
Oxaliplatin-Induced Damage to the Gastric Innervation: Role in Nausea and Vomiting
 , ,
, ,  and
and 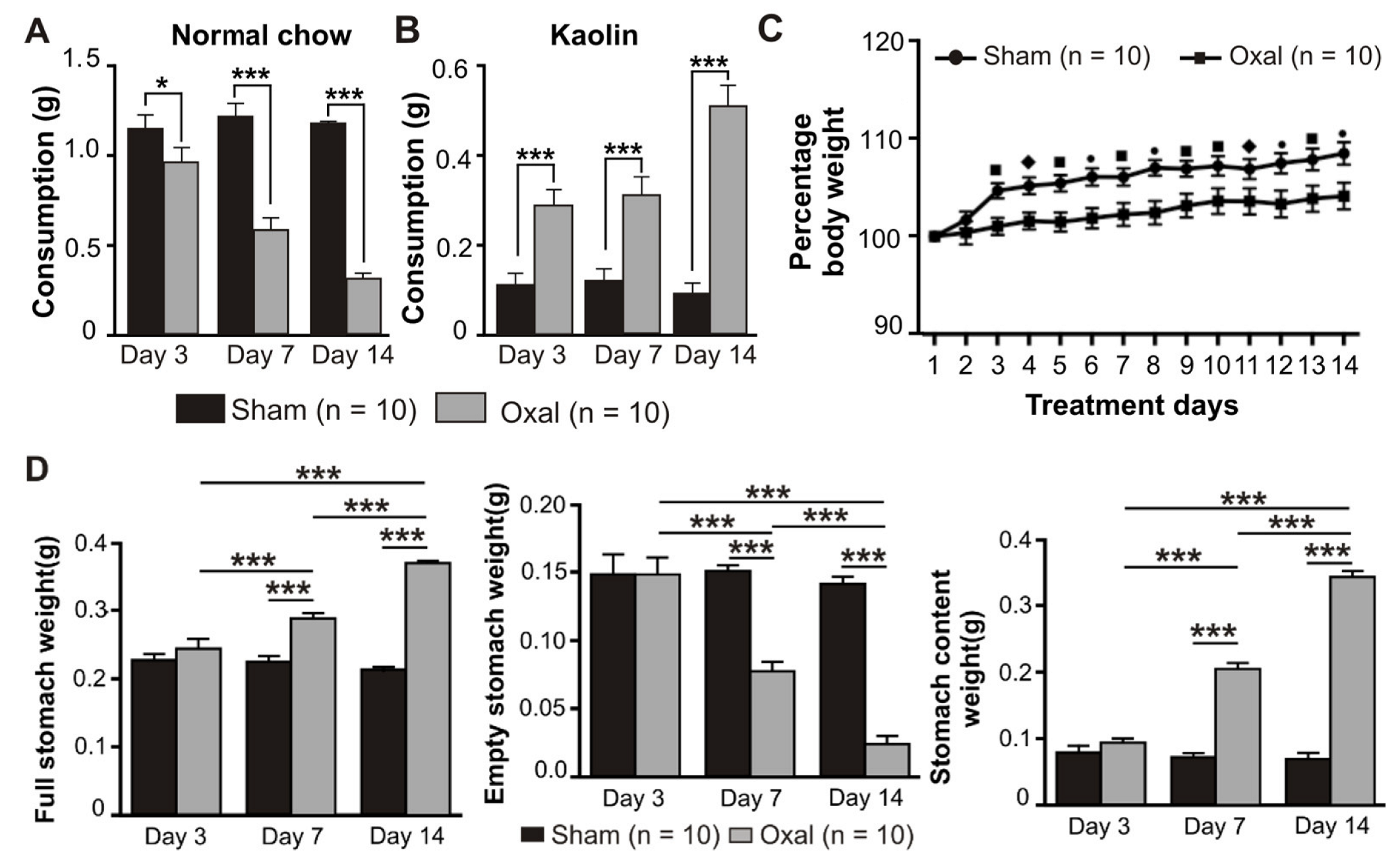{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. The Animals
2.2. In Vivo Oxaliplatin Treatment
2.3. Assessment of Feeding Behaviour: Pica
2.4. Stomach Contents
2.5. Tissue Preparation
2.6. Immunohistochemistry
2.7. Imaging
2.8. Quantitative Analysis of Immunoreactivity and Histology
2.9. The 5-HT Measurements
2.10. Electrophysiological Experiments
2.11. Statistical Analysis
3. Results
3.1. Effects of Oxaliplatin Treatment on Pica, Body Weight, and Stomach Content
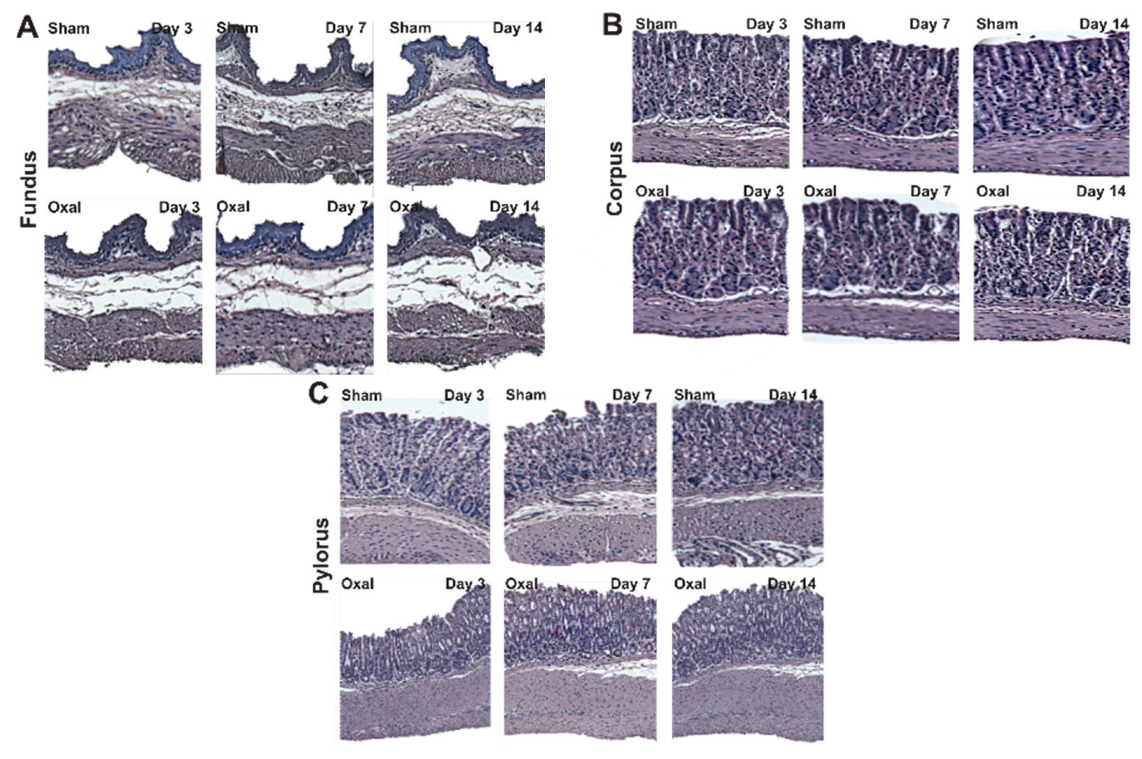
3.2. Effects of Oxaliplatin Treatment in the Morphology of the Stomach
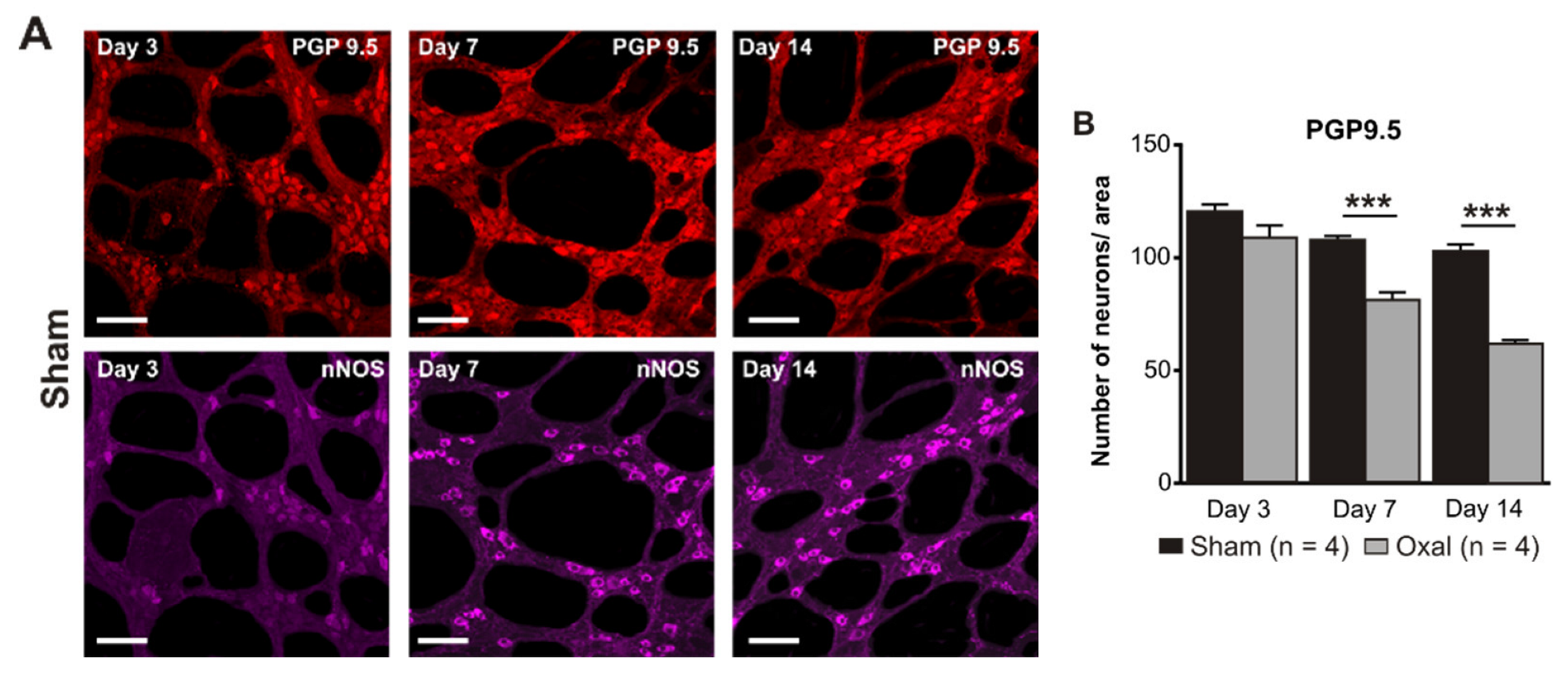
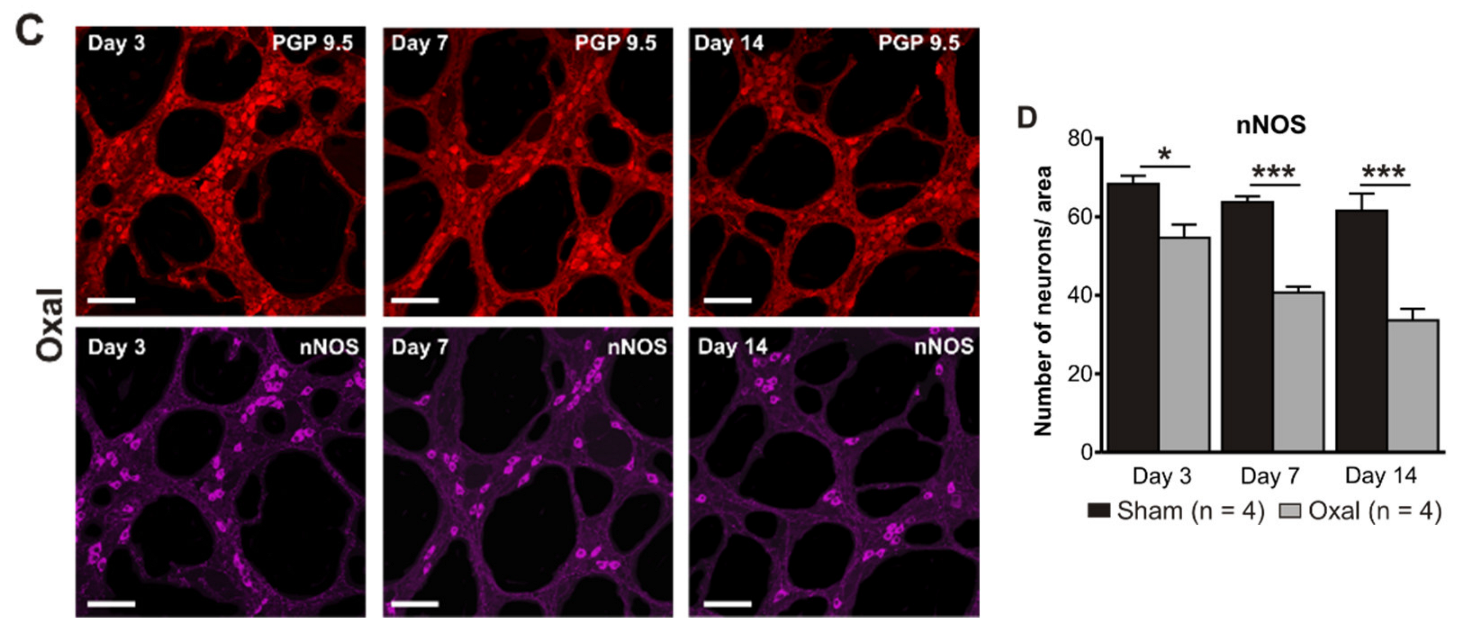
3.3. Effects of Oxaliplatin Treatment on the Myenteric Neurons and Subpopulation of nNOS-IR Neurons in the Stomach
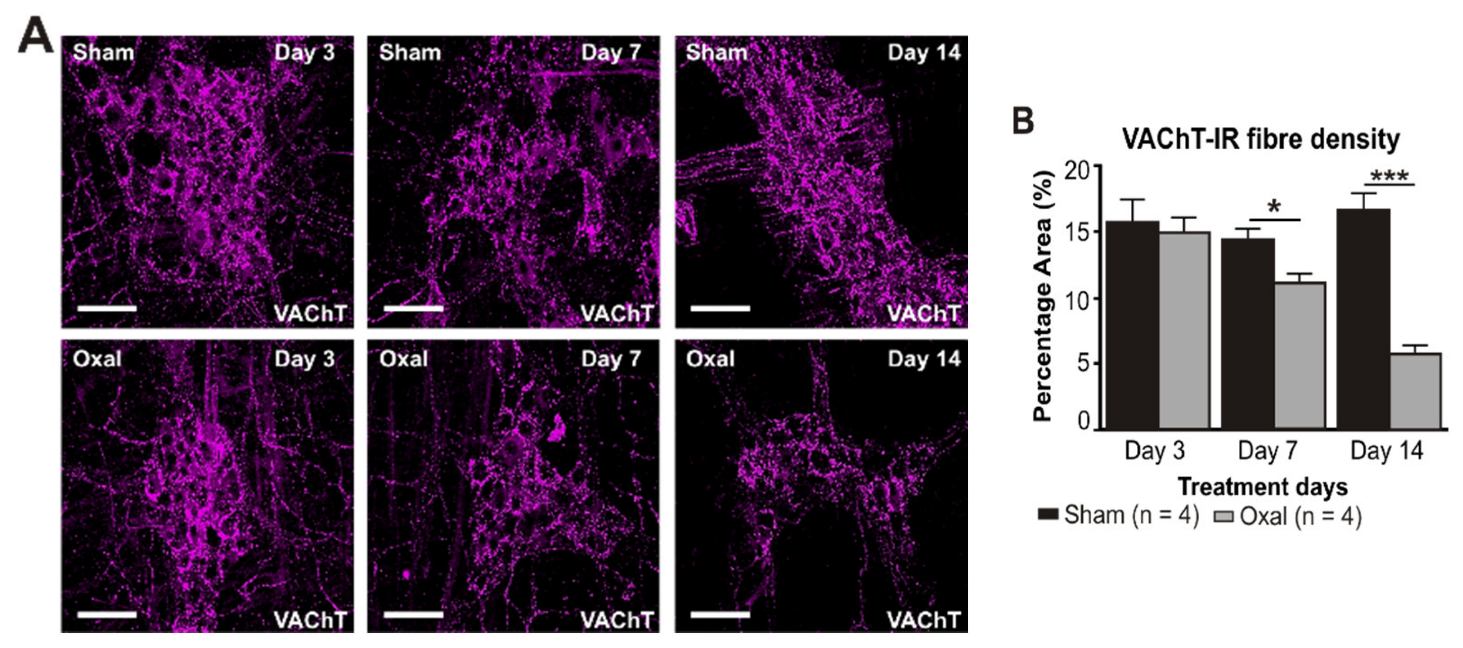
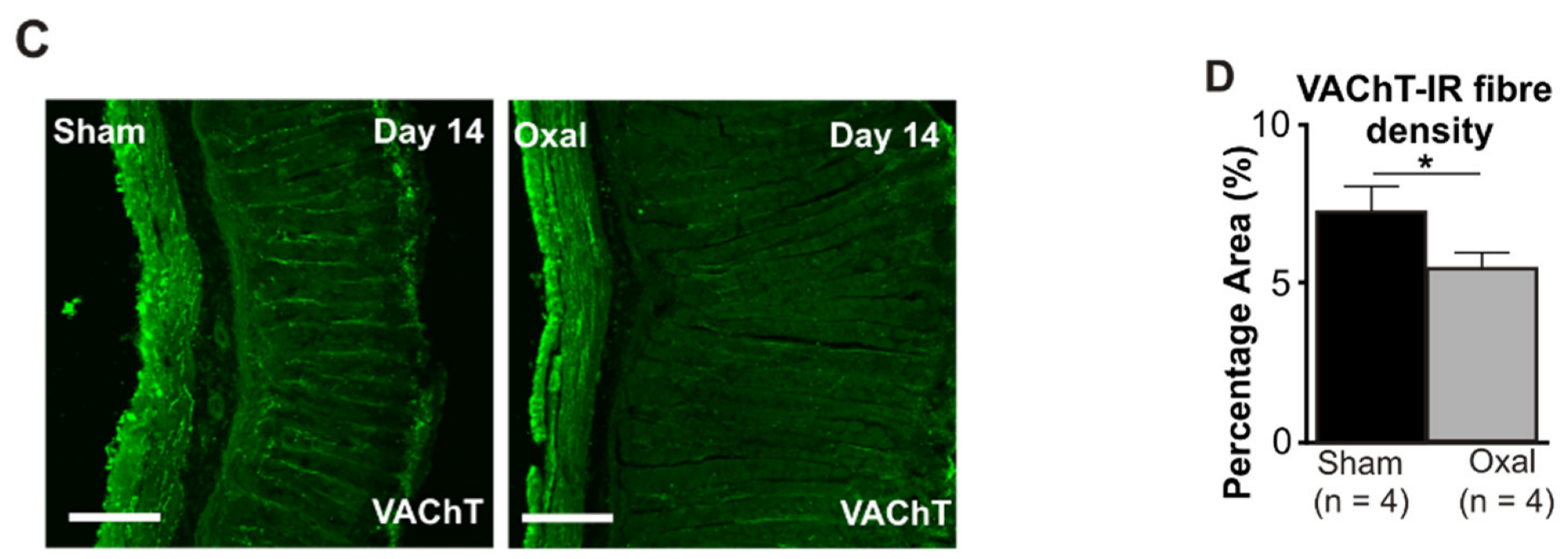
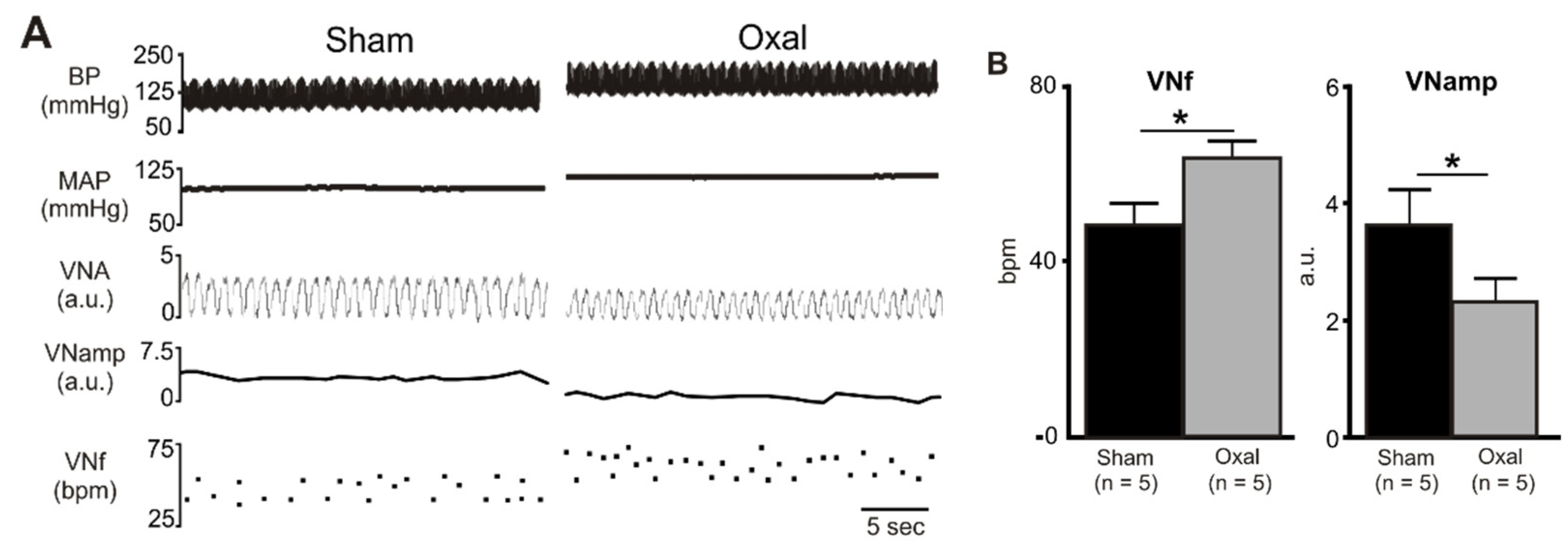
3.4. Effects of Oxaliplatin Treatment on the Expression of Cholinergic Fibres in the Stomach and Vagal Efferent Nerve Activity
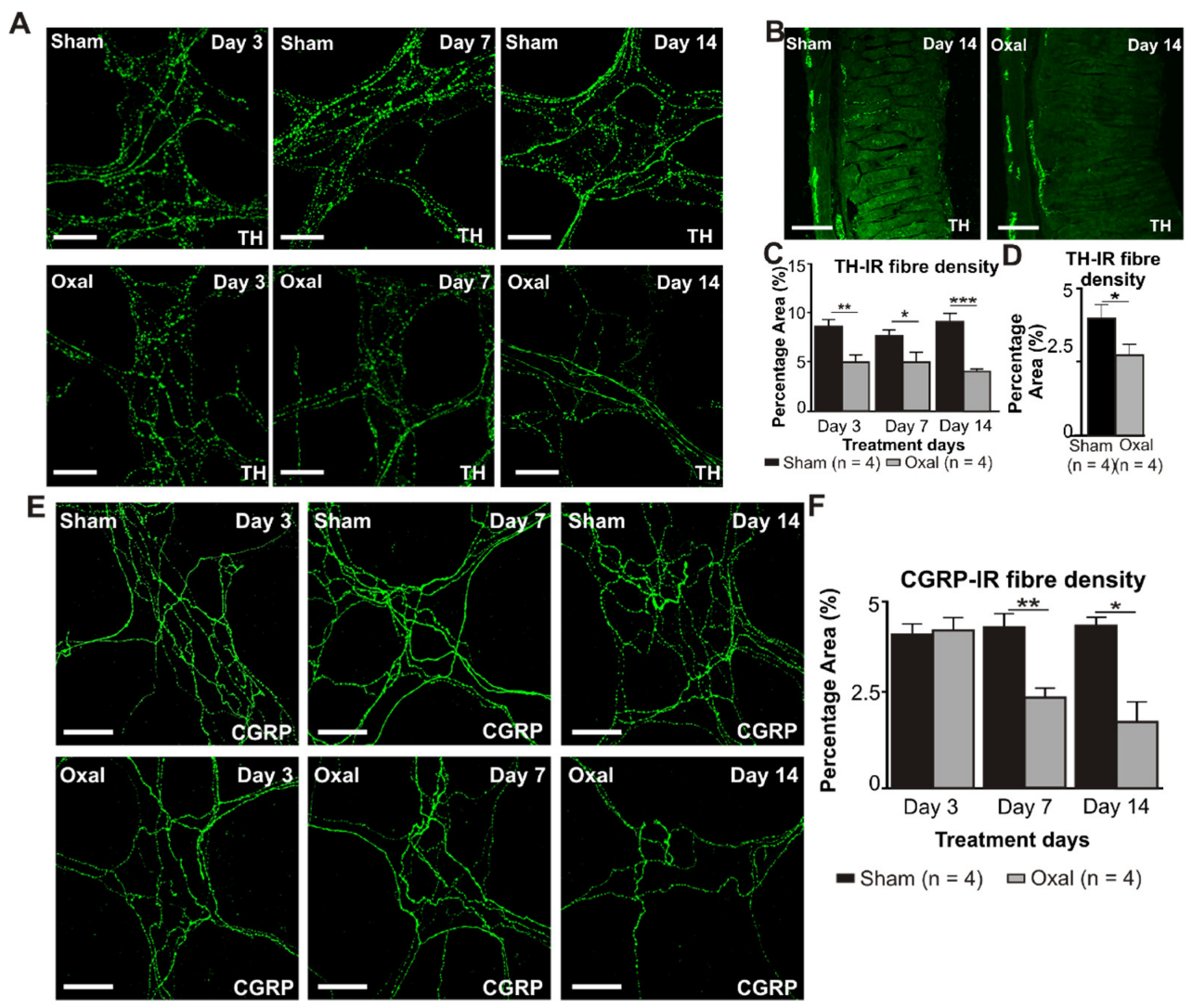
3.5. Effects of Oxaliplatin Treatment on Noradrenergic and Sensory Nerve Fibres in the Stomach
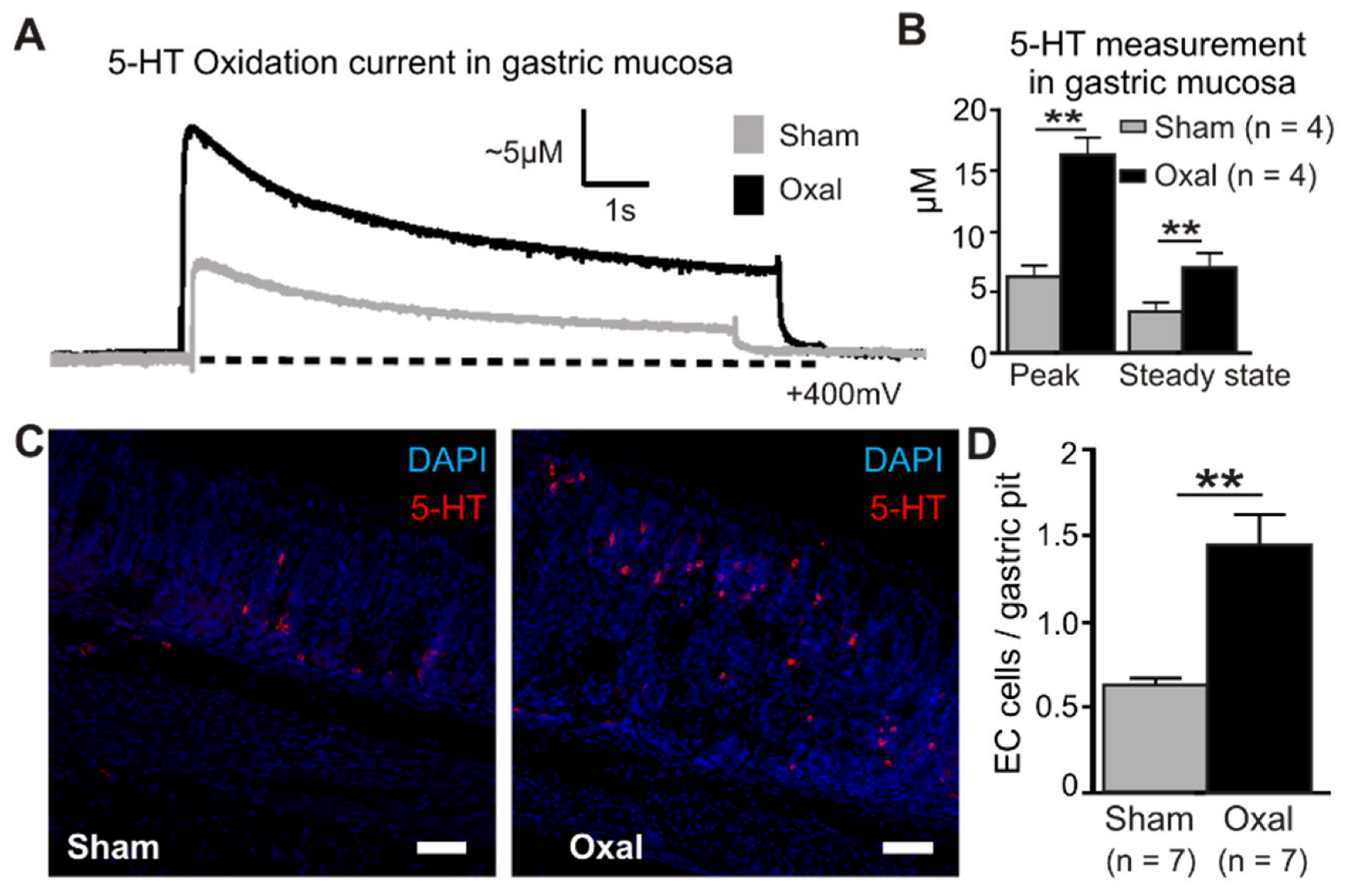
3.6. Effects of Oxaliplatin Treatment on Serotonin Level in the Stomach
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Labianca, R.; Beretta, G.D.; Kildani, B.; Milesi, L.; Merlin, F.; Mosconi, S.; Pessi, M.A.; Prochilo, T.; Quadri, A.; Gatta, G.; et al. Colon cancer. Crit. Rev. Oncol. Hematol. 2010, 74, 106–133. [Google Scholar] [CrossRef] [PubMed]
- Juul, J.S.; Hornung, N.; Andersen, B.; Laurberg, S.; Olesen, F.; Vedsted, P. The value of using the faecal immunochemical test in general practice on patients presenting with non-alarm symptoms of colorectal cancer. Br. J. Cancer 2018, 119, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Astin, M.; Griffin, T.; Neal, R.D.; Rose, P.; Hamilton, W. The diagnostic value of symptoms for colorectal cancer in primary care: A systematic review. Br. J. Gen. Pract. 2011, 61, e231–e243. [Google Scholar] [CrossRef] [PubMed]
- Annese, V.; Bassotti, G.; Napolitano, G.; Usai, P.; Andriulli, A.; Vantrappen, G. Gastrointestinal motility disorders in patients with inactive Crohn’s disease. Scand. J. Gastroenterol. 1997, 32, 1107–1117. [Google Scholar] [CrossRef] [PubMed]
- Alcindor, T.; Beauger, N. Oxaliplatin: A review in the era of molecularly targeted therapy. Curr. Oncol. 2011, 18, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Tobin, P.; Clarke, S.J. Management of chemotherapy-induced nausea, vomiting, oral mucositis, and diarrhoea. Lancet Oncol. 2005, 6, 93–102. [Google Scholar] [CrossRef]
- Jordan, K.; Kasper, C.; Schmoll, H.J. Chemotherapy-induced nausea and vomiting: Current and new standards in the antiemetic prophylaxis and treatment. Eur. J. Cancer 2005, 41, 199–205. [Google Scholar] [CrossRef]
- Becker, D.E. Nausea, vomiting, and hiccups: A review of mechanisms and treatment. Anesth. Prog. 2010, 57, 150–157. [Google Scholar] [CrossRef]
- Karthaus, M.; Schiel, X.; Ruhlmann, C.H.; Celio, L. Neurokinin-1 receptor antagonists: Review of their role for the prevention of chemotherapy-induced nausea and vomiting in adults. Expert Rev. Clin. Pharmacol. 2019, 12, 661–680. [Google Scholar] [CrossRef] [PubMed]
- Babic, T.; Browning, K.N. The role of vagal neurocircuits in the regulation of nausea and vomiting. Eur. J. Pharmacol. 2014, 722, 38–47. [Google Scholar] [CrossRef]
- Rojas, C.; Slusher, B.S. Pharmacological mechanisms of 5-HT(3) and tachykinin NK(1) receptor antagonism to prevent chemotherapy-induced nausea and vomiting. Eur. J. Pharmacol. 2012, 684, 1–7. [Google Scholar] [CrossRef]
- Zhong, W.; Shahbaz, O.; Teskey, G.; Beever, A.; Kachour, N.; Venketaraman, V.; Darmani, N.A. Mechanisms of Nausea and Vomiting: Current Knowledge and Recent Advances in Intracellular Emetic Signaling Systems. Int. J. Mol. Sci. 2021, 22, 5797. [Google Scholar] [CrossRef]
- Renn, C.L.; Carozzi, V.A.; Rhee, P.; Gallop, D.; Dorsey, S.G.; Cavaletti, G. Multimodal assessment of painful peripheral neuropathy induced by chronic oxaliplatin-based chemotherapy in mice. Mol. Pain 2011, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Vera, G.; Chiarlone, A.; Martin, M.I.; Abalo, R. Altered feeding behaviour induced by long-term cisplatin in rats. Auton. Neurosci. Basic Clin. 2006, 126–127, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Matsunaga, S.; Matsui, M.; Takeda, N.; Yamatodani, A. Pica in mice as a new model for the study of emesis. Methods Find. Exp. Clin. Pharmacol. 2002, 24, 135–138. [Google Scholar] [CrossRef]
- Robinson, A.M.; Stojanovska, V.; Rahman, A.A.; McQuade, R.M.; Senior, P.V.; Nurgali, K. Effects of Oxaliplatin Treatment on the Enteric Glial Cells and Neurons in the Mouse Ileum. J. Histochem. Cytochem. 2016, 64, 530–545. [Google Scholar] [CrossRef] [PubMed]
- McQuade, R.M.; Carbone, S.E.; Stojanovska, V.; Rahman, A.; Gwynne, R.M.; Robinson, A.M.; Goodman, C.A.; Bornstein, J.C.; Nurgali, K. Role of oxidative stress in oxaliplatin-induced enteric neuropathy and colonic dysmotility in mice. Br. J. Pharmacol. 2016, 173, 3502–3521. [Google Scholar] [CrossRef] [PubMed]
- Kajimoto, Y.; Nishino, N.; Hashimoto, T.; Shirai, Y.; Kuno, T.; Tanaka, C. cDNA cloning and tissue distribution of a rat ubiquitin carboxyl-terminal hydrolase PGP9.5. J. Biochem. 1992, 112, 28–32. [Google Scholar] [CrossRef]
- Qu, Z.-D.; Thacker, M.; Castelucci, P.; Bagyánszki, M.; Epstein, M.L.; Furness, J.B. Immunohistochemical analysis of neuron types in the mouse small intestine. Cell Tissue Res. 2008, 334, 147–161. [Google Scholar] [CrossRef]
- Kressel, M.; Berthoud, H.R.; Neuhuber, W.L. Vagal innervation of the rat pylorus: An anterograde tracing study using carbocyanine dyes and laser scanning confocal microscopy. Cell Tissue Res. 1994, 275, 109–123. [Google Scholar] [CrossRef]
- Grider, J.R. Neurotransmitters mediating the intestinal peristaltic reflex in the mouse. J. Pharmacol. Exp. Ther. 2003, 307, 460–467. [Google Scholar] [CrossRef]
- Weihe, E.; Tao-Cheng, J.H.; Schäfer, M.K.; Erickson, J.D.; E Eiden, L. Visualization of the vesicular acetylcholine transporter in cholinergic nerve terminals and its targeting to a specific population of small synaptic vesicles. Proc. Natl. Acad. Sci. USA 1996, 93, 3547–3552. [Google Scholar] [CrossRef] [PubMed]
- Nagatsu, T. The human tyrosine hydroxylase gene. Cell. Mol. Neurobiol. 1989, 9, 313–321. [Google Scholar] [CrossRef]
- Stavely, R.; Fraser, S.; Sharma, S.; Rahman, A.A.; Stojanovska, V.; Sakkal, S.; Apostolopoulos, V.; Bertrand, P.; Nurgali, K. The Onset and Progression of Chronic Colitis Parallels Increased Mucosal Serotonin Release via Enterochromaffin Cell Hyperplasia and Downregulation of the Serotonin Reuptake Transporter. Inflamm. Bowel Dis. 2018, 24, 1021–1034. [Google Scholar] [CrossRef]
- Rahman, A.A.; Shahid, I.Z.; Pilowsky, P.M. Intrathecal neuromedin U induces biphasic effects on sympathetic vasomotor tone, increases respiratory drive and attenuates sympathetic reflexes in rat. Br. J. Pharmacol. 2011, 164, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Malik, N.; Sanger, G.J.; Friedman, M.I.; Andrews, P.L. Pica—A model of nausea? Species differences in response to cisplatin. Physiol. Behav. 2005, 85, 271–277. [Google Scholar] [CrossRef]
- Vera, G.; Castillo, M.; Cabezos, P.A.; Chiarlone, A.; Martin, M.I.; Gori, A.; Pasquinelli, G.; Barbara, G.; Stanghellini, V.; Corinaldesi, R.; et al. Enteric neuropathy evoked by repeated cisplatin in the rat. Neurogastroenterol. Motil. 2011, 23, 370–378. [Google Scholar] [CrossRef]
- Cabezos, P.A.; Vera, G.; Castillo, M.; Fernandez-Pujol, R.; Martin, M.I.; Abalo, R. Radiological study of gastrointestinal motor activity after acute cisplatin in the rat. Temporal relationship with pica. Auton. Neurosci. Basic Clin. 2008, 141, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Zarate, N.; Mearin, F.; Wang, X.Y.; Hewlett, B.; Huizinga, J.D.; Malagelada, J.R. Severe idiopathic gastroparesis due to neuronal and interstitial cells of Cajal degeneration: Pathological findings and management. Gut 2003, 52, 966–970. [Google Scholar] [CrossRef]
- Pini, A.; Garella, R.; Idrizaj, E.; Calosi, L.; Baccari, M.C.; Vannucchi, M.G. Glucagon-like peptide 2 counteracts the mucosal damage and the neuropathy induced by chronic treatment with cisplatin in the mouse gastric fundus. Neurogastroenterol. Motil. 2016, 28, 206–216. [Google Scholar] [CrossRef]
- Powley, T.L.; Phillips, R.J. Gastric satiation is volumetric, intestinal satiation is nutritive. Physiol. Behav. 2004, 82, 69–74. [Google Scholar] [CrossRef]
- Phillips, R.J.; Powley, T.L. Gastric volume detection after selective vagotomies in rats. Am. J. Physiol. 1998, 274, R1626–R1638. [Google Scholar] [CrossRef] [PubMed]
- De Jonghe, B.C.; Horn, C.C. Chemotherapy-induced pica and anorexia are reduced by common hepatic branch vagotomy in the rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R756–R765. [Google Scholar] [CrossRef]
- Basile, D.; Di Nardo, P.; Corvaja, C.; Garattini, S.K.; Pelizzari, G.; Lisanti, C.; Bortot, L.; Da Ros, L.; Bartoletti, M.; Borghi, M.; et al. Mucosal Injury during Anti-Cancer Treatment: From Pathobiology to Bedside. Cancers 2019, 11, 857. [Google Scholar] [CrossRef] [PubMed]
- Pasricha, P.J.; Pehlivanov, N.D.; Gomez, G.; Vittal, H.; Lurken, M.S.; Farrugia, G. Changes in the gastric enteric nervous system and muscle: A case report on two patients with diabetic gastroparesis. BMC Gastroenterol. 2008, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Lucas, M.L. Diarrhoeal disease through enterocyte secretion: A doctrine untroubled by proof. Exp. Physiol. 2010, 95, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Wafai, L.; Taher, M.; Jovanovska, V.; Bornstein, J.C.; Dass, C.R.; Nurgali, K. Effects of oxaliplatin on mouse myenteric neurons and colonic motility. Front. Neurosci. 2013, 7, 30. [Google Scholar] [CrossRef]
- McQuade, R.M.; Stojanovska, V.; Donald, E.; Abalo, R.; Bornstein, J.C.; Nurgali, K. Gastrointestinal dysfunction and enteric neurotoxicity following treatment with anticancer chemotherapeutic agent 5-fluorouracil. Neurogastroenterol. Motil. 2016, 28, 1861–1875. [Google Scholar] [CrossRef]
- Rivera, L.R.; Poole, D.P.; Thacker, M.; Furness, J.B. The involvement of nitric oxide synthase neurons in enteric neuropathies. Neurogastroenterol. Motil. 2011, 23, 980–988. [Google Scholar] [CrossRef]
- Schemann, M.; Reiche, D.; Michel, K. Enteric pathways in the stomach. Anat. Rec. 2001, 262, 47–57. [Google Scholar] [CrossRef]
- Demedts, I.; Masaoka, T.; Kindt, S.; De Hertogh, G.; Geboes, K.; Farre, R.; Vanden Berghe, P.; Tack, J. Gastrointestinal motility changes and myenteric plexus alterations in spontaneously diabetic biobreeding rats. J. Neurogastroenterol. Motil. 2013, 19, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Browning, K.N.; Travagli, R.A. Central nervous system control of gastrointestinal motility and secretion and modulation of gastrointestinal functions. Compr. Physiol. 2014, 4, 1339–1368. [Google Scholar] [CrossRef] [PubMed]
- Kuebler, J.P.; Colangelo, L.; O’Connell, M.J.; Smith, R.E.; Yothers, G.; Begovic, M.; Robinson, B.; Seay, T.E.; Wolmark, N. Severe enteropathy among patients with stage II/III colon cancer treated on a randomized trial of bolus 5-fluorouracil/leucovorin plus or minus oxaliplatin: A prospective analysis. Cancer 2007, 110, 1945–1950. [Google Scholar] [CrossRef]
- Rahman, A.A.; Stojanovska, V.; Pilowsky, P.; Nurgali, K. Platinum accumulation in the brain and alteration in the central regulation of cardiovascular and respiratory functions in oxaliplatin-treated rats. Pflug. Arch. 2021, 473, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekharan, B.; Anitha, M.; Blatt, R.; Shahnavaz, N.; Kooby, D.; Staley, C.; Mwangi, S.; Jones, D.P.; Sitaraman, S.V.; Srinivasan, S. Colonic motor dysfunction in human diabetes is associated with enteric neuronal loss and increased oxidative stress. Neurogastroenterol. Motil. 2011, 23, 131–138. [Google Scholar] [CrossRef]
- Linden, D.R.; Couvrette, J.M.; Ciolino, A.; McQuoid, C.; Blaszyk, H.; Sharkey, K.A.; Mawe, G.M. Indiscriminate loss of myenteric neurones in the TNBS-inflamed guinea-pig distal colon. Neurogastroenterol. Motil. 2005, 17, 751–760. [Google Scholar] [CrossRef]
- Lomax, A.E.; O’Hara, J.R.; Hyland, N.P.; Mawe, G.M.; Sharkey, K.A. Persistent alterations to enteric neural signaling in the guinea pig colon following the resolution of colitis. Am. J. Physiol. 2007, 292, G482–G491. [Google Scholar] [CrossRef]
- Nurgali, K.; Qu, Z.; Hunne, B.; Thacker, M.; Pontell, L.; Furness, J.B. Morphological and functional changes in guinea-pig neurons projecting to the ileal mucosa at early stages after inflammatory damage. J. Physiol. 2011, 589, 325–339. [Google Scholar] [CrossRef]
- Phillips, R.J.; Powley, T.L. Innervation of the gastrointestinal tract: Patterns of aging. Auton. Neurosci. Basic Clin. 2007, 136, 1–19. [Google Scholar] [CrossRef]
- Khurana, R.K.; Petras, J.M. Sensory innervation of the canine esophagus, stomach, and duodenum. Am. J. Anat. 1991, 192, 293–306. [Google Scholar] [CrossRef]
- Cavaletti, G.; Tredici, G.; Petruccioli, M.G.; Donde, E.; Tredici, P.; Marmiroli, P.; Minoia, C.; Ronchi, A.; Bayssas, M.; Etienne, G.G. Effects of different schedules of oxaliplatin treatment on the peripheral nervous system of the rat. Eur. J. Cancer 2001, 37, 2457–2463. [Google Scholar] [CrossRef] [PubMed]
- Apostolidis, L.; Schwarz, D.; Xia, A.; Weiler, M.; Heckel, A.; Godel, T.; Heiland, S.; Schlemmer, H.-P.; Jäger, D.; Bendszus, M.; et al. Dorsal root ganglia hypertrophy as in vivo correlate of oxaliplatin-induced polyneuropathy. PLoS ONE 2017, 12, e0183845. [Google Scholar] [CrossRef] [PubMed]
- Ta, L.E.; Espeset, L.; Podratz, J.; Windebank, A.J. Neurotoxicity of oxaliplatin and cisplatin for dorsal root ganglion neurons correlates with platinum-DNA binding. Neurotoxicology 2006, 27, 992–1002. [Google Scholar] [CrossRef]
- Stillman, M.; Cata, J.P. Management of chemotherapy-induced peripheral neuropathy. Curr. Pain Headache Rep. 2006, 10, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, K.A. Substance P and Calcitonin Gene-Related Peptide (CGRP) in Gastrointestinal Inflammationa. Ann. N. Y. Acad. Sci. 1992, 664, 425–442. [Google Scholar] [CrossRef]
- Ishii, M.; Igarashi, M.; Patel, S.; Himi, T.; Kulecz, W. Autonomic effects on R-R variations of the heart rate in the squirrel monkey: An indicator of autonomic imbalance in conflict sickness. Am. J. Otolaryngol. 1987, 8, 144–148. [Google Scholar] [CrossRef]
- Schlegel, T.T.; Brown, T.E.; Wood, S.J.; Benavides, E.W.; Bondar, R.L.; Stein, F.; Moradshahi, P.; Harm, D.L.; Fritsch-Yelle, J.M.; Low, P.A. Orthostatic intolerance and motion sickness after parabolic flight. J. Appl. Physiol. 2001, 90, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Floyd, J.D.; Nguyen, D.T.; Lobins, R.L.; Bashir, Q.; Doll, D.C.; Perry, M.C. Cardiotoxicity of cancer therapy. J. Clin. Oncol. 2005, 23, 7685–7696. [Google Scholar] [CrossRef]
- Yeh, E.T.; Tong, A.T.; Lenihan, D.J.; Yusuf, S.W.; Swafford, J.; Champion, C.; Durand, J.B.; Gibbs, H.; Zafarmand, A.A.; Ewer, M.S. Cardiovascular complications of cancer therapy: Diagnosis, pathogenesis, and management. Circulation 2004, 109, 3122–3131. [Google Scholar] [CrossRef]
- Dermitzakis, E.V.; Kimiskidis, V.K.; Eleftheraki, A.; Lazaridis, G.; Konstantis, A.; Basdanis, G.; Tsiptsios, I.; Georgiadis, G.; Fountzilas, G. The impact of oxaliplatin-based chemotherapy for colorectal cancer on the autonomous nervous system. Eur. J. Neurol. 2014, 21, 1471–1477. [Google Scholar] [CrossRef]
- Raybould, H.E. Visceral perception: Sensory transduction in visceral afferents and nutrients. Gut 2002, 51, i11–i14. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.X.; Zhu, X.Y.; Owyang, C.; Li, Y. Intestinal serotonin acts as a paracrine substance to mediate vagal signal transmission evoked by luminal factors in the rat. J. Physiol. 2001, 530, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Andrews, P.L.; Horn, C.C. Signals for nausea and emesis: Implications for models of upper gastrointestinal diseases. Auton. Neurosci. Basic Clin. 2006, 125, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Andrews, P.L.; Davis, C.J.; Bingham, S.; Davidson, H.I.; Hawthorn, J.; Maskell, L. The abdominal visceral innervation and the emetic reflex: Pathways, pharmacology, and plasticity. Can. J. Physiol. Pharmacol. 1990, 68, 325–345. [Google Scholar] [CrossRef]
- Hawthorn, J.; Ostler, K.J.; Andrews, P.L. The role of the abdominal visceral innervation and 5-hydroxytryptamine M-receptors in vomiting induced by the cytotoxic drugs cyclophosphamide and cis-platin in the ferret. Q. J. Exp. Physiol. 1988, 73, 7–21. [Google Scholar] [CrossRef]
- Yasuda, M.; Kato, S.; Yamanaka, N.; Iimori, M.; Matsumoto, K.; Utsumi, D.; Kitahara, Y.; Amagase, K.; Horie, S.; Takeuchi, K. 5-HT₃ receptor antagonists ameliorate 5-fluorouracil-induced intestinal mucositis by suppression of apoptosis in murine intestinal crypt cells. Br. J. Pharmacol. 2013, 168, 1388–1400. [Google Scholar] [CrossRef]
- Hagbom, M.; Istrate, C.; Engblom, D.; Karlsson, T.; Rodriguez-Diaz, J.; Buesa, J.; Taylor, J.A.; Loitto, V.M.; Magnusson, K.E.; Ahlman, H.; et al. Rotavirus stimulates release of serotonin (5-HT) from human enterochromaffin cells and activates brain structures involved in nausea and vomiting. PLoS Pathog. 2011, 7, e1002115. [Google Scholar] [CrossRef]
- Blakelock, R.T.; Beasley, S.W. Infection and the gut. Semin. Pediatr. Surg. 2003, 12, 265–274. [Google Scholar] [CrossRef]
- Minami, M.; Endo, T.; Hirafuji, M.; Hamaue, N.; Liu, Y.; Hiroshige, T.; Nemoto, M.; Saito, H.; Yoshioka, M. Pharmacological aspects of anticancer drug-induced emesis with emphasis on serotonin release and vagal nerve activity. Pharmacol. Ther. 2003, 99, 149–165. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, A.A.; Masango, P.; Stavely, R.; Bertrand, P.; Page, A.; Nurgali, K. Oxaliplatin-Induced Damage to the Gastric Innervation: Role in Nausea and Vomiting. Biomolecules 2023, 13, 276. https://doi.org/10.3390/biom13020276
Rahman AA, Masango P, Stavely R, Bertrand P, Page A, Nurgali K. Oxaliplatin-Induced Damage to the Gastric Innervation: Role in Nausea and Vomiting. Biomolecules. 2023; 13(2):276. https://doi.org/10.3390/biom13020276
Chicago/Turabian StyleRahman, Ahmed A., Philenkosini Masango, Rhian Stavely, Paul Bertrand, Amanda Page, and Kulmira Nurgali. 2023. "Oxaliplatin-Induced Damage to the Gastric Innervation: Role in Nausea and Vomiting" Biomolecules 13, no. 2: 276. https://doi.org/10.3390/biom13020276
APA StyleRahman, A. A., Masango, P., Stavely, R., Bertrand, P., Page, A., & Nurgali, K. (2023). Oxaliplatin-Induced Damage to the Gastric Innervation: Role in Nausea and Vomiting. Biomolecules, 13(2), 276. https://doi.org/10.3390/biom13020276