Osmolytes: A Possible Therapeutic Molecule for Ameliorating the Neurodegeneration Caused by Protein Misfolding and Aggregation




{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
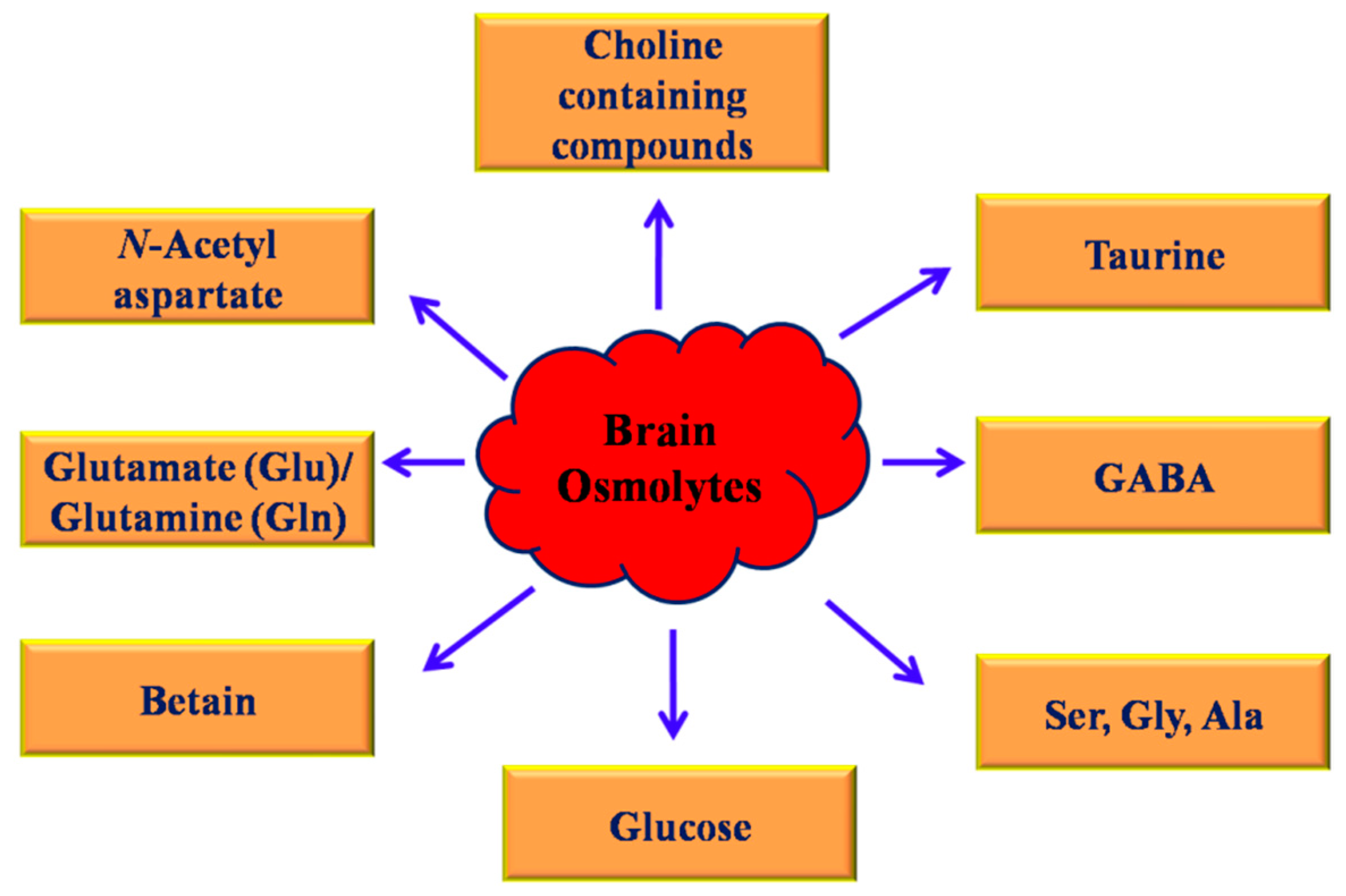
2. Osmolytes
3. Classification of Osmolytes
4. Mechanisms of Actions of Osmolytes
5. Osmolyte Prevents Protein Misfolding, Aggregation, and Fibrillization
6. Osmolytes as Therapeutic Target Against Neurological Disorders
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bantle, C.M.; Phillips, A.T.; Smeyne, R.J.; Rocha, S.M.; Olson, K.E.; Tjalkens, R.B. Infection with mosquito-borne alphavirus induces selective loss of dopaminergic neurons, neuroinflammation and widespread protein aggregation. NPJ Parkinsons Dis. 2019, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Umeda, T.; Ono, K.; Sakai, A.; Yamashita, M.; Mizuguchi, M.; Klein, W.L.; Yamada, M.; Mori, H.; Tomiyama, T. Rifampicin is a candidate preventive medicine against amyloid-beta and tau oligomers. Brain 2016, 139, 1568–1586. [Google Scholar] [CrossRef] [PubMed]
- Seidler, P.M.; Boyer, D.R.; Murray, K.A.; Yang, T.P.; Bentzel, M.; Sawaya, M.R.; Rosenberg, G.; Cascio, D.; Williams, C.K.; Newell, K.L.; et al. Structure-based inhibitors halt prion-like seeding by alzheimer’s disease-and tauopathy-derived brain tissue samples. J. Biol. Chem. 2019, 294, 16451–16464. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.R.; Sweeney, M.D.; Sagare, A.P.; Zlokovic, B.V. Neurovascular dysfunction and neurodegeneration in dementia and alzheimer’s disease. Biochim. Biophys. Acta 2016, 1862, 887–900. [Google Scholar] [CrossRef]
- Chen, G.-F.; Xu, T.-H.; Yan, Y.; Zhou, Y.-R.; Jiang, Y.; Melcher, K.; Xu, H.E. Amyloid beta: Structure, biology and structure-based therapeutic development. Acta Pharmacol. Sin. 2017, 38, 1205–1235. [Google Scholar] [CrossRef]
- Tamagno, E.; Guglielmotto, M.; Monteleone, D.; Manassero, G.; Vasciaveo, V.; Tabaton, M. The unexpected role of abeta1-42 monomers in the pathogenesis of alzheimer’s disease. J. Alzheimers Dis. 2018, 62, 1241–1245. [Google Scholar] [CrossRef]
- Ciechanover, A.; Kwon, Y.T. Degradation of misfolded proteins in neurodegenerative diseases: Therapeutic targets and strategies. Exp. Mol. Med. 2015, 47, e147. [Google Scholar] [CrossRef]
- Cai, Z.; Zhao, B.; Li, K.; Zhang, L.; Li, C.; Quazi, S.H.; Tan, Y. Mammalian target of rapamycin: A valid therapeutic target through the autophagy pathway for alzheimer’s disease? J. Neurosci. Res. 2012, 90, 1105–1118. [Google Scholar] [CrossRef]
- Bhat, M.Y.; Singh, L.R.; Dar, T.A. Trimethylamine n-oxide abolishes the chaperone activity of alpha-casein: An intrinsically disordered protein. Sci. Rep. 2017, 7, 6572. [Google Scholar] [CrossRef]
- Singh, L.R.; Poddar, N.K.; Dar, T.A.; Kumar, R.; Ahmad, F. Protein and DNA destabilization by osmolytes: The other side of the coin. Life Sci. 2011, 88, 117–125. [Google Scholar] [CrossRef]
- Yancey, P.H. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J. Exp. Biol. 2005, 208, 2819–2830. [Google Scholar] [CrossRef]
- Yancey, P.H.; Siebenaller, J.F. Co-evolution of proteins and solutions: Protein adaptation versus cytoprotective micromolecules and their roles in marine organisms. J. Exp. Biol. 2015, 218, 1880–1896. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.H.; Ahmad, N.; Ahmad, F.; Kumar, R. Naturally occurring organic osmolytes: From cell physiology to disease prevention. IUBMB Life 2010, 62, 891–895. [Google Scholar] [CrossRef]
- Yancey, P.H.; Clark, M.E.; Hand, S.C.; Bowlus, R.D.; Somero, G.N. Living with water stress: Evolution of osmolyte systems. Science 1982, 217, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Street, T.O.; Bolen, D.W.; Rose, G.D. A molecular mechanism for osmolyte-induced protein stability. Proc. Natl. Acad. Sci. USA 2006, 103, 13997–14002. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, J.K.; Bhat, R. Thermal stability of proteins in aqueous polyol solutions: Role of the surface tension of water in the stabilizing effect of polyols. J. Phys. Chem. B 1998, 102, 7058–7066. [Google Scholar] [CrossRef]
- Gekko, K.; Timasheff, S.N. Thermodynamic and kinetic examination of protein stabilization by glycerol. Biochemistry 1981, 20, 4677–4686. [Google Scholar] [CrossRef]
- Chandel, T.I.; Khan, M.V.; Khan, R.H. Impact of osmolytes in conformational modulation of protein and its applications in biotechnology. In Cellular Osmolytes; Singh, L.R., Dar, T.A., Eds.; Springer: Singapore, 2017; pp. 143–160. [Google Scholar]
- Knierbein, M.; Venhuis, M.; Held, C.; Sadowski, G. Thermodynamic properties of aqueous osmolyte solutions at high-pressure conditions. Biophys. Chem. 2019, 106211. [Google Scholar] [CrossRef]
- Jaworek, M.W.; Schuabb, V.; Winter, R. The effects of glycine, tmao and osmolyte mixtures on the pressure dependent enzymatic activity of α-chymotrypsin. Phys. Chem. Chem. Phys. 2018, 20, 1347–1354. [Google Scholar] [CrossRef]
- Bachand, G.D.; Jain, R.; Ko, R.; Bouxsein, N.F.; VanDelinder, V. Inhibition of microtubule depolymerization by osmolytes. Biomacromolecules 2018, 19, 2401–2408. [Google Scholar] [CrossRef]
- Powers, E.T.; Morimoto, R.I.; Dillin, A.; Kelly, J.W.; Balch, W.E. Biological and chemical approaches to diseases of proteostasis deficiency. Annu. Rev. Biochem. 2009, 78, 959–991. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.Y.; Timasheff, S.N. Why do some organisms use a urea-methylamine mixture as osmolyte? Thermodynamic compensation of urea and trimethylamine n-oxide interactions with protein. Biochemistry 1994, 33, 12695–12701. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.M.; Liu, Y.; Khan, S.M.; Hou, L.X.; Bolen, D.W. Increased thermal stability of proteins in the presence of naturally occurring osmolytes. Biochemistry 1992, 31, 5278–5283. [Google Scholar] [CrossRef] [PubMed]
- Bajorunaite, E.; Sereikaite, J.; Bumelis, V.-A. L-arginine suppresses aggregation of recombinant growth hormones in refolding process from e. Coli inclusion bodies. Protein J. 2007, 26, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Das, U.; Hariprasad, G.; Ethayathulla, A.S.; Manral, P.; Das, T.K.; Pasha, S.; Mann, A.; Ganguli, M.; Verma, A.K.; Bhat, R.; et al. Inhibition of protein aggregation: Supramolecular assemblies of arginine hold the key. PLoS ONE 2007, 2, e1176. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, E.; Ceman, S. Arginine methylation of rna-binding proteins regulates cell function and differentiation. Mol. Reprod. Dev. 2012, 79, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Berke, S.J.; Paulson, H.L. Protein aggregation and the ubiquitin proteasome pathway: Gaining the upper hand on neurodegeneration. Curr. Opin. Genet. Dev. 2003, 13, 253–261. [Google Scholar] [CrossRef]
- Rani, A.; Venkatesu, P. Changing relations between proteins and osmolytes: A choice of nature. PCCP 2018, 20, 20315–20333. [Google Scholar] [CrossRef]
- Tao, Y.X.; Conn, P.M. Pharmacoperones as novel therapeutics for diverse protein conformational diseases. Physiol. Rev. 2018, 98, 697–725. [Google Scholar] [CrossRef]
- Russo, A.T.; Rösgen, J.; Bolen, D.W. Osmolyte effects on kinetics of fkbp12 c22a folding coupled with prolyl isomerization. J. Mol. Biol. 2003, 330, 851–866. [Google Scholar] [CrossRef]
- Brown, C.R.; Hong-Brown, L.Q.; Biwersi, J.; Verkman, A.S.; Welch, W.J. Chemical chaperones correct the mutant phenotype of the delta f508 cystic fibrosis transmembrane conductance regulator protein. Cell Stress Chaperones 1996, 1, 117–125. [Google Scholar] [CrossRef]
- Edington, B.V.; Whelan, S.A.; Hightower, L.E. Inhibition of heat shock (stress) protein induction by deuterium oxide and glycerol: Additional support for the abnormal protein hypothesis of induction. J. Cell. Physiol. 1989, 139, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Eleutherio, E.C.; Silva, J.T.; Panek, A.D. Identification of an integral membrane 80 kda protein of saccharomyces cerevisiae induced in response to dehydration. Cell Stress Chaperones 1998, 3, 37–43. [Google Scholar] [CrossRef]
- Singh, L.R.; Chen, X.; Kozich, V.; Kruger, W.D. Chemical chaperone rescue of mutant human cystathionine beta-synthase. Mol. Genet. Metab. 2007, 91, 335–342. [Google Scholar] [CrossRef]
- Tatzelt, J.; Prusiner, S.B.; Welch, W.J. Chemical chaperones interfere with the formation of scrapie prion protein. EMBO J. 1996, 15, 6363–6373. [Google Scholar] [CrossRef]
- Tamarappoo, B.K.; Yang, B.; Verkman, A.S. Misfolding of mutant aquaporin-2 water channels in nephrogenic diabetes insipidus. J. Biol. Chem. 1999, 274, 34825–34831. [Google Scholar] [CrossRef]
- Khan, S.; Mueed, Z.; Deval, R.; Rai, P.K.; Prajapati, D.K.; Poddar, N.K. Role of osmolytes in amyloidosis. In Synucleins-Biochemistry and Role in Diseases; Surguchov, A., Ed.; IntechOpen: London, UK, 2019. [Google Scholar]
- Muchowski, P.J.; Wacker, J.L. Modulation of neurodegeneration by molecular chaperones. Nat. Rev. Neurosci. 2005, 6, 11–22. [Google Scholar] [CrossRef]
- Inayathullah, M.; Rajadas, J. Effect of osmolytes on the conformation and aggregation of some amyloid peptides: Cd spectroscopic data. Data Brief 2016, 7, 1643–1651. [Google Scholar] [CrossRef][Green Version]
- Choudhary, S.; Kishore, N. Addressing mechanism of fibrillization/aggregation and its prevention in presence of osmolytes: Spectroscopic and calorimetric approach. PLoS ONE 2014, 9, e104600. [Google Scholar] [CrossRef]
- Mishra, R.; Seckler, R.; Bhat, R. Efficient refolding of aggregation-prone citrate synthase by polyol osmolytes: How well are protein folding and stability aspects coupled? J. Biol. Chem. 2005, 280, 15553–15560. [Google Scholar] [CrossRef]
- Mittal, S.; Singh, L.R. Macromolecular crowding decelerates aggregation of a beta-rich protein, bovine carbonic anhydrase: A case study. J. Biochem. 2014, 156, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Ignatova, Z.; Gierasch, L.M. Inhibition of protein aggregation in vitro and in vivo by a natural osmoprotectant. Proc. Natl. Acad. Sci. USA 2006, 103, 13357–13361. [Google Scholar] [CrossRef]
- Walker, F.O. Huntington’s disease. Lancet 2007, 369, 218–228. [Google Scholar] [CrossRef]
- Tanaka, M.; Machida, Y.; Niu, S.; Ikeda, T.; Jana, N.R.; Doi, H.; Kurosawa, M.; Nekooki, M.; Nukina, N. Trehalose alleviates polyglutamine-mediated pathology in a mouse model of huntington disease. Nat. Med. 2004, 10, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Barkhordarian, H.; Emadi, S.; Park, C.B.; Sierks, M.R. Trehalose differentially inhibits aggregation and neurotoxicity of beta-amyloid 40 and 42. Neurobiol. Dis. 2005, 20, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Nagata, M.; Monji, A.; Yoshida, I.; Tashiro, N.; Imoto, T. Effect of sucrose on formation of the beta-amyloid fibrils and d-aspartic acids in abeta 1-42. Biol. Pharm. Bull. 2002, 25, 375–378. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tomiyama, T.; Asano, S.; Furiya, Y.; Shirasawa, T.; Endo, N.; Mori, H. Racemization of asp23 residue affects the aggregation properties of alzheimer amyloid beta protein analogues. J. Biol. Chem. 1994, 269, 10205–10208. [Google Scholar]
- Papadopoulos, M.C.; Verkman, A.S. Aquaporin water channels in the nervous system. Nat. Rev. Neurosci. 2013, 14, 265–277. [Google Scholar] [CrossRef]
- Woo, J.; Kim, J.E.; Im, J.J.; Lee, J.; Jeong, H.S.; Park, S.; Jung, S.Y.; An, H.; Yoon, S.; Lim, S.M.; et al. Astrocytic water channel aquaporin-4 modulates brain plasticity in both mice and humans: A potential gliogenetic mechanism underlying language-associated learning. Mol. Psychiatry 2018, 23, 1021–1030. [Google Scholar] [CrossRef]
- Nilius, B.; Eggermont, J.; Voets, T.; Buyse, G.; Manolopoulos, V.; Droogmans, G. Properties of volume-regulated anion channels in mammalian cells. Prog. Biophys. Mol. Biol. 1997, 68, 69–119. [Google Scholar] [CrossRef]
- Parkerson, K.A.; Sontheimer, H. Biophysical and pharmacological characterization of hypotonically activated chloride currents in cortical astrocytes. Glia 2004, 46, 419–436. [Google Scholar] [CrossRef]
- Hazama, A.; Okada, Y. Ca2+ sensitivity of volume-regulatory k+ and cl- channels in cultured human epithelial cells. J. Physiol. 1988, 402, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.E.; Kwon, J.; Won, J.; An, H.; Jang, M.W.; Woo, J.; Lee, J.S.; Park, M.G.; Yoon, B.E.; Lee, S.E.; et al. Tweety-homolog (ttyh) family encodes the pore-forming subunits of the swelling-dependent volume-regulated anion channel (vracswell) in the brain. Exp. Neurobiol. 2019, 28, 183–215. [Google Scholar] [CrossRef]
- Haussinger, D.; Schliess, F. Pathogenetic mechanisms of hepatic encephalopathy. Gut 2008, 57, 1156–1165. [Google Scholar] [CrossRef]
- Knight, L.S.; Piibe, Q.; Lambie, I.; Perkins, C.; Yancey, P.H. Betaine in the brain: Characterization of betaine uptake, its influence on other osmolytes and its potential role in neuroprotection from osmotic stress. Neurochem. Res. 2017, 42, 3490–3503. [Google Scholar] [CrossRef] [PubMed]
- Oeltzschner, G.; Butz, M.; Baumgarten, T.J.; Hoogenboom, N.; Wittsack, H.-J.; Schnitzler, A. Low visual cortex gaba levels in hepatic encephalopathy: Links to blood ammonia, critical flicker frequency, and brain osmolytes. Metab. Brain Dis. 2015, 30, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Atamna, H. Therapeutic approaches to delay the onset of alzheimer’s disease. J. Aging Res. 2011, 2011, 820903. [Google Scholar] [CrossRef]
- Haass, C.; Selkoe, D.J. Soluble protein oligomers in neurodegeneration: Lessons from the alzheimer’s amyloid beta-peptide. Nat. Rev. Mol. Cell Biol. 2007, 8, 101–112. [Google Scholar] [CrossRef]
- Scherzinger, E.; Lurz, R.; Turmaine, M.; Mangiarini, L.; Hollenbach, B.; Hasenbank, R.; Bates, G.P.; Davies, S.W.; Lehrach, H.; Wanker, E.E. Huntingtin-encoded polyglutamine expansions form amyloidlike protein aggregates in vitro and in vivo. Cell 1997, 90, 549–558. [Google Scholar] [CrossRef]
- Davies, S.W.; Turmaine, M.; Cozens, B.A.; DiFiglia, M.; Sharp, A.H.; Ross, C.A.; Scherzinger, E.; Wanker, E.E.; Mangiarini, L.; Bates, G.P. Formation of neuronal intranuclear inclusions underlies the neurological dysfunction in mice transgenic for the HD mutation. Cell 1997, 90, 537–548. [Google Scholar] [CrossRef]
- Chen, M.; Ona, V.O.; Li, M.; Ferrante, R.J.; Fink, K.B.; Zhu, S.; Bian, J.; Guo, L.; Farrell, L.A.; Hersch, S.M.; et al. Minocycline inhibits caspase-1 and caspase-3 expression and delays mortality in a transgenic mouse model of Huntington disease. Nat. Med. 2000, 6, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, R.J.; Andreassen, O.A.; Jenkins, B.G.; Dedeoglu, A.; Kuemmerle, S.; Kubilus, J.K.; Kaddurah-Daouk, R.; Hersch, S.M.; Beal, M.F. Neuroprotective effects of creatine in a transgenic mouse model of Huntington’s disease. J. Neurosci. 2000, 20, 4389–4397. [Google Scholar] [CrossRef] [PubMed]
- Hockly, E.; Richon, V.M.; Woodman, B.; Smith, D.L.; Zhou, X.; Rosa, E.; Sathasivam, K.; Ghazi-Noori, S.; Mahal, A.; Lowden, P.A.; et al. Suberoylanilide hydroxamic acid, a histone deacetylase inhibitor, ameliorates motor deficits in a mouse model of Huntington’s disease. Proc. Natl. Acad. Sci. USA 2003, 100, 2041–2046. [Google Scholar] [CrossRef]
- Hughes, R.E.; Olson, J.M. Therapeutic opportunities in polyglutamine disease. Nat. Med. 2001, 7, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Warrick, J.M.; Paulson, H.L.; Gray-Board, G.L.; Bui, Q.T.; Fischbeck, K.H.; Pittman, R.N.; Bonini, N.M. Expanded polyglutamine protein forms nuclear inclusions and causes neural degeneration in drosophila. Cell 1998, 93, 939–949. [Google Scholar] [CrossRef]
- Soto, C. Protein misfolding and disease; protein refolding and therapy. FEBS Lett. 2001, 498, 204–207. [Google Scholar] [CrossRef]
- Chow, M.K.; Devlin, G.L.; Bottomley, S.P. Osmolytes as modulators of conformational changes in serpins. Biol. Chem. 2001, 382, 1593–1599. [Google Scholar] [CrossRef]
- Dobson, C.M. Protein folding and disease: A view from the first horizon symposium. Nat. Rev. Drug Discov. 2003, 2, 154–160. [Google Scholar] [CrossRef]
- Bolen, D.W.; Baskakov, I.V. The osmophobic effect: Natural selection of a thermodynamic force in protein folding. J. Mol. Biol. 2001, 310, 955–963. [Google Scholar] [CrossRef]
- Ebel, C.; Eisenberg, H.; Ghirlando, R. Probing protein-sugar interactions. Biophys. J. 2000, 78, 385–393. [Google Scholar] [CrossRef]
- Wendorf, J.R.; Radke, C.J.; Blanch, H.W. Reduced protein adsorption at solid interfaces by sugar excipients. Biotechnol. Bioeng. 2004, 87, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Davis-Searles, P.R.; Saunders, A.J.; Erie, D.A.; Winzor, D.J.; Pielak, G.J. Interpreting the effects of small uncharged solutes on protein-folding equilibria. Annu. Rev. Biophys. Biomol. Struct. 2001, 30, 271–306. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kushwah, N.; Jain, V.; Yadav, D. Osmolytes: A Possible Therapeutic Molecule for Ameliorating the Neurodegeneration Caused by Protein Misfolding and Aggregation. Biomolecules 2020, 10, 132. https://doi.org/10.3390/biom10010132
Kushwah N, Jain V, Yadav D. Osmolytes: A Possible Therapeutic Molecule for Ameliorating the Neurodegeneration Caused by Protein Misfolding and Aggregation. Biomolecules. 2020; 10(1):132. https://doi.org/10.3390/biom10010132
Chicago/Turabian StyleKushwah, Neetu, Vishal Jain, and Dhananjay Yadav. 2020. "Osmolytes: A Possible Therapeutic Molecule for Ameliorating the Neurodegeneration Caused by Protein Misfolding and Aggregation" Biomolecules 10, no. 1: 132. https://doi.org/10.3390/biom10010132
APA StyleKushwah, N., Jain, V., & Yadav, D. (2020). Osmolytes: A Possible Therapeutic Molecule for Ameliorating the Neurodegeneration Caused by Protein Misfolding and Aggregation. Biomolecules, 10(1), 132. https://doi.org/10.3390/biom10010132