
Crosstalk between Acidosis and Iron Metabolism: Data from In Vivo Studies



Abstract
1. Introduction
2. Results
2.1. Both ATP4A and ATP4B Are Stimulated by Acidosis
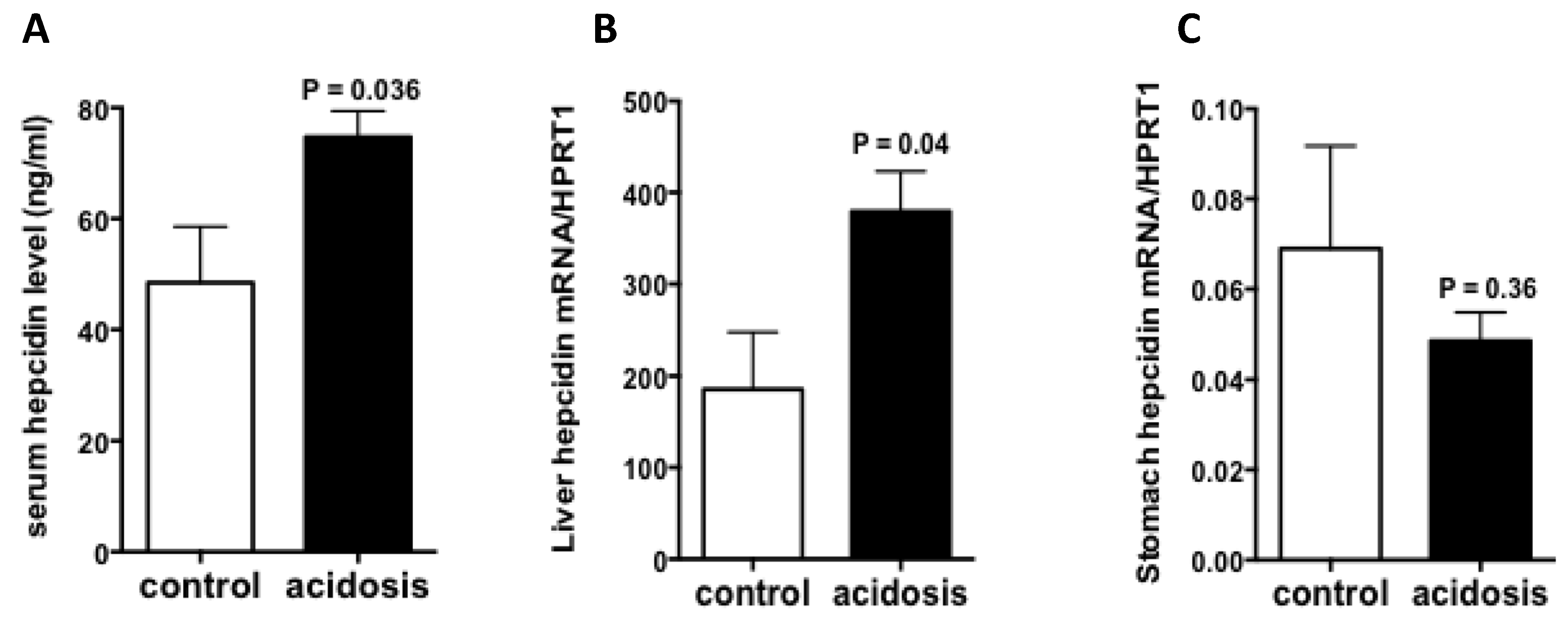
2.2. Hepcidin Expression Is Increased in Acidosis Condition
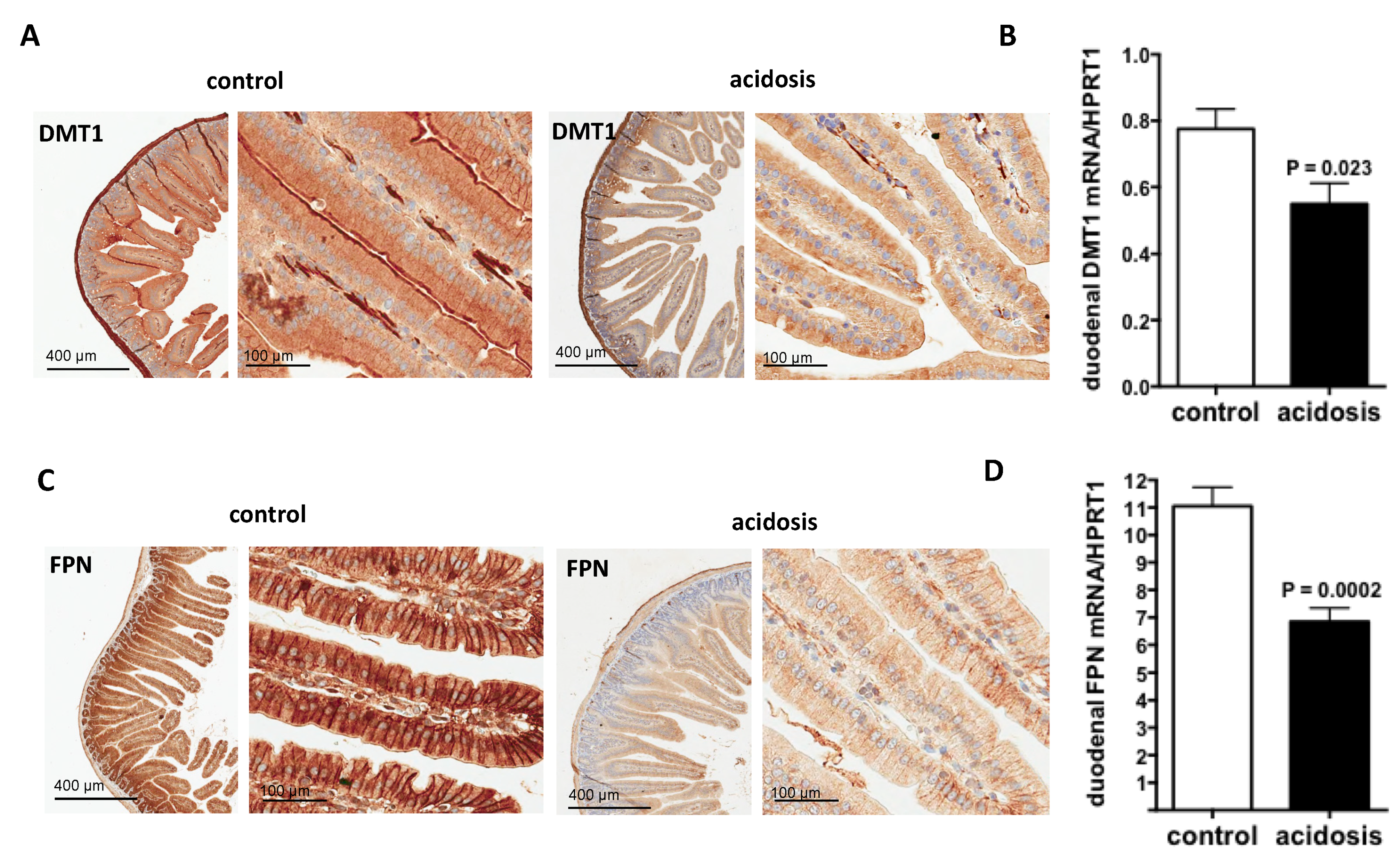
2.3. Both Duodenal Iron Transporters DMT1 and FPN Are Downregulated under Acidosis Condition
2.4. Regulation of Iron Status by Acidosis
2.5. ATP4A Expression Is Stimulated in Iron-Rich Diet
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experimental Animal Models
4.3. Biochemical Analyses
4.4. Immunohistochemistry
4.5. Quantitative RT-PCR
4.6. Protein Extraction and Immunoblotting (WB)
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McKie, A.T.; Barrow, D.; Latunde-Dada, G.O.; Rolfs, A.; Sager, G.; Mudaly, E.; Mudaly, M.; Richardson, C.; Barlow, D.; Bomford, A.; et al. An Iron-Regulated Ferric Reductase Associated with the Absorption of Dietary Iron. Science 2001, 291, 1755–1759. [Google Scholar] [CrossRef] [PubMed]
- McKie, A.T.; Marciani, P.; Rolfs, A.; Brennan, K.; Wehr, K.; Barrow, D.; Miret, S.; Bomford, A.; Peters, T.J.; Farzaneh, F.; et al. A Novel Duodenal Iron-Regulated Transporter, IREG1, Implicated in the Basolateral Transfer of Iron to the Circulation. Mol. Cell 2000, 5, 299–309. [Google Scholar] [CrossRef]
- Abboud, S.; Haile, D.J. A Novel Mammalian Iron-Regulated Protein Involved in Intracellular Iron Metabolism. J. Biol. Chem. 2000, 275, 19906–19912. [Google Scholar] [CrossRef]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and Characterization of a Mammalian Proton-Coupled Metal-Ion Transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Fleming, M.D.; Trenor, C.C.; Su, M.A.; Foernzler, D.; Beier, D.R.; Dietrich, W.F.; Andrews, N.C. Microcytic Anaemia Mice Have a Mutation in Nramp2, a Candidate Iron Transporter Gene. Nat. Genet. 1997, 16, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, P.; Bothwell, T.; Charlton, R.W. Role of hydrochloric acid in iron absorption. J. Appl. Physiol. 1964, 19, 187–188. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; Schubert, M.L. Gastric Secretion. Curr. Opin. Gastroenterol. 2012, 28, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.L. Gastric Exocrine and Endocrine Secretion. Curr. Opin. Gastroenterol. 2009, 25, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Spicer, Z.; Miller, M.L.; Andringa, A.; Riddle, T.M.; Duffy, J.J.; Doetschman, T.; Shull, G.E. Stomachs of Mice Lacking the Gastric H,K-ATPase Alpha -Subunit Have Achlorhydria, Abnormal Parietal Cells, and Ciliated Metaplasia. J. Biol. Chem. 2000, 275, 21555–21565. [Google Scholar] [CrossRef]
- Asano, S.; Kawada, K.; Kimura, T.; Grishin, A.V.; Caplan, M.J.; Takeguchi, N. The Roles of Carbohydrate Chains of the Beta-Subunit on the Functional Expression of Gastric H(+),K(+)-ATPase. J. Biol. Chem. 2000, 275, 8324–8330. [Google Scholar] [CrossRef] [PubMed]
- Mandal, U.; Ali, K.M.; Chatterjee, K.; De, D.; Biswas, A.; Ghosh, D. Management of Experimental Hypochlorhydria with Iron Deficiency by the Composite Extract of Fumaria Vaillantii L. and Benincasa Hispida T. in Rat. J. Nat. Sci. Biol. Med. 2014, 5, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.M.; Rjater, R.G.; Kale-Pradhan, P.B. Perils and Pitfalls of Long-Term Effects of Proton Pump Inhibitors. Expert Rev. Clin. Pharmacol. 2013, 6, 443–451. [Google Scholar] [CrossRef] [PubMed]
- van Aerts, R.M.M.; van Deursen, C.T.B.M.; Koek, G.H. Proton Pump Inhibitors Reduce the Frequency of Phlebotomy in Patients With Hereditary Hemochromatosis. Clin. Gastroenterol. Hepatol. 2016, 14, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, C.; Geissler, C.A.; Powell, J.J.; Bomford, A. Proton Pump Inhibitors Suppress Absorption of Dietary Non-Haem Iron in Hereditary Haemochromatosis. Gut 2007, 56, 1291–1295. [Google Scholar] [CrossRef] [PubMed]
- Krieg, L.; Milstein, O.; Krebs, P.; Xia, Y.; Beutler, B.; Du, X. Mutation of the Gastric Hydrogen-Potassium ATPase Alpha Subunit Causes Iron-Deficiency Anemia in Mice. Blood 2011, 118, 6418–6425. [Google Scholar] [CrossRef] [PubMed]
- Pigeon, C.; Ilyin, G.; Courselaud, B.; Leroyer, P.; Turlin, B.; Brissot, P.; Loréal, O. A New Mouse Liver-Specific Gene, Encoding a Protein Homologous to Human Antimicrobial Peptide Hepcidin, Is Overexpressed during Iron Overload. J. Biol. Chem. 2001, 276, 7811–7819. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, G.; Bennoun, M.; Devaux, I.; Beaumont, C.; Grandchamp, B.; Kahn, A.; Vaulont, S. Lack of Hepcidin Gene Expression and Severe Tissue Iron Overload in Upstream Stimulatory Factor 2 (USF2) Knockout Mice. Proc. Natl. Acad. Sci. USA 2001, 98, 8780–8785. [Google Scholar] [CrossRef]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin Regulates Cellular Iron Efflux by Binding to Ferroportin and Inducing Its Internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed]
- Delaby, C.; Pilard, N.; Gonçalves, A.S.; Beaumont, C.; Canonne-Hergaux, F. Presence of the Iron Exporter Ferroportin at the Plasma Membrane of Macrophages Is Enhanced by Iron Loading and Down-Regulated by Hepcidin. Blood 2005, 106, 3979–3984. [Google Scholar] [CrossRef] [PubMed]
- Brasse-Lagnel, C.; Karim, Z.; Letteron, P.; Bekri, S.; Bado, A.; Beaumont, C. Intestinal DMT1 Cotransporter Is Down-Regulated by Hepcidin via Proteasome Internalization and Degradation. Gastroenterology 2011, 140, 1261–1271.e1. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, S.; Sharp, P.; Ramesh, B.; Srai, S.K. Inhibition of Iron Transport across Human Intestinal Epithelial Cells by Hepcidin. Blood 2004, 104, 2178–2180. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chaston, T.; Chung, B.; Mascarenhas, M.; Marks, J.; Patel, B.; Srai, S.K.; Sharp, P. Evidence for Differential Effects of Hepcidin in Macrophages and Intestinal Epithelial Cells. Gut 2008, 57, 374–382. [Google Scholar] [CrossRef]
- Mena, N.P.; Esparza, A.; Tapia, V.; Valdés, P.; Núñez, M.T. Hepcidin Inhibits Apical Iron Uptake in Intestinal Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G192–G198. [Google Scholar] [CrossRef]
- Schwarz, P.; Kübler, J.A.M.; Strnad, P.; Müller, K.; Barth, T.F.E.; Gerloff, A.; Feick, P.; Peyssonnaux, C.; Vaulont, S.; Adler, G.; et al. Hepcidin Is Localised in Gastric Parietal Cells, Regulates Acid Secretion and Is Induced by Helicobacter Pylori Infection. Gut 2012, 61, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Frazier, M.D.; Mamo, L.B.; Ghio, A.J.; Turi, J.L. Hepcidin Expression in Human Airway Epithelial Cells Is Regulated by Interferon-γ. Respir. Res. 2011, 12, 100. [Google Scholar] [CrossRef] [PubMed]
- Gnana-Prakasam, J.P.; Martin, P.M.; Mysona, B.A.; Roon, P.; Smith, S.B.; Ganapathy, V. Hepcidin Expression in Mouse Retina and Its Regulation via Lipopolysaccharide/Toll-like Receptor-4 Pathway Independent of Hfe. Biochem. J. 2008, 411, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Kulaksiz, H.; Theilig, F.; Bachmann, S.; Gehrke, S.G.; Rost, D.; Janetzko, A.; Cetin, Y.; Stremmel, W. The Iron-Regulatory Peptide Hormone Hepcidin: Expression and Cellular Localization in the Mammalian Kidney. J. Endocrinol. 2005, 184, 361–370. [Google Scholar] [CrossRef]
- Moulouel, B.; Houamel, D.; Delaby, C.; Tchernitchko, D.; Vaulont, S.; Letteron, P.; Thibaudeau, O.; Puy, H.; Gouya, L.; Beaumont, C.; et al. Hepcidin Regulates Intrarenal Iron Handling at the Distal Nephron. Kidney Int. 2013, 84, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Houamel, D.; Ducrot, N.; Lefebvre, T.; Daher, R.; Moulouel, B.; Sari, M.-A.; Letteron, P.; Lyoumi, S.; Millot, S.; Tourret, J.; et al. Hepcidin as a Major Component of Renal Antibacterial Defenses against Uropathogenic Escherichia coli. J. Am. Soc. Nephrol. JASN 2016, 27, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Ashby, D.R.; Gale, D.P.; Busbridge, M.; Murphy, K.G.; Duncan, N.D.; Cairns, T.D.; Taube, D.H.; Bloom, S.R.; Tam, F.W.K.; Chapman, R.S.; et al. Plasma Hepcidin Levels Are Elevated but Responsive to Erythropoietin Therapy in Renal Disease. Kidney Int. 2009, 75, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Tomosugi, N.; Kawabata, H.; Wakatabe, R.; Higuchi, M.; Yamaya, H.; Umehara, H.; Ishikawa, I. Detection of Serum Hepcidin in Renal Failure and Inflammation by Using ProteinChip System. Blood 2006, 108, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- van der Weerd, N.C.; Grooteman, M.P.C.; Bots, M.L.; van den Dorpel, M.A.; den Hoedt, C.H.; Mazairac, A.H.A.; Nubé, M.J.; Penne, E.L.; Gaillard, C.A.; Wetzels, J.F.M.; et al. Hepcidin-25 in Chronic Hemodialysis Patients Is Related to Residual Kidney Function and Not to Treatment with Erythropoiesis Stimulating Agents. PloS ONE 2012, 7, e39783. [Google Scholar] [CrossRef]
- Coyne, D.W. Hepcidin: Clinical Utility as a Diagnostic Tool and Therapeutic Target. Kidney Int. 2011, 80, 240–244. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mizumoto, C.; Kawabata, H.; Uchiyama, T.; Sakamoto, S.; Kanda, J.; Tomosugi, N.; Takaori-Kondo, A. Acidic Milieu Augments the Expression of Hepcidin, the Central Regulator of Iron Homeostasis. Int. J. Hematol. 2012, 96, 701–709. [Google Scholar] [CrossRef]
- Chan, R.; Wong, V.W.-S.; Chu, W.C.-W.; Wong, G.L.-H.; Li, L.S.; Leung, J.; Chim, A.M.-L.; Yeung, D.K.-W.; Sea, M.M.-M.; Woo, J.; et al. Higher Estimated Net Endogenous Acid Production May Be Associated with Increased Prevalence of Nonalcoholic Fatty Liver Disease in Chinese Adults in Hong Kong. PLoS ONE 2015, 10, e0122406. [Google Scholar] [CrossRef] [PubMed]
- Galy, B.; Ferring-Appel, D.; Kaden, S.; Gröne, H.-J.; Hentze, M.W. Iron Regulatory Proteins Are Essential for Intestinal Function and Control Key Iron Absorption Molecules in the Duodenum. Cell Metab. 2008, 7, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, N.; Pantopoulos, K. The IRP/IRE System in Vivo: Insights from Mouse Models. Front. Pharmacol. 2014, 5, 176. [Google Scholar] [CrossRef]
- Tybl, E.; Gunshin, H.; Gupta, S.; Barrientos, T.; Bonadonna, M.; Celma Nos, F.; Palais, G.; Karim, Z.; Sanchez, M.; Andrews, N.C.; et al. Control of Systemic Iron Homeostasis by the 3’ Iron-Responsive Element of Divalent Metal Transporter 1 in Mice. HemaSphere 2020, 4, e459. [Google Scholar] [CrossRef] [PubMed]
- Rajamäki, K.; Nordström, T.; Nurmi, K.; Åkerman, K.E.O.; Kovanen, P.T.; Öörni, K.; Eklund, K.K. Extracellular Acidosis Is a Novel Danger Signal Alerting Innate Immunity via the NLRP3 Inflammasome. J. Biol. Chem. 2013, 288, 13410–13419. [Google Scholar] [CrossRef] [PubMed]
- Agoro, R.; Mura, C. Inflammation-Induced up-Regulation of Hepcidin and down-Regulation of Ferroportin Transcription Are Dependent on Macrophage Polarization. Blood Cells. Mol. Dis. 2016, 61, 16–25. [Google Scholar] [CrossRef]
- Harada, N.; Kanayama, M.; Maruyama, A.; Yoshida, A.; Tazumi, K.; Hosoya, T.; Mimura, J.; Toki, T.; Maher, J.M.; Yamamoto, M.; et al. Nrf2 Regulates Ferroportin 1-Mediated Iron Efflux and Counteracts Lipopolysaccharide-Induced Ferroportin 1 MRNA Suppression in Macrophages. Arch. Biochem. Biophys. 2011, 508, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, N.; Dudas, J.; Ramadori, G. Changes of Gene Expression of Iron Regulatory Proteins during Turpentine Oil-Induced Acute-Phase Response in the Rat. Lab. Investig. J. Tech. Methods Pathol. 2007, 87, 713–725. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ludwiczek, S.; Aigner, E.; Theurl, I.; Weiss, G. Cytokine-Mediated Regulation of Iron Transport in Human Monocytic Cells. Blood 2003, 101, 4148–4154. [Google Scholar] [CrossRef] [PubMed]
- Attmane-Elakeb, A.; Mount, D.B.; Sibella, V.; Vernimmen, C.; Hebert, S.C.; Bichara, M. Stimulation by in Vivo and in VitroMetabolic Acidosis of Expression of RBSC-1, the Na+-K+(NH4+)-2Cl−Cotransporter of the Rat Medullary Thick Ascending Limb. J. Biol. Chem. 1998, 273, 33681–33691. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, T.; Dessendier, N.; Houamel, D.; Ialy-Radio, N.; Kannengiesser, C.; Manceau, H.; Beaumont, C.; Nicolas, G.; Gouya, L.; Puy, H.; et al. LC-MS/MS Method for Hepcidin-25 Measurement in Human and Mouse Serum: Clinical and Research Implications in Iron Disorders. Clin. Chem. Lab. Med. 2015, 53, 1557–1567. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| Mouse Hamp | CGATACCAATGCAGAAGAGAAGG | TTTGCAACAGATACCACACTGGG |
| Mouse Atp4a | AGCACCAGGCACCATGGGGAAG | CACCAGGGCCAGACCCCAGTT |
| Mouse Atp4b | ACCCCTACACCCCAGACTAC | CCATACACGTCCGGTCTCAA |
| Mouse Dmt1 | GGCTTTCTTATGAGCATTGCCTA | GGAGCACCCAGAGCAGCTTA |
| Mouse Fpn | CCCATAGTCTCTGTCAGCCTGC | CCGTCAAATCAAAGGACCAAA |
| Mouse Hprt1 | AGCTACTGTAATGATCAGTCAACG | AGAGGTCCTTTTCACCAGCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daher, R.; Ducrot, N.; Lefebvre, T.; Zineeddine, S.; Ausseil, J.; Puy, H.; Karim, Z. Crosstalk between Acidosis and Iron Metabolism: Data from In Vivo Studies. Metabolites 2022, 12, 89. https://doi.org/10.3390/metabo12020089
Daher R, Ducrot N, Lefebvre T, Zineeddine S, Ausseil J, Puy H, Karim Z. Crosstalk between Acidosis and Iron Metabolism: Data from In Vivo Studies. Metabolites. 2022; 12(2):89. https://doi.org/10.3390/metabo12020089
Chicago/Turabian StyleDaher, Raêd, Nicolas Ducrot, Thibaud Lefebvre, Sofia Zineeddine, Jérome Ausseil, Hervé Puy, and Zoubida Karim. 2022. "Crosstalk between Acidosis and Iron Metabolism: Data from In Vivo Studies" Metabolites 12, no. 2: 89. https://doi.org/10.3390/metabo12020089
APA StyleDaher, R., Ducrot, N., Lefebvre, T., Zineeddine, S., Ausseil, J., Puy, H., & Karim, Z. (2022). Crosstalk between Acidosis and Iron Metabolism: Data from In Vivo Studies. Metabolites, 12(2), 89. https://doi.org/10.3390/metabo12020089