
Pro-Inflammatory and Cytotoxic Effects of Polystyrene Microplastics on Human and Murine Intestinal Cell Lines

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Cultures and PS-MPs Treatment, Cell Viability and Oxidative Stress
2.1.1. Cell Cultures and PS-MPs Treatment
2.1.2. Cell Viability—MTT Assay
2.1.3. Oxidative Stress—SOD Assay
2.2. Magnetic-Beads Panel MultiplexPlex Assay
2.3. RNA Extraction, Retrotranscription and Quantitative Real-Time PCR
2.4. Statistical Analysis
2.4.1. MTT and SOD Assays Statistical Analysis
2.4.2. Magnetic-Beads Panel MultiplexPlex Assay Statistical Analysis
2.4.3. Real-Time PCR Statistical Analysis
3. Results
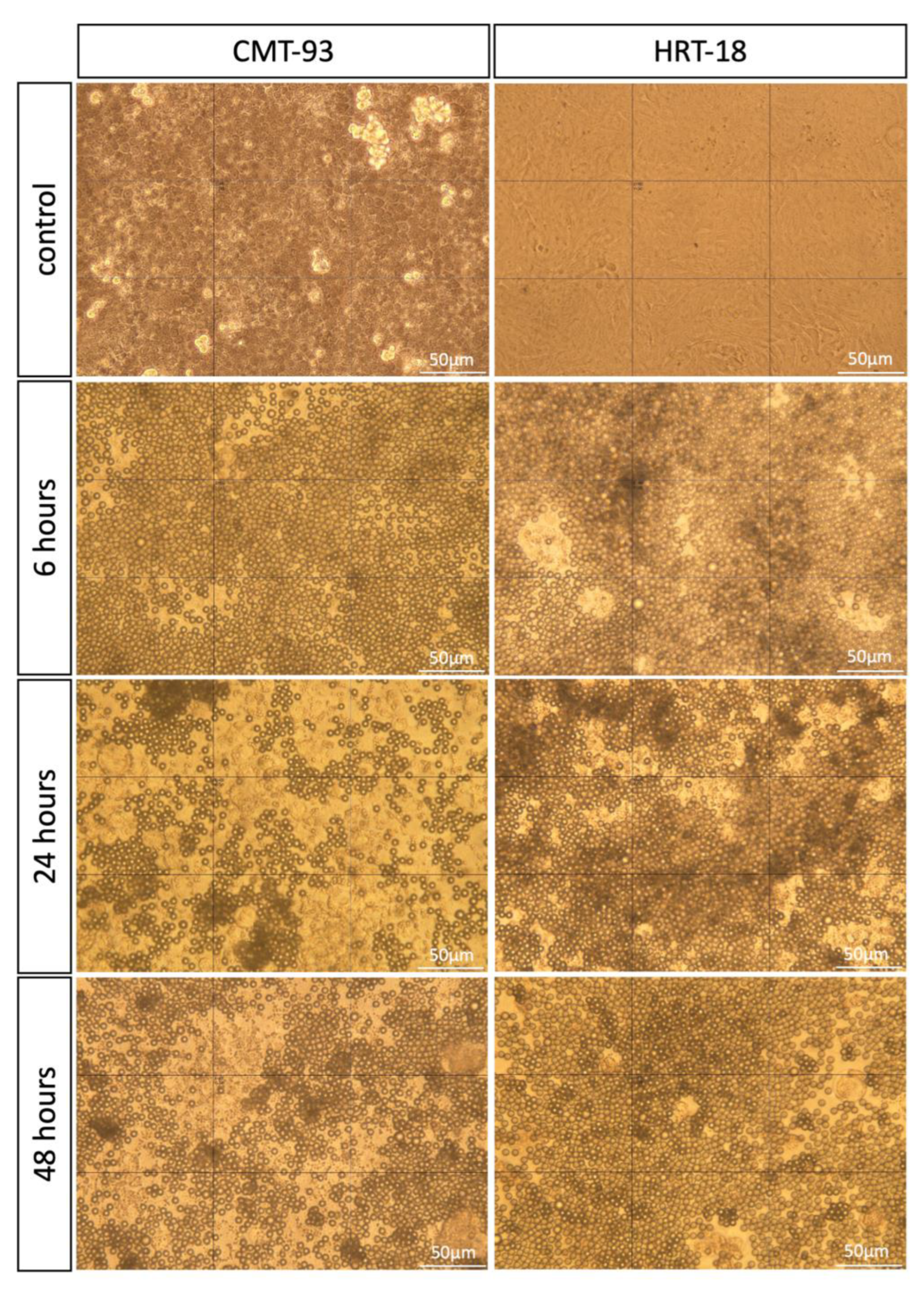
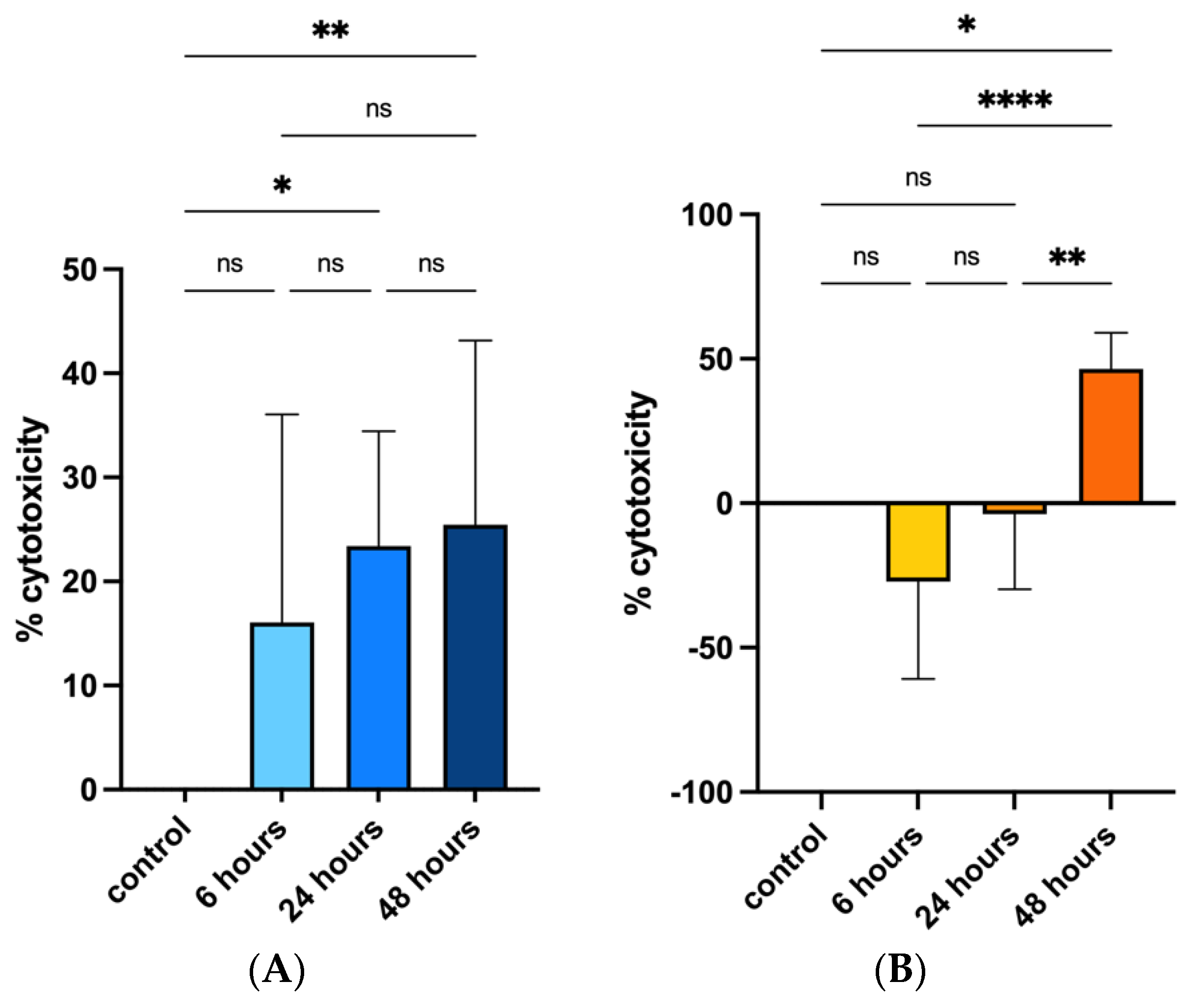
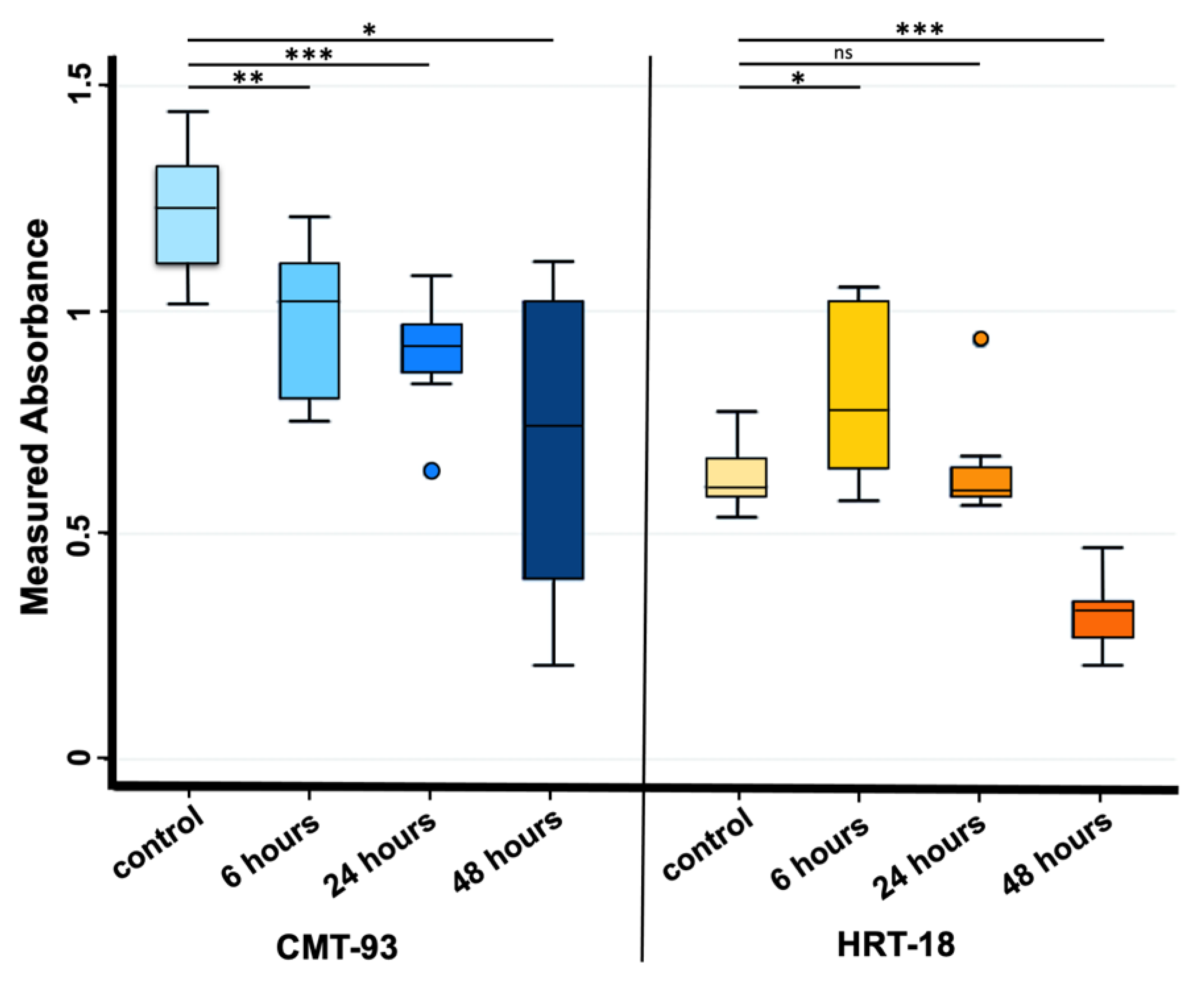
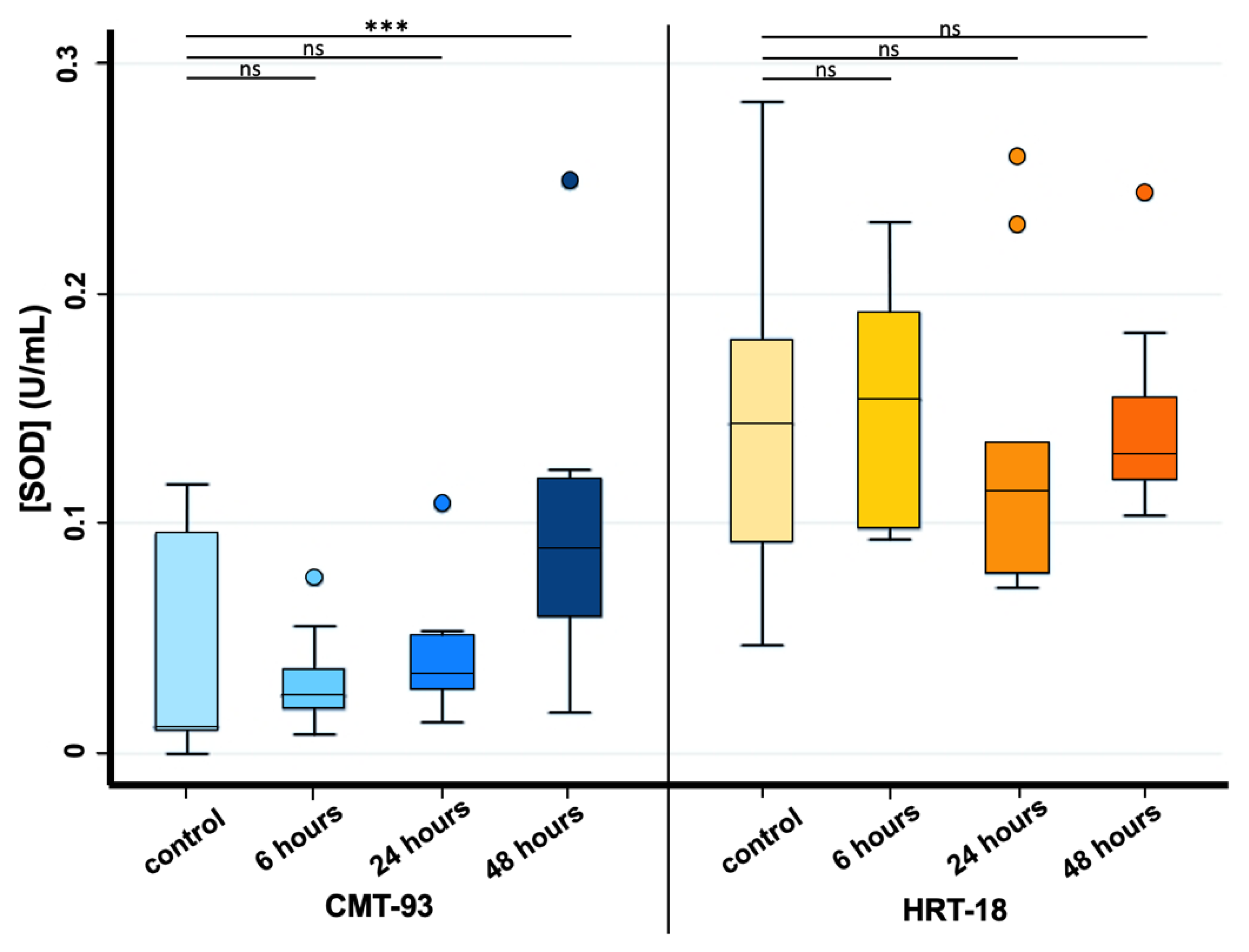
3.1. Cell Cultures, Cell Viability and Oxidative Stress after PS-MPs Treatment
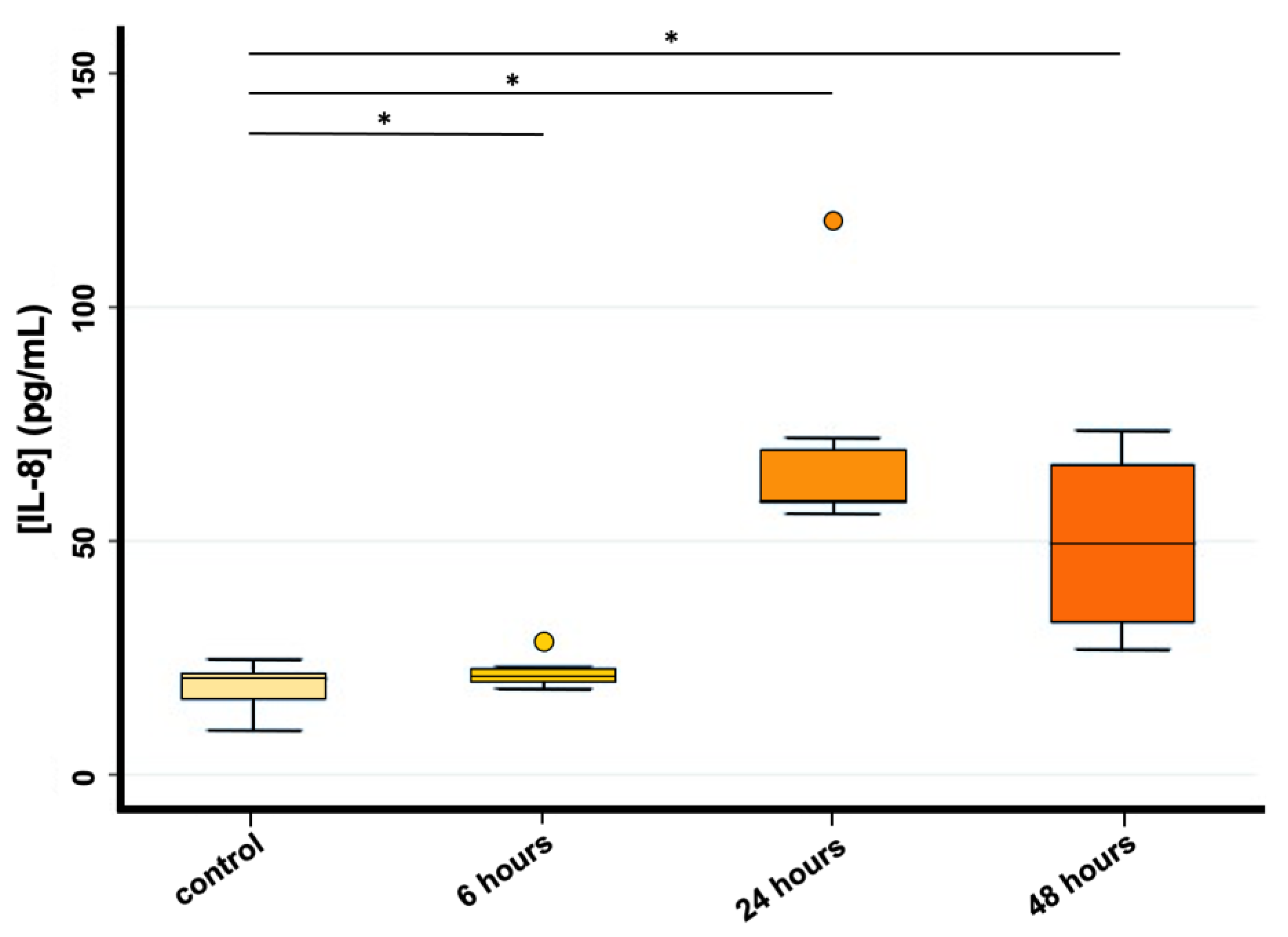
3.2. Magnetic-Beads Panel MultiplexPlex Luminex Assay
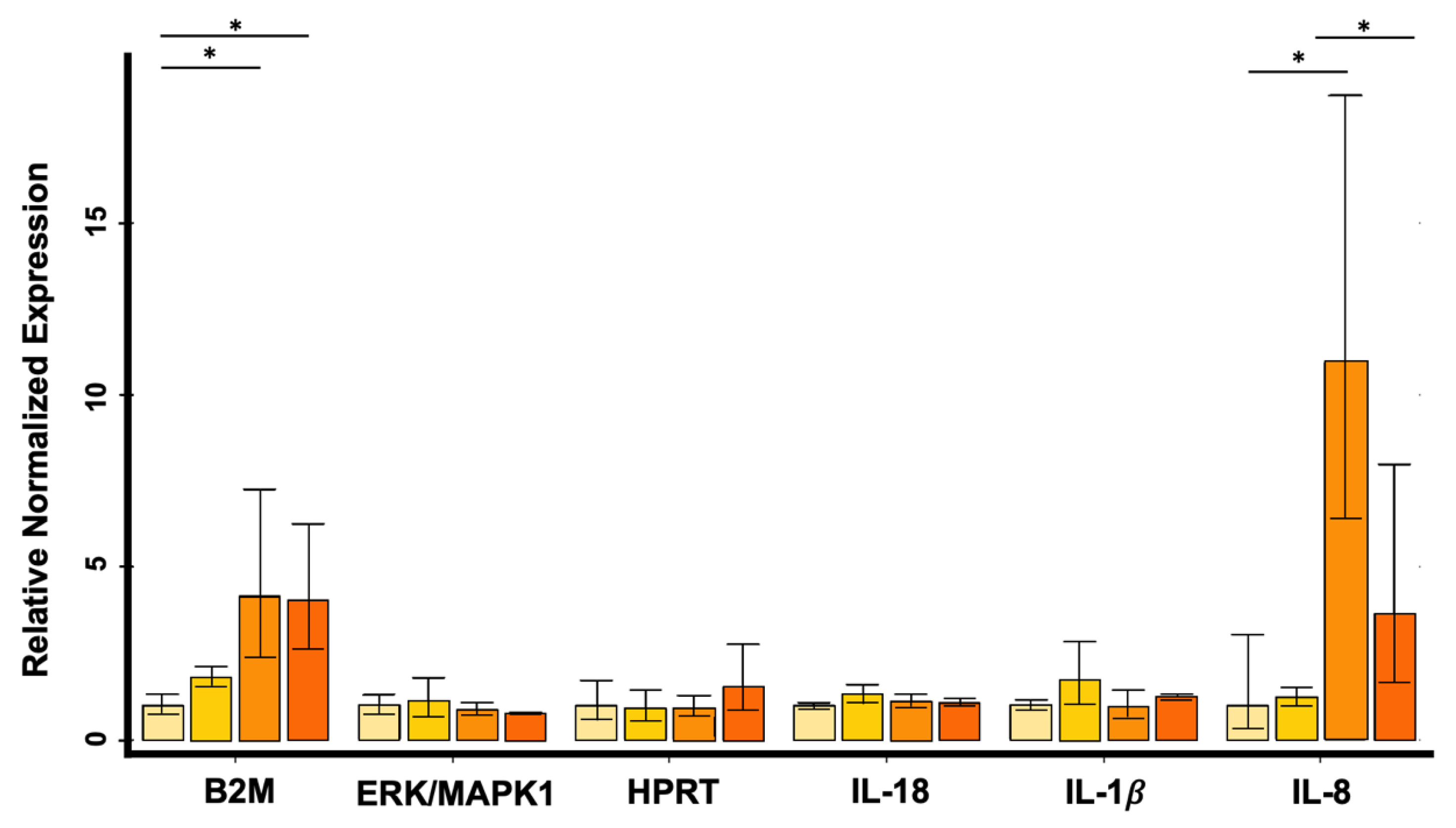
3.3. Quantitative Real-Time PCR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MPs | Microplastics |
| PS-MPs | Polystyrene microplastics |
| GIT | Gastro-intestinal tract |
| EFSA | European Food Safety Authority |
| CREDIMA | National Reference Center for Marine Mammals Diagnostics |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-2H-tetrazolium bromide |
| SOD | Superoxide dismutase |
| ROS | Reactive oxygen species |
References
- Schlatter, C. Environmental Pollution and Human Health. Sci. Total Environ. 1994, 143, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Briggs, D. Environmental Pollution and the Global Burden of Disease. Br. Med. Bull. 2003, 68, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Forleo, A.; Radogna, A.V.; Siciliano, P.; Notari, T.; Pappalardo, S.; Piscopo, M.; Montano, L.; Capone, S. A Novel Human Biomonitoring Study by Semiconductor Gas Sensors in Exposomics: Investigation of Health Risk in Contaminated Sites. Environ. Pollut. 2022, 304, 119119. [Google Scholar] [CrossRef]
- Montano, L.; Pironti, C.; Pinto, G.; Ricciardi, M.; Buono, A.; Brogna, C.; Venier, M.; Piscopo, M.; Amoresano, A.; Motta, O. Polychlorinated Biphenyls (PCBs) in the Environment: Occupational and Exposure Events, Effects on Human Health and Fertility. Toxics 2022, 10, 365. [Google Scholar] [CrossRef] [PubMed]
- Waring, R.H.; Harris, R.M.; Mitchell, S.C. Plastic Contamination of the Food Chain: A Threat to Human Health? Maturitas 2018, 115, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy Metal Pollution in the Environment and Their Toxicological Effects on Humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef]
- Manisalidis, I.; Stavropoulou, E.; Stavropoulos, A.; Bezirtzoglou, E. Environmental and Health Impacts of Air Pollution: A Review. Front. Public Health 2020, 8, 14. [Google Scholar] [CrossRef]
- PlascticsEurope. Plastics-the Facts 2021—An Analysis of European Plastics Production, Demand and Waste Data. 2021. Available online: https://plasticseurope.org/wp-content/uploads/2021/12/Plastics-the-Facts-2021-web-final.pdf (accessed on 26 December 2022).
- Meaza, I.; Toyoda, J.H.; Wise, J.P. Microplastics in Sea Turtles, Marine Mammals and Humans: A One Environmental Health Perspective. Front. Environ. Sci. 2021, 8, 575614. [Google Scholar] [CrossRef]
- Mü, R.-J.; Kleeberg, I.; Deckwer, W.-D. Biodegradation of Polyesters Containing Aromatic Constituents. J. Biotechnol. 2001, 86, 87–95. [Google Scholar]
- Edge, M.; Hayes, M.; Mohammadian, M.; Allen, N.S.; Jewitt, T.S.; Brems, K.; Jones, K. Aspects of Poly(Ethylene Terephthalate) Degradation for Archival Life and Environmental Degradation. Polym. Degrad. Stab. 1991, 32, 131–153. [Google Scholar] [CrossRef]
- Allen, N.S.; Edge, M.; Mohammadian, M.; Jones, K. Physicochemical Aspects of the Environmental Degradation of Poly(Ethylene Terephthalate). Polym. Degrad. Stab. 1994, 43, 229–237. [Google Scholar] [CrossRef]
- Webb, H.K.; Arnott, J.; Crawford, R.J.; Ivanova, E.P. Plastic Degradation and Its Environmental Implications with Special Reference to Poly(Ethylene Terephthalate). Polymers 2013, 5, 1–18. [Google Scholar] [CrossRef]
- Paul, M.B.; Stock, V.; Cara-Carmona, J.; Lisicki, E.; Shopova, S.; Fessard, V.; Braeuning, A.; Sieg, H.; Böhmert, L. Micro- And Nanoplastics-Current State of Knowledge with the Focus on Oral Uptake and Toxicity. Nanoscale Adv. 2020, 2, 4350–4367. [Google Scholar] [CrossRef] [PubMed]
- Kutralam-Muniasamy, G.; Shruti, V.C.; Pérez-Guevara, F.; Roy, P.D. Microplastic Diagnostics in Humans: “The 3Ps” Progress, Problems, and Prospects. Sci. Total Environ. 2023, 856, 159164. [Google Scholar] [CrossRef]
- Liu, P.; Zhan, X.; Wu, X.; Li, J.; Wang, H.; Gao, S. Effect of Weathering on Environmental Behavior of Microplastics: Properties, Sorption and Potential Risks. Chemosphere 2020, 242, 125193. [Google Scholar] [CrossRef] [PubMed]
- Presence of Microplastics and Nanoplastics in Food, with Particular Focus on Seafood. EFSA J. 2016, 14, e04501. [CrossRef]
- Zantis, L.J.; Carroll, E.L.; Nelms, S.E.; Bosker, T. Marine Mammals and Microplastics: A Systematic Review and Call for Standardisation. Environ. Pollut. 2021, 269, 116142. [Google Scholar] [CrossRef]
- Panti, C.; Baini, M.; Lusher, A.; Hernandez-Milan, G.; Bravo Rebolledo, E.L.; Unger, B.; Syberg, K.; Simmonds, M.P.; Fossi, M.C. Marine Litter: One of the Major Threats for Marine Mammals. Outcomes from the European Cetacean Society Workshop. Environ. Pollut. 2019, 247, 72–79. [Google Scholar]
- Corazzola, G.; Baini, M.; Grattarola, C.; Panti, C.; Marcer, F.; Garibaldi, F.; Berio, E.; Mancusi, C.; Galli, M.; Mazzariol, S.; et al. Analysis of the Gastro-Intestinal Tract of Marine Mammals: A Multidisciplinary Approach with a New Multi-Sieves Tool. Animals 2021, 11, 1824. [Google Scholar] [CrossRef]
- Pearce, S.C.; Coia, H.G.; Karl, J.P.; Pantoja-Feliciano, I.G.; Zachos, N.C.; Racicot, K. Intestinal in Vitro and Ex Vivo Models to Study Host-Microbiome Interactions and Acute Stressors. Front. Physiol. 2018, 9, 1584. [Google Scholar] [CrossRef]
- Lei, L.; Wu, S.; Lu, S.; Liu, M.; Song, Y.; Fu, Z.; Shi, H.; Raley-Susman, K.M.; He, D. Microplastic Particles Cause Intestinal Damage and Other Adverse Effects in Zebrafish Danio Rerio and Nematode Caenorhabditis Elegans. Sci. Total Environ. 2018, 619–620, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Wu, X.; Liu, S.; Wang, Z.; Chen, L. Size-Dependent Effects of Polystyrene Microplastics on Cytotoxicity and Efflux Pump Inhibition in Human Caco-2 cells. Chemosphere 2019, 221, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Hesler, M.; Aengenheister, L.; Ellinger, B.; Drexel, R.; Straskraba, S.; Jost, C.; Wagner, S.; Meier, F.; von Briesen, H.; Büchel, C.; et al. Multi-Endpoint Toxicological Assessment of Polystyrene Nano- and Microparticles in Different Biological Models in Vitro. Toxicol. Vitr. 2019, 61, 104610. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Wu, M.; Tian, D.; Qiu, L.; Li, T. Effects of Polystyrene Microbeads on Cytotoxicity and Transcriptomic Profiles in Human Caco-2 Cells. Env. Toxicol. 2020, 35, 495–506. [Google Scholar] [CrossRef]
- Dong, C.-D.; Chen, C.W.; Chen, Y.C.; Chen, H.H.; Lee, J.S.; Lin, C.H. Polystyrene Microplastic Particles: In Vitro Pulmonary Toxicity Assessment. J. Hazard Mater. 2020, 385, 121575. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.; Hwang, J.; Bang, J.; Han, S.; Kim, T.; Oh, Y.; Hwang, Y.; Choi, J.; Hong, J. In Vitro Toxicity from a Physical Perspective of Polyethylene Microplastics Based on Statistical Curvature Change Analysis. Sci. Total Environ. 2021, 752, 142242. [Google Scholar] [CrossRef] [PubMed]
- Stock, V.; Laurisch, C.; Franke, J.; Dönmez, M.H.; Voss, L.; Böhmert, L.; Braeuning, A.; Sieg, H. Uptake and Cellular Effects of PE, PP, PET and PVC Microplastic Particles. Toxicol. Vitr. 2021, 70, 105021. [Google Scholar] [CrossRef]
- Koressaar, T.; Remm, M. Enhancements and Modifications of Primer Design Program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef]
- Benedetto, A.; Squadrone, S.; Prearo, M.; Elia, A.C.; Giorgi, I.; Abete, M.C. Evaluation of ABC Efflux Transporters Genes Expression in Kidney of Rainbow Trout (Oncorhynchus Mykiss) Fed with Melamine and Cyanuric Acid Diets. Chemosphere 2011, 84, 727–730. [Google Scholar] [CrossRef]
- González-Bermúdez, L.; Anglada, T.; Genescà, A.; Martín, M.; Terradas, M. Identification of Reference Genes for RT-QPCR Data Normalisation in Aging Studies. Sci. Rep. 2019, 9, 13970. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome. Biol. 2002, 3, 1–12. [Google Scholar] [CrossRef]
- Aves, A.R.; Revell, L.E.; Gaw, S.; Ruffell, H.; Schuddeboom, A.; Wotherspoon, N.E.; Larue, M.; Mcdonald, A.J. First Evidence of Microplastics in Antarctic Snow. Cryosphere 2022, 16, 2127–2145. [Google Scholar] [CrossRef]
- Wang, F.; Lai, Z.; Peng, G.; Luo, L.; Liu, K.; Huang, X.; Xu, Y.; Shen, Q.; Li, D. Microplastic Abundance and Distribution in a Central Asian Desert. Sci. Total Environ. 2021, 800, 149529. [Google Scholar] [CrossRef]
- Gardon, T.; el Rakwe, M.; Paul-Pont, I.; le Luyer, J.; Thomas, L.; Prado, E.; Boukerma, K.; Cassone, A.L.; Quillien, V.; Soyez, C.; et al. Microplastics Contamination in Pearl-Farming Lagoons of French Polynesia. J. Hazard Mater. 2021, 419, 126396. [Google Scholar] [CrossRef] [PubMed]
- Gambino, I.; Bagordo, F.; Grassi, T.; Panico, A.; de Donno, A. Occurrence of Microplastics in Tap and Bottled Water: Current Knowledge. Int. J. Environ. Res. Public Health 2022, 19, 5283. [Google Scholar] [CrossRef]
- Gasperi, J.; Wright, S.L.; Dris, R.; Collard, F.; Mandin, C.; Guerrouache, M.; Langlois, V.; Kelly, F.J.; Tassin, B. Microplastics in Air: Are We Breathing It In? Curr. Opin. Environ. Sci. Health 2018, 1, 1–5. [Google Scholar] [CrossRef]
- Ragusa, A.; Svelato, A.; Santacroce, C.; Catalano, P.; Notarstefano, V.; Carnevali, O.; Papa, F.; Rongioletti, M.C.A.; Baiocco, F.; Draghi, S.; et al. Plasticenta: First Evidence of Microplastics in Human Placenta. Environ. Int. 2021, 146, 106274. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, A.; Notarstefano, V.; Svelato, A.; Belloni, A.; Gioacchini, G.; Blondeel, C.; Zucchelli, E.; de Luca, C.; D’avino, S.; Gulotta, A.; et al. Raman Microspectroscopy Detection and Characterisation of Microplastics in Human Breastmilk. Polymers 2022, 14, 2700. [Google Scholar] [CrossRef] [PubMed]
- Leslie, H.A.; van Velzen, M.J.M.; Brandsma, S.H.; Vethaak, A.D.; Garcia-Vallejo, J.J.; Lamoree, M.H. Discovery and Quantification of Plastic Particle Pollution in Human Blood. Environ. Int. 2022, 163, 107199. [Google Scholar] [CrossRef]
- Wang, Q.; Bai, J.; Ning, B.; Fan, L.; Sun, T.; Fang, Y.; Wu, J.; Li, S.; Duan, C.; Zhang, Y.; et al. Effects of Bisphenol A and Nanoscale and Microscale Polystyrene Plastic Exposure on Particle Uptake and Toxicity in Human Caco-2 Cells. Chemosphere 2020, 254, 126788. [Google Scholar] [CrossRef] [PubMed]
- Lehner, R.; Wohlleben, W.; Septiadi, D.; Landsiedel, R.; Petri-Fink, A.; Rothen-Rutishauser, B. A Novel 3D Intestine Barrier Model to Study the Immune Response upon Exposure to Microplastics. Arch. Toxicol. 2020, 94, 2463–2479. [Google Scholar] [CrossRef]
- Khalid, N.; Aqeel, M.; Noman, A.; Khan, S.M.; Akhter, N. Interactions and Effects of Microplastics with Heavy Metals in Aquatic and Terrestrial Environments. Environ. Pollut. 2021, 290, 118104. [Google Scholar] [CrossRef] [PubMed]
- Lettieri, G.; Carusone, N.; Notariale, R.; Prisco, M.; Ambrosino, A.; Perrella, S.; Manna, C.; Piscopo, M. Morphological, Gene, and Hormonal Changes in Gonads and In-Creased Micrococcal Nuclease Accessibility of Sperm Chromatin Induced by Mercury. Biomolecules 2022, 12, 87. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Shelver, W.L. Micro- and Nanoplastic Induced Cellular Toxicity in Mammals: A Review. Sci. Total Environ. 2021, 755, 142518. [Google Scholar] [CrossRef] [PubMed]
- Stock, V.; Böhmert, L.; Lisicki, E.; Block, R.; Cara-Carmona, J.; Pack, L.K.; Selb, R.; Lichtenstein, D.; Voss, L.; Henderson, C.J.; et al. Uptake and Effects of Orally Ingested Polystyrene Microplastic Particles in Vitro and in Vivo. Arch. Toxicol. 2019, 93, 1817–1833. [Google Scholar] [CrossRef]
- Palaniappan, S.; Sadacharan, C.M.; Rostama, B. Polystyrene and Polyethylene Microplastics Decrease Cell Viability and Dysregulate Inflammatory and Oxidative Stress Markers of MDCK and L929 Cells In Vitro. Expo. Health 2022, 14, 75–85. [Google Scholar] [CrossRef]
- Stock, V.; Böhmert, L.; Dönmez, M.H.; Lampen, A.; Sieg, H. An Inverse Cell Culture Model for Floating Plastic Particles. Anal. Biochem. 2020, 591, 113545. [Google Scholar] [CrossRef]
- Schirinzi, G.F.; Pérez-Pomeda, I.; Sanchís, J.; Rossini, C.; Farré, M.; Barceló, D. Cytotoxic Effects of Commonly Used Nanomaterials and Microplastics on Cerebral and Epithelial Human Cells. Environ. Res. 2017, 159, 579–587. [Google Scholar] [CrossRef]
- Bhattacharya, P. A review on the impacts of microplastic beads used in cosmetics. Acta Biomed. 2016, 3, 47–52. [Google Scholar]
- Rubio, L.; Marcos, R.; Hernández, A. Potential Adverse Health Effects of Ingested Micro- and Nanoplastics on Humans. Lessons Learned from in Vivo and in Vitro Mammalian Models. J. Toxicol. Environ. Health B Crit. Rev. 2020, 23, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Hirt, N.; Body-Malapel, M. Immunotoxicity and Intestinal Effects of Nano- and Microplastics: A Review of the Literature. Part Fibre Toxicol. 2020, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- Donkers, J.M.; Höppener, E.M.; Grigoriev, I.; Will, L.; Melgert, B.N.; van der Zaan, B.; van de Steeg, E.; Kooter, I.M. Advanced Epithelial Lung and Gut Barrier Models Demonstrate Passage of Microplastic Particles. Microplastics Nanoplastics 2022, 2, 6. [Google Scholar] [CrossRef]
- Zinkernagel, A.S.; Timmer, A.M.; Pence, M.A.; Locke, J.B.; Buchanan, J.T.; Turner, C.E.; Mishalian, I.; Sriskandan, S.; Hanski, E.; Nizet, V. The IL-8 Protease SpyCEP/ScpC of Group A Streptococcus Promotes Resistance to Neutrophil Killing. Cell Host. Microbe 2008, 4, 170–178. [Google Scholar] [CrossRef]
- Quan, J.M.; Martin, T.R.; Rosenberg, G.B.; Foster, D.C.; Whitmore, T.; Goodman, R.B. Antibodies against the N-Terminus of IL-8 Receptor A Inhibit Neutrophil Chemotaxis. Biochem. Biophys. Res. Commun. 1996, 219, 405–411. [Google Scholar] [CrossRef]
- Cotton, J.A.; Platnich, J.M.; Muruve, D.A.; Jijon, H.B.; Buret, A.G.; Beck, P.L. Interleukin-8 in Gastrointestinal Inflammation and Malignancy: Induction and Clinical Consequences. Int. J. Interferon Cytokine Mediat. Res. 2016, 8, 13–34. [Google Scholar]
- Prata, J.C.; da Costa, J.P.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Environmental Exposure to Microplastics: An Overview on Possible Human Health Effects. Sci. Total Environ. 2020, 702, 134455. [Google Scholar] [CrossRef]
- Fournier, E.; Etienne-Mesmin, L.; Grootaert, C.; Jelsbak, L.; Syberg, K.; Blanquet-Diot, S.; Mercier-Bonin, M. Microplastics in the Human Digestive Environment: A Focus on the Potential and Challenges Facing in Vitro Gut Model Development. J. Hazard Mater. 2021, 415, 125632. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | Bead Region | Bead Region | Standard Curve (pg/mL) | Cell Culture | Sensitivity (pg/mL) |
|---|---|---|---|---|---|
| IL-1β/IL-1F2 | 28 | A | 17.7–4300 | 1:2 | 0.8 |
| IL-6 | 13 | A | 4.53–1100 | 1:2 | 1.7 |
| IL-7 | 29 | K | 5.14–1250 | 1:2 | 0.410 |
| IL-8/CXCL8 | 18 | A | 4.12–1000 | 1:2 | 1.8 |
| IL-10 | 22 | A | 4.12–1000 | 1:2 | 1.6 |
| IL-15 | 63 | J | 6.3–1550 | 1:2 | 1.01 |
| IL-18/IL-1F4 | 78 | C | 7.12–1730 | 1:2 | 1.93 |
| IL-23 | 76 | C | 144–35,000 | 1:2 | 11.4 |
| IL-33 | 14 | C | 12.3–3000 | 1:2 | 1.8 |
| Analyte | Bead Region | Bead Region | Standard Curve (pg/mL) | Cell Culture | Sensitivity (pg/mL) |
|---|---|---|---|---|---|
| IL-1β/IL-1F2 | 19 | Mouse A | 247–60,000 | 1:2 | 41.8 |
| IL-6 | 27 | Mouse A | 28.8–7000 | 1:2 | 2.30 |
| IL-7 | 14 | Mouse C | 267–65,000 | 1:2 | 35.4 |
| IL-10 | 28 | Mouse A | 12.8–3100 | 1:2 | 8.20 |
| IL-33 | 43 | Mouse B | 82.3–20,000 | 1:2 | 57.1 |
| Gene | Reference Number | Forward Sequence (5′-3′) | Reverse Sequence (5′-3′) | Species |
|---|---|---|---|---|
| IL-8 | NM_001310420.1 | CTCTTGGCAGCCTTCCTGAT | TTTGGGGTGGAAAGGTTTGGA | Human |
| IL-18 | NM_001386420.1 | AAGATGGCTGCTGAACCAGT | TGCCAAAGTAATCTGATTCCAGG | Human |
| IL-1β | NM_000576.3 | TCGCCAGTGAAATGATGGCT | GGTCGGAGATTCGTAGCTGG | Human |
| Cyclin D1 | NM_053056 | AGCTGTGCATCTACACCGAC | GAAATCGTGCGGGGTCATTG | Human |
| MAPK1/ERK | NM_002745.5 | CGTGTTGCAGATCCAGACCA | GCCAGAATGCAGCCTACAGA | Human |
| β-Actin | NM_001101.5 | ACAGAGCCTCGCCTTTGC | CGCGGCGATATCATCATCCA | Human |
| B2M | NM_004048.4 | CTGCCGTGTGAACCATGTGA | TCAAACCTCCATGATGCTGC | Human |
| HPRT | NM_000194.3 | TGCTGAGGATTTGGAAAGGGT | GGGCTACAATGTGATGGCCT | Human |
| IL-8/CXCL8 | NM_011339.2 | TGATGCTCCATGGGTGAAGG | CAGAAGCTTCATTGCCGGTG | Murine |
| IL-18 | NM_001357221.1 | GGCTGCCATGTCAGAAGACT | ACAGTGAAGTCGGCCAAAGT | Murine |
| IL-1β | NM_008361.4 | GCCACCTTTTGACAGTGATGAG | GACAGCCCAGGTCAAAGGTT | Murine |
| Cyclin D1 | NM_001379248.1 | AAACAAGGACCCCCTCCATC | GGCTTCAATCTGTTCCTGGC | Murine |
| MAPK1/ERK | NM_001038663.1 | CCTCCTGCTGAACACCACTT | ATCTGGATCTGCAACACGGG | Murine |
| β-Actin | NM_007393.5 | CTGTCGAGTCGCGTCCACC | CGCAGCGATATCGTCATCCAT | Murine |
| B2M | NM_009735.3 | GAGCCCAAGACCGTCTACTG | GGTTCAAATGAATCTTCAGAGCATC | Murine |
| HPRT | NM_013556.2 | TTCTTTGCTGACCTGCTGGA | TTATGTCCCCCGTTGACTGA | Murine |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mattioda, V.; Benedetti, V.; Tessarolo, C.; Oberto, F.; Favole, A.; Gallo, M.; Martelli, W.; Crescio, M.I.; Berio, E.; Masoero, L.; et al. Pro-Inflammatory and Cytotoxic Effects of Polystyrene Microplastics on Human and Murine Intestinal Cell Lines. Biomolecules 2023, 13, 140. https://doi.org/10.3390/biom13010140
Mattioda V, Benedetti V, Tessarolo C, Oberto F, Favole A, Gallo M, Martelli W, Crescio MI, Berio E, Masoero L, et al. Pro-Inflammatory and Cytotoxic Effects of Polystyrene Microplastics on Human and Murine Intestinal Cell Lines. Biomolecules. 2023; 13(1):140. https://doi.org/10.3390/biom13010140
Chicago/Turabian StyleMattioda, Virginia, Valerio Benedetti, Carlotta Tessarolo, Francesca Oberto, Alessandra Favole, Marina Gallo, Walter Martelli, Maria Ines Crescio, Enrica Berio, Loretta Masoero, and et al. 2023. "Pro-Inflammatory and Cytotoxic Effects of Polystyrene Microplastics on Human and Murine Intestinal Cell Lines" Biomolecules 13, no. 1: 140. https://doi.org/10.3390/biom13010140
APA StyleMattioda, V., Benedetti, V., Tessarolo, C., Oberto, F., Favole, A., Gallo, M., Martelli, W., Crescio, M. I., Berio, E., Masoero, L., Benedetto, A., Pezzolato, M., Bozzetta, E., Grattarola, C., Casalone, C., Corona, C., & Giorda, F. (2023). Pro-Inflammatory and Cytotoxic Effects of Polystyrene Microplastics on Human and Murine Intestinal Cell Lines. Biomolecules, 13(1), 140. https://doi.org/10.3390/biom13010140