Cardioprotective Effect of Glycyrrhizin on Myocardial Remodeling in Diabetic Rats



Abstract
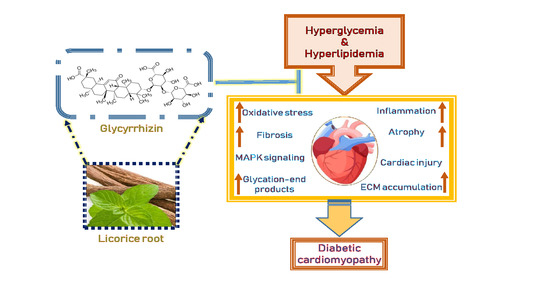
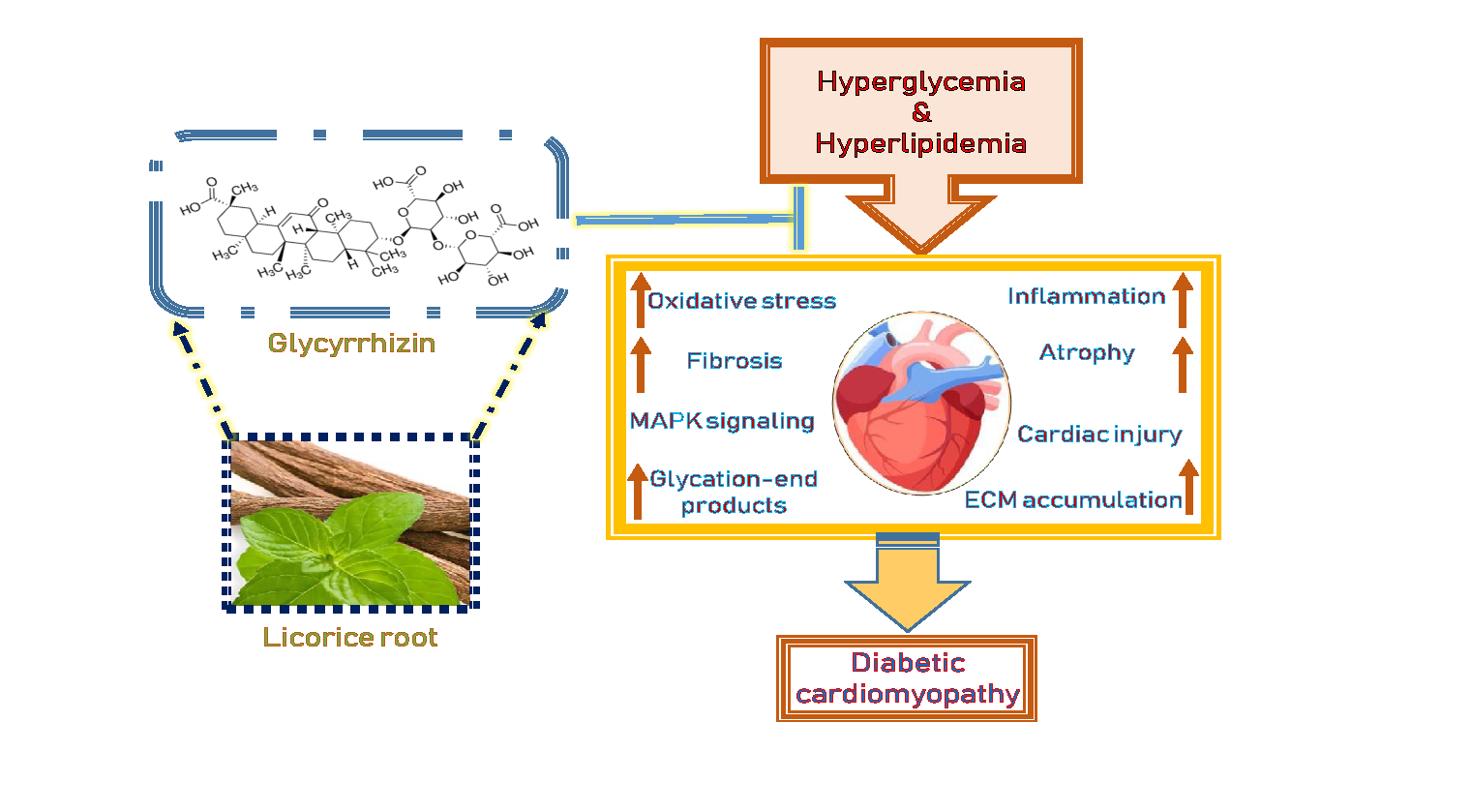{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Treatment and Tissue Collection
2.3. Western Blot
2.4. Histopathological Analysis
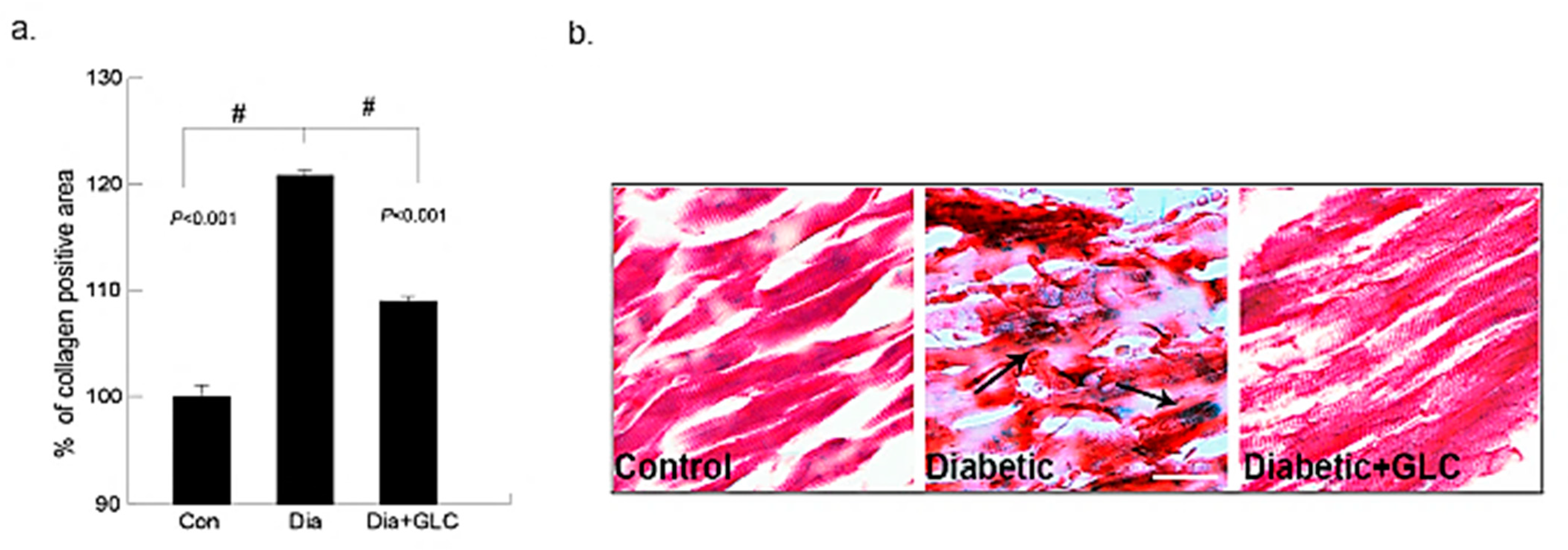
2.5. Collagen Deposition Quantification
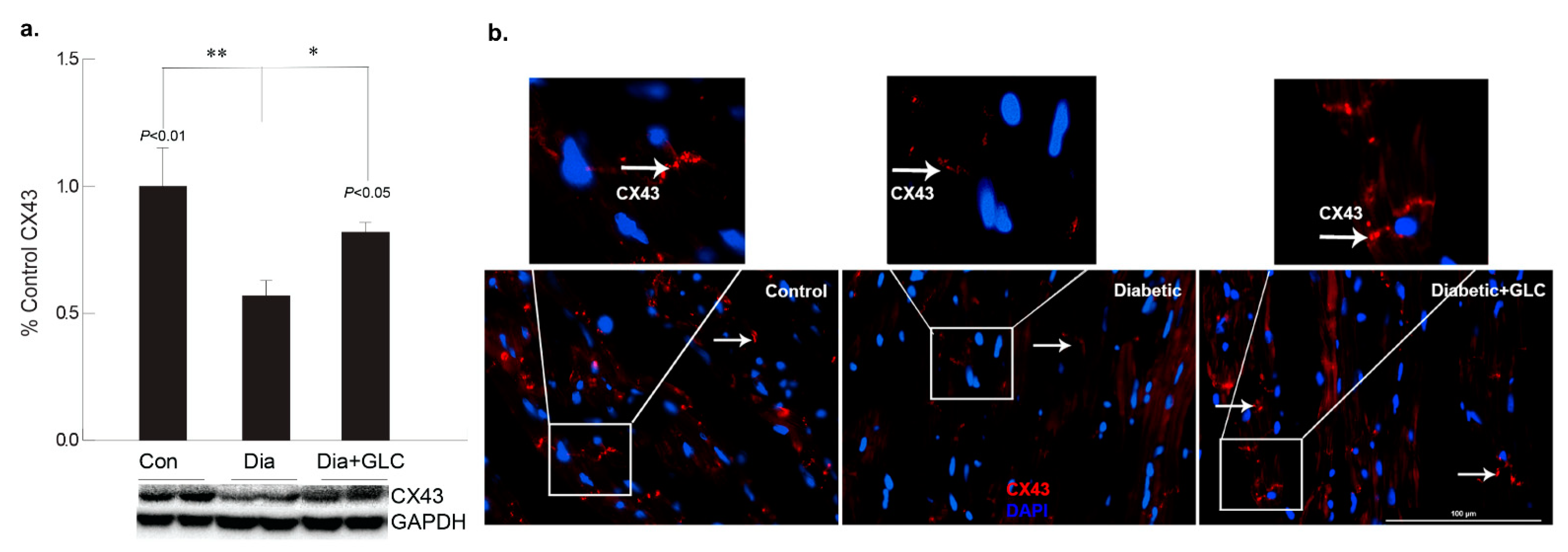
2.6. Immunohistochemistry and Immuno-Histochemical Analysis
2.7. Cell Culture and Treatment
2.8. Statistical Analysis
3. Results
3.1. Type 2 Diabetic Animals Treated with GLC Exhibited a Significant Change in Collagen Deposition without Any Change in Blood Glucose Levels
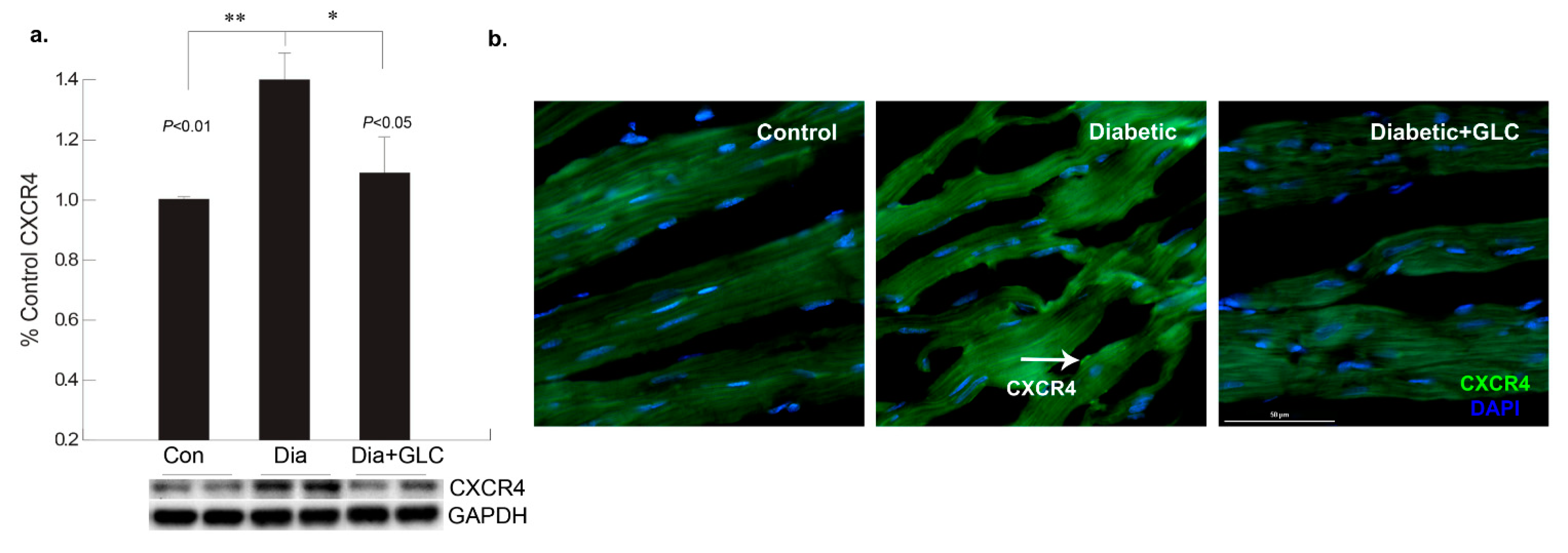
3.2. GLC Altered Chemokine Receptor CXCR4 and Connexin-43 Expression in Cardiac Tissue of Type 2 Diabetic Rats
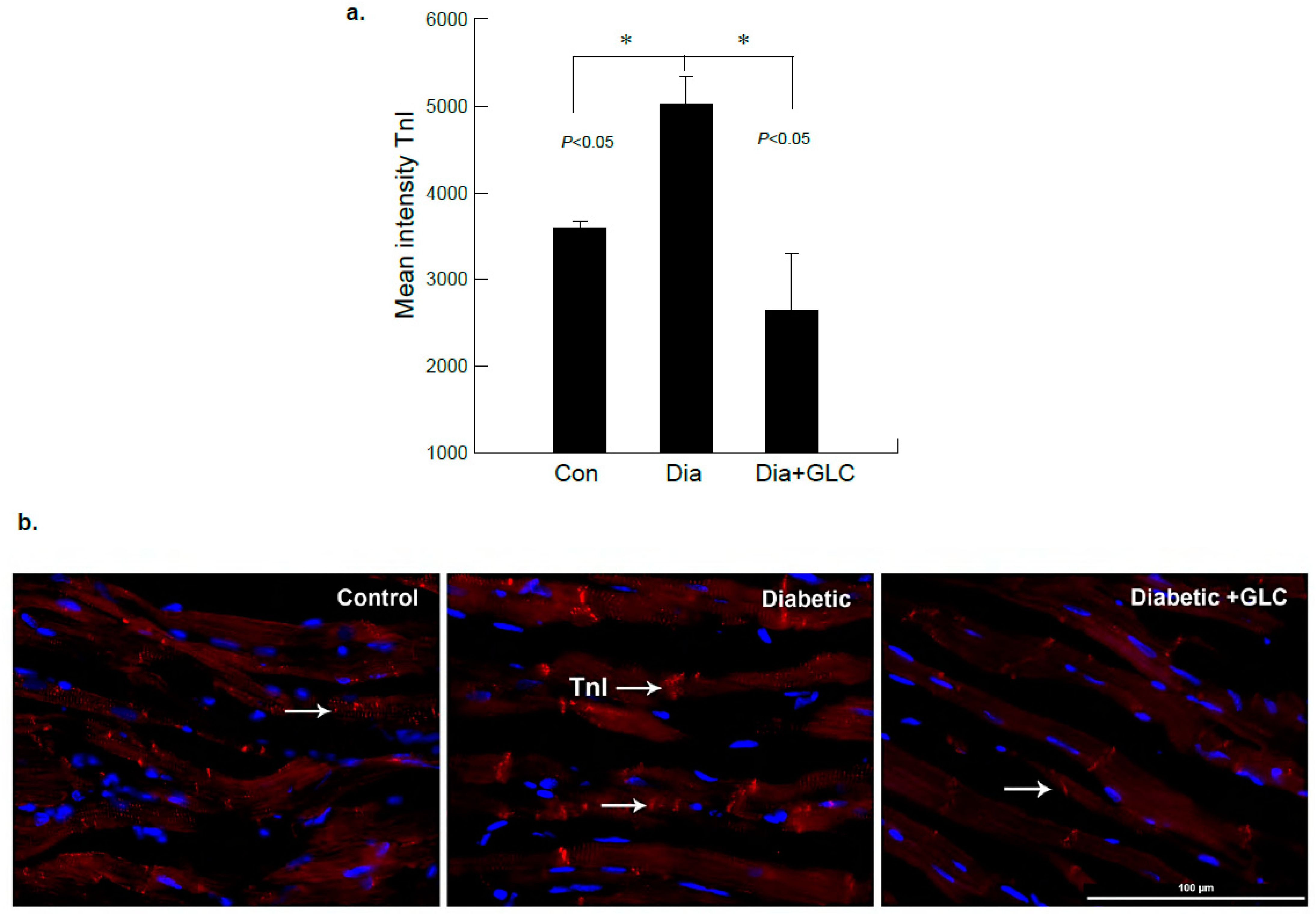
3.3. ZDF Rats Exhibited Increased Troponin-I in the Cardiac Tissue which Was Alleviated with GLC Treatment
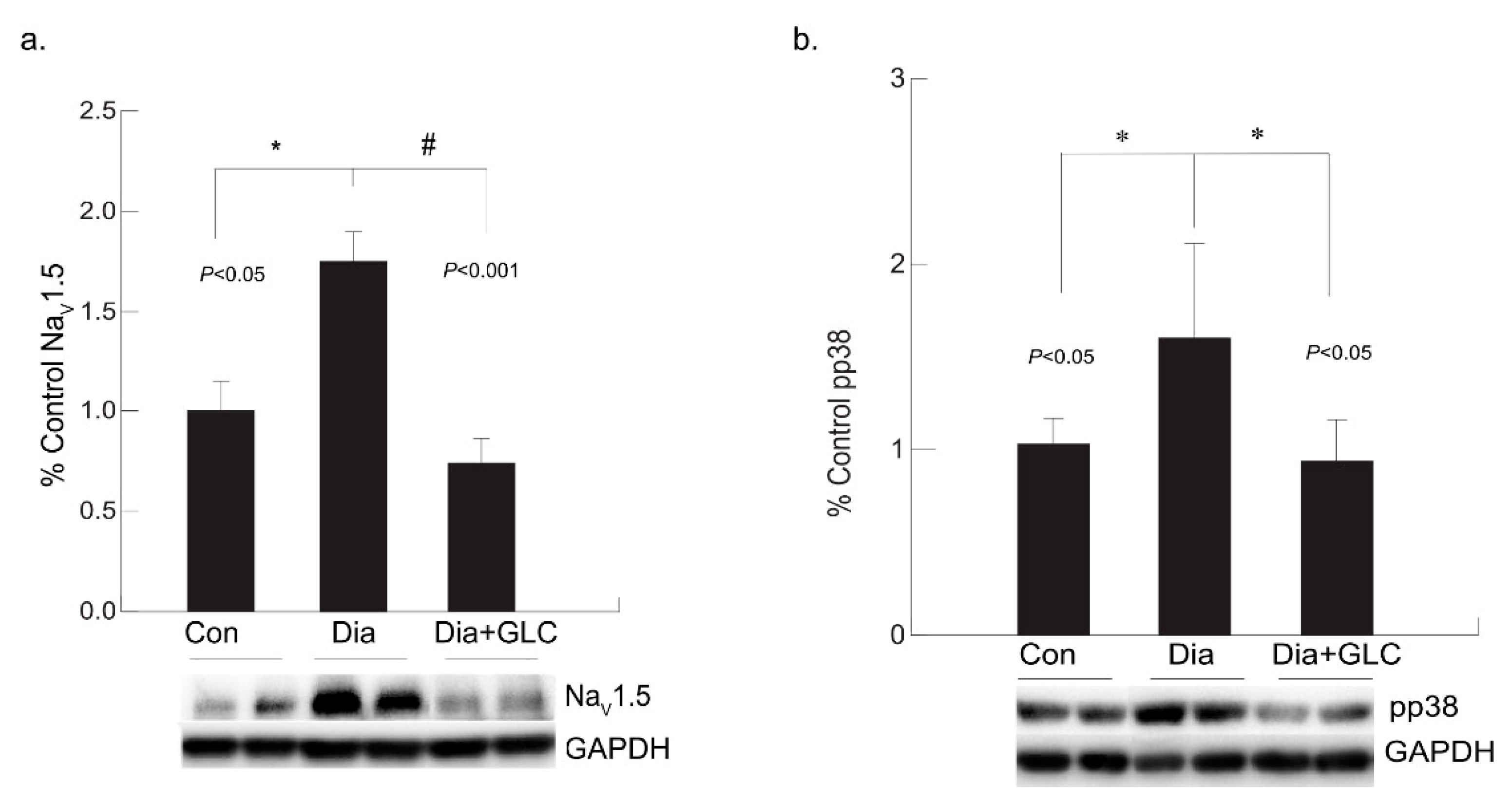
3.4. Diabetic Cardiac Tissue of ZDF Rats Exhibited an Increase in the Expression of Voltage-gated Sodium Channel NaV1.5 and Increased Phosphorylation of p38 MAPK
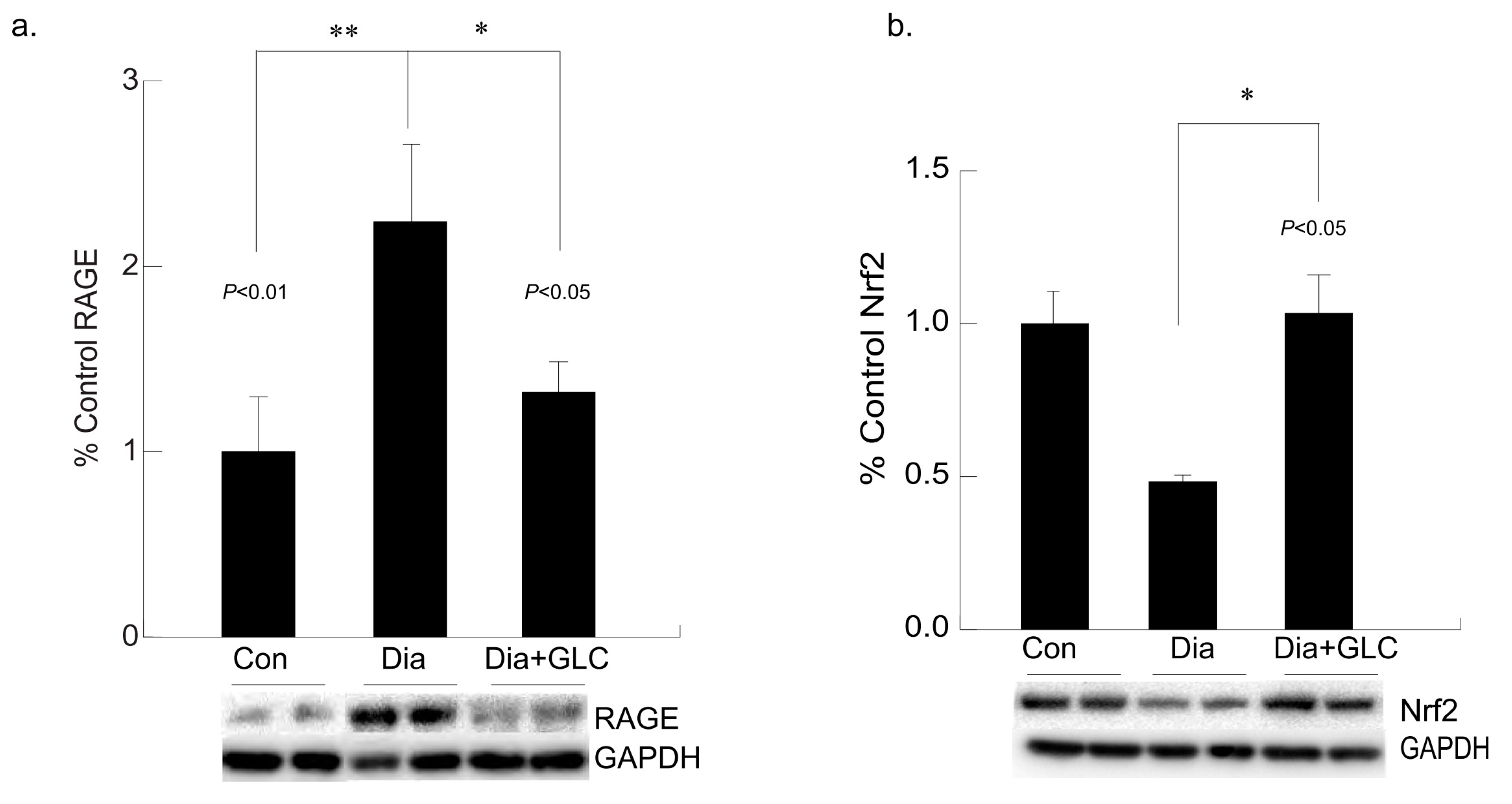
3.5. Hyperglycemia Mediated Expression Levels of Oxidative Stress Marker and Antioxidant Transcription Factor Were Altered with GLC Treatment in Cardiac Tissue of Type 2 Diabetic Animals
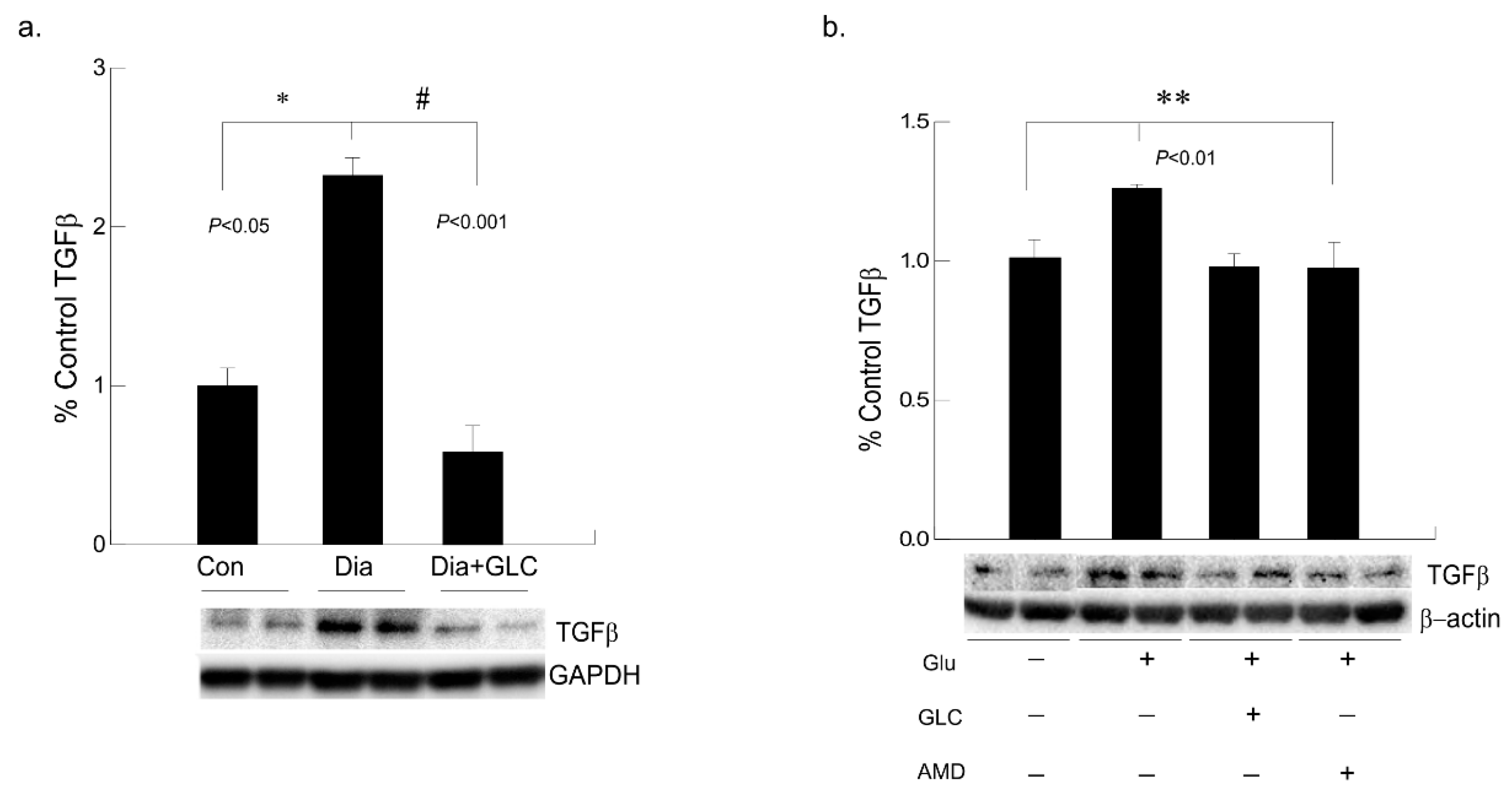
3.6. Alleviation of Hyperglycemia-Mediated Increase in the Expression of Fibrotic Marker TGFβ by GLC Treatment and CXCR4 Antagonist
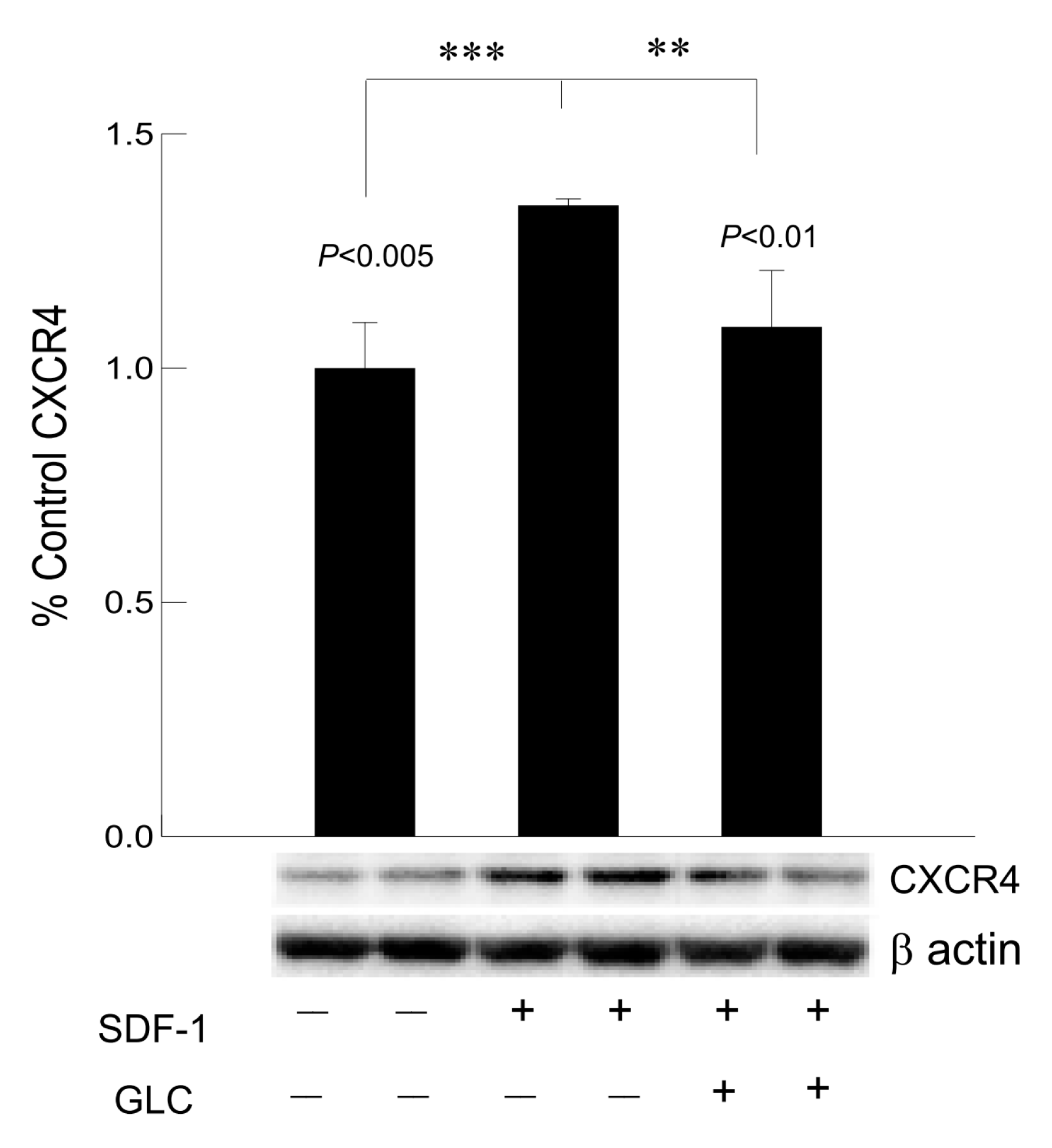
3.7. Hyperglycemia Mediated Increased Expression of Chemokine Receptor CXCR4 Was Mimicked by SDF1 and Treatment with GLC Could Block the Effect in Cardiomyocytes In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CXCR4 | Chemokine C-X-C motif receptor 4 |
| RAGE | Receptor for advanced glycation end products |
| ZDF | Zucker diabetic fatty |
| GLC | Glycyrrhizin |
| TGF-β | Transforming growth factor beta |
| CX43 | Connexin-43 |
| Nrf2 | Nuclear factor erythroid-2-related factor 2 |
| TnI | Troponin I |
References
- Williams, L.J.; Nye, B.G.; Wende, A.R. Diabetes-Related Cardiac Dysfunction. Endocrinol. Metab. 2017, 32, 171–179. [Google Scholar] [CrossRef]
- Zhao, C.T.; Wang, M.; Siu, C.W.; Hou, Y.L.; Wang, T.; Tse, H.F.; Yiu, K.H. Myocardial dysfunction in patients with type 2 diabetes mellitus: Role of endothelial progenitor cells and oxidative stress. Cardiovasc. Diabetol. 2012, 11, 147. [Google Scholar] [CrossRef] [PubMed]
- Stratmann, B.; Tschoepe, D. Heart in Diabetes: Not Only a Macrovascular Disease. Diabetes Care 2011, 34, S138–S144. [Google Scholar] [CrossRef] [PubMed]
- Laakso, M. Heart in Diabetes: A Microvascular Disease. Diabetes Care 2011, 34, S145–S149. [Google Scholar] [CrossRef]
- Gulsin, G.S.; Athithan, L.; McCann, G.P. Diabetic cardiomyopathy: Prevalence, determinants and potential treatments. Ther. Adv. Endocrinol. Metab. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Chiha, M.; Njeim, M.; Chedrawy, E.G. Diabetes and Coronary Heart Disease: A Risk Factor for the Global Epidemic. Int. J. Hypertens. 2012, 2012, 1–7. [Google Scholar] [CrossRef]
- Jia, G.; Hill, M.A.; Sowers, J.R. Diabetic Cardiomyopathy: An Update of Mechanisms Contributing to This Clinical Entity. Circ. Res. 2018, 122, 624–638. [Google Scholar] [CrossRef]
- Marfella, R.; Siniscalchi, M.; Esposito, K.; Sellitto, A.; De Fanis, U.; Romano, C.; Portoghese, M.; Siciliano, S.; Nappo, F.; Sasso, F.C.; et al. Effects of Stress Hyperglycemia on Acute Myocardial Infarction: Role of inflammatory immune process in functional cardiac outcome. Diabetes Care 2003, 26, 3129–3135. [Google Scholar] [CrossRef]
- Pyo, R.T.; Sui, J.; Dhume, A.; Palomeque, J.; Blaxall, B.C.; Diaz, G.; Tunstead, J.; Logothetis, D.E.; Hajjar, R.J.; Schecter, A.D. CXCR4 modulates contractility in adult cardiac myocytes. J. Mol. Cell Cardiol. 2006, 41, 834–844. [Google Scholar] [CrossRef]
- Russo, I.; Frangogiannis, N.G. Diabetes-associated cardiac fibrosis: Cellular effectors, molecular mechanisms and therapeutic opportunities. J. Mol. Cell Cardiol. 2016, 90, 84–93. [Google Scholar] [CrossRef]
- Tao, A.; Song, J.; Lan, T.; Xu, X.; Kvietys, P.; Kao, R.; Martin, C.; Rui, T. Cardiomyocyte-fibroblast interaction contributes to diabetic cardiomyopathy in mice: Role of HMGB1/TLR4/IL-33 axis. Biochim. Biophys. Acta 2015, 1852, 2075–2085. [Google Scholar] [CrossRef]
- Chu, P.Y.; Joshi, M.S.; Horlock, D.; Kiriazis, H.; Kaye, D.M. CXCR4 Antagonism Reduces Cardiac Fibrosis and Improves Cardiac Performance in Dilated Cardiomyopathy. Front. Pharmacol. 2019, 10, 117. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, Z.; Cai, L. Diabetic Cardiomyopathy and Its Prevention by Nrf2: Current Status. Diabetes Metab. J. 2014, 38, 337–345. [Google Scholar] [CrossRef]
- Xu, C.; Liang, C.; Sun, W.; Chen, J.; Chen, X. Glycyrrhizic acid ameliorates myocardial ischemic injury by the regulation of inflammation and oxidative state. Drug Des. Dev. Ther. 2018, 12, 1311–1319. [Google Scholar] [CrossRef]
- Wang, S.; Ding, L.; Ji, H.; Xu, Z.; Liu, Q.; Zheng, Y. The Role of p38 MAPK in the Development of Diabetic Cardiomyopathy. Int. J. Mol. Sci. 2016, 17, 1037. [Google Scholar] [CrossRef]
- Mapanga, R.F.; Essop, M.F. Damaging effects of hyperglycemia on cardiovascular function: Spotlight on glucose metabolic pathways. Am. J. Physiol. Circ. Physiol. 2016, 310, H153–H173. [Google Scholar] [CrossRef]
- Wu, L.; Derynck, R. Essential role of TGF-beta signaling in glucose-induced cell hypertrophy. Dev. Cell 2009, 17, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Candales, A.; Burgos, P.M.H.; Hernandez-Suarez, D.F.; Harris, D. Linking Chronic Inflammation with Cardiovascular Disease: From Normal Aging to the Metabolic Syndrome. S. Pac. J. Nat. Appl. Sci. 2017, 3, e341. [Google Scholar]
- Ruparelia, N.; Chai, J.T.; Fisher, E.A.; Choudhury, R.P. Inflammatory processes in cardiovascular disease: A route to targeted therapies. Nat. Rev. Cardiol. 2017, 14, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Zethelius, B.; Johnston, N.; Venge, P. Troponin I as a predictor of coronary heart disease and mortality in 70-year-old men: A community-based cohort study. Circulation 2006, 113, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Göser, S.; Andrassy, M.; Buss, S.J.; Leuschner, F.; Volz, C.H.; Ottl, R.; Zittrich, S.; Blaudeck, N.; Hardt, S.E.; Pfitzer, G.; et al. Cardiac Troponin I but Not Cardiac Troponin T Induces Severe Autoimmune Inflammation in the Myocardium. Circulation 2006, 114, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.L.; Liang, B.; Wang, X.W.; Fan, F.G.; Jin, J.; Lan, R.; Yang, J.H.; Wang, X.C.; Jin, L.; Cao, Q. Glycyrrhizic acid attenuates CCl(4)-induced hepatocyte apoptosis in rats via a p53-mediated pathway. World J. Gastroenterol. 2013, 19, 3781–3791. [Google Scholar] [CrossRef]
- Liang, B.; Guo, X.-L.; Jin, J.; Ma, Y.-C.; Feng, Z.-Q. Glycyrrhizic acid inhibits apoptosis and fibrosis in carbon-tetrachloride-induced rat liver injury. World J. Gastroenterol. 2015, 21, 5271–5280. [Google Scholar] [CrossRef]
- Rahnama, M.; Mehrabani, D.; Japoni, S.; Edjtehadi, M.; Firoozi, M.S. The healing effect of licorice (Glycyrrhiza glabra) on Helicobacter pylori infected peptic ulcers. J. Res. Med. Sci. 2013, 18, 532–533. [Google Scholar]
- Cai, X.; Wang, X.; Li, J.; Chen, S. Protective effect of glycyrrhizin on myocardial ischemia/reperfusion injury-induced oxidative stress, inducible nitric oxide synthase and inflammatory reactions through high-mobility group box 1 and mitogen-activated protein kinase expression. Exp. Ther. Med. 2017, 14, 1219–1226. [Google Scholar] [CrossRef][Green Version]
- Zhai, C.-L.; Zhang, M.-Q.; Zhang, Y.; Xu, H.-X.; Wang, J.-M.; An, G.-P.; Wang, Y.-Y.; Li, L. Glycyrrhizin protects rat heart against ischemia-reperfusion injury through blockade of HMGB1-dependent phospho-JNK/Bax pathway. Acta Pharmacol. Sin. 2012, 33, 1477–1487. [Google Scholar] [CrossRef]
- Thakur, V.; Sadanandan, J.; Chattopadhyay, M. High-Mobility Group Box 1 Protein Signaling in Painful Diabetic Neuropathy. Int. J. Mol. Sci. 2020, 21, 881. [Google Scholar] [CrossRef]
- Gong, G.; Xiang, L.; Yuan, L.; Hu, L.; Wu, W.; Cai, L.; Yin, L.; Dong, H. Protective Effect of Glycyrrhizin, a Direct HMGB1 Inhibitor, on Focal Cerebral Ischemia/Reperfusion-Induced Inflammation, Oxidative Stress, and Apoptosis in Rats. PLoS ONE 2014, 9, e89450. [Google Scholar] [CrossRef]
- Asleh, R.; Briasoulis, A.; Pereira, N.L.; Edwards, B.S.; Frantz, R.P.; Daly, R.C.; Lerman, A.; Kushwaha, S.S. Hypercholesterolemia after conversion to sirolimus as primary immunosuppression and cardiac allograft vasculopathy in heart transplant recipients. J. Hear. Lung Transplant. 2018, 37, 1372–1380. [Google Scholar] [CrossRef] [PubMed]
- Pignone, M.; Phillips, C.; Mulrow, C. Use of lipid lowering drugs for primary prevention of coronary heart disease: Meta-analysis of randomised trials. BMJ 2000, 321, 983–986. [Google Scholar] [CrossRef] [PubMed]
- Kosmas, C.E.; Silverio, D.; Sourlas, A.; Montan, P.D.; Guzman, E.; Garcia, M.J. Anti-inflammatory therapy for cardiovascular disease. Ann. Transl. Med. 2019, 7, 147. [Google Scholar] [CrossRef] [PubMed]
- Senoner, T.; Dichtl, W. Oxidative Stress in Cardiovascular Diseases: Still a Therapeutic Target? Nutrients 2019, 11, 2090. [Google Scholar] [CrossRef] [PubMed]
- du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar]
- Chattopadhyay, M.; Mata, M.; Fink, D.J. Continuous delta-opioid receptor activation reduces neuronal voltage-gated sodium channel (NaV1.7) levels through activation of protein kinase C in painful diabetic neuropathy. J. Neurosci. 2008, 28, 6652–6658. [Google Scholar] [CrossRef] [PubMed]
- Alonzo, M.; Delgado, M.; Cleetus, C.; Kumar, S.A.; Thakur, V.; Chattopadhyay, M.; Joddar, B. Methods for histological characterization of cryo-induced myocardial infarction in a rat model. Acta. Histochem. 2020, 122, 151624. [Google Scholar] [CrossRef]
- Hegab, Z.; Gibbons, S.; Neyses, L.; Mamas, M.A. Role of advanced glycation end products in cardiovascular disease. World J. Cardiol. 2012, 4, 90–102. [Google Scholar] [CrossRef]
- Lancefield, T.F.; Patel, S.K.; Freeman, M.; Velkoska, E.; Wai, B.; Srivastava, P.M.; Horrigan, M.; Farouque, O.; Burrell, L.M. The Receptor for Advanced Glycation End Products (RAGE) Is Associated with Persistent Atrial Fibrillation. PLoS ONE 2016, 11, e0161715. [Google Scholar] [CrossRef]
- Egaña-Gorroño, L.; López-Díez, R.; Yepuri, G.; Ramirez, L.S.; Reverdatto, S.; Gugger, P.F.; Shekhtman, A.; Ramasamy, R.; Schmidt, A.M. Receptor for Advanced Glycation End Products (RAGE) and Mechanisms and Therapeutic Opportunities in Diabetes and Cardiovascular Disease: Insights From Human Subjects and Animal Models. Front. Cardiovasc. Med. 2020, 7, 37. [Google Scholar] [CrossRef]
- Bangert, A.; Andrassy, M.; Müller, A.-M.; Bockstahler, M.; Fischer, A.; Volz, C.H.; Leib, C.; Göser, S.; Korkmaz-Icöz, S.; Zittrich, S.; et al. Critical role of RAGE and HMGB1 in inflammatory heart disease. Proc. Natl. Acad. Sci. USA 2016, 113, E155–E164. [Google Scholar] [CrossRef]
- Yamagishi, S.; Takeuchi, M.; Inagaki, Y.; Nakamura, K.; Imaizumi, T. Role of advanced glycation end products (AGEs) and their receptor (RAGE) in the pathogenesis of diabetic microangiopathy. Int. J. Clin. Pharmacol. Res. 2003, 23, 129–134. [Google Scholar]
- Yan, S.F.; Ramasamy, R.; Naka, Y.; Schmidt, A.M. Glycation, inflammation, and RAGE: A scaffold for the macrovascular complications of diabetes and beyond. Circ. Res. 2003, 93, 1159–1169. [Google Scholar] [CrossRef]
- Li, C.-J.; Lv, L.; Li, H.; Yu, D.-M. Cardiac fibrosis and dysfunction in experimental diabetic cardiomyopathy are ameliorated by alpha-lipoic acid. Cardiovasc. Diabetol. 2012, 11, 73. [Google Scholar] [CrossRef] [PubMed]
- Westermann, D.; Rutschow, S.; Van Linthout, S.; Linderer, A.; Bücker-Gärtner, C.; Sobirey, M.; Riad, A.; Pauschinger, M.; Schultheiss, H.-P.; Tschöpe, C. Inhibition of p38 mitogen-activated protein kinase attenuates left ventricular dysfunction by mediating pro-inflammatory cardiac cytokine levels in a mouse model of diabetes mellitus. Diabetologia 2006, 49, 2507–2513. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.-Y.; Walder, K.; Horlock, D.; Williams, D.; Nelson, E.; Byrne, M.; Jandeleit-Dahm, K.; Zimmet, P.; Kaye, D.M. CXCR4 Antagonism Attenuates the Development of Diabetic Cardiac Fibrosis. PLoS ONE 2015, 10, e0133616. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Yu, H.; Li, X.; Jin, C.; Zhao, Y.; Xu, S.; Sheng, X. P38 MAPK/miR-1 are involved in the protective effect of EGCG in high glucose-induced Cx43 downregulation in neonatal rat cardiomyocytes. Cell Biol. Int. 2016, 40, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-Y.; Ke, H.-H.; He, Y.; Wen, L.-N.; Xu, W.-Y.; Wu, Z.-F.; Zhao, Y.-M.; Zhong, G.-Q. Transplantation of mesenchymal stem cells modulated Cx43 and Cx45 expression in rats with myocardial infarction. Cytotechnology 2018, 70, 225–234. [Google Scholar] [CrossRef]
- Wright, J.A.; Richards, T.; Becker, D.L. Connexins and Diabetes. Cardiol. Res. Pr. 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- Biernacka, A.; Dobaczewski, M.; Frangogiannis, N.G. TGF-beta signaling in fibrosis. Growth Factors 2011, 29, 196–202. [Google Scholar] [CrossRef]
- Dobaczewski, M.; Chen, W.; Frangogiannis, N.G. Transforming growth factor (TGF)-beta signaling in cardiac remodeling. J. Mol. Cell Cardiol. 2011, 51, 600–606. [Google Scholar] [CrossRef]
- Dobaczewski, M.; Frangogiannis, N.G. Chemokines and cardiac fibrosis. Front. Biosci. Schol. Ed. 2009, 1, 391–405. [Google Scholar] [CrossRef]
- Thakur, V.; Nargis, S.; Gonzalez, M.; Pradhan, S.; Terreros, D.; Chattopadhyay, M. Role of Glycyrrhizin in the Reduction of Inflammation in Diabetic Kidney Disease. Nephron 2017, 137, 137–147. [Google Scholar] [CrossRef]
- Hammarsten, O.; Mair, J.; Möckel, M.; Lindahl, B.; Jaffe, A.S. Possible mechanisms behind cardiac troponin elevations. Biomarkers 2018, 23, 725–734. [Google Scholar] [CrossRef]
- Park, K.C.; Gaze, D.C.; Collinson, P.O.; Marber, M.S. Cardiac troponins: From myocardial infarction to chronic disease. Cardiovasc. Res. 2017, 113, 1708–1718. [Google Scholar] [CrossRef]
- Babuin, L.; Jaffe, A.S. Troponin: The biomarker of choice for the detection of cardiac injury. CMAJ 2005, 173, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- D’Oria, R.; Schipani, R.; Leonardini, A.; Natalicchio, A.; Perrini, S.; Cignarelli, A.; Laviola, L.; Giorgino, F. The Role of Oxidative Stress in Cardiac Disease: From Physiological Response to Injury Factor. Oxidative Med. Cell. Longev. 2020, 2020, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.M.; Maltagliati, A.J. Nrf2 at the heart of oxidative stress and cardiac protection. Physiol. Genom. 2018, 50, 77–97. [Google Scholar] [CrossRef] [PubMed]
- Rook, M.B.; Evers, M.M.; Vos, M.A.; Bierhuizen, M.F. Biology of cardiac sodium channel Nav1.5 expression. Cardiovasc. Res. 2012, 93, 12–23. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thakur, V.; Alcoreza, N.; Delgado, M.; Joddar, B.; Chattopadhyay, M. Cardioprotective Effect of Glycyrrhizin on Myocardial Remodeling in Diabetic Rats. Biomolecules 2021, 11, 569. https://doi.org/10.3390/biom11040569
Thakur V, Alcoreza N, Delgado M, Joddar B, Chattopadhyay M. Cardioprotective Effect of Glycyrrhizin on Myocardial Remodeling in Diabetic Rats. Biomolecules. 2021; 11(4):569. https://doi.org/10.3390/biom11040569
Chicago/Turabian StyleThakur, Vikram, Narah Alcoreza, Monica Delgado, Binata Joddar, and Munmun Chattopadhyay. 2021. "Cardioprotective Effect of Glycyrrhizin on Myocardial Remodeling in Diabetic Rats" Biomolecules 11, no. 4: 569. https://doi.org/10.3390/biom11040569
APA StyleThakur, V., Alcoreza, N., Delgado, M., Joddar, B., & Chattopadhyay, M. (2021). Cardioprotective Effect of Glycyrrhizin on Myocardial Remodeling in Diabetic Rats. Biomolecules, 11(4), 569. https://doi.org/10.3390/biom11040569