β-sheet. Sci. Rep. 2020, 10, 5720. [Google Scholar] [CrossRef]
- Lewis, R.N.; Zhang, Y.P.; McElhaney, R.N. Calorimetric and spectroscopic studies of the phase behavior and organization of lipid bilayer model membranes composed of binary mixtures of dimyristoylphosphatidylcholine and dimyristoylphosphatidylglycerol. Biochim. Biophys. Acta 2005, 1668, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Anthis, N.J.; Clore, G.M. Sequence-specific determination of protein and peptide concentrations by absorbance at 205 nm. Protein Sci. 2013, 22, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, L.; Khurana, R.; Coats, A.; Frokjaer, S.; Brange, J.; Vyas, S.; Uversky, V.N.; Fink, A.L. Effect of environmental factors on the kinetics of insulin fibril formation: Elucidation of the molecular mechanism. Biochemistry 2001, 40, 6036–6046. [Google Scholar] [CrossRef] [PubMed]
- Carija, A.; Pinheiro, F.; Pujols, J.; Brás, I.C.; Lázaro, D.F.; Santambrogio, C.; Grandori, R.; Outeiro, T.F.; Navarro, S.; Ventura, S. Biasing the native α-synuclein conformational ensemble towards compact states abolishes aggregation and neurotoxicity. Redox Biol. 2019, 22, 101135. [Google Scholar] [CrossRef]
- Matsuo, K.; Sakai, K.; Matsushima, Y.; Fukuyama, T.; Gekko, K. Optical cell with a temperature-control unit for a vacuum-ultraviolet circular dichroism spectrophotometer. Anal. Sci. 2003, 19, 129–132. [Google Scholar] [CrossRef]
- Sawada, M.; Namatame, H.; Taniguchi, M. Optical design of a compact and practical UV beamline at HiSOR-BL12. J. Phys. Conf. Ser. 2013, 425, 162010. [Google Scholar] [CrossRef]
- Sreerama, N.; Woody, R.W. Estimation of protein secondary structure from circular dichroism spectra: Comparison of CONTIN, SELCON, and CDSSTR methods with an expanded reference set. Anal. Biochem. 2000, 287, 252–260. [Google Scholar] [CrossRef]
- Matsuo, K.; Watanabe, H.; Gekko, K. Improved sequence-based prediction of protein secondary structures by combining vacuum-ultraviolet circular dichroism spectroscopy with neural network. Proteins Struct. Funct. Bioinf. 2008, 73, 104–112. [Google Scholar] [CrossRef]
- Sreerama, N.; Woody, R.W. Computation and analysis of protein circular dichroism spectra. Methods Enzymol. 2004, 383, 318–351. [Google Scholar] [CrossRef]
- Terakawa, M.S.; Lee, Y.H.; Kinoshita, M.; Lin, Y.X.; Sugiki, T.; Fukui, N.; Ikenoue, T.; Kawata, Y.; Goto, Y. Membrane-induced initial structure of α-synuclein control its amyloidogenesis on model membranes. Biochim. Biophys. Acta Biomembr. 2018, 1860, 757–766. [Google Scholar] [CrossRef]
- Nordén, B.; Rodger, A.; Dafforn, T. Linear Dichroism and Circular Dichroism: A Textbook on Polarized-Light Spectroscopy; RSC Publishing: Cambridge, UK, 2019. [Google Scholar]
- Dafforn, T.R.; Rodger, A. Linear dichroism of biomolecules: Which way is up? Curr. Opin. Struct. Biol. 2004, 14, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Hicks, M.R.; Damianoglou, A.; Rodger, A.; Dafforn, T.R. Folding and Membrane Insertion of the Pore-Forming Peptide Gramicidin Occur as a Concerted Process. J. Mol. Biol. 2008, 383, 358–366. [Google Scholar] [CrossRef]
- Marsh, D. Quantitation of secondary structure in ATR infrared spectroscopy. Biophys. J. 1999, 77, 2630–2637. [Google Scholar] [CrossRef] [PubMed]
- Marsh, D. Spin-label electron spin resonance and Fourier transform infrared spectroscopy for structural/dynamic measurements on ion channels. Methods Enzymol. 1999, 294, 59–92. [Google Scholar] [CrossRef]
- Lenk, T.J.; Horbett, T.A.; Ratner, B.D.; Chittur, K.K. Infrared spectroscopic studies of time-dependent changes in fibrinogen adsorbed to polyurethanes. Langmuir 1991, 7, 1755–1764. [Google Scholar] [CrossRef]
- Stani, C.; Vaccari, L.; Mitri, E.; Birarda, G. FTIR investigation of the secondary structure of type I collagen: New insight into the amide III band. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2020, 229, 118006. [Google Scholar] [CrossRef]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Qi, Y.F.; Cheng, X.; Lee, J.; Vermaas, J.V.; Pogorelov, T.V.; Tajkhorshid, E.; Park, S.; Klauda, J.B.; Im, W. CHARMM-GUI HMMM Builder for Membrane Simulations with the Highly Mobile Membrane-Mimetic Model. Biophys. J. 2015, 109, 2012–2022. [Google Scholar] [CrossRef]
- Monje-Galvan, V.; Warburton, L.; Klauda, J.B. Setting Up All-Atom Molecular Dynamics Simulations to Study the Interactions of Peripheral Membrane Proteins with Model Lipid Bilayers. Methods Mol. Biol. 2019, 1949, 325–339. [Google Scholar] [CrossRef]
- Mark James, A.; Teemu, M.; Roland, S.; Szilárd, P.; Jeremy, C.S.; Berk, H.; Erik, L. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]
- Huang, J.; Rauscher, S.; Nawrocki, G.; Ran, T.; Feig, M.; de Groot, B.L.; Grubmüller, H.; MacKerell, A.D. CHARMM36m: An improved force field for folded and intrinsically disordered proteins. Nat. Methods 2017, 14, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Bu, B.; Crowe, M.; Li, D.C.; Diao, J.J.; Ji, B.H. Membrane packing defects in synaptic vesicles recruit complexin and synuclein. Phys. Chem. Chem. Phys. 2021, 23, 2117–2125. [Google Scholar] [CrossRef] [PubMed]
- Martens, C.; Shekhar, M.; Lau, A.M.; Tajkhorshid, E.; Politis, A. Integrating hydrogen-deuterium exchange mass spectrometry with molecular dynamics simulations to probe lipid-modulated conformational changes in membrane proteins. Nat. Protoc. 2019, 14, 3183–3204. [Google Scholar] [CrossRef]
- Hoover, W.G. Canonical dynamics: Equilibrium phase-space distributions. Phys. Rev. A 1985, 31, 1695–1697. [Google Scholar] [CrossRef]
- Nose, S. A unified formulation of the constant temperature molecular dynamics methods. J. Chem. Phys. 1984, 81, 511–519. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N⋅log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Hess, B. P-LINCS: A parallel linear constraint solver for molecular simulation. J. Chem. Theory Comput. 2008, 4, 116–122. [Google Scholar] [CrossRef]
- Pedersen, J.S.; Otzen, D.E. Amyloid-a state in many guises: Survival of the fittest fibril fold. Protein Sci. 2008, 17, 2–10. [Google Scholar] [CrossRef]
- Rocha, S.; Kumar, R.; Nordén, B.; Wittung-Stafshede, P. Orientation of α-Synuclein at Negatively Charged Lipid Vesicles: Linear Dichroism Reveals Time-Dependent Changes in Helix Binding Mode. J. Am. Chem. Soc. 2021, 143, 18899–18906. [Google Scholar] [CrossRef]
- Dikiy, I.; Eliezer, D. Folding and misfolding of alpha-synuclein on membranes. Biochim. Biophys. Acta Biomembr. 2012, 1818, 1013–1018. [Google Scholar] [CrossRef]
- Sreerama, N.; Woody, R.W. On the analysis of membrane protein circular dichroism spectra. Protein Sci. 2004, 13, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Yonehara, R.; Gekko, K. Improved estimation of the secondary structures of proteins by vacuum-ultraviolet circular dichroism spectroscopy. J. Biochem. 2005, 138, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Sakurada, Y.; Tate, S.; Namatame, H.; Taniguchi, M.; Gekko, K. Secondary-structure analysis of alcohol-denatured proteins by vacuum-ultraviolet circular dichroism spectroscopy. Proteins 2012, 80, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Munishkina, L.A.; Henriques, J.; Uversky, V.N.; Fink, A.L. Role of protein-water interactions and electrostatics in α-synuclein fibril formation. Biochemistry 2004, 43, 3289–3300. [Google Scholar] [CrossRef]
- Matsuo, K.; Namatame, H.; Taniguchi, M.; Gekko, K. Membrane-induced conformational change of alpha1-acid glycoprotein characterized by vacuum-ultraviolet circular dichroism spectroscopy. Biochemistry 2009, 48, 9103–9111. [Google Scholar] [CrossRef]
- Matsuo, K.; Maki, Y.; Namatame, H.; Taniguchi, M.; Gekko, K. Conformation of membrane-bound proteins revealed by vacuum-ultraviolet circular-dichroism and linear-dichroism spectroscopy. Proteins Struct. Funct. Bioinf. 2016, 84, 349–359. [Google Scholar] [CrossRef]
- Shvadchak, V.V.; Subramaniam, V. A four-amino acid linker between repeats in the α-synuclein sequence is important for fibril formation. Biochemistry 2014, 53, 279–281. [Google Scholar] [CrossRef]
- Martin, I.; Goormaghtigh, E.; Ruysschaert, J.M. Attenuated total reflection IR spectroscopy as a tool to investigate the orientation and tertiary structure changes in fusion proteins. Biochim. Biophys. Acta 2003, 1614, 97–103. [Google Scholar] [CrossRef]
- Poojari, C.; Kukol, A.; Strodel, B. How the amyloid-β peptide and membranes affect each other: An extensive simulation study. Biochim. Biophys. Acta 2013, 1828, 327–339. [Google Scholar] [CrossRef]
- Sode, K.; Ochiai, S.; Kobayashi, N.; Usuzaka, E. Effect of reparation of repeat sequences in the human alpha-synuclein on fibrillation ability. Int. J. Biol. Sci. 2006, 3, 1–7. [Google Scholar] [CrossRef]
- Xia, P.; Cao, Y.; Zhao, Q.; Li, H. Energy gap of conformational transition related with temperature for the NACore of α-synuclein. Phys. Chem. Chem. Phys. 2024, 26, 23062–23072. [Google Scholar] [CrossRef] [PubMed]
- Kiechle, M.; Grozdanov, V.; Danzer, K.M. The Role of Lipids in the Initiation of α-Synuclein Misfolding. Front. Cell Dev. Biol. 2020, 8, 562241. [Google Scholar] [CrossRef]
- Havemeister, F.; Ghaeidamini, M.; Esbjörner, E.K. Monovalent cations have different effects on the assembly kinetics and morphology of α-synuclein amyloid fibrils. Biochem. Biophys. Res. Commun. 2023, 679, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Malinchik, S.B.; Inouye, H.; Szumowski, K.E.; Kirschner, D.A. Structural analysis of Alzheimer’s beta(1-40) amyloid: Protofilament assembly of tubular fibrils. Biophys. J. 1998, 74, 537–545. [Google Scholar] [CrossRef]
- Agopian, A.; Guo, Z. Structural origin of polymorphism of Alzheimer’s amyloid β-fibrils. Biochem. J. 2012, 447, 43–50. [Google Scholar] [CrossRef]
- Nassar, R.; Wong, E.; Gsponer, J.; Lamour, G. Inverse Correlation between Amyloid Stiffness and Size. J. Am. Chem. Soc. 2019, 141, 58–61. [Google Scholar] [CrossRef]

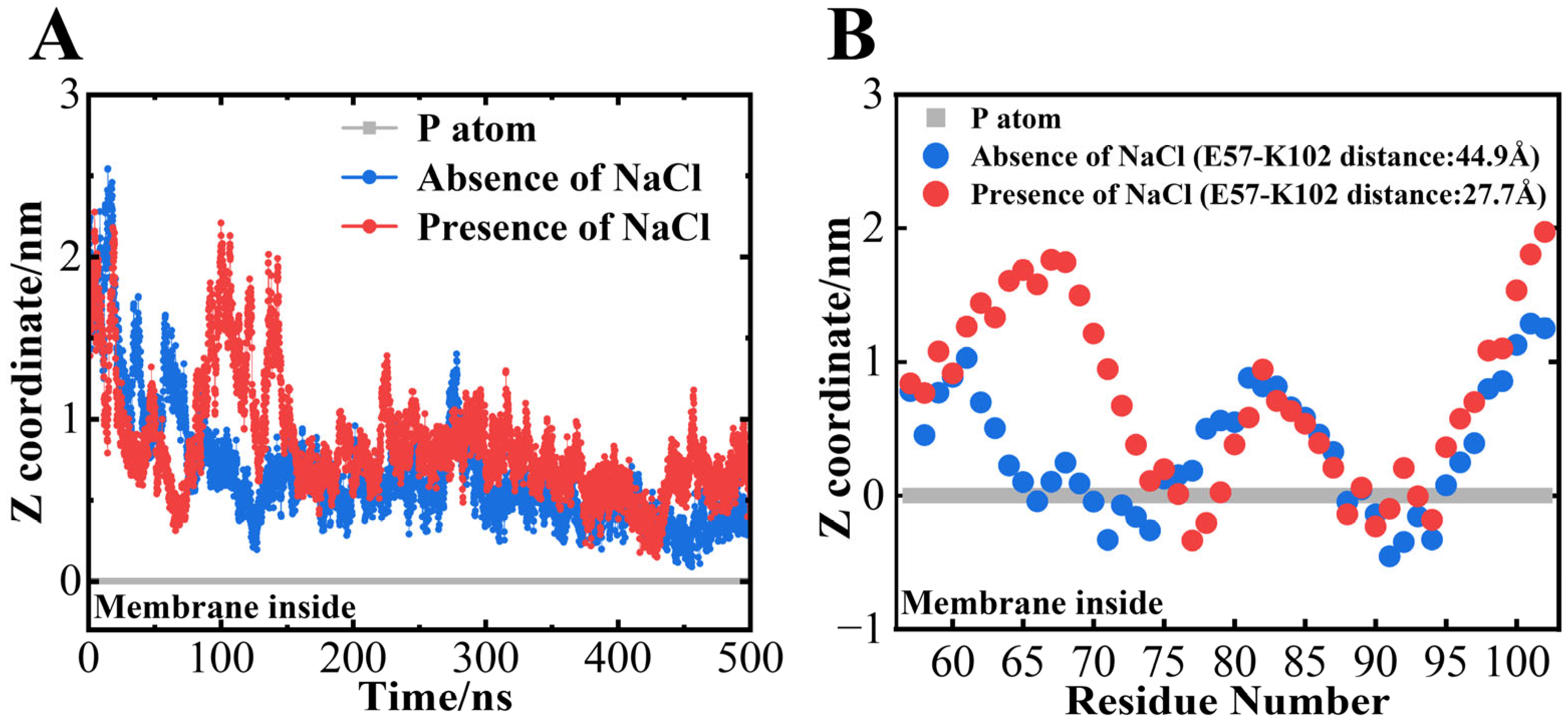
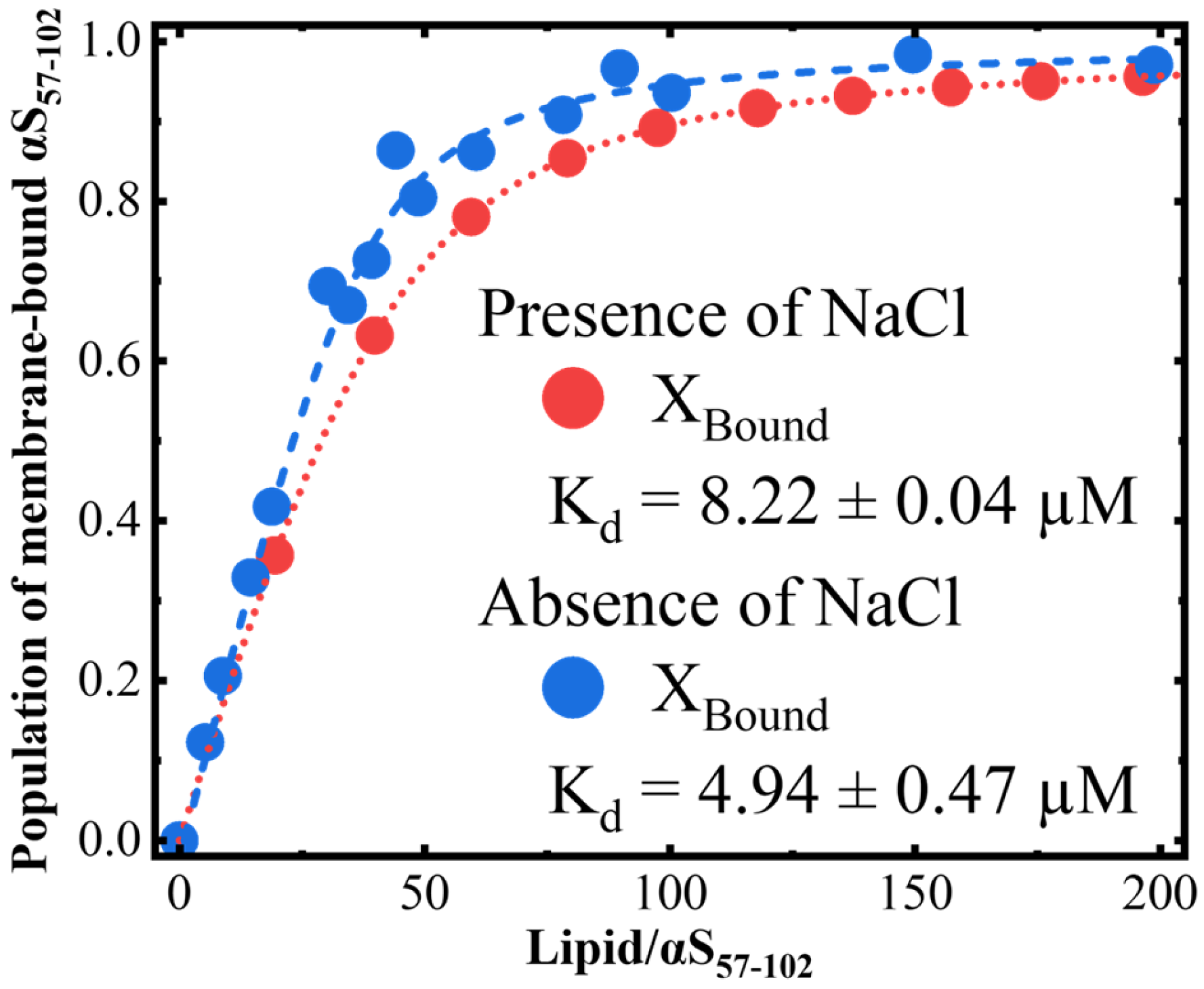

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α-Helix | β-Strand | Turn | Unordered | |||
|---|---|---|---|---|---|---|
| Contents (%) | Numbers | Contents (%) | Numbers | Contents (%) | Contents (%) | |
| Native State (L/P = 0) | ||||||
| Absence of NaCl | 0 | 0 | 36.2 ± 1.7 | 4 | 16.9 ± 0.5 | 46.9 ± 0.7 |
| Presence of NaCl | 2.3 ± 0.5 | 1 | 29.1 ± 2.6 | 3 | 20.0 ± 1.2 | 48.6 ± 1.1 |
| Membrane-bound State (L/P = 200) | ||||||
| Absence of NaCl | 66.4 ± 3.3 | 2 | 1.1 ± 2.5 | 1 | 20.6 ± 1.6 | 11.9 ± 3.0 |
| Presence of NaCl | 37.6 ± 1.5 | 2 | 12.8 ± 1.7 | 2 | 19.2 ± 1.6 | 30.4 ± 1.9 |
| Absence of NaCl | Presence of NaCl | Presence of NaCl | |
|---|---|---|---|
| L/P = 20 | L/P = 20 | L/P = 100 | |
| α-Helix contents (%) | 28.7 ± 3.0 | 23.4 ± 5.9 | 33.8 ± 1.3 |
| β-Sheet contents (%) | 49.9 ± 0.4 | 55.5 ± 0.4 | 38.4 ± 1.0 |
| Angle β (deg) | 21.2 ± 5.3 | 33.2 ± 6.6 | 46.3 ± 0.9 |
| Angle α (deg) | 69.5 ± 2.2 | 67.2 ± 6.5 | 54.9 ± 2.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imaura, R.; Matsuo, K. Salt-Induced Membrane-Bound Conformation of the NAC Domain of α-Synuclein Leads to Structural Polymorphism of Amyloid Fibrils. Biomolecules 2025, 15, 506. https://doi.org/10.3390/biom15040506
Imaura R, Matsuo K. Salt-Induced Membrane-Bound Conformation of the NAC Domain of α-Synuclein Leads to Structural Polymorphism of Amyloid Fibrils. Biomolecules. 2025; 15(4):506. https://doi.org/10.3390/biom15040506
Chicago/Turabian StyleImaura, Ryota, and Koichi Matsuo. 2025. "Salt-Induced Membrane-Bound Conformation of the NAC Domain of α-Synuclein Leads to Structural Polymorphism of Amyloid Fibrils" Biomolecules 15, no. 4: 506. https://doi.org/10.3390/biom15040506
APA StyleImaura, R., & Matsuo, K. (2025). Salt-Induced Membrane-Bound Conformation of the NAC Domain of α-Synuclein Leads to Structural Polymorphism of Amyloid Fibrils. Biomolecules, 15(4), 506. https://doi.org/10.3390/biom15040506