Eicosanoid Profile of Influenza A Virus Infected Pigs



{kind=link}
{kind=link}
Abstract
1. Introduction
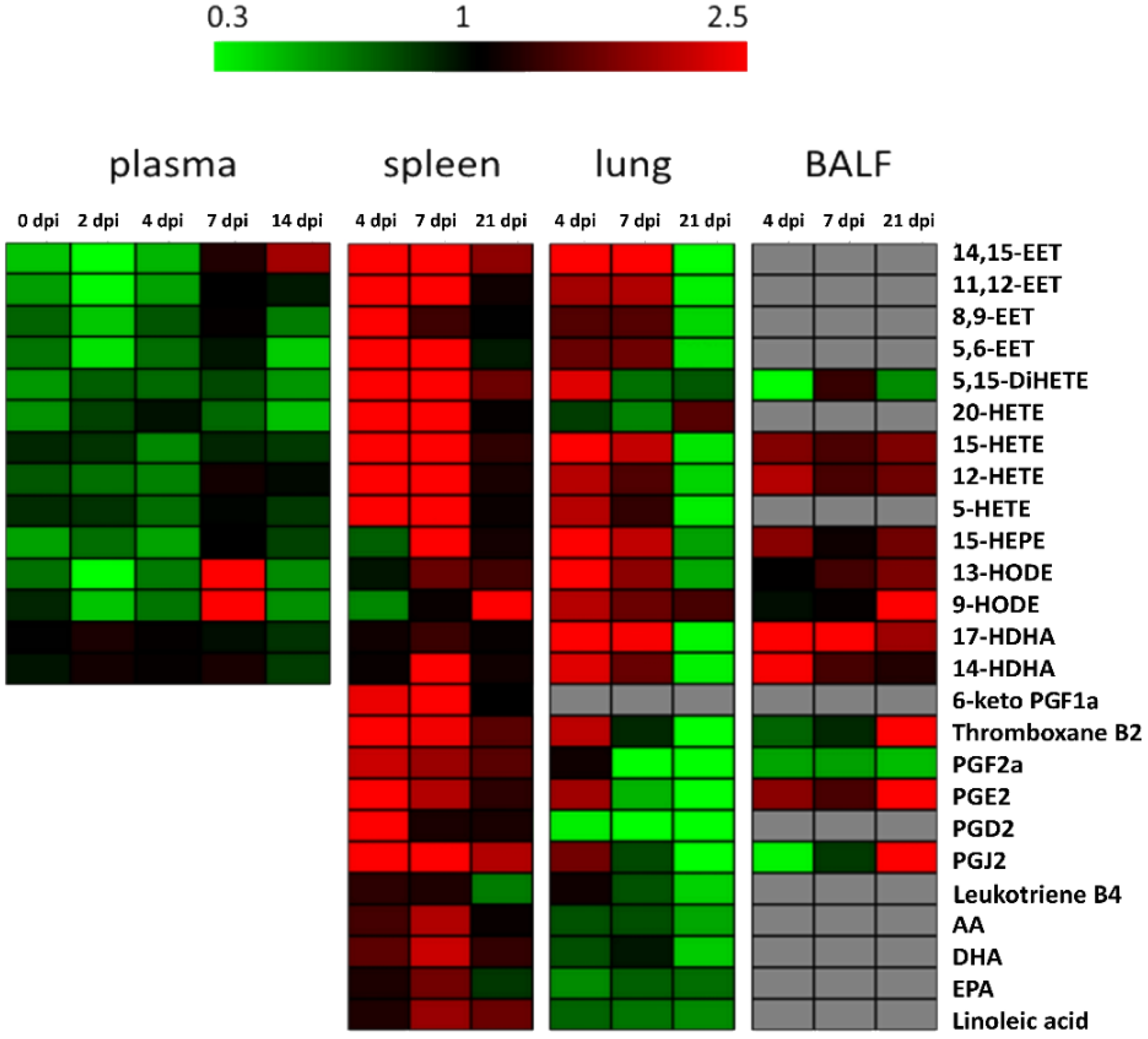
2. Results
2.1. Virus Infection
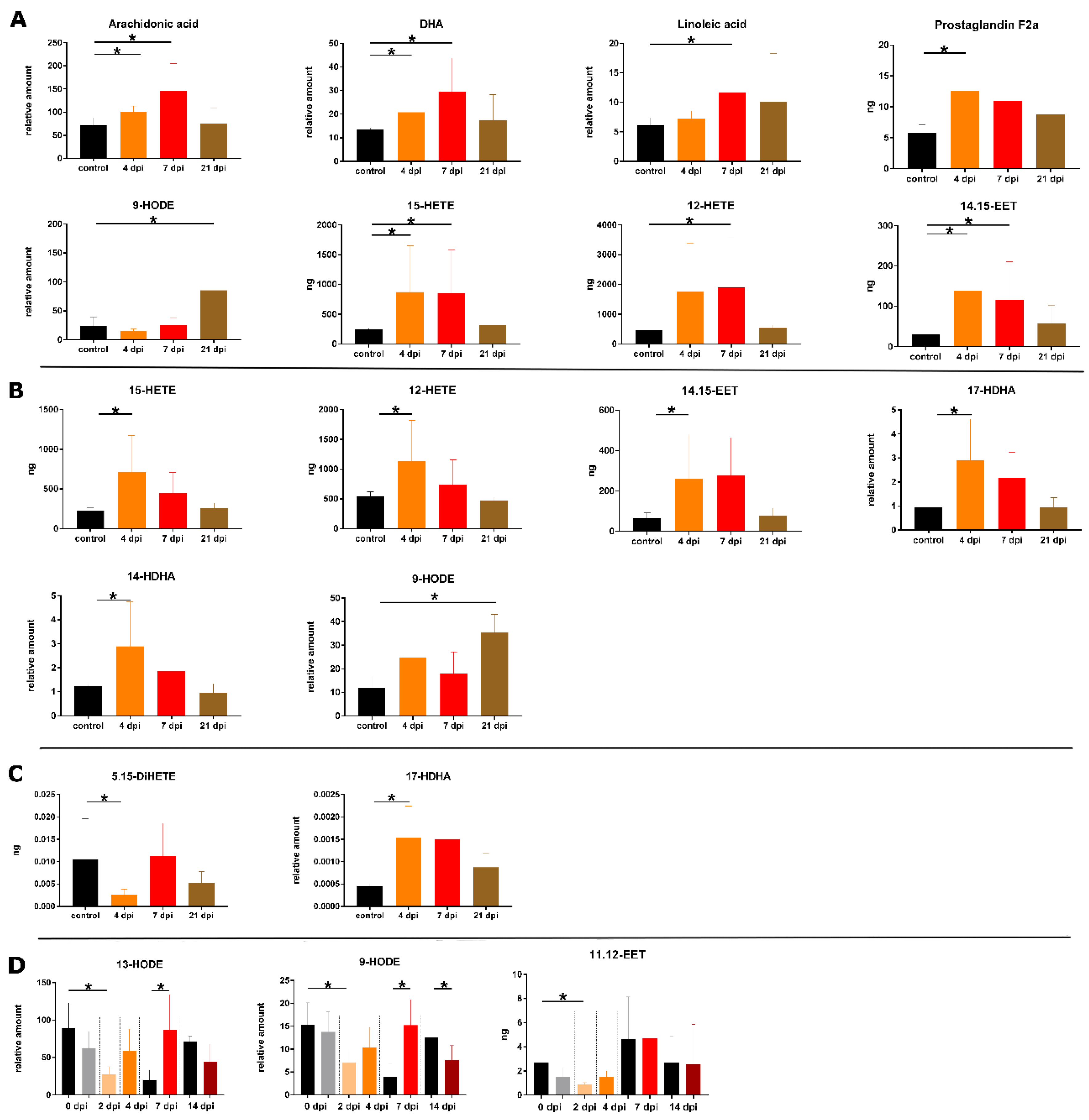
2.2. Spleen
2.3. Lung
2.4. BALF
2.5. Blood Plasma
3. Discussion
3.1. Eicosanoid Pattern of IAV-Infected Pigs Compared to Other Animal Infection Models
3.2. Sample Material of Choice To Unravel Changes in the Eicosanoid Profile
4. Materials and Methods
4.1. Chemicals and Standards
4.2. Cells and Virus
4.3. Pig Infection
4.4. Eicosanoid Extraction
4.5. Eicosanoid Measurement
4.6. Statistics and Data Visualization
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Collaborators, G.B.D.L.R.I. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of lower respiratory infections in 195 countries, 1990-2016: A systematic analysis for the global burden of disease study 2016. Lancet Infect. Dis. 2018, 18, 1191–1210. [Google Scholar]
- McCullers, J.A. The co-pathogenesis of influenza viruses with bacteria in the lung. Nat. Rev. Microbiol. 2014, 12, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Brundage, J.F.; Shanks, G.D. Deaths from bacterial pneumonia during 1918-19 influenza pandemic. Emerg. Infect. Dis. 2008, 14, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Rice, T.W.; Rubinson, L.; Uyeki, T.M.; Vaughn, F.L.; John, B.B.; Miller, R.R., 3rd; Higgs, E.; Randolph, A.G.; Smoot, B.E.; Thompson, B.T.; et al. Critical illness from 2009 pandemic influenza a virus and bacterial coinfection in the united states. Crit. Care Med. 2012, 40, 1487–1498. [Google Scholar] [CrossRef] [PubMed]
- Scholtissek, C.; Burger, H.; Kistner, O.; Shortridge, K.F. The nucleoprotein as a possible major factor in determining host specificity of influenza h3n2 viruses. Virology 1985, 147, 287–294. [Google Scholar] [CrossRef]
- Mohiuddin, M.M.; Reichart, B.; Byrne, G.W.; McGregor, C.G.A. Current status of pig heart xenotransplantation. Int. J. Surg. 2015, 23, 234–239. [Google Scholar] [CrossRef]
- Niemann, H.; Petersen, B. The production of multi-transgenic pigs: Update and perspectives for xenotransplantation. Transgenic Res. 2016, 25, 361–374. [Google Scholar] [CrossRef]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The pig: A model for human infectious diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef]
- Summerfield, A. Special issue on porcine immunology: An introduction from the guest editor. Dev. Comp. Immunol. 2009, 33, 265–266. [Google Scholar] [CrossRef]
- Summerfield, A.; McCullough, K.C. The porcine dendritic cell family. Dev. Comp. Immunol. 2009, 33, 299–309. [Google Scholar] [CrossRef]
- Sanchez, E.L.; Lagunoff, M. Viral activation of cellular metabolism. Virology 2015, 479, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Ritter, J.B.; Wahl, A.S.; Freund, S.; Genzel, Y.; Reichl, U. Metabolic effects of influenza virus infection in cultured animal cells: Intra- and extracellular metabolite profiling. BMC Syst. Biol. 2010, 4. [Google Scholar] [CrossRef] [PubMed]
- Masoodi, M.; Eiden, M.; Koulman, A.; Spaner, D.; Volmer, D.A. Comprehensive lipidomics analysis of bioactive lipids in complex regulatory networks. Anal. Chem. 2010, 82, 8176–8185. [Google Scholar] [CrossRef] [PubMed]
- Arita, M. Mediator lipidomics in acute inflammation and resolution. J. Biochem. 2012, 152, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef]
- Sugimoto, M.A.; Sousa, L.P.; Pinho, V.; Perretti, M.; Teixeira, M.M. Resolution of inflammation: What controls its onset? Front. Immunol. 2016, 7. [Google Scholar] [CrossRef]
- Fan, F.; Muroya, Y.; Roman, R.J. Cytochrome p450 eicosanoids in hypertension and renal disease. Curr. Opin. Nephrol. Hypertens. 2015, 24, 37–46. [Google Scholar] [CrossRef]
- Wagner, K.; Vito, S.; Inceoglu, B.; Hammock, B.D. The role of long chain fatty acids and their epoxide metabolites in nociceptive signaling. Prostaglandins Other Lipid Mediat. 2014, 113, 2–12. [Google Scholar] [CrossRef]
- Holtzman, M.J. Arachidonic acid metabolism in airway epithelial cells. Annu. Rev. Physiol. 1992, 54, 303–329. [Google Scholar] [CrossRef]
- Tam, V.C. Lipidomic profiling of bioactive lipids by mass spectrometry during microbial infections. Semin. Immunol. 2013, 25, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Spector, A.A.; Fang, X.; Snyder, G.D.; Weintraub, N.L. Epoxyeicosatrienoic acids (eets): Metabolism and biochemical function. Prog. Lipid Res. 2004, 43, 55–90. [Google Scholar] [CrossRef]
- Obinata, H.; Izumi, T. G2a as a receptor for oxidized free fatty acids. Prostaglandins Other Lipid Mediat. 2009, 89, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Kronke, G.; Katzenbeisser, J.; Uderhardt, S.; Zaiss, M.M.; Scholtysek, C.; Schabbauer, G.; Zarbock, A.; Koenders, M.I.; Axmann, R.; Zwerina, J.; et al. 12/15-lipoxygenase counteracts inflammation and tissue damage in arthritis. J. Immunol. 2009, 183, 3383–3389. [Google Scholar] [CrossRef] [PubMed]
- Wray, J.A.; Sugden, M.C.; Zeldin, D.C.; Greenwood, G.K.; Samsuddin, S.; Miller-Degraff, L.; Bradbury, J.A.; Holness, M.J.; Warner, T.D.; Bishop-Bailey, D. The epoxygenases cyp2j2 activates the nuclear receptor ppar alpha in vitro and in vivo. Plos ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.; Sirois, M.; Echave, V.; Gomes, M.M.; Rousseau, E. Eet displays anti-inflammatory effects in tnf-alpha stimulated human bronchi: Putative role of cpi-17. Am. J. Respir. Cell Mol. Biol. 2008, 38, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Kuba, K.; Ichikawa, A.; Nakayama, M.; Katahira, J.; Iwamoto, R.; Watanebe, T.; Sakabe, S.; Daidoji, T.; Nakamura, S.; et al. The lipid mediator protectin d1 inhibits influenza virus replication and improves severe influenza. Cell 2013, 153, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Tam, V.C.; Quehenberger, O.; Oshansky, C.M.; Suen, R.; Armando, A.M.; Treuting, P.M.; Thomas, P.G.; Dennis, E.A.; Aderem, A. Lipidomic profiling of influenza infection identifies mediators that induce and resolve inflammation. Cell 2013, 154, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Ramon, S.; Baker, S.F.; Sahler, J.M.; Kim, N.; Feldsott, E.A.; Serhan, C.N.; Martinez-Sobrido, L.; Topham, D.J.; Phipps, R.P. The specialized proresolving mediator 17-hdha enhances the antibody-mediated immune response against influenza virus: A new class of adjuvant? J. Immunol. 2014, 193, 6031–6040. [Google Scholar] [CrossRef]
- Morita, E.; Schroder, J.M.; Christophers, E. Identification of a novel and highly potent eosinophil chemotactic lipid in human eosinophils treated with arachidonic-acid. J. Immunol. 1990, 144, 1893–1900. [Google Scholar] [PubMed]
- Tejera, N.; Boeglin, W.E.; Suzuki, T.; Schneider, C. Cox-2-dependent and -independent biosynthesis of dihydroxy-arachidonic acids in activated human leukocytes. J. Lipid Res. 2012, 53, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Chavis, C.; Chanez, P.; Vachier, I.; Bousquet, J.; Michel, F.B.; Godard, P. 5-15-dihete and lipoxins generated by neutrophils from endogenous arachidonic-acid as asthma biomarkers. Biochem. Biophys. Res. Commun. 1995, 207, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Leroux, J.L.; Blotman, F.; Chavis, C. Conversion of endogenous arachidonic-acid to 5,15-dihete and lipoxins by polymorphonuclear cells from patients with rheumatoid-arthritis. Inflamm. Res. 1995, 44, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Altmann, R.; Hausmann, M.; Spottl, T.; Gruber, M.; Bull, A.W.; Menzel, K.; Vogl, D.; Herfarth, H.; Scholmerich, J.; Falk, W.; et al. 13-oxo-ode is an endogenous ligand for ppargamma in human colonic epithelial cells. Biochem. Pharmacol. 2007, 74, 612–622. [Google Scholar] [CrossRef] [PubMed]
- Schwaiger, T.; Sehl, J.; Karte, C.; Schäfer, A.; Hühr, J.; Mettenleiter, T.C.; Schröder, C.; Ulrich, R.; Köllner, B.; Blohm, U. Minor clinic, major immunology–experimental h1n1pdm09 infection in pigs mimics human seasonal influenza infections. 2019. Unpublished manuscript. [Google Scholar]
- Balas, L.; Guichardant, M.; Durand, T.; Lagarde, M. Confusion between protectin d1 (pd1) and its isomer protectin dx (pdx). An overview on the dihydroxy-docosatrienes described to date. Biochimie 2014, 99, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pace, S.; Rossi, A.; Krauth, V.; Dehm, F.; Troisi, F.; Bilancia, R.; Weinigel, C.; Rummler, S.; Werz, O.; Sautebin, L. Sex differences in prostaglandin biosynthesis in neutrophils during acute inflammation. Sci. Rep. 2017, 7, 3759. [Google Scholar] [CrossRef] [PubMed]
- Mahy, B.W.J.; Kangro, H.O. Virology methods manual; Academic Press: London, UK, 1996. [Google Scholar]
- Traxler, S.; Bischoff, A.C.; Sass, R.; Trefz, P.; Gierschner, P.; Brock, B.; Schwaiger, T.; Karte, C.; Blohm, U.; Schroder, C.; et al. Voc breath profile in spontaneously breathing awake swine during influenza a infection. Sci. Rep. 2018, 8, 14857. [Google Scholar] [CrossRef] [PubMed]
- Spackman, E.; Senne, D.A.; Myers, T.J.; Bulaga, L.L.; Garber, L.P.; Perdue, M.L.; Lohman, K.; Daum, L.T.; Suarez, D.L. Development of a real-time reverse transcriptase pcr assay for type a influenza virus and the avian h5 and h7 hemagglutinin subtypes. J. Clin. Microbiol. 2002, 40, 3256–3260. [Google Scholar] [CrossRef] [PubMed]
- Rivera, J.; Ward, N.; Hodgson, J.; Puddey, I.B.; Falck, J.R.; Croft, K.D. Measurement of 20-hydroxyeicosatetraenoic acid in human urine by gas chromatography-mass spectrometry. Clin. Chem. 2004, 50, 224–226. [Google Scholar] [CrossRef] [PubMed]
- Gomolka, B.; Siegert, E.; Blossey, K.; Schunck, W.H.; Rothe, M.; Weylandt, K.H. Analysis of omega-3 and omega-6 fatty acid-derived lipid metabolite formation in human and mouse blood samples. Prostaglandins Other Lipid Mediat. 2011, 94, 81–87. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schultz, D.; Methling, K.; KoInfekt Study Group; Rothe, M.; Lalk, M. Eicosanoid Profile of Influenza A Virus Infected Pigs. Metabolites 2019, 9, 130. https://doi.org/10.3390/metabo9070130
Schultz D, Methling K, KoInfekt Study Group, Rothe M, Lalk M. Eicosanoid Profile of Influenza A Virus Infected Pigs. Metabolites. 2019; 9(7):130. https://doi.org/10.3390/metabo9070130
Chicago/Turabian StyleSchultz, Daniel, Karen Methling, KoInfekt Study Group, Michael Rothe, and Michael Lalk. 2019. "Eicosanoid Profile of Influenza A Virus Infected Pigs" Metabolites 9, no. 7: 130. https://doi.org/10.3390/metabo9070130
APA StyleSchultz, D., Methling, K., KoInfekt Study Group, Rothe, M., & Lalk, M. (2019). Eicosanoid Profile of Influenza A Virus Infected Pigs. Metabolites, 9(7), 130. https://doi.org/10.3390/metabo9070130