
Enhanced β-carotene and Biomass Production by Induced Mixotrophy in Dunaliella salina across a Combined Strategy of Glycerol, Salinity, and Light


Abstract
:1. Introduction
2. Results
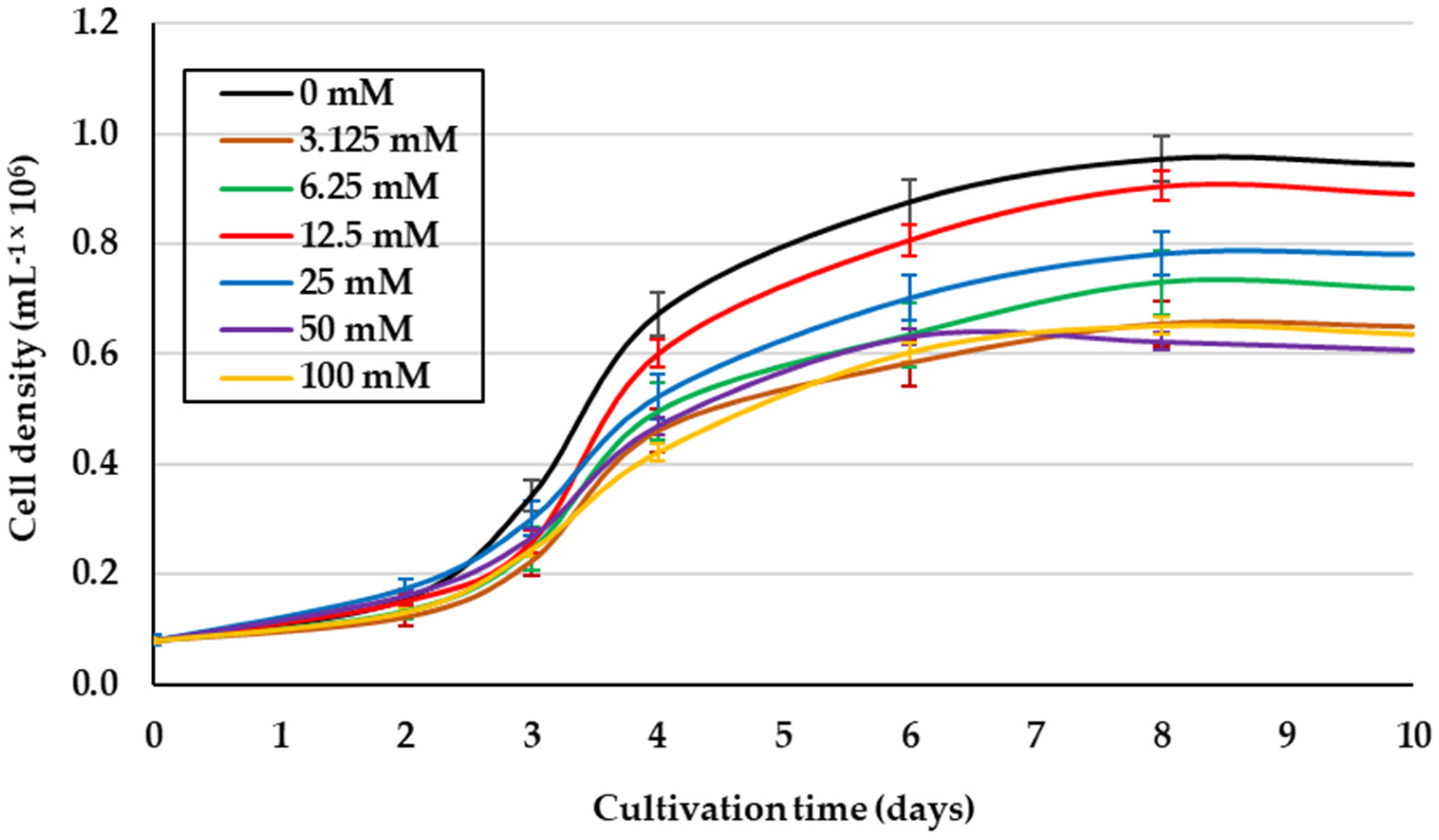
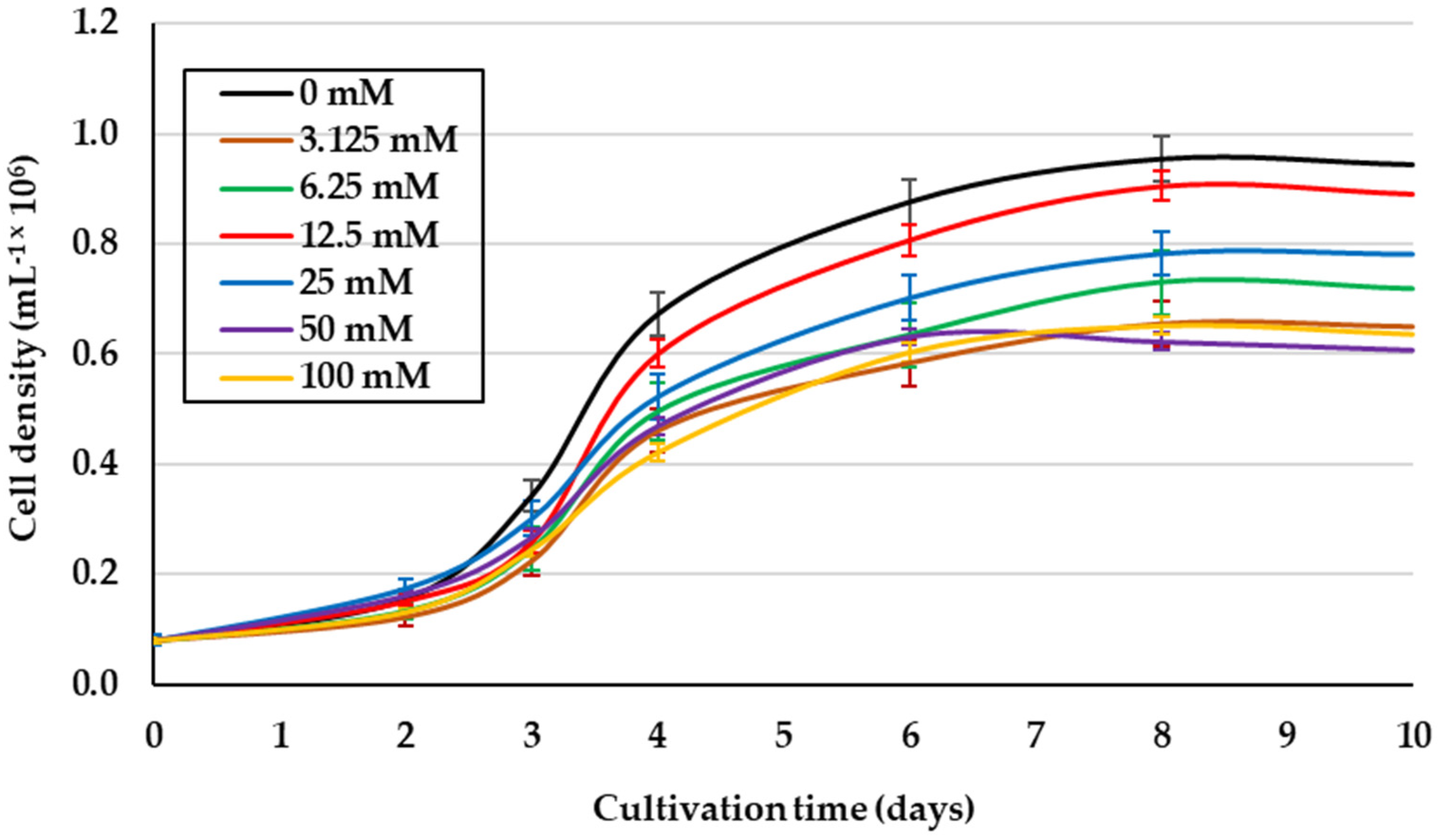
2.1. Effect of Glycerol Concentration on Mixotrophic Culture of D. salina under 1.5 M NaCl Isosmotic Salinity
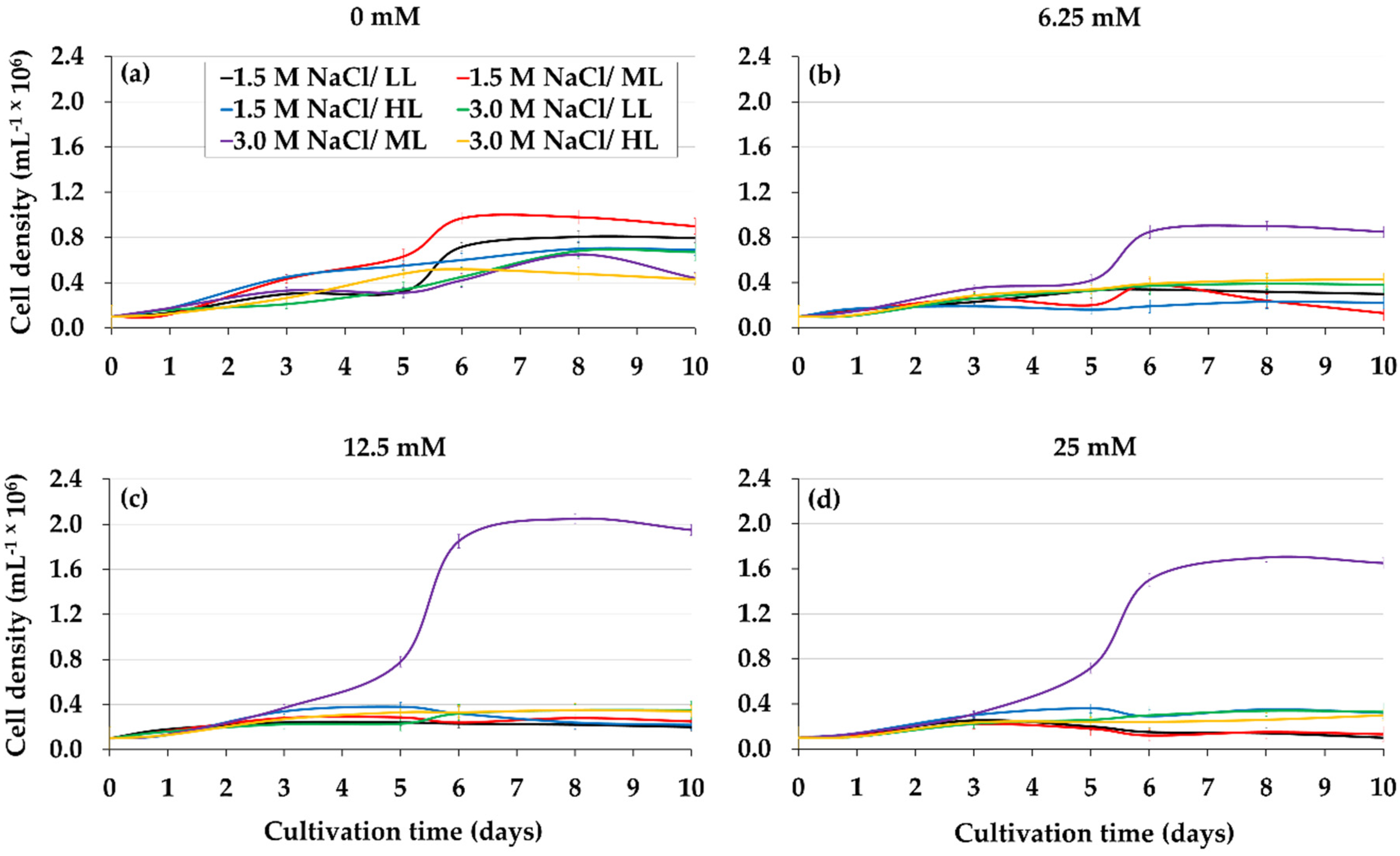
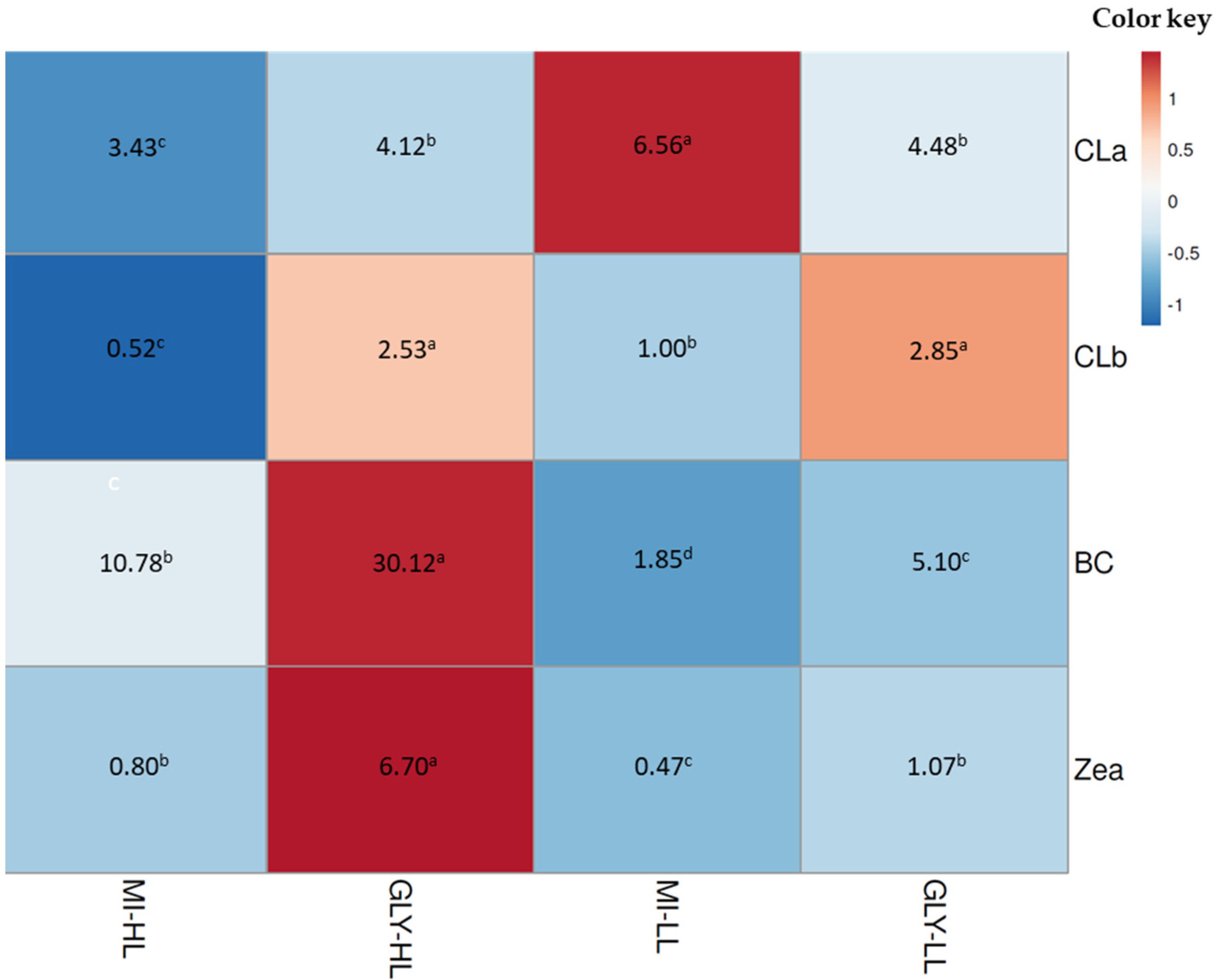
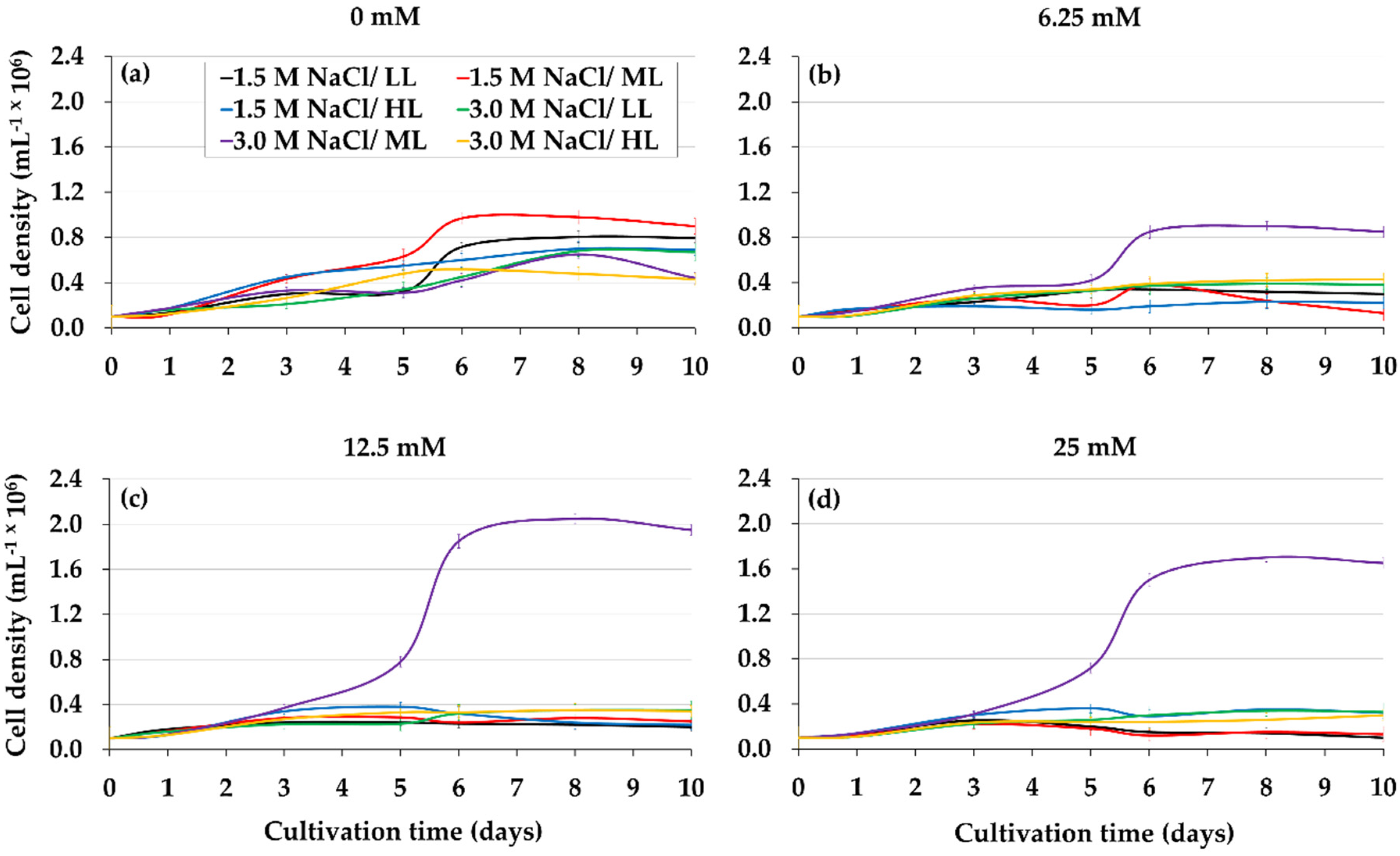
2.2. Combined Effect of Glycerol, Light, and Salinity on Mixotrophic Culture of D. salina
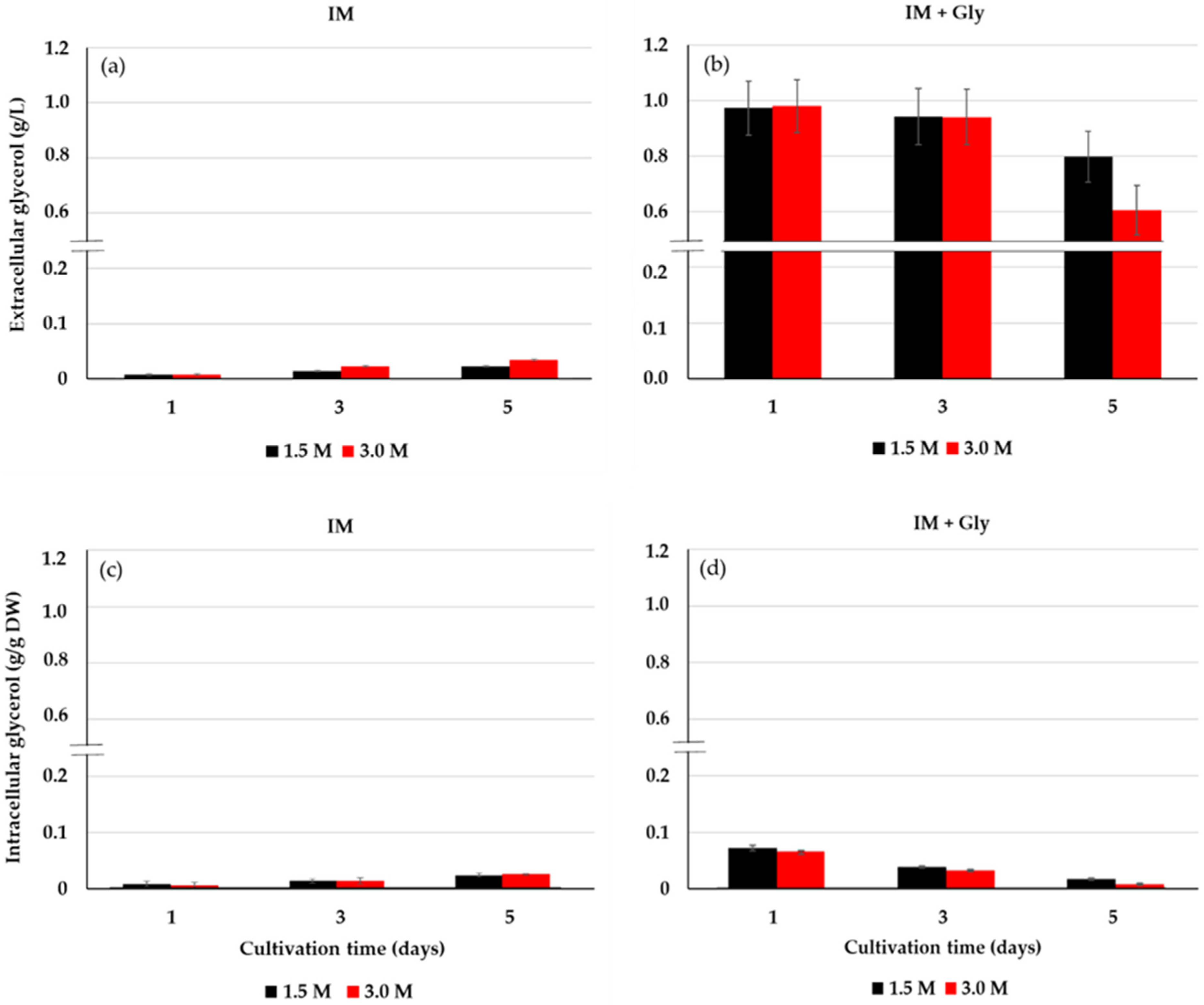
2.3. Extracellular and Intracellular Glycerol of D. salina
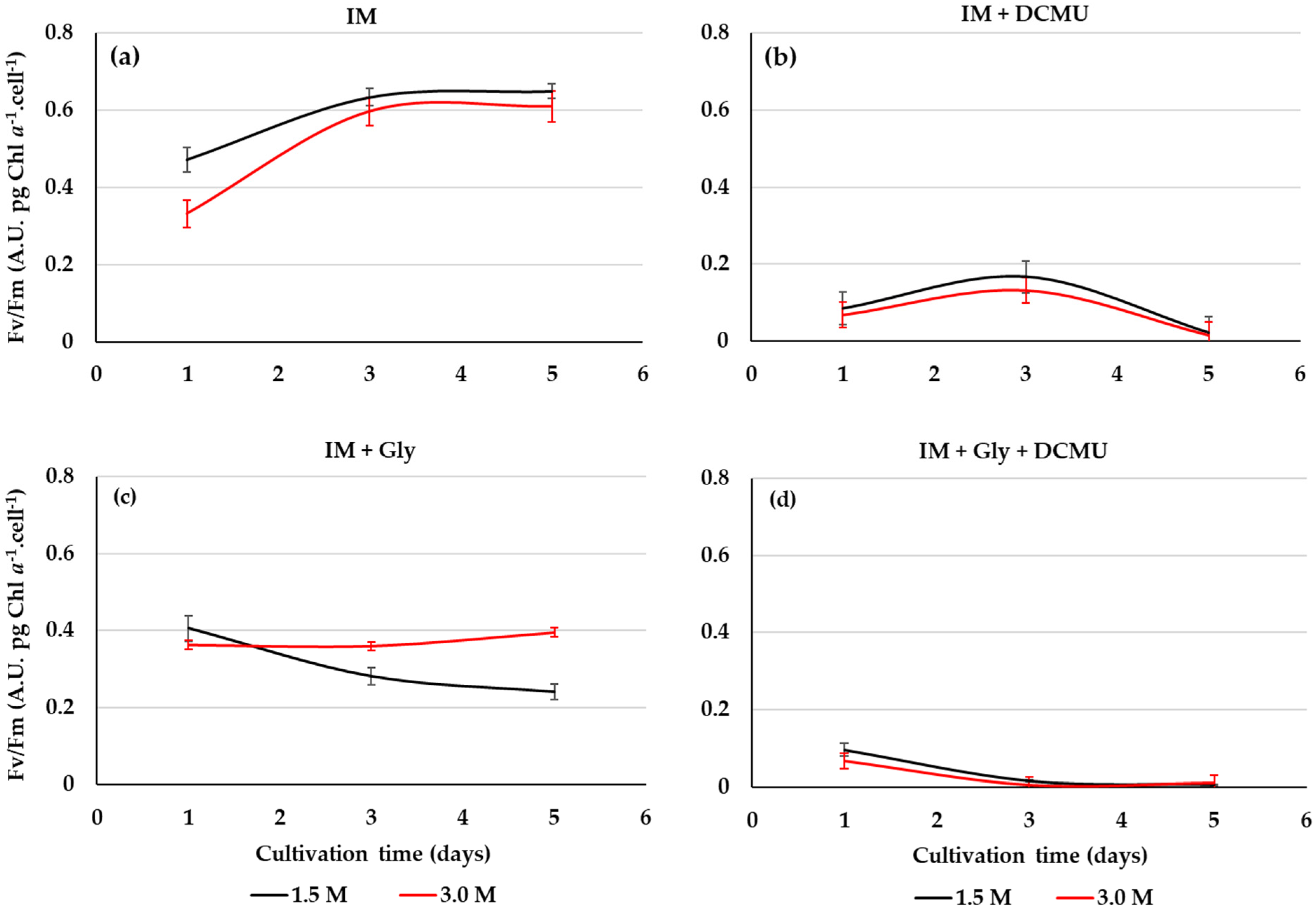
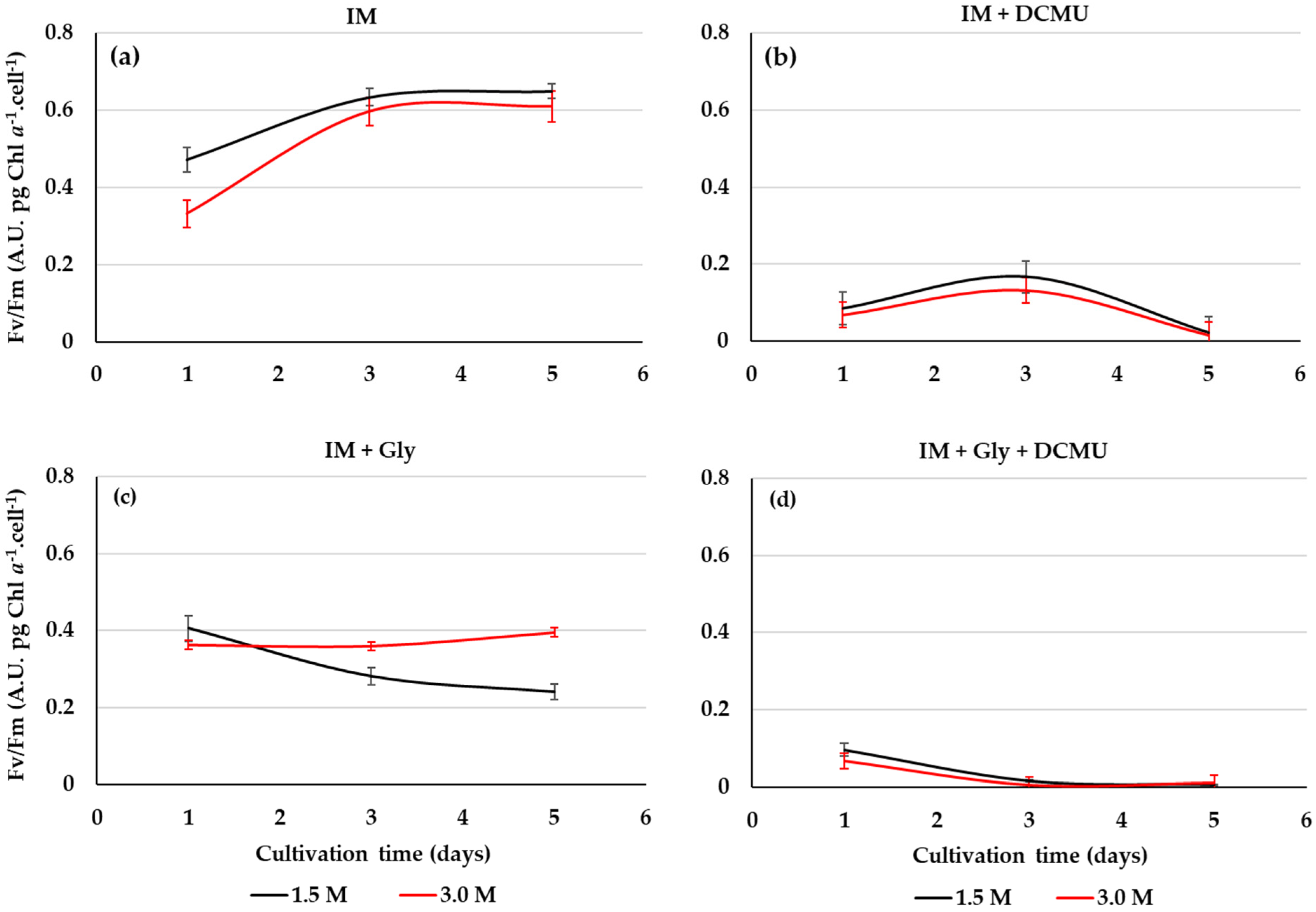
2.4. PSII Activity from D. salina Cultivated on Glycerol
3. Discussion
4. Materials and Methods
4.1. Strain and Culture Conditions
4.2. Mixotrophic Growth Conditions Set Up
4.3. Cell Growth and Dry Cell Weight
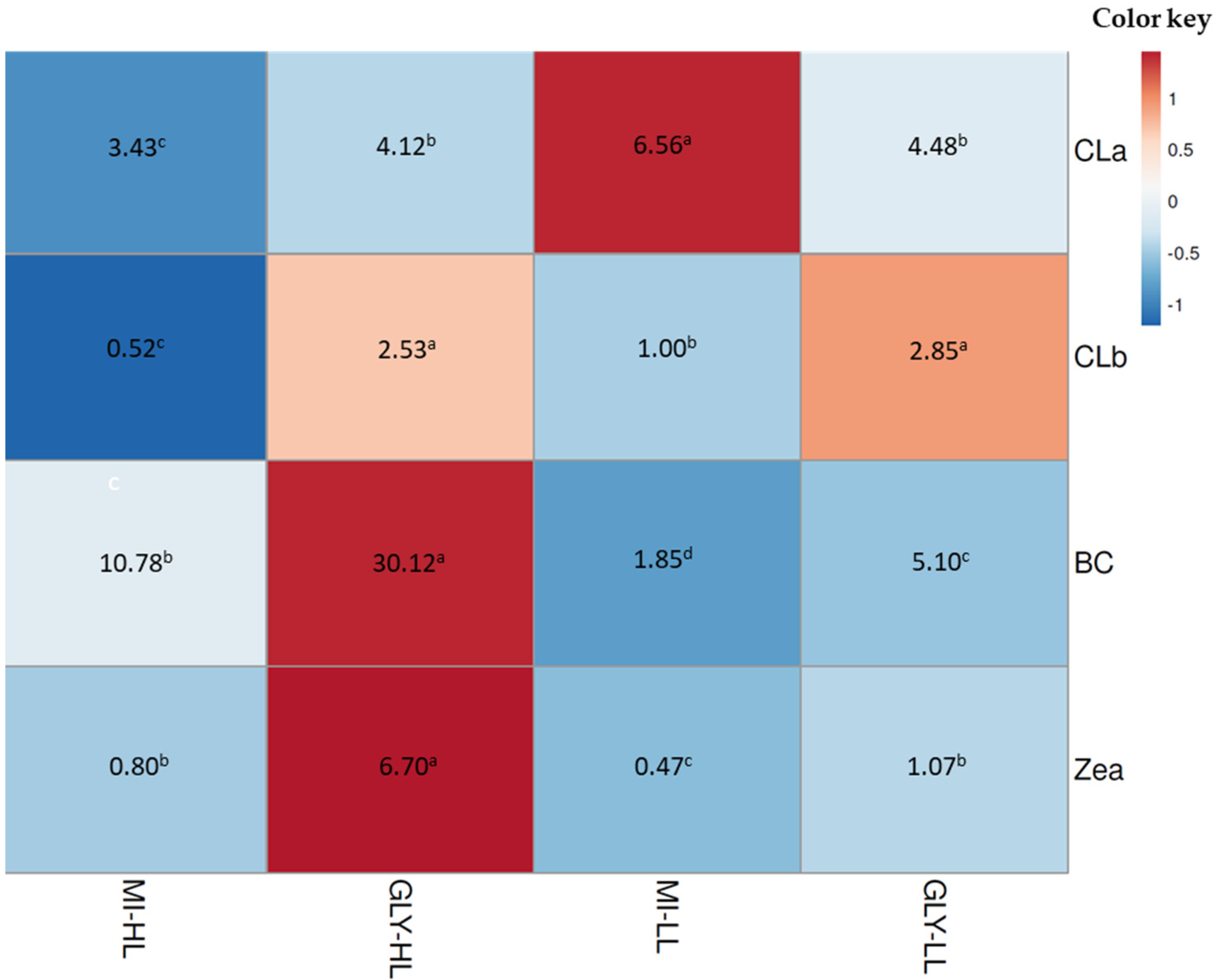
4.4. Pigment Content
4.4.1. Spectrophotometric Determination
4.4.2. HPLC Analysis
4.5. Extracellular and Intracellular Glycerol of D. salina
4.6. Photosynthetic Efficiency Measurement
4.7. Yield Parameters
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cezare-Gomes, E.A.; Mejia-da-Silva, L.; Pérez-Mora, L.S.; Matsudo, M.; Ferreira-Camargo, L.; Singh, A.; Monteiro de Carvalho, J.C. Potential of microalgae carotenoids for industrial application. Appl. Biochem. Biotech. 2019, 188, 602–634. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Z.; Jiang, H.; Mao, X. Biotechnology advances in β-carotene production by microorganisms. Trends Food Sci. Technol. 2021, 111, 322–332. [Google Scholar] [CrossRef]
- Mehariya, S.; Goswami, R.K.; Karthikeysan, O.P.; Verma, P. Microalgae for high-value products: A way towards green nutraceutical and pharmaceutical compounds. Chemosphere. 2021, 280, 130553. [Google Scholar] [CrossRef]
- Molino, A.; Mehariya, S.; Di Sanzo, G.; Larocca, V.; Martino, M.; Leone, G.P.; Marino, T.; Chianese, S.; Balducchi, R.; Musmarra, D. Recent developments in supercritical fluid extraction of bioactive compounds from microalgae: Role of key parameters, technological achievements and challenges. J. CO2 Util. 2020, 36, 196–209. [Google Scholar] [CrossRef]
- Ejike, E.C.C.; Collins, S.A.; Balasuriya, N.; Swanson, A.K.; Mason, B.; Udenigwe, C.C. Prospects of microalgae proteins in producing peptide-based functional foods for promoting cardiovascular health. Trends Food Sci. Technol. 2017, 59, 30–36. [Google Scholar] [CrossRef]
- Monte, J.; Ribeiro, C.; Parreira, C.; Costa, L.; Brive, L.; Casal, S.; Brazinha, C.; Crespo, J.G. Biorefinery of Dunaliella salina: Sustainable recovery of carotenoids, polar lipids and glycerol. Bioresour. Technol. 2020, 297, 122509. [Google Scholar] [CrossRef]
- Patel, A.K.; Singhania, R.R.; Sim, S.J.; Di Dong, C. Recent advancements in mixotrophic bioprocessing for production of high value microalgal products. Bioresour. Technol. 2021, 320, 124421. [Google Scholar] [CrossRef] [PubMed]
- Poddar, N.; Sen, R.; Martin, G. Glycerol and nitrate utilisation by marine microalgae Nannochloropsis salina and Chlorella sp. and associated bacteria during mixotrophic and heterotrophic growth. Algal Res. 2018, 33, 298–309. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, W.; Mao, X.; Li, Y.; Wu, T.; Chen, F. High-value biomass from microalgae production platforms: Strategies and progress based on carbon metabolism and energy conversion. Biotechnol. Biofuels 2018, 11, 227. [Google Scholar] [CrossRef] [Green Version]
- Chavoshi, Z.Z.; Shariati, M. Lipid production in Dunaliella salina under autotrophic, heterotrophic, and mixotrophic conditions. Biologia 2019, 74, 1579–1590. [Google Scholar] [CrossRef]
- Gonabadi, E.; Samadlouie, H.R.; Zenoozian, M.S. Optimization of culture conditions for enhanced Dunaliella salina productions in mixotrophic culture. Prep. Biochem. Biotechnol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Morowvat, M.; Ghasemi, Y. Culture medium optimization for enhanced β-carotene and biomass production by Dunaliella salina in mixotrophic culture. Biocatal. Agric. Biotechnol. 2016, 7, 217–223. [Google Scholar] [CrossRef]
- Mojaat, M.; Pruvost, J.; Foucault, A.; Legrand, J. Effect of organic carbon sources and Fe2+ ions on growth and β-carotene accumulation by Dunaliella salina. Biochem. Eng. J. 2008, 39, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Keerthi, S.; Koduru, U.; Sarma, N. A nutrient medium for development of cell dense inoculum in mixotrophic mode to seed mass culture units of Dunaliella salina. Algol. Stud. 2015, 147, 7–28. [Google Scholar] [CrossRef]
- Liu, X.; Duan, S.; Li, A.; Xu, N.; Cai, Z.; Hu, Z. Effects of organic carbon sources on growth, photosynthesis, and respiration of Phaeodactylum tricornutum. J. Appl. Phycol. 2009, 21, 239–246. [Google Scholar] [CrossRef]
- Kim, H.S.; Park, W.K.; Lee, B.; Seon, G.; Suh, W.I.; Moon, M.; Chang, Y.K. Optimization of heterotrophic cultivation of Chlorella sp. HS2 using screening, statistical assessment, and validation. Sci. Rep. 2019, 9, 19383. [Google Scholar] [CrossRef] [Green Version]
- Chalima, A.; Oliver, L.; Fernández de Castro, L.; Karnaouri, A.; Dietrich, T.; Topakas, A. Utilization of Volatile Fatty Acids from Microalgae for the Production of High Added Value Compounds. Fermentation 2017, 3, 54. [Google Scholar] [CrossRef] [Green Version]
- Baldisserotto, C.; Sabia, A.; Guerrini, A.; Demaria, S.; Maglie, M.; Ferroni, L.; Pancaldi, S. Mixotrophic cultivation of Thalassiosira pseudonana with pure and crude glycerol: Impact on lipid profile. Algal Res. 2021, 54, 102194. [Google Scholar] [CrossRef]
- Rajendran, L.; Nagarajan, N.G.; Karuppan, M. Enhanced biomass and lutein production by mixotrophic cultivation of Scenedesmus sp. using crude glycerol in an airlift photobioreactor. Biochem. Eng. J. 2020, 161, 107684. [Google Scholar] [CrossRef]
- Sohrabi, D.; Jazini, M.H.; Shariati, M. Mixotrophic cultivation of Dunaliella salina on crude glycerol obtained from calcinated fatty acid production process. Russ. J. Mar. Biol. 2019, 45, 470–480. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, C.; Liu, J.; Yang, N. A strategy for stimulating astaxanthin and lipid production in Haematococcus pluvialis by exogenous glycerol application under low light. Algal Res. 2020, 46, 101779. [Google Scholar] [CrossRef]
- Clomburg, J.M.; Gonzalez, R. Anaerobic fermentation of glycerol: A platform for renewable fuels and chemicals. Trends Biotechnol. 2013, 31, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Taghavi, N.; Robinson, G. Improving the optimum yield and growth of Chlamydomonas reinhardtii CC125 and CW15 using various carbon sources and growth regimes. Afr. J. Biotechnol. 2016, 15, 1083–1100. [Google Scholar]
- Bindea, M.; Rusu, B.; Rusu, A.; Trif, M.; Leopold, L.F.; Dulf, F.; Vodnar, D.C. Valorification of crude glycerol for pure fractions of docosahexaenoic acid and β-carotene production by using Schizochytrium limacinum and Blakeslea trispora. Microb. Cell Fact. 2018, 17, 97. [Google Scholar] [CrossRef]
- Borowitzka, M.; Borowitzka, L. Dunaliella. In Micro-Algal Biotechnology; Borowitzka, M., Borowitzka, L., Eds.; Cambridge University Press: New York, NY, USA, 1988; pp. 27–58. [Google Scholar]
- Chen, H.; Jiang, J. Osmotic Responses of Dunaliella to the Changes of Salinity. J. Cell. Physiol. 2009, 219, 251–258. [Google Scholar] [CrossRef]
- D’Imporzano, G.; Salati, S.; Veronesi, D.; Scaglia, B.; Adani, F. Microalgae Mixotrophic Growth: Opportunity for Stream Depuration and Carbon Recovery. In Prospects and Challenges in Algal Biotechnology; Tripathi, B.N., Kumar, D., Eds.; Springer Nature: Singapore, 2017; pp. 141–177. [Google Scholar]
- Suarez, G.; Romero, T.; Borowitzka, M. Cultivo de Dunaliella salina en medio orgánico. Boletín Cent. Investig. Biológicas Maracaibo 1999, 33, 211–225. [Google Scholar]
- Farhat, N.; Rabhi, M.; Falleh, H.; Jouini, J.; Abdelly, C.; Smaoui, A. Optimization of salt concentrations for a higher carotenoid production in Dunaliella salina (chlorophyceae). J. Phycol. 2011, 47, 1072–1077. [Google Scholar] [CrossRef]
- Guevara, M.; Lodeiros, C.; Gómez, O.; Lemus, N.; Núñez, P.; Romero, L.; Vásquez, A.; Rosales, N. Carotenogénesis de cinco cepas del alga Dunaliella sp. (Chlorophyceae) aisladas de lagunas hipersalinas de Venezuela. Rev. Biol. Trop. 2005, 53, 331–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grama, B.S.; Agathos, S.N.; Jeffryes, C.S. Balancing photosynthesis and respiration increase microalgal biomass productivity during photoheterotrophy on glycerol. ACS Sustain. Chem. Eng. 2016, 4, 1611–1618. [Google Scholar] [CrossRef]
- Cecchin, M.; Benfatto, B.; Griggio, F.; Mori, A.; Cazzaniga, S.; Vitulo, N.; Delledonne, M.; Ballottari, M. Molecular basis of autotrophic vs mixotrophic growth in Chlorella sorokiniana. Sci. Rep. 2018, 8, 6465. [Google Scholar] [CrossRef]
- Marchello, A.E.; Dos Santos, A.C.; Lombardi, A.T.; Oliveira de Souza, C.W.; Montanhim, G.C. Physiological and ecological aspects of Chlorella sorokiniana (Trebouxiophyceae) under photoautotrophic and mixotrophic conditions. Microb. Ecol. 2018, 76, 791–800. [Google Scholar] [CrossRef]
- Abiusi, F.; Wijffels, R.H.; Janssen, M. Doubling of microalgae productivity by oxygen balanced mixotrophy. ACS Sustain. Chem. Eng. 2020, 8, 6065–6074. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.K.P.; Hughes, A.D.; McEvoy, L.; Thornton, B.; Day, J.G. The carbon partitioning of glucose and DIC in mixotrophic, heterotrophic and photoautotrophic cultures of Tetraselmis suecica. Biotechnol. Lett. 2021, 43, 729–743. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, O.; Bashan, Y. Microalgal heterotrophic and mixotrophic culturing for bio-refining: From metabolic routes to techno-economics. In Algal Biorefineries: Products and Refinery Design; Prokop, A., Bajpai, R., Zappi, M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 61–131. [Google Scholar]
- Lin, H.; Fang, L.; Low, C.S.; Chow, Y.; Lee, Y.K. Occurrence of glycerol uptake in Dunaliella tertiolecta under hyperosmotic stress. FEBS J. 2013, 280, 1064–1072. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, T.; Mori, N.; Sato, N. Activation of oxidative carbon metabolism by nutritional enrichment by photosynthesis and exogenous organic compounds in the red alga Cyanidioschyzon merolae: Evidence for heterotrophic growth. SpringerPlus 2015, 4, 559. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, A.; Schreiber, U.; Avron, M. Salt-induced metabolic changes in Dunaliella salina. Plant Physiol. 1980, 65, 810–813. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Jiang, J.; Wu, G. Effects of salinity changes on the growth of Dunaliella salina and its isozyme activities of Glycerol-3-phosphate dehydrogenase. J. Agric. Food Chem. 2009, 57, 6178–6182. [Google Scholar] [CrossRef]
- Yazdani, S.S.; González, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhuge, J.; Fang, H.; Prior, B.A. Glycerol production by microbial fermentation: A review. Biotechnol. Adv. 2001, 19, 201–223. [Google Scholar] [CrossRef]
- Han, D.; Li, Y.; Hu, Q. Astaxanthin in microalgae: Pathways, functions and biotechnological implications. Algae 2013, 28, 131–147. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Show, P.L.; Juan, J.C.; Chang, J.; Ling, T.C. Enhancing biomass and lipid productions of microalgae in palm oil mil effluent using carbon and nutrient supplementation. Energy Convers. Manag. 2018, 164, 188–197. [Google Scholar] [CrossRef]
- Bardavid, R.E.; Khristo, P.; Oren, A. Interrelationships between Dunaliella and halophilic prokaryotes in saltern crystallizer ponds. Extremophiles 2008, 12, 5–14. [Google Scholar] [CrossRef]
- White, S.; Anandraj, A.; Bux, F. PAM fluorometry as a tool to assess microalgal nutrient stress and monitor cellular neutral lipids. Bioresour. Technol. 2011, 102, 1675–1682. [Google Scholar] [CrossRef]
- Xu, Y.; Ibrahim, I.M.; Harvey, P.J. The influence of photoperiod and light intensity on the growth and photosynthesis of Dunaliella salina (chlorophyta) CCAP 19/30. Plant Physiol. Biochem. 2016, 106, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, M.M.; Liu, S.F.; Xu, R.L.; Mou, J.H.; Qin, Z.H.; Zhou, Z.G.; Li, H.Y.; Lin, C.S.; Sun, Z. Synergistic bioconversion of lipids and carotenoids from food waste by Dunaliella salina with fulvic acid via a two-stage cultivation strategy. Energy Convers. Manag. 2021, 234, 113908. [Google Scholar] [CrossRef]
- Xu, S.; Elsayed, M.; Ismail, G.A.; Li, C.; Wang, S.; Abomohra, A.E. Evaluation of bioethanol and biodiesel production from Scenedesmus obliquus grown in biodiesel waste glycerol: A sequential integrated route for enhanced energy recovery. Energy Convers. Manag. 2019, 197, 111907. [Google Scholar] [CrossRef]
- Tan, K.W.; Lee, Y.K. The dilemma for lipid productivity in green microalgae: Importance of substrate provision in improving oil yield without sacrificing growth. Biotechnol Biofuels 2016, 9, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashir, K.M.I.; Mansoor, S.; Kim, N.R.; Grohmann, F.R.; Shah, A.A.; Cho, M.G. Effect of organic carbon sources and environmental factors on cell growth and lipid content of Pavlova lutheri. Ann. Microbiol. 2019, 69, 353–368. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as Sources of Carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef]
- Cuaresma, M.; Casal, C.; Forján, E.; Vílchez, C. Productivity and selective accumulation of carotenoids of the novel extremophile microalga Chlamydomonas acidophila grown with different carbon sources in batch systems. J. Ind. Microbiol. Biotechnol. 2011, 38, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Mantzouridou, F.; Naziri, E.; Tsimidou, M.Z. Industrial glycerol as a supplementary carbon source in the production of beta-carotene by Blakeslea trispora. J. Agric. Food Chem. 2008, 56, 2668–2675. [Google Scholar] [CrossRef]
- Lao, Y.M.; Xiao, L.; Luo, L.X.; Jiang, J.G. Hypoosmotic expression of Dunaliella bardawil ζ-carotene desaturase is attributed to a hypoosmolarity-responsive element different from other key carotenogenic genes. Plant Physiol. 2014, 165, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Mousavi, P.; Morowvat, M.H.; Montazeri-Najafabady, N.; Abolhassanzadeh, Z.; Mohagheghzadeh, A.; Hamidi, M.; Niazi, A.; Ghasemi, Y. Investigating the effects of phytohormones on growth and β-carotene production in a naturally isolates stain of Dunaliella salina. J. Appl. Pharm. Sci. 2016, 6, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Kasahara, H. Current aspects of auxin biosynthesis in plants. Biosci. Biotechnol. Biochem. 2016, 80, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes, A.S.; Gonzalez, M.; Conejeros, M.; Dellarossa, V.; Parra, O. Growth and carotenogenesis in eight strains of Dunaliella salina Teodoresco from Chile. J. Appl. Phycol. 1992, 4, 111–118. [Google Scholar] [CrossRef]
- Pourkarimi, S.; Hallajisani, A.; Alizadehdakhel, A.; Nouralishahi, A.; Golzary, A. Factors affecting production of beta-carotene from Dunaliella salina microalgae. Biocatal. Agric. Biotechnol. 2020, 29, 101771. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Avron, M. On the factors which determine massive β-carotene accumulation in the halotolerant alga Dunaliella bardawil. Plant Physiol. 1983, 72, 593–597. [Google Scholar] [CrossRef] [Green Version]
- Imamoglu, E.; Demirel, Z.; Dalay, M.C. Evaluation of culture conditions of locally isolated Dunaliella salina strain EgeMacc-024. Biochem. Eng. J. 2014, 92, 22–27. [Google Scholar] [CrossRef]
- Sforza, E.; Pastore, M.; Barbera, E.; Bertucco, A. Respirometry as a tool to quantify kinetic parameters of microalgal mixotrophic growth. Bioprocess Biosyst. Eng. 2019, 42, 839–851. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Division rates. In Handbook of Phycological Methods. Culture Methods and Growth Measurements; Stein, J.R., Ed.; Cambridge University Press: Cambridge, UK, 1973; pp. 289–313. [Google Scholar]
- Zhu, C.J.; Lee, Y.K. Determination of biomass dry weight of marine microalgae. J. Appl. Phycol. 1997, 9, 189–194. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determination of total carotenoids and chlorophylls a and b of leaf in different solvents. Biochem. Soc. Trans. 1985, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Van Heukelem, L.; Thomas, C.S. Computer-assisted high-performance liquid chromatography meted development with applications to the isolation and analysis of phytoplankton pigments. J. Chromatogr. 2001, 910, 31–49. [Google Scholar] [CrossRef]
- Almazán-Becerril, A.; García-Mendoza, E. Maximum efficiency of charge separation of photosystem II of the phytoplankton community in the Eastern Tropical North Pacific of Mexico: A nutrient stress diagnostic tool? Cienc. Mar. 2008, 34, 29–43. [Google Scholar] [CrossRef]
- Wright, S.; Montoura, R.F.C. Guidelines for collection and pigment analysis of marine samples. In Phytoplankton Pigments in Oceanography: Guidelines to Modern Methods; Jeffrey, S.W., Mantoura, R.F.C., Wright, S., Eds.; UNESCO Publishing: Paris, France, 1997; pp. 429–445. [Google Scholar]
- Petrovic, U.; Plemenitas, A. Determination of intra-and extracellular concentration of glycerol in the halophilic black yeast Hortaea werneckii grown at different environmental salinities. In Non-Conventional Yeasts in Genetics, Biochemistry and Biotechnology; Wolf, K., Breunig, K., Barth, G., Eds.; Springer: New York, NY, USA, 2003; pp. 127–130. [Google Scholar]
- Guenther, J.E.; Melis, A. Dynamics of Photosystem II heterogeneity in Dunaliella salina (green algae). Photosynth. Res. 1990, 23, 195–203. [Google Scholar] [CrossRef]
- Gupta, S.; Pawar, S.B. Strategic mixed substrate cultivation of microalgae: Productivity, respiration, yield, and lipid quality. J. Appl. Phycol. 2019, 31, 1573–1588. [Google Scholar] [CrossRef]
- Montgomery, D. Diseño y Análisis de Experimentos; LIMUSA Wiley: Mexico, D.F., Mexico, 2004; p. 692. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Medium | Chl a (pg cell−1) | Chl b (pg cell−1) | BCar (pg cell−1) | Final pH |
|---|---|---|---|---|
| IM | 2.48 ± 0.18 a | 0.65 ± 0.05 a | 1.24 ± 0.09 b | 8.32 ± 0.11 a |
| IM + 12.5 mM Gly | 1.22 ± 0.09 b | 0.62 ± 0.05 a | 1.90 ± 0.14 a | 7.24 ± 0.05 b |
| p-value | p < 0.00012 | p > 0.495 | p < 0.002 | p < 0.00008 |
| Glycerol Level (mM) | Salinity (M NaCl) | Light Intensity (μmol Photons m−2 s−1) | Chl a (pg cell−1) | Chl b (pg cell−1) | Bcar (pg cell−1) | Final pH |
|---|---|---|---|---|---|---|
| 0 | 1.5 | 10 | 3.95 ± 0.19 b | 1.21 ± 0.06 c | 1.56 ± 0.08 e | 8.55 ± 0.42 ab |
| 6.25 | 1.5 | 10 | 2.47 ± 0.12 c | 1.33 ± 0.07 c | 2.28 ± 0.11 d | 7.65 ± 0.33 b |
| 12.5 | 1.5 | 10 | 2.24 ± 0.11 d | 1.71 ± 0.08 b | 2.61 ± 0.13 c | 7.50 ± 0.37 b |
| 25.0 | 1.5 | 10 | 2.75 ± 0.13 c | 2.36 ± 0.11 a | 2.19 ± 0.11 d | 7.42 ± 0.38 b |
| 0 | 1.5 | 50 | 2.18 ± 0.11 d | 0.59 ± 0.02 d | 1.37 ± 0.07 e | 9.05 ± 0.45 a |
| 6.25 | 1.5 | 50 | 2.90 ± 0.14 c | 1.30 ± 0.06 c | 1.67 ± 0.08 b | 7.68 ± 0.38 b |
| 12.5 | 1.5 | 50 | 3.53 ± 017 b | 1.33 ± 0.06 c | 2.20 ± 0.11 d | 7.58 ± 0.37 b |
| 25.0 | 1.5 | 50 | 3.27 ± 0.16 b | 1.57 ± 0.07 b | 1.63 ± 0.08 e | 7.50 ± 0.35 b |
| 0 | 1.5 | 100 | 4.03 ± 0.20 ab | 1.05 ± 0.05 cd | 2.76 ± 0.14 c | 9.25 ± 0.46 a |
| 6.25 | 1.5 | 100 | 1.56 ± 0.08 e | 0.79 ± 0.04 d | 2.28 ± 0.11 d | 7.71 ± 0.38 b |
| 12.5 | 1.5 | 100 | 3.63 ± 0.18 b | 1.54 ± 0.07 b | 2.82 ± 0.14 c | 7.82 ± 0.39 b |
| 25.0 | 1.5 | 100 | 2.51 ± 0.12 c | 1.00 ± 0.05 cd | 2.66 ± 0.13 c | 7.63 ± 0.38 b |
| 0 | 3.0 | 10 | 2.47 ± 0.13 c | 1.64 ± 0.08 b | 2.16 ± 0.11 d | 8.38 ± 0.42 ab |
| 6.25 | 3.0 | 10 | 2.28 ± 0.11 d | 2.02 ± 0.10 a | 2.77 ± 0.14 c | 7.84 ± 0.39 b |
| 12.5 | 3.0 | 10 | 2.90 ± 0.15 c | 1.58 ± 0.08 b | 2.11± 0.10 d | 7.67 ± 0.38 b |
| 25.0 | 3.0 | 10 | 2.87 ± 0.14 c | 1.51 ± 0.08 b | 2.12 ± 0.11 d | 7.60 ± 0.38 b |
| 0 | 3.0 | 50 | 1.82 ± 0.09 e | 0.45 ± 0.02 d | 1.28 ± 0.06 f | 8.42 ± 0.42 ab |
| 6.25 | 3.0 | 50 | 2.48 ± 0.12 d | 0.75 ± 0.04 d | 2.22 ± 0.11 e | 7.28 ± 0.36 b |
| 12.5 | 3.0 | 50 | 4.58 ± 0.22 a | 1.40 ± 0.07 c | 4.43 ± 0.17 c | 8.10 ± 0.40 ab |
| 25.0 | 3.0 | 50 | 3.91 ± 0.19 b | 1.22 ± 0.06 c | 2.44 ± 0.12 d | 8.12 ± 0.42 ab |
| 0 | 3.0 | 100 | 2.77 ± 0.14 c | 0.63 ± 0.03 d | 4.54 ± 0.21 b | 8.82 ± 0.44 ab |
| 6.25 | 3.0 | 100 | 2.62 ± 0.13 c | 0.49 ± 0.02 c | 4.73 ± 0.23 ab | 8.84 ± 0.40 ab |
| 12.5 | 3.0 | 100 | 2.54 ± 0.12 c (1) | 0.44 ± 0.02 c | 4.51 ± 0.22 b | 8.78 ± 0.43 ab |
| 25.0 | 3.0 | 100 | 2.60 ± 0.13 c (2) | 0.57 ± 0.03 b | 5.03 ± 0.25 a | 8.72 ± 0.44 ab |
| p-value | p < 0.00002 | p < 0.0001 | p < 0.0002 | p < 0.00002 | ||
| Culture Medium | Cell Density (mL−1 × 106) | Chl a (pg cell−1) | Chl b (pg cell−1) | BCar (pg cell−1) | Final pH |
|---|---|---|---|---|---|
| IM | 0.82 ± 0.07 a | 1.02 ± 0.09 a | 0.15 ± 0.01 b | 2.39 ± 0.22 a | 7.85 ± 0.32 a |
| IM + 12.5 mM Gly | 0.98 ± 0.09 a | 1.05 ± 0.09 a | 0.57 ± 0.05 a | 2.24 ± 0.20 a | 6.52 ± 0.09 b |
| p-value | p > 0.070 | p > 0.712 | p < 0.000 | p > 0.426 | p < 0.001 |
| Organic Carbon Substrate | Yx/s (g g−1) | Yp/s (mg g−1) | Reference |
|---|---|---|---|
| Glucose | -- | 8.91 ± 0.06 | [11] |
| Glycerol | 2.50 ± 0.08 | 1.16 ± 0.02 | [20] |
| 2.33 ± 0.03 | 43.5 ± 0.08 | In this work |
| Substrate | Biomass | β-Carotene | Reference |
|---|---|---|---|
| Inorganic | 1.0–5.0 × 106 cells mL−1 | 4–8 pg cell−1 | [58,59] |
| Glucose | 0.99 g L−1 | 8.03 mg g−1 | [12] |
| Acetate | 1.0–1.2 × 106 cells mL−1 | <5 pg cell−1 | [13] |
| Glycerol | 1.0 × 106 cells mL−1 | 0.94 pg cell−1 | [28] |
| 4.0–5.0 × 106 cells mL−1 | 0.8–1.0 pg cell−1 | [20] | |
| 2.15 × 106 cells mL−1 | 30 pg cell−1 | In this work | |
| (2.0 g L−1) | (50 mg g−1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capa-Robles, W.; García-Mendoza, E.; Paniagua-Michel, J.d.J. Enhanced β-carotene and Biomass Production by Induced Mixotrophy in Dunaliella salina across a Combined Strategy of Glycerol, Salinity, and Light. Metabolites 2021, 11, 866. https://doi.org/10.3390/metabo11120866
Capa-Robles W, García-Mendoza E, Paniagua-Michel JdJ. Enhanced β-carotene and Biomass Production by Induced Mixotrophy in Dunaliella salina across a Combined Strategy of Glycerol, Salinity, and Light. Metabolites. 2021; 11(12):866. https://doi.org/10.3390/metabo11120866
Chicago/Turabian StyleCapa-Robles, Willian, Ernesto García-Mendoza, and José de Jesús Paniagua-Michel. 2021. "Enhanced β-carotene and Biomass Production by Induced Mixotrophy in Dunaliella salina across a Combined Strategy of Glycerol, Salinity, and Light" Metabolites 11, no. 12: 866. https://doi.org/10.3390/metabo11120866
APA StyleCapa-Robles, W., García-Mendoza, E., & Paniagua-Michel, J. d. J. (2021). Enhanced β-carotene and Biomass Production by Induced Mixotrophy in Dunaliella salina across a Combined Strategy of Glycerol, Salinity, and Light. Metabolites, 11(12), 866. https://doi.org/10.3390/metabo11120866