Abstract
Hepatocellular carcinoma (HCC) is the third most common cancer worldwide, widely prevalent across many countries, and poses a serious threat to human health. With changes in its epidemiology, the incidence of HCC is expected to continue rising. As a class of organic molecules widely distributed in nature, quinone compounds possess notable antioxidant, antibacterial, and antitumor properties. This article selects several quinone compounds that have shown notable research progress in recent years and artificially categorizes them into “plant-derived quinone compounds” and “non-plant-derived quinone compounds.” We then provide a detailed review of the research findings regarding HCC in vitro and in vivo experiments and clinical trials, including their potential toxic side effects. Additionally, based on the varying toxicity reduction of several selected plant-derived quinones when combined with doxorubicin, we further hypothesize that these plant-derived quinone compounds may also exert detoxifying effects on other non-plant-derived quinones discussed in this article. In summary, quinone compounds still hold significant research value and development potential in the fight against HCC. At the same time, we hope our review will provide valuable insights and inspiration for future research in this field.
1. Introduction
Hepatocellular carcinoma (HCC) is the third most common cancer worldwide, with an estimated 865,000 new cases diagnosed in 2022 and approximately 757,938 associated deaths [1,2]. As a significant threat to human health, HCC ranks among the top five leading causes of cancer-related mortality in 90 countries worldwide [3]. The epidemiology of HCC is currently undergoing significant shifts. While changes in susceptibility factors have contributed to a decline in hepatitis B and C as predominant etiological drivers, the rising prevalence of non-alcoholic fatty liver disease (NAFLD), also referred to as metabolic-associated fatty liver disease (MAFLD), is expected to drive a continued increase in HCC incidence over the next three decades [4].
Although the incidence of HCC remains concerning, significant advancements have been made in the clinical management of HCC over the past decade. Currently, treatment strategies for HCC primarily follow the Barcelona Clinic Liver Cancer (BCLC) staging system, which allocates different therapeutic approaches based on tumor staging. In principle, early-stage HCC patients are primarily treated with surgical resection, local ablation, or transplantation. For patients with intermediate-stage HCC, transarterial chemoembolization (TACE) is the preferred option, while advanced-stage patients are more likely to opt for systemic therapies. For those with compensated liver disease, surgical resection and systemic treatment are commonly employed to prevent liver function deterioration. Statistically, these treatment modalities have significantly improved the median survival time of various HCC patient groups [5].
Quinones are a class of organic compounds widely distributed in nature, characterized by their unique conjugated cyclic diketone structure, which imparts distinct physicochemical and biological properties. These compounds exhibit various bioactivities, including antioxidant, antibacterial, and antitumor effects. Due to their diverse functional potential, quinones have found promising applications in various fields such as industrial manufacturing, healthcare, and food technology. Consequently, research on quinone compounds has attracted growing interest in recent years [6,7]. However, despite numerous studies demonstrating the potential therapeutic efficacy of various quinone compounds against hepatocellular carcinoma (HCC), a comprehensive review summarizing their anti-HCC effects and mechanisms of action in HCC-related cellular models is still lacking.
This review categorizes representative quinone compounds into non-plant-derived and plant-derived quinones. We evaluate each category’s therapeutic advantages and limitations in treating hepatocellular carcinoma (HCC). We draw from evidence across three key domains: in vivo and in vitro studies, clinical research, and reported adverse effects. In addition, we examine the detoxifying properties of several plant-derived quinones in mitigating doxorubicin-induced toxicity. Finally, we provide a forward-looking perspective on the potential synergistic application of plant- and non-plant-derived quinones in HCC therapy.
2. Quinone Compounds
Quinonoid compounds constitute a broadly distributed class of organic molecules across various organisms, including plants, fungi, bacteria, and animals [8]. These compounds are secondary metabolites derived from the oxidative transformation of phenolic substances. Structurally, they are defined by a core framework featuring at least two carbonyl groups conjugated with adjacent double bonds, typically arranged within an aromatic or cyclic system. In higher plants, their biosynthesis involves multiple metabolic pathways. The structural diversity of these compounds is chiefly influenced by the degree of extension of their conjugated double-bond systems and the periodic arrangement of carbonyl groups. Accordingly, they can be categorized into four subclasses: benzoquinones, naphthoquinones, anthraquinones, and phenanthrenequinones. Existing literature data indicate that the distribution of quinonoid compounds in the plant kingdom exhibits significant family- and genus-specificity. These compounds are predominantly enriched in families such as Rubiaceae, Boehmeriaceae, and Polygonaceae, while secondary accumulation is observed in taxa including Liliaceae, Oleaceae, Meliaceae, Rosaceae, and Loranthaceae [9].
Regarding biological activity, quinonoid compounds exhibit significant antibacterial, antimalarial, and antitumor effects. Current studies suggest that these compounds may inhibit cancer progression and malignancy through multiple mechanisms, including inhibition of glycoproteins, DNA damage, cell cycle arrest induction, and apoptosis promotion [10,11]. Furthermore, quinonoid compounds may exert potent cytotoxicity, directly killing tumor cells. For example, Lee et al. isolated three phenanthrenequinone derivatives (calanquinone A, B, and C), among which calanquinone A demonstrated potent in vitro cytotoxicity against seven human cancer cell lines [12]. For instance, the quinonoid compounds Morindaparvin E and Morindaparvin F have demonstrated potent cytotoxic activity against various cancer cell lines, including HeLa cervical cancer cells and ovarian, renal, and breast cancer cells [13]. Although quinonoid compounds have been extensively investigated for their anticancer properties, the number of reported compounds with confirmed efficacy against primary hepatocellular carcinoma remains remarkably limited.
Based on a comprehensive search of databases including PubMed, Google Scholar, and ScienceDirect, we categorized nine predominantly reported quinonoid compounds with anti-HCC activity into plant-derived and non-plant-derived quinonoid compounds. We focused on highlighting the research progress of several key plant-derived quinonoids—emodin, shikonin, plumbagin (PLB), thymoquinone (TQ), tanshinone IIA (Tan IIA), and tanshinone I (Tan I)—and, among the non-plant-derived quinonoids, doxorubicin (DOX), mitoxantrone (MTX), idarubicin (IDA), and mitomycin (MMC). Figure 1 illustrates these quinonoid compounds’ sources, classifications, and chemical structures.
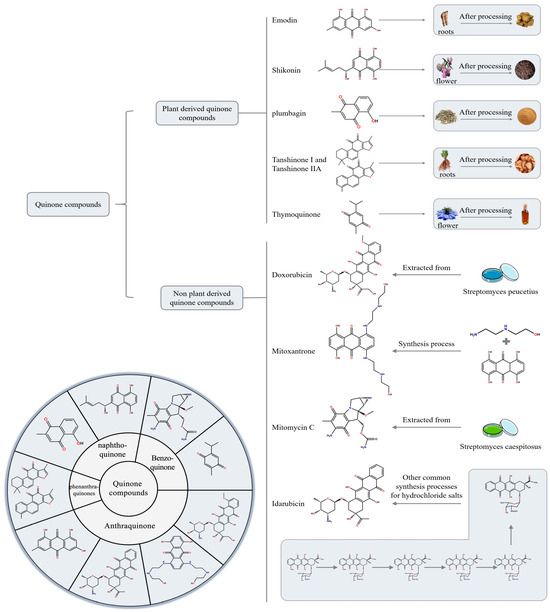
Figure 1.
Classification of quinone compounds and their synthesis routes or source.
3. Research Progress of Plant-Derived Quinonoid Compounds in the Treatment of HCC
3.1. Experimental Studies of Plant-Derived Quinone Compounds in the Field of Anti-HCC
As one of the most lethal cancers worldwide, HCC remains only partially understood in terms of its biological and molecular mechanisms, despite extensive research efforts. The development of novel therapeutic agents for HCC must therefore be grounded in a deeper understanding of its underlying biology and molecular pathways. As the following section will discuss therapeutic strategies of quinone compounds against liver cancer, it will involve several key mechanisms, including apoptosis, oxidative stress, and epithelial–mesenchymal transition (EMT); to facilitate readers’ comprehension, we briefly summarize the major anti-HCC mechanisms of the quinone-based compounds discussed in this paper. Apoptosis, a form of programmed cell death, has long been a focal point in cancer research. Notably, the evasion of apoptosis is a key mechanism underlying drug resistance and disease progression in HCC. Thus, elucidating the molecular mechanisms responsible for dysregulated apoptosis in HCC cells is of critical importance [14]. Oxidative stress is a state of imbalance between pro-oxidant and antioxidant processes in the body, and it plays a distinct role in the development of HCC. During oxidative stress, large amounts of reactive oxygen species (ROS) are produced, acting as a double-edged sword. On the one hand, ROS can promote the proliferation and migration of tumor cells; on the other hand, excessive oxidative stress can lead to tumor cell death [15]. EMT is a multistep biological process that commonly occurs during embryonic development. In the progression of HCC, the harsh microenvironment within the tumor core induces EMT in hepatocytes, enabling them to acquire enhanced resistance to apoptosis and increased migratory capacity. Furthermore, EMT facilitates angiogenesis in HCC cells, allowing them to obtain oxygen and nutrients necessary for tumor growth. Therefore, the development of novel therapeutic agents should be based on a thorough understanding of the biological and molecular mechanisms underlying EMT [16]. Among the various cancer-related biological mechanisms, cuproptosis represents a relatively unique form of cell death. First defined in detail in 2022, cuproptosis has since been reported as a mechanism that can be exploited to induce cell death in lung and colorectal cancer cells. Given its distinct mode of action, the exploration of cuproptosis-related pathways holds great potential for future research and therapeutic development in the field of HCC [17].
Natural products have long served as valuable sources for developing novel therapeutics for various diseases, including cancer. Emodin, a naturally occurring anthraquinone derivative extracted from medicinal plants such as Rheum (rhubarb), Polygonum cuspidatum, and Fallopia multiflora, has demonstrated notable anti-hepatocellular carcinoma (HCC) activity. Qin, Hassan, et al. [18,19] demonstrated that emodin inhibits the proliferation of Hep-G2 cells by inducing cell cycle arrest at the S/G2-M phase and promoting apoptosis. In addition, emodin significantly suppresses these cells’ migratory and invasive capacities, improves the survival rate of HCC-bearing rats, and reduces the number of hepatic nodules. At the molecular level, emodin downregulates the elevated mRNA and protein expression of Protein Kinase C (PKC), Extracellular signal-regulated kinase 5 (ERK5), A Disintegrin And Metalloproteinase With Thrombospondin 4 (ADAMTS4), Matrix Metalloproteinase 3 (MMP3), and Vascular Endothelial Growth Factor (VEGF) in the livers of affected rats. Beyond its intrinsic anti-HCC activity, emodin exhibits promising potential with conventional chemotherapeutic agents for treating HCC. A study conducted by Kim et al. [20] demonstrated that emodin inhibits the transcriptional activity of sterol regulatory element-binding protein 2 (SREBP-2), thereby suppressing cholesterol biosynthesis and downstream signaling of the oncogenic protein kinase B (AKT). Furthermore, when combined with sorafenib, emodin synergistically enhances sorafenib-induced G1 phase cell cycle arrest and apoptosis. In vivo, emodin exhibits superior tumor growth inhibition in animal models xenografted with Hep-G2 or SK-HEP-1 cells. Similarly, cisplatin is a cytotoxic anticancer drug with broad-spectrum efficacy. It exerts its effects by inhibiting DNA replication in cancer cells and disrupting structural components of their cell membranes [21,22]. However, its clinical application is limited due to the development of drug resistance and inherent toxicity [23]. YANG et al. [24] investigated the combined application of cisplatin and emodin and found that the combination exerted significantly stronger inhibitory effects on cell proliferation than either agent alone. Moreover, the co-treatment more effectively suppressed cell migration in wound healing assays, exhibiting both time- and dose-dependent responses. This enhanced antitumor efficacy is likely attributed to emodin’s ability to inhibit epithelial–mesenchymal transition (EMT), thereby increasing the sensitivity of Hep-G2 cells to cisplatin. In summary, emodin inhibits HCC progression by modulating key signaling pathways, such as PKC/ERK5/MMP3. Combined with agents like sorafenib or cisplatin, it synergistically enhances cell cycle arrest, apoptosis, and chemosensitivity. Nevertheless, further mechanistic studies and comprehensive safety evaluations must support its clinical translation.
Shikonin is a naturally occurring red-colored quinonoid compound extracted from the roots of Lithospermum erythrorhizon (commonly known as purple gromwell). It exhibits a broad spectrum of pharmacological activities, including anti-inflammatory, antioxidant, and anticancer effects [25]. Mitochondria play a critical role in regulating numerous aspects of cellular function, including energy metabolism, apoptosis, and redox balance. Mitochondrial dysfunction has been shown to significantly affect cell growth and proliferation, contributing to the pathogenesis of HCC [26]. Shikonin has been reported to induce apoptosis in HCC cells by increasing reactive oxygen species (ROS) production by inhibiting pyruvate kinase M2 (PKM2) activity. However, the sensitivity to shikonin varies among different HCC cell lines, suggesting cell line-specific mechanisms of action or resistance. Therefore, Yang et al. [27] evaluated the effects of shikonin on oxidative stress and metabolic pathways in sensitive and resistant HCC cell lines. They sought to identify the mechanisms underlying the differences in sensitivity. The study results indicate that Hep-G2 cells are more sensitive to shikonin than HCCLM3 cells. Shikonin induces nuclear expression of PKM2 and HIF1α, activating glycolysis in HCCLM3 cells but not in Hep-G2 cells, while downregulating PGC1α-mediated mitochondrial biogenesis to enhance cell survival. This may be the primary reason HCCLM3 cells are less sensitive to shikonin than Hep-G2 cells. In summary, shikonin disrupts the energy metabolic homeostasis of HCC cells through multiple molecular targets, thereby providing a critical theoretical foundation for developing metabolism-oriented precision therapies for HCC.
Plumbagin (PLB) is a naturally occurring naphthoquinone isolated from the plant Plumbago zeylanica L. It has been demonstrated to possess anticancer activity against hepatocellular carcinoma and various other types of cancer [28]. The findings of Liu et al. [28] demonstrate that PLB exerts inhibitory effects on the viability of both Huh-7 and Hep-G2 HCC cell lines, concurrently reducing their colony formation capacity. Moreover, PLB induces oxidative stress and DNA damage in HCC cells by elevating intracellular ROS levels and activating cell cycle checkpoints. Building upon the observation that PLB inhibits the proliferation of HCC cells, though the underlying mechanisms remained unclear, Yao et al. [29] conducted a series of in vitro and in vivo experiments to elucidate the molecular basis of this effect. Their findings revealed that PLB suppresses the deubiquitinase activity of USP31, thereby promoting the ubiquitination and subsequent destabilization of glutathione peroxidase 4 (GPX4). The reduced stability of GPX4 leads to a marked accumulation of reactive oxygen species (ROS), ultimately inducing apoptosis in HCC cells. In a separate study, Du et al. [30] investigated the impact of PLB on EMT in HCC cells. Their results revealed that PLB significantly upregulates epithelial markers while downregulating mesenchymal markers. Western blot analysis of EMT-related proteins further confirmed the potent inhibitory effect of PLB on the EMT process in HCC cells. Additionally, in vivo experiments demonstrated that PLB markedly reduces the expression of Snail, vimentin, and N-cadherin. Histopathological examination showed that the number of hepatic tumor foci and pulmonary metastatic lesions in the PLB-treated group was substantially lower than in the control group, providing more direct evidence of PLB’s antitumor efficacy against HCC. Cuproptosis is a novel mode of cell death first comprehensively characterized in 2022. The accumulation of copper within mitochondria facilitates the aggregation of lipoylated dihydrolipoamide S-acetyltransferase (DLAT), triggering this distinct form of cell death. Against the backdrop of cuproptosis research, Wang et al. [17] sought to elucidate the potential involvement of this novel cell death pathway in the antitumor effects of PLB on HCC. Their findings demonstrated that PLB treatment leads to the transcriptional downregulation of ATP7B, a key regulator of intracellular copper homeostasis. This downregulation results in copper accumulation within HCC cells, ultimately triggering cuproptosis. Further mechanistic investigations revealed that PLB induces the downregulation of DNA methyltransferase 1 (DNMT1), thereby promoting the transcription of miR-302a-3p. The upregulated miR-302a-3p subsequently binds to the 3′untranslated region (3′UTR) of ATP7B mRNA, leading to its silencing. This molecular cascade culminates in copper overload and the induction of cuproptosis in HCC cells. In the context of combination therapy with other anti-HCC agents, PLB has been shown to act synergistically with non-plant-derived quinone compounds, such as DOX, to suppress HCC progression. Cao et al. [31] initially confirmed the synergistic cytotoxic effects of DOX and PLB when co-administered in HCC chemotherapy. They subsequently developed a targeted nanoformulation incorporating DOX, PLB, and PLGA-PEG-AEAA. They evaluated its efficacy using both in vitro cell models and a subcutaneous DOX-resistant HCC mouse model. The results demonstrated that, compared to monotherapy, the combination of DOX and PLB synergistically induced apoptosis in DOX-resistant Hepa1-6 cells (Hepa1-6-R cells) and significantly inhibited their migratory capacity. This enhanced efficacy may be attributed to PLB-mediated suppression of aberrant STAT3 activation. Furthermore, compared to non-targeted formulations and the combination of free DOX and PLB, the targeted formulation induced greater apoptosis and more effectively reduced cell migration in Hepa1-6-R cells. In subsequent in vivo experiments, the targeted formulation significantly reduced tumor weight and STAT3 activity within tumor tissues. This study established a mechanistic and therapeutic link between plant-derived and non-plant-derived quinones in the combinatorial treatment of HCC, thereby advancing research on quinone-based combination strategies for liver cancer therapy. Dan Shen is one of China’s most popular traditional herbs and contains various natural quinone compounds. Tanshinone I (Tan I) is one of the well-known major active constituents of Salvia miltiorrhiza. In recent years, it has attracted considerable attention in the research community due to emerging studies. It has been shown to possess potent activities such as antioxidative stress effects, autophagy and apoptosis regulation, and inflammation inhibition [32]. Tanshinone IIA (Tan IIA), a quinonoid bioactive compound from Salvia miltiorrhiza (Danshen), has attracted significant scholarly attention due to its unique structure and potent pharmacological effects. Extensive research has revealed its wide-ranging therapeutic properties, including anti-inflammatory, antioxidant, antiarrhythmic, cardioprotective, and antitumor activities. In recent years, notable progress has been made in understanding its mechanisms and efficacy, particularly in antitumor applications [33]. In studies investigating the therapeutic effects of Tan I on HCC, this compound has been shown to induce G0/G1 phase cell cycle arrest and subsequent apoptosis in liver cancer cells by downregulating cyclin D1 and upregulating p21 expression. Additionally, Tan I triggers apoptosis in Hep-G2 and Huh-7 cells by promoting endoplasmic reticulum (ER) stress through generating reactive oxygen species (ROS) and inhibiting p53-mediated autophagy. Furthermore, another study demonstrated that Tan I impairs genomic stability in HCC cells by suppressing non-homologous end joining (NHEJ) and homologous recombination (HR) DNA repair pathways in a dose-dependent manner [34,35]. Epirubicin (EADM) is a commonly used chemotherapeutic agent for treating hepatocellular carcinoma (HCC); however, like many other anti-HCC drugs, it often leads to the gradual development of drug resistance in cancer cells. The accumulation of hypoxia-inducible factor-1 alpha (HIF-1α) has been identified as a key contributor to EADM resistance in HCC. Several studies have reported that certain tanshinones may modulate HIF-1α expression by targeting the PI3K/AKT signaling pathway [36]. Building upon this research background, Zhao et al. demonstrated that the combined application of Tan I and epirubicin (EADM) enhances the cytotoxicity and growth inhibitory effects of EADM against HCC cells by targeting the PI3K/AKT/HIF-1α signaling pathway. Notably, the combination synergistically reverses HIF-1α-mediated drug resistance, partially overcoming EADM resistance in hepatocellular carcinoma. Regarding safety, in vivo studies in mouse models confirmed that the combination therapy is both safe and effective, and it also mitigates the body weight loss typically induced by EADM treatment [37]. In recent years, PD-1 inhibitors have gained widespread popularity as a treatment option for various tumors. However, their efficacy in treating HCC has been limited [38]. This phenomenon may be attributed to the limited infiltration capacity of tumor-infiltrating lymphocytes (TILs) within the tumor microenvironment, potentially due to abnormal tumor vasculature. Although previous studies have shown that Tan IIA can enhance vascular integrity and promote vascular normalization, the precise molecular mechanisms underlying these effects remain poorly understood [39]. Mao et al. [40] reported that tanshinone IIA (Tan IIA) enhances vascular integrity in situ by promoting the expression of tight junction proteins—ZO-1, Occludin, and Claudin-5—through inhibition of ELTD1 expression, while concurrently suppressing JAK1 and JAK2 signaling pathways. These effects contribute to the remodeling of the immunosuppressive tumor microenvironment and the inhibition of tumor growth. Furthermore, combining Tan IIA with PD-1 inhibitors resulted in significantly greater tumor suppression than PD-1 inhibitor monotherapy in animal models. However, relatively few studies have explored the synergistic effects of Tan IIA in combination with conventional chemotherapeutic agents. Although these studies remain preliminary, emerging evidence suggests that combining Tan IIA with sorafenib or its derivative SC-1 significantly suppresses cell migration, invasion, and sorafenib resistance in HCC cells [41]. The observed reduction in drug resistance is likely attributable to the combination’s potent inhibition of STAT3 signaling. Furthermore, this combined treatment increases the sub-G1 cell population in Huh-7 and Hep-G2 cells, accompanied by pronounced activation of caspases, indicating enhanced apoptosis.
TQ is a plant-derived natural quinone compound extracted from Nigella sativa [42]. Due to its antioxidant, hepatoprotective, anticancer, and anti-inflammatory properties, there is growing interest in its pharmacological mechanisms and therapeutic effects [43]. Studies by Ibrahim, Tadros, and Farghaly [44,45,46] suggest that TQ may inhibit angiogenesis in HCC by regulating miR-1-3p. Additionally, data from related animal models indicate that TQ effectively prevents changes in liver enzymes and serum proteins while reducing the expression levels of HCC markers such as AFP, AFP-L3, and GPC3. Furthermore, TQ significantly decreases BCL-2 expression and upregulates the BAX/BCL-2 ratio and CASP3 expression levels. Regarding combination therapies involving common anti-HCC chemotherapeutic agents, TQ combined with cisplatin has significantly improved liver pathology. TQ enhances the efficacy of cisplatin by mitigating oxidative stress and downregulating the endoplasmic reticulum stress marker GRP78, thereby substantially reducing cisplatin-induced oxidative damage. Moreover, TQ inhibits angiogenesis in HCC and slows disease progression by modulating multiple signaling pathways and molecular targets. Significantly, TQ not only potentiates the therapeutic effects of cisplatin but also attenuates some of the drug’s adverse side effects. The relevant mechanisms underlying the anti-HCC effects of these plant-derived quinone compounds are summarized in Figure 2. Additionally, the sources of these compounds and the outcomes of corresponding animal and cellular studies are briefly detailed in Table 1.
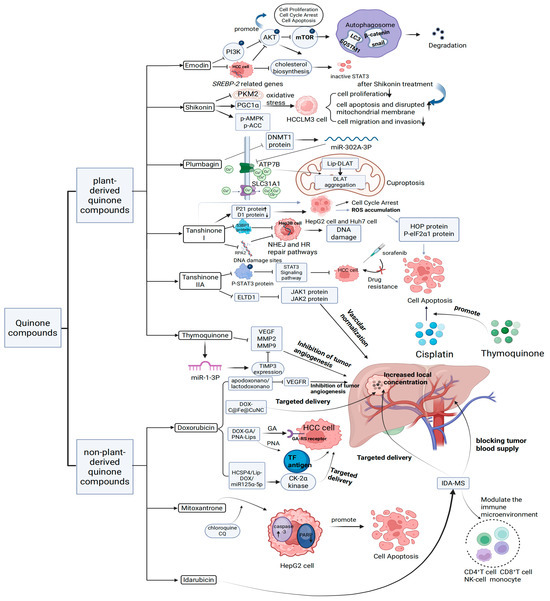
Figure 2.
Mechanism of action of plant-derived quinones and non-plant-derived quinones on HCC.

Table 1.
The source of plant-derived quinone compounds and their animal and cell experiments in the treatment of HCC.
3.2. Clinical Studies on Plant-Derived Quinonoid Compounds
Investigations into the anti-HCC effects of the aforementioned plant-derived quinonoid compounds have primarily been confined to in vitro cellular assays and animal models, resulting in relatively limited clinical data on their efficacy as individual agents. Nevertheless, the source plants of these quinones are frequently incorporated into compound formulations currently used in clinical management of HCC, offering valuable insights and reference points for future research and therapeutic innovation. For example, in formulations such as the Da Huang Zhe Chong Formula and rhubarb-based sodium tablets studied by Hou, Tian, Deng, et al. [47,48,49], the emodin present in rhubarb may contribute to therapeutic effects in patients with HCC. Notably, post-embolization syndrome following TACE was significantly reduced in HCC patients treated with the Da Huang Zhe Chong Formula. Additionally, patients receiving interventional therapy for liver cancer exhibited marked improvements in liver function and enhanced immune responses. Furthermore, treatment with rhubarb-based sodium tablets effectively improved serum biomarkers, including ALT, TBIL, and AFP, while significantly reducing complications such as upper gastrointestinal bleeding.
Danshen injection is a traditional Chinese medicinal preparation primarily derived from Salvia miltiorrhiza, the primary source of Tan I and Tan IIA. Clinical studies conducted by Zhu, Zhang, et al. [50,51] have demonstrated that Danshen injection may enhance immune function in patients with intermediate to advanced HCC by increasing levels of CD4+ T cells, CD3+ T cells, the CD4+/CD8+ T cell ratio, and serum immunoglobulins IgM, IgG, and IgA. Furthermore, Danshen injection has been reported to improve clinical symptoms and signs and to prolong patient survival (Table 2).
Table 2.
The source of plant-derived quinone compounds and their clinical experiments in the treatment of HCC.
3.3. Adverse Reactions of Plant-Derived Quinonoid Compounds
Although plant-derived quinone compounds have shown preliminary potential to inhibit HCC via multiple pathways and targets, they are not without specific toxicities. The toxic effects of these compounds may pose significant challenges to their advancement into clinical trials. Among them, emodin—a natural plant-derived quinone with anticancer properties—has been reported to exhibit fewer side effects compared to conventional chemotherapeutic agents [52]. The primary toxicities associated with emodin include reproductive toxicity, genotoxicity, and mutagenicity. Oshida et al. found that emodin can impair spermatogenic function in mice and interfere with the expression of genes related to spermatogenesis. Drinking an emodin solution at a concentration of 20–40 μM can reduce the maturation rate of mouse oocytes and the in vitro fertilization rate, thereby damaging early embryonic development. When emodin concentration exceeds 0.93 μM, the compound adversely affects the embryo survival rate and hatching rate of zebrafish [53,54,55]. The toxicity of shikonin primarily manifests as skin toxicity, cytotoxicity, hepatotoxicity, and metabolic toxicity. Cheng et al. [56] and Figat et al. [57] reported that shikonin acts as an inhibitor of uridine 5′-diphosphate glucuronosyltransferase (UGT), contributing to hepatotoxicity in both humans and rats. Additionally, shikonin exhibits potent cytotoxicity against the conventional V79 cell line. Tang et al. [58] further demonstrated that shikonin may inhibit multiple cytochrome P450 enzymes in mammals. Acting as a mixed and competitive inhibitor of CYP1A2, CYP2C9, CYP2B6, CYP2D6, CYP3A4, and CYP2E1, shikonin has the potential to interfere with drug metabolism and cause drug–drug and food–drug interaction toxicities. The toxicity profile of PLB is primarily characterized by reproductive toxicity in female animals and, to a lesser extent, hepatotoxicity. In addition, a limited number of studies have reported that PLB may also exhibit considerable genotoxicity and antigenic toxicity [59]. Kumar et al. [60] administered three crude extracts of PLB to Wistar rats, using petroleum ether, acetone, and 70% ethanol as extraction solvents. The experimental results showed that all PLB extracts induced hepatic and renal congestion in the animals, accompanied by elevated levels of biochemical markers such as AST and urea. Notably, since the petroleum ether extract contained the highest concentration of PLB, rats receiving this extract at equivalent maximum doses exhibited more rapid mortality. Lim et al. [61] focused specifically on the reproductive toxicity of PLB. To assess their developmental toxicity, they investigated four 1,4-naphthoquinone derivatives, including PLB, by exposing both wild-type and transgenic zebrafish embryos. The study found that PLB treatment significantly increased embryo lethality and reduced hatching rates. Notably, when zebrafish embryos were exposed to 0.13 µM PLB at 4 h post-fertilization, severe impairment of brain differentiation and the manifestation of pronounced cyclopia were observed. Embryos exhibiting cyclopia ultimately failed to survive. The toxicity of TQ primarily manifests as genotoxicity and teratogenicity. Abukhader et al. [62] demonstrated that TQ administered at doses of 35 mg/kg and 50 mg/kg induced significant adverse effects on embryonic development during pregnancy in rats. Toxicological data and side effect profiles for Tan I, both in vivo and in vitro, remain insufficient and require further investigation [63]. However, Huang et al. [64] reported that all lipophilic tanshinones cause various developmental defects in zebrafish. Specifically, Tan IIA has been associated with teratogenic effects and potential cardiotoxicity. Wang et al. [65] observed that zebrafish embryos exposed to different concentrations of Tan IIA for four days exhibited pronounced pericardial edema, spinal curvature, and tail loss. Tan IIA showed evidence of cardiotoxicity at higher concentrations in zebrafish embryos. A summary of the sources, toxicities, and adverse effects of these plant-derived quinones is provided in Table 3.
Table 3.
Possible adverse reactions of plant-derived quinones.
4. Research Progress of Non-Plant-Derived Quinone Compounds in the Field of Anti-HCC
4.1. Experimental Studies of Non-Plant-Derived Quinone Compounds in the Field of Anti-HCC
Anthracyclines represent a class of aromatic type II polyketide compounds characterized by a linear tetracyclic 7,8,9,10-tetrahydro-5,12-naphthacenedione core. This scaffold features a polyhydroxylated anthraquinone structure fused with a fourth saturated substituted ring, forming the aglycone moiety typically conjugated to one or more sugar residues. The history of anthracyclines in cancer therapy dates back to the 1960s with the discovery of daunomycin, whose remarkable efficacy against lymphoma significantly propelled research into this class of molecules. Since then, continuous structural optimization and biosynthetic engineering—particularly through modified bacterial strains—have produced hundreds of anthracycline analogs. Beyond the three clinically relevant agents discussed in this paper—DOX and IDA, commonly used in HCC therapy—numerous other anthracyclines have been approved for clinical application. For instance, amrubicin (Amr) is employed in the treatment of lung cancer, while epirubicin is widely used in adjuvant therapy for hormone receptor-positive breast cancer. However, a major limitation of this class, particularly in the case of DOX, is its pronounced cardiotoxicity. Therefore, a central focus of current research is the development of strategies to mitigate this adverse effect while simultaneously enhancing the therapeutic efficacy of anthracyclines [66].
Doxorubicin is a cytotoxic anthracycline antibiotic isolated from Streptomyces peucetius cultures. Its anticancer activity primarily arises from intercalation into DNA nucleotide bases. Specifically, DOX disrupts the DNA replication process by stabilizing the topoisomerase II-DNA cleavage complex, thereby preventing the re-ligation of the DNA double helix and effectively terminating DNA replication [67]. Although its severe side effects limit the clinical application of DOX, recent advances in nanocarrier technologies may help mitigate these issues while enhancing its therapeutic efficacy. Bhaskar, Saddik, and colleagues [68,69] independently developed deferroprotein- and nano-lactoferrin-loaded DOX formulations and a DOX-loaded C@Fe@Cu nanocomposite. Their studies demonstrated that both types of protein-based DOX nanoparticles effectively treated HCC in rat models. Notably, the nanoprotein-treated groups exhibited significantly higher hepatic DOX concentrations than those receiving free DOX. Furthermore, these groups showed greater upregulation of p53 and p21 expression, suggesting enhanced apoptotic activity relative to DOX alone. In parallel, the C@Fe@Cu nanocomposite formulation markedly reduced the IC50 of DOX, indicating increased cytotoxic potency against liver cancer cells. Cells treated with the DOX-C@Fe@Cu nanocomposite also exhibited a higher proportion of late apoptotic cells than those treated with an equivalent dose of free DOX. These findings suggest that nanocarrier systems based on nanoproteins or metal–organic composites may offer significant advantages in augmenting the anti-HCC efficacy of DOX, presenting a promising new strategy for liver cancer therapy. Similarly, liposomes—widely regarded as non-toxic, biocompatible, and non-immunogenic drug carriers—also hold considerable potential [70,71]. Liposomes are widely utilized to encapsulate lipophilic and hydrophilic drugs, enhancing their pharmacokinetic and pharmacodynamic profiles to improve therapeutic efficacy and reduce systemic toxicity [72]. Li, Zhao, and colleagues [72,73] respectively developed DOX-loaded liposomal formulations modified with glycyrrhetinic acid (GA) and peanut agglutinin (PNA) as targeting ligands (DOX-GA/PNA-Lips), as well as an HCC-targeted liposomal delivery system designed to overcome multidrug resistance (MDR), termed HCSP4/Lipo-DOX/miR125a-5p. Their studies showed that DOX-GA/PNA-Lips significantly enhanced targeting efficiency toward the MUC1-positive HCC cell line SMMC-7721. In vivo, mice treated with the liposomal DOX formulation exhibited reduced tumor volume and weight and less body weight loss compared to those receiving free DOX. Additionally, the HCSP4/Lipo-DOX/miR125a-5p formulation effectively inhibited MDR development by downregulating the expression of MDR-associated genes. These findings highlight the potential of ligand-modified and gene-regulatory liposomal systems in enhancing DOX delivery and overcoming resistance mechanisms in HCC therapy. After treatment with HCSP4/Lipo-DOX/miR125a-5p, the expression levels of ABCB1, ABCC5, ATP1B1, and EZH2 were significantly reduced in Hep-G2 and Hep-G2/ADR cells. In summary, this study provides a liposome-based DOX delivery strategy that enhances the targeting efficiency of HCC treatment and effectively inhibits the development of MDR, offering new ideas and experimental evidence for HCC therapy.
Due to the severe cardiotoxicity associated with DOX, significant research efforts have focused on mitigating its cardiac side effects through various strategies. They also pursue the development of novel anticancer agents with reduced cardiotoxic profiles. In the 1980s, MTX was introduced as a synthetic analogue of DOX [74]. As a member of the anthracycline class, MTX is widely used to treat malignancies such as lymphoma and bone marrow cancers. However, its clinical efficacy in HCC, particularly in patients with advanced-stage disease, has been limited [75]. Autophagy, an evolutionarily conserved cellular process, enables the degradation and recycling of intracellular components in response to stress and environmental stimuli [76]. This mechanism may contribute to the survival of HCC cells during MTX treatment by mitigating drug-induced cytotoxic stress [77]. Using HCC cells as a model, Xie et al. [77] observed a significant increase in LC3-II levels and a concomitant decrease in p62 levels following MTX treatment, indicating enhanced autophagic activity. In the Hep-G2 group treated with a low dose of MTX, an increase in Beclin-1 was observed, whereas in the high-dose MTX-treated Hep-G2 group, Beclin-1 levels slightly decreased. Additionally, in MTX-treated Hep-G2 cells, phosphorylation of p70S6K at Thr389 and total protein levels were significantly reduced. They then used chloroquine (CQ) to inhibit autophagy in MTX-treated Hep-G2 cells and measured apoptosis. Compared to the MTX-only group, the combined treatment group (MTX + CQ) showed a significantly higher cleavage rate, indicating that CQ enhanced MTX-induced apoptosis in Hep-G2 cells by inhibiting autophagy.
In the ongoing effort to develop anthracycline-based antitumor agents with reduced cardiotoxicity, IDA was introduced in 1976 [78]. Structurally, IDA differs from daunorubicin by substituting a hydrogen atom for the C4 methoxy group on the aglycone D-ring. This seemingly minor modification significantly increases the drug’s lipophilicity, leading to substantially higher plasma concentrations of its metabolites than daunorubicin [79]. Due to its improved lipophilicity and extended biological half-life, IDA is frequently employed in TACE to treat HCC. However, conventional TACE formulations often suffer from emulsion instability, which leads to rapid phase separation and inadequate local drug accumulation within the tumor site, thereby limiting therapeutic efficacy. To overcome the above limitations, Zheng et al. [80] developed a biodegradable microsphere loaded with idarubicin (IDA-MS). They then used IDA-MS for TACE treatment in VX2 rabbit and C57BL/6 mouse models. In the rabbit VX2 HCC model, TACE with IDA-MS significantly inhibited tumor growth and blocked tumor blood vessels more effectively. In the mouse HCC model, IDA-MS markedly reduced tumor volume and weight and improved the pathological condition of tumor tissues. This study indicates that a biodegradable microsphere-based IDA delivery system has potential advantages in TACE treatment of HCC, providing new experimental evidence for optimizing chemotherapy embolization strategies for HCC. The dosing form of the above non-plant-derived quinones and the associated mechanisms of anti-HCC action can be seen in Figure 2. The source, mode of administration, or form of the above non-plant-derived quinones and the corresponding efficacy are briefly documented in Table 4.
Table 4.
The source of non-plant-derived quinone compounds and their animal and cell experiments in the treatment of HCC.
4.2. Clinical Studies of Non-Plant-Derived Quinone Compounds in the Field of Anti-HCC
DOX, a widely used anthracycline chemotherapeutic agent, exhibits potent antitumor activity against various malignancies. However, its clinical utility is severely limited by its significant systemic toxicity. In a study by Lai et al. [81], 60 patients with inoperable HCC were randomly assigned to receive DOX every three weeks, while 46 comparable patients received no anticancer therapy. Although the DOX-treated group demonstrated a higher median survival compared to the untreated group, approximately 25% of patients experienced fatal complications, underscoring the drug’s severe toxicity. To mitigate these adverse effects, researchers have explored liposomal formulations of DOX in treating HCC. Valle, Tak, and colleagues [82,83] evaluated two such approaches: pegylated liposomal doxorubicin (PLD) and lyso-thermosensitive liposomal doxorubicin (LTLD) in combination with radiofrequency ablation (RFA). Patients treated with PLD exhibited improved tolerance to several of DOX’s toxic side effects. Although the combination of RFA and LTLD did not show a significant safety advantage over RFA alone, its therapeutic efficacy was notably enhanced—particularly when RFA was applied to isolated lesions for 45 min or longer.
MTX has been used in the clinical treatment of HCC for the last century. Shepherd, Farres et al. [84,85] conducted hepatic arterial infusion of MTX and minimally invasive intratumoral injection of MTX in patients with HCC. Under the hepatic arterial infusion treatment, MTX demonstrated potent activity against primary HCC; However, the drug appeared to exhibit toxic effects similar to those of epirubicin. Meanwhile, in malignant hepatic lesions with no other therapeutic options, minimally invasive intratumoral injection of MTX promoted tumor necrosis and showed a high safety profile. With the rapid advancement of nanocarrier technology, clinical research on MTX treatment for HCC has also achieved breakthroughs. Zhou et al. [86] compared the therapeutic effects of Mitoxantrone (Dihydroxyanthracenedione, DHAD) delivered via polybutylcyanoacrylate nanoparticles (DHAD-PBCA-NPs) to those of free DHAD in patients with HCC. The DHAD-PBCA-NP group demonstrated improved clinical outcomes, including greater disease stabilization, reduced disease progression, and a lower incidence of leukopenia compared to the group receiving DHAD alone. However, the incidence of anemia was lower in the DHAD-treated group than in the DHAD-PBCA-NP group. Mitomycin C (MMC), a cytotoxic antibiotic originally isolated from Streptomyces caespitosus, has shown broad-spectrum antitumor activity and has long been investigated in the clinical management of HCC [87]. Clinical studies by Cheirsilpa et al. [88] revealed that high-dose MMC exhibited selective antitumor efficacy against HCC with a more favorable toxicity profile than doxorubicin. Idarubicin (IDA), a second-generation anthracycline, has also demonstrated superior cytotoxicity against HCC cells in preclinical in vitro studies [89]. Growing clinical evidence further supports the potential of IDA to replace DOX in transarterial chemoembolization (TACE) regimens for HCC. Boulin et al. [90] administered microspheres loaded with varying doses of IDA to patients via TACE. They observed that the maximum tolerated dose of IDA loaded on the drug-eluting beads was 10 mg per TACE session. This clinical study’s median overall survival was significantly higher than conventional TACE treatment. Furthermore, their findings demonstrated that IDA-loaded drug-eluting beads were more effective in treating HCC than intravenous administration of IDA. Teyssier et al. [91] retrospectively reviewed the medical records and imaging studies of HCC patients who consecutively underwent Doxorubicin-TACE (Dox-TACE) or Idarubicin-TACE (Ida-TACE) treatments at their hospital from 2012 to 2014. Comparing the objective response rates, complete response rates, and safety profiles of the two TACE treatments, the results indicated that Ida-TACE has promising potential as a superior alternative to Dox-TACE for treating intermediate-stage HCC.
In summary, although the aforementioned non-plant-derived quinones have demonstrated vigorous anti-HCC activity and have been combined with other antitumor therapies in subsequent studies to maximize their clinical effectiveness, their side effects, toxicity, and inability to achieve a cure for HCC remain significant obstacles to the future development of these drugs. The clinical efficacy of the above non-plant-derived quinones is briefly documented in Table 5.
Table 5.
The source of non-plant-derived quinone compounds and their clinical experiments in the treatment of HCC.
4.3. Adverse Reactions and Toxicity of Non-Plant-Derived Quinone Compounds
In the early years of doxorubicin (DOX) clinical use, its most common side effects were identified as acute nausea and vomiting, stomatitis, alopecia, and gastrointestinal disturbances. With broader clinical application, DOX was found to exert varying degrees of toxicity on multiple organs, including the heart, brain, kidneys, and liver. Among these, DOX-induced cardiotoxicity is widely recognized as a major dose-limiting factor, as the heart is particularly sensitive to its toxic effects. DOX can induce cardiomyocyte hypertrophy and ultrastructural alterations, notably affecting mitochondrial integrity. The underlying mechanisms may involve enhanced redox cycling with NADH dehydrogenase, altered expression of mitochondrial proteins, DNA damage, and the activation of autophagic pathways. Regarding neurotoxicity, DOX has been shown to stimulate the production of tumor necrosis factor-alpha (TNF-α), activating microglial cells in the brain, releasing pro-inflammatory cytokines. Elevated TNF-α upregulates inducible nitric oxide synthase (iNOS), increasing reactive nitrogen species (RNS) levels and promoting nitration of manganese superoxide dismutase (MnSOD). This nitration reduces MnSOD activity, producing elevated reactive oxygen species (ROS). Excess ROS facilitates the opening of the mitochondrial permeability transition pore (PTP), triggering cytochrome c release and initiating apoptosis. Regarding nephrotoxicity, DOX can damage glomerular podocytes, contributing to renal pathologies. Clinically, this manifests as severe proteinuria, renal enlargement, and increased glomerular capillary permeability. Mitochondrial dysfunction and lipid peroxidation are also implicated in the pathogenesis of DOX-induced nephropathy. With respect to hepatotoxicity, DOX-induced ROS generation leads to elevated activities of superoxide dismutase (SOD) and glutathione peroxidase (GPX), increased DNA damage, and depletion of key antioxidants such as glutathione (GSH) and vitamin E. These effects contribute to oxidative stress and liver injury. Moreover, ROS may activate IκB kinase (IKK), which promotes the expression of pro-inflammatory cytokines and ultimately leads to hepatocyte death [92]. In monitoring adverse reactions during DOX treatment for HCC, mild vomiting and hair loss are relatively common. At the same time, cardiotoxicity occurs more frequently and is generally more severe compared to other toxicities [81,93,94].
In the clinical monitoring of adverse reactions to MTX in the treatment of HCC, typical side effects include vomiting and alopecia. However, bone marrow suppression and cardiotoxicity are also frequently observed and may be life-threatening [84,86,95]. Although the precise mechanisms underlying MTX-induced cardiotoxicity remain incompletely understood, several contributing factors have been identified. These include generating reactive ROS through interactions between MTX and iron, inhibition of topoisomerase II, and mitochondrial damage—all of which contribute to oxidative stress and cardiac dysfunction [96].
The most common toxic side effect of MMC is delayed bone marrow suppression. Additionally, diarrhea, anorexia, and hair loss are also relatively common. Hair loss, rash, and stomatitis occur occasionally [97]. Other rare but potentially fatal side effects include hemolytic uremic syndrome, interstitial pneumonia, and heart failure. In clinical studies of MMC treatment for HCC, patients were observed to experience varying degrees of bone marrow suppression, and the majority of patients developed symptoms of nausea and vomiting [88,98].
In clinical trials evaluating IDA for treating various malignancies, bone marrow suppression has been identified as the primary dose-limiting toxicity. Gastrointestinal toxicity is the second most frequently reported adverse effect and has also proven dose-limiting in specific phase I studies. In addition, IDA is associated with a broad spectrum of dermatological toxicities and exhibits some degree of cardiotoxicity. Notably, extravasation of IDA during administration can result in severe local tissue damage, including ulceration and necrosis [99]. The adverse effects and toxicities associated with the above non-plant-derived quinones are summarized in Table 6.
Table 6.
Possible adverse reactions of non-plant-derived quinones.
5. Investigation of the Attenuation Effects of Plant-Derived Quinone Compounds on the Toxicity of the Non-Plant-Derived Quinone Compound Dox
Due to the severe side effects and high toxicity of non-plant-derived quinone-based anti-HCC drugs, recent years have witnessed notable progress in research on plant-derived quinone compounds for treating HCC. Simultaneously, some researchers have investigated whether these plant-derived quinones can help mitigate the toxic side effects associated with non-plant-derived quinone anti-HCC drugs. Most studies have centered on the detoxifying effects of DOX. One study on emodin demonstrated its ability to restore the reduced ejection fraction (EF) and end-systolic volume (ES) in DOX-treated mouse hearts. In addition, emodin significantly lowered serum levels of CK-MB, LDH, and IL-1β, alleviated cellular disarray, and corrected disorganized myocardial fiber structures, thereby improving DOX-induced cardiac dysfunction and myocardial injury. Likewise, shikonin was shown to reduce DOX-induced elevations in MDA, 4-HNE, and NADPH oxidase activity, while increasing the suppressed levels of GSH and SOD, thereby mitigating DOX-induced oxidative stress damage [100,101]. Compared to other plant-derived quinones, baicalein exhibits a broader detoxifying effect against doxorubicin (DOX)-induced toxicity. Karabulut et al. [102,103,104] investigated the detoxifying effects of thymoquinone (TQ) on DOX-induced toxicity from three perspectives: cardiotoxicity, nephrotoxicity, and reproductive toxicity. Regarding cardiotoxicity, TQ restored the myocardial fiber tissue damaged by DOX to near-normal levels and significantly reduced the expression of molecular chaperones HSP70, HSP90, and GRP78 in the cytoplasm of rat myocardial fibers. Regarding nephrotoxicity, TQ alleviated the severity of DOX-induced kidney injury and significantly increased total antioxidant status (TAS). In the context of reproductive toxicity, TQ mitigated the reduction in testicular weight caused by DOX and markedly decreased the expression levels of Caspase-3 and HSP90 in rat reproductive cells. Compared to TQ, tanshinone I (Tan I) and tanshinone IIA (Tan IIA) display a more targeted detoxifying effect against DOX-induced cardiotoxicity. Jiang, Xu, and colleagues [105,106] conducted experimental evaluations on the cardioprotective properties of Tan I and Tan IIA. Tan I was shown to dose-dependently restore the reduced ejection fraction (EF) and end-systolic volume (ES) caused by DOX and attenuate DOX-induced cardiomyocyte apoptosis. Meanwhile, Tan IIA significantly enhanced cell viability and prevented cardiomyocyte apoptosis induced by DOX. Furthermore, Tan IIA preserved myocardial architecture and prevented fiber disarray, restoring cardiac function impaired by DOX. The protective effects of the above plant-derived quinones against DOX-induced toxicity are summarized in Table 7.
Table 7.
The detoxifying effect of plant-derived quinone compounds on doxorubicin.
6. Clinical Translation of Quinonoid Compounds—Challenges and Limitations
Overall, quinone-based compounds currently face significant challenges in clinical translation and further development. Although anthraquinone derivatives can alter cellular redox states through iron-dependent lipid peroxidation, increasing ROS production, and inducing oxidative stress in tumor cells, their pharmacokinetic profiles reveal poor oral absorption. As a result, these compounds are predominantly administered via intravenous injection. Moreover, their tendency to accumulate in cardiac tissue raises concerns regarding potentially severe cardiotoxicity [66]. Naphthoquinone derivatives, represented by shikonin, can undergo redox cycling in isolated mitochondria, generating ROS. However, naphthoquinones’ generally poor water solubility limits their utility in cancer therapy. Pharmacokinetically, naphthoquinones exhibit considerable variability across different compounds. For instance, shikonin demonstrates low bioavailability. To address this, approaches such as encapsulation in nanogels have been explored to enhance its stability both in vitro and in vivo [25,107]. TQ, the reduced form of thymoquinone, a representative plant-derived benzoquinone, exhibits potent antioxidant activity through its high free radical scavenging capacity. It has shown promise in treating chronic hepatic and renal injuries. However, TQ’s high lipophilicity and limited hydrophilicity present significant formulation challenges, impeding its progression into clinical trials [108]. Tan I and Tan IIA, representative plant-derived phenanthrenequinones, have relatively low toxicity profiles. Tan IIA, in particular, contains conjugated double bonds, a dihydrofuran ring, and quinone carbonyl groups that enable it to scavenge free radicals and interrupt lipid peroxidation chain reactions, thereby exerting antioxidant effects and protecting the DNA of normal cells. Nevertheless, Tan IIA’s high lipophilicity, coupled with its hydrophilic derivatives’ rapid metabolism and short half-life, compromises its biosafety and limits its clinical applicability. Similarly, Tan I benefits from a structure conducive to free radical scavenging and modulation of intracellular antioxidant signaling pathways. Unfortunately, its pronounced lipophilicity also poses significant barriers to clinical translation, necessitating further research and development [33,63].
7. Conclusions and Prospects
The selected non-plant-derived quinone compounds discussed in this article are widely used chemotherapeutic agents in clinical settings. Although they exhibit consistent anti-HCC efficacy and remain integral to current treatment protocols, their associated toxic side effects can significantly compromise patients’ quality of life during therapy. Consequently, current research focuses on reducing these toxicities while maintaining or enhancing therapeutic effectiveness. In addition to exploring combination therapies that achieve synergistic antitumor effects and reduced toxicity, applying advanced drug delivery technologies—such as nanoparticle-based systems—also plays a critical role in optimizing clinical outcomes. Among the four non-plant-derived quinone compounds, DOX has garnered significant attention from researchers in recent years. Numerous studies have focused on the development of drug delivery systems based on nanoproteins or metal–organic frameworks to enhance the anti-HCC efficacy of DOX while minimizing its associated side effects. IDA, often regarded as a clinical “counterpart” to DOX, has been the subject of multiple comparative clinical studies in recent years. Notably, IDA has demonstrated increasing advantages over DOX in TACE therapy, gradually emerging as a potential substitute [91,109,110,111]. In contrast, in recent years, mitoxantrone and mitomycin have received relatively limited clinical research attention in the context of HCC treatment. Further optimization of their therapeutic efficacy and safety profiles is warranted to fully explore their potential in anti-HCC strategies. Research on plant-derived quinone compounds for the treatment of HCC has steadily increased in recent years. These studies aim to demonstrate such compounds’ inhibitory or therapeutic effects on HCC cells and liver tumors through in vitro and in vivo experiments, pathological evaluations, and biochemical marker analyses. In parallel, growing attention is being directed toward their potential to enhance the efficacy and reduce the toxicity of conventional chemotherapeutic agents. However, due to the limited depth of investigation into the anti-HCC properties of these compounds, most available studies remain restricted to cell-based and animal models, and their overall number is still relatively small. Among the six plant-derived quinone compounds, only emodin and Tan I and Tan II are potentially present in traditional Chinese medicinal formulations such as Dahuang Zhechong Formula and Danshen Injection, respectively. However, current clinical evidence does not conclusively attribute the therapeutic effects of these formulations to the presence of these specific compounds. As a result, the pharmacological profiles of plant-derived quinones remain largely underexplored, and further investigation is warranted. Nonetheless, PLB has attracted increasing academic interest in recent years, particularly in the context of HCC. A growing body of research has focused on elucidating its pharmacodynamics and underlying mechanisms of action, including novel pathways such as cuproptosis [17]. This surge in mechanistic studies suggests that PLB may hold greater developmental potential than other plant-derived quinones, positioning it as a promising candidate for future anti-HCC drug development. Moreover, the number of plant-derived quinones conclusively shown to possess anti-HCC activity remains limited. Therefore, it is essential to expand the scope of investigation into plant-derived quinone compounds to identify additional candidates with potential for anti-HCC applications. Simultaneously, more extensive and in-depth in vitro and in vivo studies are required to elucidate further the mechanisms by which plant-derived quinones with established research backgrounds exert their effects, aiming to advance these compounds toward clinical evaluation. Additionally, considering that several of the plant-derived quinones discussed in this article have demonstrated varying degrees of toxicity mitigation when used in combination with DOX—a non-plant-derived quinone with well-documented anti-HCC efficacy—future research should also explore the potential of these compounds as adjuvants to reduce associated toxicity while maintaining or enhancing their therapeutic effects. Moreover, the combination of PLB and DOX synergistically inhibited the migration and proliferation of drug-resistant HCC cells [31]. We hypothesize that combining plant-derived quinones with DOX may mitigate DOX-induced toxicity and enhance the overall anti-HCC efficacy of both agents.
Furthermore, we speculate that these plant-derived quinone compounds may also exert detoxifying effects on other non-plant-derived quinones mentioned in this study. Their combined use could potentially lead to improved therapeutic outcomes in treating HCC. In summary, quinone compounds—especially those derived from plants—still hold significant research value and development potential in HCC treatment.
Author Contributions
Y.Y. reviewed the literature and wrote the text, while Y.M., M.L. and K.P. were responsible for drawing and submitting the manuscript. Z.L., Y.W., X.H., S.L. and G.L. provided guidance for this article. All authors have read and agreed to the published version of the manuscript.
Funding
Natural Science Foundation of Jilin Province [grant numbers YDZJ202501ZYTS161].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Figures have been created with BioRender.com.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Abbreviations
| Tan IIA | Tanshinone IIA |
| Tan I | Tanshinone I |
| PLB | Plumbagin |
| DOX | Doxorubicin |
| MTX | Mitoxantrone |
| IDA | Idarubicin |
| MMC | Mitomycin |
| PKC | Protein Kinase C |
| ERK5 | Extracellular signal-regulated kinase 5 |
| ADAMTS4 | A Disintegrin And Metalloproteinase With Thrombospondin 4 |
| MMP3 | Matrix Metalloproteinase 3 |
| VEGF | Vascular Endothelial Growth Factor |
| SREBP-2 | Sterol Regulatory Element-Binding Protein 2 |
| ROS | Reactive Oxygen Species |
| PKM2 | Pyruvate kinase isozyme type M2 |
| HIF1α | Hypoxia-inducible factor 1 alpha |
| ZO-1 | Zona Occludens 1 |
| ELTD1 | EGF, latrophilin and seven transmembrane domain containing 1 |
| JAK1 | Janus Kinase 1 |
| JAK2 | Janus Kinase 2 |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| AFP | Alpha-Fetoprotein |
| AFP-L3 | Alpha-Fetoprotein isoform L3 |
| GPC3 | Glypican 3 |
| Bcl-2 | B-cell lymphoma-2 |
| Bax | Bcl-2 associated X protein |
| TACE | Transcatheter Arterial Chemoembolization |
| ALT | Alanine Aminotransferase |
| TBIL | Total Bilirubin |
| DHAD | Dihydroxyanthracenedione |
| DHAD-loaded PBCA nanoparticles | Mitoxantrone-loaded Polybutylcyanacrylate Nanoparticles |
| HCC | Hepatocellular Carcinoma |
| TNF-α | Tumor Necrosis Factor-alpha |
| iNOS | Inducible Nitric Oxide Synthase |
| RNS | Reactive Nitrogen Species |
| MnSOD | Manganese Superoxide Dismutase |
| PTP | Permeability Transition Pore |
| SOD | Superoxide Dismutase |
| GPX | Glutathione Peroxidase |
| GSH | Glutathione |
| IKK | IκB kinase |
| HSP70 | Heat Shock Protein 70 |
| HSP90 | Heat Shock Protein 90 |
| TAS | Total Antioxidant Status |
| TQ | Thymoquinone |
| UGT | Uridine 5′-diphosphate Glucuronosyltransferase |
| MDR | Multidrug Resistance |
| IDA-MS | Biodegradable Microsphere Loaded with Idarubicin |
| PLD | Pegylated Liposomal Doxorubicin |
| RFA | Radiofrequency Ablation |
| LTLD | Lyso-thermosensitive Liposomal Doxorubicin |
| EF | Ejection FractionES: End-systolic Volume |
| CK-MB | Creatine Kinase-MB |
| LDH | Lactate Dehydrogenase |
| IL-1β | Interleukin-1 beta |
| GRP78 | Glucose Regulated Protein 78 |
| AKT | Oncogenic protein kinase B |
| EMT | Epithelial-mesenchymal transition |
| NHEJ | Non-homologous end joining |
| HR | Homologous recombination |
| TILs | Tumor-infiltrating lymphocytes |
| GA | Glycyrrhetinic acid |
| PNA | Peanut agglutininCQ: Chloroquine |
| GPX4 | Glutathione peroxidase 4 |
| DLAT | Lipoylated dihydrolipoamide S-acetyltransferase |
| DNMT1 | DNA methyltransferase 1 |
| Amr | Amrubicin |
| Epi | Epirubicin |
| Hepa1-6-R cells | DOX-resistant Hepa1-6 cells |
| BCLC | Barcelona Clinic Liver Cancer |
| MMP-2 | Matrix metalloproteinase-2 |
| MMP-9 | Matrix metalloproteinase-9 |
| p53 | Tumor protein p53 |
| MSP | Methylation-specific PCR |
| Hb | Hemoglobin |
| ECG | Electrocardiography |
| 2D Echo | Two-dimensional echocardiography |
| LVEF | Left ventricular ejection fraction |
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Qu, L.; Ma, X.; Fan, D. Ginsenoside Rk3 Suppresses Hepatocellular Carcinoma Development through Targeting the Gut-Liver Axis. J. Agric. Food Chem. 2021, 69, 10121–10137. [Google Scholar] [CrossRef]
- Rumgay, H.; Arnold, M.; Ferlay, J.; Lesi, O.; Cabasag, C.J.; Vignat, J.; Laversanne, M.; McGlynn, K.A.; Soerjomataram, I. Global burden of primary liver cancer in 2020 and predictions to 2040. J. Hepatol. 2022, 77, 1598–1606. [Google Scholar] [CrossRef]
- Koshy, A. Evolving Global Etiology of Hepatocellular Carcinoma (HCC): Insights and Trends for 2024. J. Clin. Exp. Hepatol. 2025, 15, 102406. [Google Scholar] [CrossRef]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nature reviews. Dis. Primers 2021, 7, 6. [Google Scholar] [CrossRef]
- Dong, M.; Ming, X.; Xiang, T.; Feng, N.; Zhang, M.; Ye, X.; He, Y.; Zhou, M.; Wu, Q. Recent research on the physicochemical properties and biological activities of quinones and their practical applications: A comprehensive review. Food Funct. 2024, 15, 8973–8997. [Google Scholar] [CrossRef] [PubMed]
- Schieber, A. Reactions of Quinones-Mechanisms, Structures, and Prospects for Food Research. J. Agric. Food Chem. 2018, 66, 13051–13055. [Google Scholar] [CrossRef] [PubMed]
- Lemos, T.L.; Monte, F.J.; Santos, A.K.; Fonseca, A.M.; Santos, H.S.; Oliveira, M.F.; Costa, S.M.; Pessoa, O.D.; Braz-Filho, R. Quinones from plants of northeastern Brazil: Structural diversity, chemical transformations, NMR data and biological activities. Nat. Prod. Res. 2007, 21, 529–550. [Google Scholar] [CrossRef]
- Gomes de Carvalho, N.K.; Wellisson da Silva Mendes, J.; Martins da Costa, J.G. Quinones: Biosynthesis, Characterization of (13) C Spectroscopical Data and Pharmacological Activities. Chem. Biodivers. 2023, 20, e202301365. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ding, X.; Feng, S.X.; Guan, Q.; Zhang, X.P.; Du, C.; Di, Y.T.; Chen, T. Seven New Tetrahydroanthraquinones from the Root of Prismatomeris connata and Their Cytotoxicity against Lung Tumor Cell Growth. Molecules 2015, 20, 22565–22577. [Google Scholar] [CrossRef]
- Tian, W.; Wang, C.; Li, D.; Hou, H. Novel anthraquinone compounds as anticancer agents and their potential mechanism. Future Med. Chem. 2020, 12, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.L.; Chang, F.R.; Yen, M.H.; Yu, D.; Liu, Y.N.; Bastow, K.F.; Morris-Natschke, S.L.; Wu, Y.C.; Lee, K.H. Cytotoxic phenanthrenequinones and 9,10-dihydrophenanthrenes from Calanthe arisanensis. J. Nat. Prod. 2009, 72, 210–213. [Google Scholar] [CrossRef]
- Kang, J.; Zhang, P.; Gao, Z.; Zhang, J.; Yan, Z.; Wang, H.; Chen, R. Naphthohydroquinones, naphthoquinones, anthraquinones, and a naphthohydroquinone dimer isolated from the aerial parts of Morinda parvifolia and their cytotoxic effects through up-regulation of p53. Phytochemistry 2016, 130, 144–151. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, J.; Yu, Z.; Zhai, F.; Li, H.; Jin, X. Regulation of apoptosis by ubiquitination in liver cancer. Am. J. Cancer Res. 2023, 13, 4832–4871. [Google Scholar]
- Liu, Y.; Hao, C.; Li, L.; Zhang, H.; Zha, W.; Ma, L.; Chen, L.; Gan, J. The Role of Oxidative Stress in the Development and Therapeutic Intervention of Hepatocellular Carcinoma. Curr. Cancer Drug Targets 2023, 23, 792–804. [Google Scholar] [CrossRef]
- Giannelli, G.; Koudelkova, P.; Dituri, F.; Mikulits, W. Role of epithelial to mesenchymal transition in hepatocellular carcinoma. J. Hepatol. 2016, 65, 798–808. [Google Scholar] [CrossRef]
- Wang, C.; Wang, H.; Wang, C.; Tian, T.; Jin, A.; Liu, Y.; Huo, R.; Liu, T.; Pan, B.; Guo, W.; et al. Plumbagin Triggers Cuproptosis in Hepatocellular Carcinoma (HCC) via the DNA-Methyltransferase 1 (DNMT1)/microRNA-302a-3p (miR-302a-3p)/ATPase Copper Transporting Beta (ATP7B) Axis. MedComm 2025, 6, e70312. [Google Scholar] [PubMed]
- Qin, B.; Zeng, Z.; Xu, J.; Shangwen, J.; Ye, Z.J.; Wang, S.; Wu, Y.; Peng, G.; Wang, Q.; Gu, W.; et al. Emodin inhibits invasion and migration of hepatocellular carcinoma cells via regulating autophagy-mediated degradation of snail and β-catenin. BMC Cancer 2022, 22, 671. [Google Scholar] [CrossRef]
- Hassan, H.M.; Hamdan, A.M.; Alattar, A.; Alshaman, R.; Bahattab, O.; Al-Gayyar, M.M.H. Evaluating anticancer activity of emodin by enhancing antioxidant activities and affecting PKC/ADAMTS4 pathway in thioacetamide-induced hepatocellular carcinoma in rats. Redox Rep. Commun. Free Radic. Res. 2024, 29, 2365590. [Google Scholar]
- Kim, Y.S.; Lee, Y.M.; Oh, T.I.; Shin, D.H.; Kim, G.H.; Kan, S.Y.; Kang, H.; Kim, J.H.; Kim, B.M.; Yim, W.J.; et al. Emodin Sensitizes Hepatocellular Carcinoma Cells to the Anti-Cancer Effect of Sorafenib through Suppression of Cholesterol Metabolism. Int. J. Mol. Sci. 2018, 19, 3127. [Google Scholar] [CrossRef]
- Gentilin, E.; Simoni, E.; Candito, M.; Cazzador, D.; Astolfi, L. Cisplatin-Induced Ototoxicity: Updates on Molecular Targets. Trends Mol. Med. 2019, 25, 1123–1132. [Google Scholar] [CrossRef]
- Giacomini, I.; Ragazzi, E.; Pasut, G.; Montopoli, M. The Pentose Phosphate Pathway and Its Involvement in Cisplatin Resistance. Int. J. Mol. Sci. 2020, 21, 937. [Google Scholar] [CrossRef]
- Sheng, J.; Shen, L.; Sun, L.; Zhang, X.; Cui, R.; Wang, L. Inhibition of PI3K/mTOR increased the sensitivity of hepatocellular carcinoma cells to cisplatin via interference with mitochondrial-lysosomal crosstalk. Cell Prolif. 2019, 52, e12609. [Google Scholar] [CrossRef]
- Yang, M.; Xiong, Z.; Deng, H.; Chen, X.; Lai, Q.; Wang, H.; Leng, Y. Effect of emodin combined with cisplatin on the invasion and migration of HepG2 hepatoma cells. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2023, 74, 403–411. [Google Scholar]
- Wang, F.; Yao, X.; Zhang, Y.; Tang, J. Synthesis, biological function and evaluation of Shikonin in cancer therapy. Fitoterapia 2019, 134, 329–339. [Google Scholar] [CrossRef]
- Liu, B.; Jin, J.; Zhang, Z.; Zuo, L.; Jiang, M.; Xie, C. Shikonin exerts antitumor activity by causing mitochondrial dysfunction in hepatocellular carcinoma through PKM2-AMPK-PGC1α signaling pathway. Biochem. Cell Biol. 2019, 97, 397–405. [Google Scholar] [CrossRef]
- Yang, W.; Liu, J.; Hou, L.; Chen, Q.; Liu, Y. Shikonin differentially regulates glucose metabolism via PKM2 and HIF1α to overcome apoptosis in a refractory HCC cell line. Life Sci. 2021, 265, 118796. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, W.; Jin, L.; Liu, S.; Liang, L.; Wei, Y. Plumbagin Exhibits Genotoxicity and Induces G2/M Cell Cycle Arrest via ROS-Mediated Oxidative Stress and Activation of ATM-p53 Signaling Pathway in Hepatocellular Cells. Int. J. Mol. Sci. 2023, 24, 6279. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Yan, D.; Jiang, B.; Xue, Q.; Chen, X.; Huang, Q.; Qi, L.; Tang, D.; Chen, X.; Liu, J. Plumbagin is a novel GPX4 protein degrader that induces apoptosis in hepatocellular carcinoma cells. Free Radic. Biol. Med. 2023, 203, 1–10. [Google Scholar] [CrossRef]
- Du, Y.Q.; Yuan, B.; Ye, Y.X.; Zhou, F.L.; Liu, H.; Huang, J.J.; Wei, Y.F. Plumbagin Regulates Snail to Inhibit Hepatocellular Carcinoma Epithelial-Mesenchymal Transition in vivo and in vitro. J. Hepatocell. Carcinoma 2024, 11, 565–580. [Google Scholar] [CrossRef]
- Cao, C.; Li, Y.; Shi, F.; Jiang, S.; Li, Y.; Yang, L.; Zhou, X.; Gao, Y.; Tang, F.; Li, H.; et al. Nano co-delivery of doxorubicin and plumbagin achieves synergistic chemotherapy of hepatocellular carcinoma. Int. J. Pharm. 2024, 661, 124424. [Google Scholar] [CrossRef]
- Bai, Y.; Wen, H.; Lin, J.; Liu, X.; Yu, H.; Wu, M.; Wang, L.; Chen, D. Tanshinone I improves renal fibrosis by promoting gluconeogenesis through upregulation of peroxisome proliferator-activated receptor-γ coactivator 1α. Ren. Fail. 2024, 46, 2433710. [Google Scholar] [CrossRef]
- Huang, X.; Deng, H.; Shen, Q.K.; Quan, Z.S. Tanshinone IIA: Pharmacology, Total Synthesis, and Progress in Structure-modifications. Curr. Med. Chem. 2022, 29, 1959–1989. [Google Scholar] [CrossRef]
- Liu, X.; Liu, J. Tanshinone I induces cell apoptosis by reactive oxygen species-mediated endoplasmic reticulum stress and by suppressing p53/DRAM-mediated autophagy in human hepatocellular carcinoma. Artif. Cells Nanomed. Biotechnol. 2020, 48, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Feng, N.; Geng, A. Tanshinone I suppresses hepatocellular carcinoma cells growth through targeting DNA double-strand break repair. Cancer Biol. Ther. 2023, 24, 2229958. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Zhang, X.; Li, Y.; Zhou, H.; Wang, H.; Wang, F.; Ma, H.; Cao, L. Identification of the molecular mechanisms of Salvia miltiorrhiza relevant to the treatment of osteoarthritis based on network pharmacology. Discov. Med. 2020, 30, 83–95. [Google Scholar] [PubMed]
- Zhao, J.; Lin, E.; Cai, C.; Zhang, M.; Li, D.; Cai, S.; Zeng, G.; Yin, Z.; Wang, B.; Li, P.; et al. Combined Treatment of Tanshinone I and Epirubicin Revealed Enhanced Inhibition of Hepatocellular Carcinoma by Targeting PI3K/AKT/HIF-1α. Drug Des. Dev. Ther. 2022, 16, 3197–3213. [Google Scholar] [CrossRef]
- Santoni, M.; Rizzo, A.; Mollica, V.; Matrana, M.R.; Rosellini, M.; Faloppi, L.; Marchetti, A.; Battelli, N.; Massari, F. The impact of gender on The efficacy of immune checkpoint inhibitors in cancer patients: The MOUSEION-01 study. Crit. Rev. Oncol./Hematol. 2022, 170, 103596. [Google Scholar] [CrossRef]
- Wang, W.Q.; Liu, L.; Sun, H.C.; Fu, Y.L.; Xu, H.X.; Chai, Z.T.; Zhang, Q.B.; Kong, L.Q.; Zhu, X.D.; Lu, L.; et al. Tanshinone IIA inhibits metastasis after palliative resection of hepatocellular carcinoma and prolongs survival in part via vascular normalization. J. Hematol. Oncol. 2012, 5, 69. [Google Scholar] [CrossRef]
- Mao, D.; Wang, H.; Guo, H.; Che, X.; Chen, M.; Li, X.; Liu, Y.; Huo, J.; Chen, Y. Tanshinone IIA normalized hepatocellular carcinoma vessels and enhanced PD-1 inhibitor efficacy by inhibiting ELTD1. Phytomed. Int. J. Phytother. Phytopharm. 2024, 123, 155191. [Google Scholar] [CrossRef]
- Chiu, C.M.; Huang, S.Y.; Chang, S.F.; Liao, K.F.; Chiu, S.C. Synergistic antitumor effects of tanshinone IIA and sorafenib or its derivative SC-1 in hepatocellular carcinoma cells. OncoTargets Ther. 2018, 11, 1777–1785. [Google Scholar] [CrossRef]
- Mohammed, N.K.; Abd Manap, M.Y.; Tan, C.P.; Muhialdin, B.J.; Alhelli, A.M.; Meor Hussin, A.S. The Effects of Different Extraction Methods on Antioxidant Properties, Chemical Composition, and Thermal Behavior of Black Seed (Nigella sativa L.) Oil. Evid.-Based Complement. Altern. Med. eCAM 2016, 2016, 6273817. [Google Scholar] [CrossRef] [PubMed]
- Entok, E.; Ustuner, M.C.; Ozbayer, C.; Tekin, N.; Akyuz, F.; Yangi, B.; Kurt, H.; Degirmenci, I.; Gunes, H.V. Anti-inflammatuar and anti-oxidative effects of Nigella sativa L.: 18FDG-PET imaging of inflammation. Mol. Biol. Rep. 2014, 41, 2827–2834. [Google Scholar] [CrossRef]
- Ibrahim, S.; Fahim, S.A.; Tadros, S.A.; Badary, O.A. Suppressive effects of thymoquinone on the initiation stage of diethylnitrosamine hepatocarcinogenesis in rats. J. Biochem. Mol. Toxicol. 2022, 36, e23078. [Google Scholar] [CrossRef]
- Tadros, S.A.; Attia, Y.M.; Maurice, N.W.; Fahim, S.A.; Abdelwahed, F.M.; Ibrahim, S.; Badary, O.A. Thymoquinone Suppresses Angiogenesis in DEN-Induced Hepatocellular Carcinoma by Targeting miR-1-3p. Int. J. Mol. Sci. 2022, 23, 15904. [Google Scholar] [CrossRef]
- Farghaly, M.E.; Khowailed, A.A.; Aboulhoda, B.E.; Rashed, L.A.; Gaber, S.S.; Ashour, H. Thymoquinone Potentiated the Anticancer Effect of Cisplatin on Hepatic Tumorigenesis by Modulating Tissue Oxidative Stress and Endoplasmic GRP78/CHOP Signaling. Nutr. Cancer 2022, 74, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Hou, B.S.; Wang, H.F.; XU, H.J.; Tan, Z.X.; Zhan, W.Y.; Song, Y.Q.; Zhang, Y.M.; Liu, X. Effect of Rhubarb Aphid Fanghe Xiaoyaofang Decoction on Postoperative TACE Syndrome of Primary Hepatocellular Carcinoma. Med. Inf. 2018, 31, 53–56. [Google Scholar]
- Tian, D.X.; Liu, W. Effects of Dahuang Zhechong Pill on liver function and immune function after interventional therapy for liver cancer. Chin. J. Mod. Drug Appl. 2022, 16, 11–14. [Google Scholar]
- Deng, Z.F.; Zhao, C. Clinical Observation of Rheum officinale Soda Tablets for treating advanced primary liver cancer. J. Pract. Med. 2007, 11, 1750–1751. [Google Scholar]
- Zhu, Y.L.; Yi, F.T. The effect of γ knife the combined application of danshen injections on immune function in patients with primary liver cancer. Chin. J. Integr. Tradit. West. Med. Liver Dis. 2020, 30, 111–113. [Google Scholar]
- Zhang, Q.H.; Shao, X.W. The clinical efficacy of compound Danshen injection in liver cancer patients and its impact on T cell subsets. Chin. J. Integr. Tradit. West. Med. Liver Dis. 2004, 3, 184. [Google Scholar]
- Semwal, R.B.; Semwal, D.K.; Combrinck, S.; Viljoen, A. Emodin—A natural anthraquinone derivative with diverse pharmacological activities. Phytochemistry 2021, 190, 112854. [Google Scholar] [CrossRef]
- Oshida, K.; Hirakata, M.; Maeda, A.; Miyoshi, T.; Miyamoto, Y. Toxicological effect of emodin in mouse testicular gene expression profile. J. Appl. Toxicol. JAT 2011, 31, 790–800. [Google Scholar] [CrossRef]
- He, Q.; Liu, K.; Wang, S.; Hou, H.; Yuan, Y.; Wang, X. Toxicity induced by emodin on zebrafish embryos. Drug Chem. Toxicol. 2012, 35, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.H.; Chang, S.C.; Chan, W.H. Injurious effects of emodin on maturation of mouse oocytes, fertilization and fetal development via apoptosis. Int. J. Mol. Sci. 2012, 13, 13911–13925. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Tang, S.; Chen, A.; Zhang, Y.; Liu, M.; Wang, X. Evaluation of the inhibition risk of shikonin on human and rat UDP-glucuronosyltransferases (UGT) through the cocktail approach. Toxicol. Lett. 2019, 312, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Figat, R.; Zgadzaj, A.; Geschke, S.; Sieczka, P.; Pietrosiuk, A.; Sommer, S.; Skrzypczak, A. Cytotoxicity and antigenotoxicity evaluation of acetylshikonin and shikonin. Drug Chem. Toxicol. 2021, 44, 140–147. [Google Scholar] [CrossRef]
- Tang, S.; Chen, A.; Zhou, X.; Zeng, L.; Liu, M.; Wang, X. Assessment of the inhibition risk of shikonin on cytochrome P450 via cocktail inhibition assay. Toxicol. Lett. 2017, 281, 74–83. [Google Scholar] [CrossRef]
- Thakor, N.; Janathia, B. Plumbagin: A Potential Candidate for Future Research and Development. Curr. Pharm. Biotechnol. 2022, 23, 1800–1812. [Google Scholar] [CrossRef]
- Kumar, D.; Patil, P.A.; Roy, S.; Kholkute, S.D.; Hegde, H.V.; Nair, V. Comparative toxicity profiles of Plumbago zeylanica L. root petroleum ether, acetone and hydroalcoholic extracts in Wistar rats. AYU 2015, 36, 329–334. [Google Scholar]
- Lim, Y.; Kim, C.; Kim, D.; Kim, M.J.; Yu, J.W.; Song, M.H.; Kim, Y.; Son, J.; Lee, J.H.; Lee, S.E. Plumbagin, a natural derivative of 1,4-naphthoquinone, induces cyclopic phenomenon via increased apoptosis and ROS generation in the early stage of zebrafish embryos. Ecotoxicol. Environ. Saf. 2025, 302, 118535. [Google Scholar] [CrossRef] [PubMed]
- AbuKhader, M.M.; Khater, S.H.; Al-Matubsi, H.Y. Acute effects of thymoquinone on the pregnant rat and embryo-fetal development. Drug Chem. Toxicol. 2013, 36, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Ke, L.; Zhong, C.; Chen, Z.; Zheng, Z.; Li, S.; Chen, B.; Wu, Q.; Yao, H. Tanshinone I: Pharmacological activities, molecular mechanisms against diseases and future perspectives. Phytomed. Int. J. Phytother. Phytopharm. 2023, 110, 154632. [Google Scholar]
- Huang, Y.; Yu, S.H.; Zhen, W.X.; Cheng, T.; Wang, D.; Lin, J.B.; Wu, Y.H.; Wang, Y.F.; Chen, Y.; Shu, L.P.; et al. Tanshinone I, a new EZH2 inhibitor restricts normal and malignant hematopoiesis through upregulation of MMP9 and ABCG2. Theranostics 2021, 11, 6891–6904. [Google Scholar] [CrossRef]
- Wang, T.; Wang, C.; Wu, Q.; Zheng, K.; Chen, J.; Lan, Y.; Qin, Y.; Mei, W.; Wang, B. Evaluation of Tanshinone IIA Developmental Toxicity in Zebrafish Embryos. Molecules 2017, 22, 660. [Google Scholar] [CrossRef]
- Mattioli, R.; Ilari, A.; Colotti, B.; Mosca, L.; Fazi, F.; Colotti, G. Doxorubicin and other anthracyclines in cancers: Activity, chemoresistance and its overcoming. Mol. Asp. Med. 2023, 93, 101205. [Google Scholar] [CrossRef]
- Tam, K. The Roles of Doxorubicin in Hepatocellular Carcinoma. ADMET DMPK 2013, 1, 29–44. [Google Scholar] [CrossRef]
- Golla, K.; Bhaskar, C.; Ahmed, F.; Kondapi, A.K. A target-specific oral formulation of Doxorubicin-protein nanoparticles: Efficacy and safety in hepatocellular cancer. J. Cancer 2013, 4, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Saddik, M.S.; Elsayed, M.M.A.; Abdel-Rheem, A.A.; El-Mokhtar, M.A.; Mosa, E.S.; Al-Hakkani, M.F.; Al-Shelkamy, S.A.; Khames, A.; Daha, M.A.; Abdel-Aleem, J.A. A Novel C@Fe@Cu Nanocomposite Loaded with Doxorubicin Tailored for the Treatment of Hepatocellular Carcinoma. Pharmaceutics 2022, 14, 1845. [Google Scholar] [CrossRef]
- Chen, L.; Alrbyawi, H.; Poudel, I.; Arnold, R.D.; Babu, R.J. Co-delivery of Doxorubicin and Ceramide in a Liposomal Formulation Enhances Cytotoxicity in Murine B16BL6 Melanoma Cell Lines. AAPS PharmSciTech 2019, 20, 99. [Google Scholar] [CrossRef]
- Sun, W.; Wang, Y.; Cai, M.; Lin, L.; Chen, X.; Cao, Z.; Zhu, K.; Shuai, X. Codelivery of sorafenib and GPC3 siRNA with PEI-modified liposomes for hepatoma therapy. Biomater. Sci. 2017, 5, 2468–2479. [Google Scholar] [CrossRef]
- Li, X.; Diao, W.; Xue, H.; Wu, F.; Wang, W.; Jiang, B.; Bai, J.; Lian, B.; Feng, W.; Sun, T.; et al. Improved efficacy of doxorubicin delivery by a novel dual-ligand-modified liposome in hepatocellular carcinoma. Cancer Lett. 2020, 489, 163–173. [Google Scholar]
- Zhao, R.; Cheng, S.; Bai, X.; Zhang, D.; Fang, H.; Che, W.; Zhang, W.; Zhou, Y.; Duan, W.; Liang, Q.; et al. Development of an efficient liposomal DOX delivery formulation for HCC therapy by targeting CK2α. Biotechnol. J. 2024, 19, e2400050. [Google Scholar] [CrossRef] [PubMed]
- Fox, E.J. Mechanism of action of mitoxantrone. Neurology 2004, 63 (Suppl. S6), S15–S18. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Okusaka, T.; Sato, Y.; Furuse, J.; Mitsunaga, S.; Ueno, H.; Morizane, C.; Inaba, Y.; Kobayashi, T.; Arai, Y. A Phase I/II trial of continuous hepatic intra-arterial infusion of 5-fluorouracil, mitoxantrone and cisplatin for advanced hepatocellular carcinoma. Jpn. J. Clin. Oncol. 2017, 47, 512–519. [Google Scholar] [PubMed]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in mammalian autophagy research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef]
- Xie, B.; He, X.; Guo, G.; Zhang, X.; Li, J.; Liu, J.; Lin, Y. High-throughput screening identified mitoxantrone to induce death of hepatocellular carcinoma cells with autophagy involvement. Biochem. Biophys. Res. Commun. 2020, 521, 232–237. [Google Scholar] [CrossRef]
- Arcamone, F.; Bernardi, L.; Giardino, P.; Patelli, B.; Marco, A.; Casazza, A.M.; Pratesi, G.; Reggiani, P. Synthesis and antitumor activity of 4-demethoxydaunorubicin, 4-demethoxy-7,9-diepidaunorubicin, and their beta anomers. Cancer Treat. Rep. 1976, 60, 829–834. [Google Scholar]
- Goebel, M. Oral idarubicin--an anthracycline derivative with unique properties. Ann. Hematol. 1993, 66, 33–43. [Google Scholar] [CrossRef]
- Zheng, Z.; Ma, M.; Han, X.; Li, X.; Huang, J.; Zhao, Y.; Liu, H.; Kang, J.; Kong, X.; Sun, G.; et al. Idarubicin-loaded biodegradable microspheres enhance sensitivity to anti-PD1 immunotherapy in transcatheter arterial chemoembolization of hepatocellular carcinoma. Acta Biomater. 2023, 157, 337–351. [Google Scholar] [CrossRef]
- Lai, C.L.; Wu, P.C.; Chan, G.C.; Lok, A.S.; Lin, H.J. Doxorubicin versus no antitumor therapy in inoperable hepatocellular carcinoma. A prospective randomized trial. Cancer 1988, 62, 479–483. [Google Scholar] [CrossRef]
- Valle, J.W.; Dangoor, A.; Beech, J.; Sherlock, D.J.; Lee, S.M.; Scarffe, J.H.; Swindell, R.; Ranson, M. Treatment of inoperable hepatocellular carcinoma with pegylated liposomal doxorubicin (PLD): Results of a phase II study. Br. J. Cancer 2005, 92, 628–630. [Google Scholar] [CrossRef]
- Tak, W.Y.; Lin, S.M.; Wang, Y.; Zheng, J.; Vecchione, A.; Park, S.Y.; Chen, M.H.; Wong, S.; Xu, R.; Peng, C.Y.; et al. Phase III HEAT Study Adding Lyso-Thermosensitive Liposomal Doxorubicin to Radiofrequency Ablation in Patients with Unresectable Hepatocellular Carcinoma Lesions. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 73–83. [Google Scholar] [CrossRef]
- Shepherd, F.A.; Evans, W.K.; Blackstein, M.E.; Fine, S.; Heathcote, J.; Langer, B.; Taylor, B.; Habal, F.; Kutas, G.; Pritchard, K.I.; et al. Hepatic arterial infusion of mitoxantrone in the treatment of primary hepatocellular carcinoma. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 1987, 5, 635–640. [Google Scholar]
- Farrés, M.T.; de Baere, T.; Lagrange, C.; Ramirez, L.; Rougier, P.; Munck, J.N.; Roche, A. Percutaneous mitoxantrone injection for primary and secondary liver tumors: Preliminary results. Cardiovasc. Interv. Radiol. 1998, 21, 399–403. [Google Scholar] [CrossRef]
- Zhou, Q.; Sun, X.; Zeng, L.; Liu, J.; Zhang, Z. A randomized multicenter phase II clinical trial of mitoxantrone-loaded nanoparticles in the treatment of 108 patients with unresected hepatocellular carcinoma. Nanomed. Nanotechnol. Biol. Med. 2009, 5, 419–423. [Google Scholar]
- Mitomycin. LiverTox: Clinical and Research Information on Drug-Induced Liver Injury; National Institute of Diabetes and Digestive and Kidney Diseases: Bethesda, MD, USA, 2012.
- Cheirsilpa, A.; Leelasethakul, S.; Auethaveekiat, V.; Maoleekulpriroj, S.; Kangsumrit, N.; Thanakaravit, P.; Phanthumjida, P. High-dose mitomycin C: Activity in hepatocellular carcinoma. Cancer Chemother. Pharmacol. 1989, 24, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Boulin, M.; Guiu, S.; Chauffert, B.; Aho, S.; Cercueil, J.P.; Ghiringhelli, F.; Krause, D.; Fagnoni, P.; Hillon, P.; Bedenne, L.; et al. Screening of anticancer drugs for chemoembolization of hepatocellular carcinoma. Anti-Cancer Drugs 2011, 22, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Boulin, M.; Hillon, P.; Cercueil, J.P.; Bonnetain, F.; Dabakuyo, S.; Minello, A.; Jouve, J.L.; Lepage, C.; Bardou, M.; Wendremaire, M.; et al. Idarubicin-loaded beads for chemoembolisation of hepatocellular carcinoma: Results of the IDASPHERE phase I trial. Aliment. Pharmacol. Ther. 2014, 39, 1301–1313. [Google Scholar] [CrossRef]
- Roth, G.S.; Teyssier, Y.; Abousalihac, M.; Seigneurin, A.; Ghelfi, J.; Sengel, C.; Decaens, T. Idarubicin vs doxorubicin in transarterial chemoembolization of intermediate stage hepatocellular carcinoma. World J. Gastroenterol. 2020, 26, 324–334. [Google Scholar] [CrossRef]
- Carvalho, C.; Santos, R.X.; Cardoso, S.; Correia, S.; Oliveira, P.J.; Santos, M.S.; Moreira, P.I. Doxorubicin: The good, the bad and the ugly effect. Curr. Med. Chem. 2009, 16, 3267–3285. [Google Scholar] [CrossRef]
- Sciarrino, E.; Simonetti, R.G.; Le Moli, S.; Pagliaro, L. Adriamycin treatment for hepatocellular carcinoma. Experience with 109 patients. Cancer 1985, 56, 2751–2755. [Google Scholar] [CrossRef]
- Colombo, M.; Tommasini, M.A.; Del Ninno, E.; Rumi, M.G.; De Fazio, C.; Dioguardi, M.L. Hepatocellular carcinoma in Italy: Report of a clinical trial with intravenous doxorubicin. Liver 1985, 5, 336–341. [Google Scholar]
- Dunk, A.A.; Scott, S.C.; Johnson, P.J.; Melia, W.; Lok, A.S.; Murray-Lyon, I.; Williams, R.; Thomas, H.C. Mitozantrone as single agent therapy in hepatocellular carcinoma. A phase II study. J. Hepatol. 1985, 1, 395–404. [Google Scholar] [CrossRef]
- Damiani, R.M.; Moura, D.J.; Viau, C.M.; Caceres, R.A.; Henriques, J.A.P.; Saffi, J. Pathways of cardiac toxicity: Comparison between chemotherapeutic drugs doxorubicin and mitoxantrone. Arch. Toxicol. 2016, 90, 2063–2076. [Google Scholar] [CrossRef]
- Verweij, J.; Pinedo, H.M. Mitomycin C: Mechanism of action, usefulness and limitations. Anti-Cancer Drugs 1990, 1, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, K.; Tsuchiya, S.; Nakayama, T.; Hiyama, Y.; Iwama, S.; Goto, N.; Takashi, M.; Ohtsuki, T.; Kono, K.; Nakajima, Y.; et al. Arterial chemoembolization of hepatocellular carcinoma with mitomycin C microcapsules. Radiology 1984, 152, 51–55. [Google Scholar] [CrossRef]
- Fields, S.M.; Koeller, J.M. Idarubicin: A second-generation anthracycline. DICP Ann. Pharmacother. 1991, 25, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Chen, Y.; Fan, X.; Han, J.; Zhong, L.; Zhang, Y.; Liu, Q.; Lin, J.; Huang, W.; Su, L.; et al. Emodin attenuates cardiomyocyte pyroptosis in doxorubicin-induced cardiotoxicity by directly binding to GSDMD. Phytomed. Int. J. Phytother. Phytopharm. 2023, 121, 155105. [Google Scholar] [CrossRef]
- Tuo, H.; Li, W.; Zhao, W.; Zhao, J.; Li, D.; Jin, L. Shikonin alleviates doxorubicin-induced cardiotoxicity via Mst1/Nrf2 pathway in mice. Sci. Rep. 2024, 14, 924. [Google Scholar] [CrossRef] [PubMed]
- Karabulut, D.; Ozturk, E.; Kaymak, E.; Akin, A.T.; Yakan, B. Thymoquinone attenuates doxorubicin-cardiotoxicity in rats. J. Biochem. Mol. Toxicol. 2021, 35, e22618. [Google Scholar] [CrossRef]
- Kaymak, E.; Öztürk, E.; Akİn, A.T.; Karabulut, D.; Yakan, B. Thymoquinone alleviates doxorubicin induced acute kidney injury by decreasing endoplasmic reticulum stress, inflammation and apoptosis. Biotech. Histochem. Off. Publ. Biol. Stain Comm. 2022, 97, 622–634. [Google Scholar] [CrossRef]
- Öztürk, E.; Kaymak, E.; Akin, A.T.; Karabulut, D.; Ünsal, H.M.; Yakan, B. Thymoquinone is a protective agent that reduces the negative effects of doxorubicin in rat testis. Hum. Exp. Toxicol. 2020, 39, 1364–1373. [Google Scholar] [CrossRef]
- Jiang, Q.; Chen, X.; Tian, X.; Zhang, J.; Xue, S.; Jiang, Y.; Liu, T.; Wang, X.; Sun, Q.; Hong, Y.; et al. Tanshinone I inhibits doxorubicin-induced cardiotoxicity by regulating Nrf2 signaling pathway. Phytomed. Int. J. Phytother. Phytopharm. 2022, 106, 154439. [Google Scholar] [CrossRef]
- Xu, L.; He, D.; Wu, Y.; Shen, L.; Wang, Y.; Xu, Y. Tanshinone IIA inhibits cardiomyocyte apoptosis and rescues cardiac function during doxorubicin-induced cardiotoxicity by activating the DAXX/MEK/ERK1/2 pathway. Phytomed. Int. J. Phytother. Phytopharm. 2022, 107, 154471. [Google Scholar] [CrossRef]
- Qiu, H.Y.; Wang, P.F.; Lin, H.Y.; Tang, C.Y.; Zhu, H.L.; Yang, Y.H. Naphthoquinones: A continuing source for discovery of therapeutic antineoplastic agents. Chem. Biol. Drug Des. 2018, 91, 681–690. [Google Scholar] [PubMed]
- Malik, S.; Singh, A.; Negi, P.; Kapoor, V.K. Thymoquinone: A small molecule from nature with high therapeutic potential. Drug Discov. Today 2021, 26, 2716–2725. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fan, W.; Li, H.; Qiao, L.; Liu, Z.; Zhu, B.; Guo, J.; Huang, K.; Tang, Y.; Wen, J.; et al. Idarubicin versus Epirubicin in Transarterial Chemoembolization for Barcelona Clinic Liver Cancer Stage B Hepatocellular Carcinoma: An Open-label, Randomized, Phase IV Trial. Radiology 2025, 315, e242315. [Google Scholar] [CrossRef]
- Zhao, C.; Yan, H.; Xiang, Z.; Wang, H.; Li, M.; Huang, M. Idarubicin versus epirubicin in drug-eluting beads-transarterial chemoembolization for treating hepatocellular carcinoma: A real-world retrospective study. Investig. New Drugs 2023, 41, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Kim, H.C.; Han, J.; Jang, M.J.; Chung, J.W. Transarterial Chemoembolization Using Idarubicin Versus Doxorubicin Chemoemulsion in Patients with Hepatocellular Carcinoma (IDADOX): Protocol for a Randomized, Non-inferiority, Double-Blind Trial. Cardiovasc. Interv. Radiol. 2024, 47, 372–378. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).