
Effects of Monoterpene-Based Biostimulants on Chickpea (Cicer arietinum L.) Plants: Functional and Molecular Insights


 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
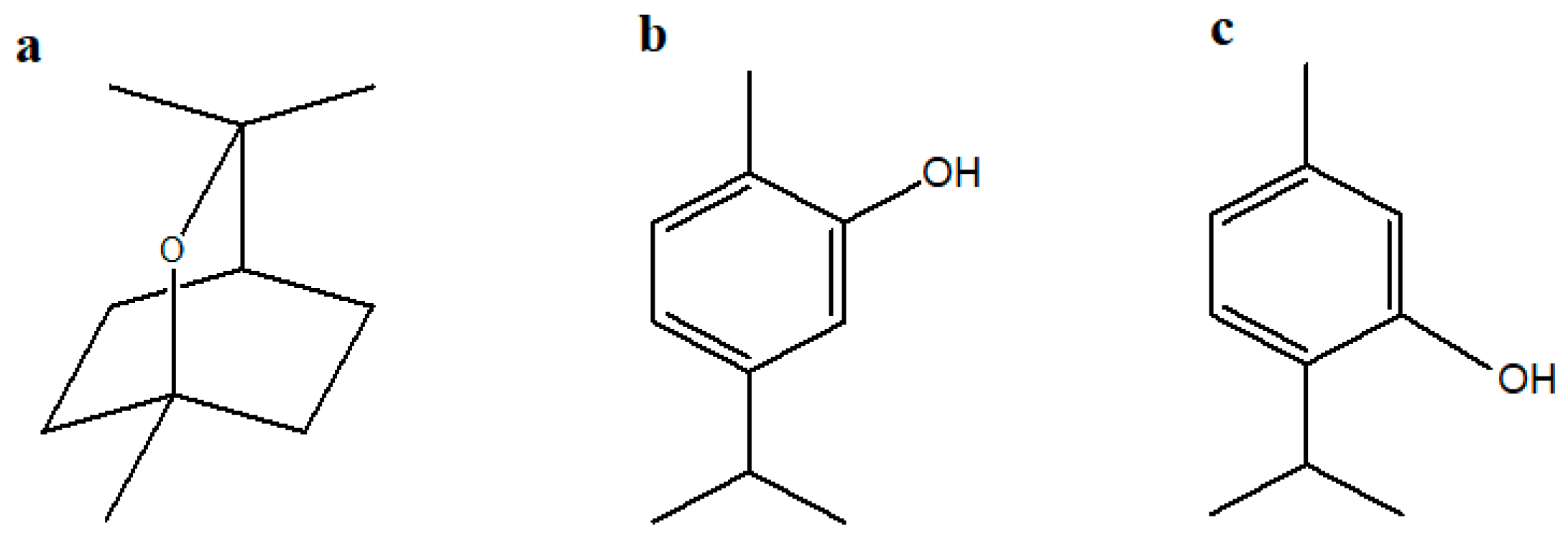
2.1. Major Compounds
2.2. Experimental Design
2.3. Biometric Parameters and Chlorophyll Content
2.4. Mineral Elements
2.5. Antioxidant Compounds
2.5.1. Total Polyphenol Content
2.5.2. Total Flavonoid Content
2.6. Total Soluble Proteins
2.7. Vitamin C (Ascorbic Acid)
2.8. PyRx-Based Molecular Docking: Preparation, Validation, and Visualization
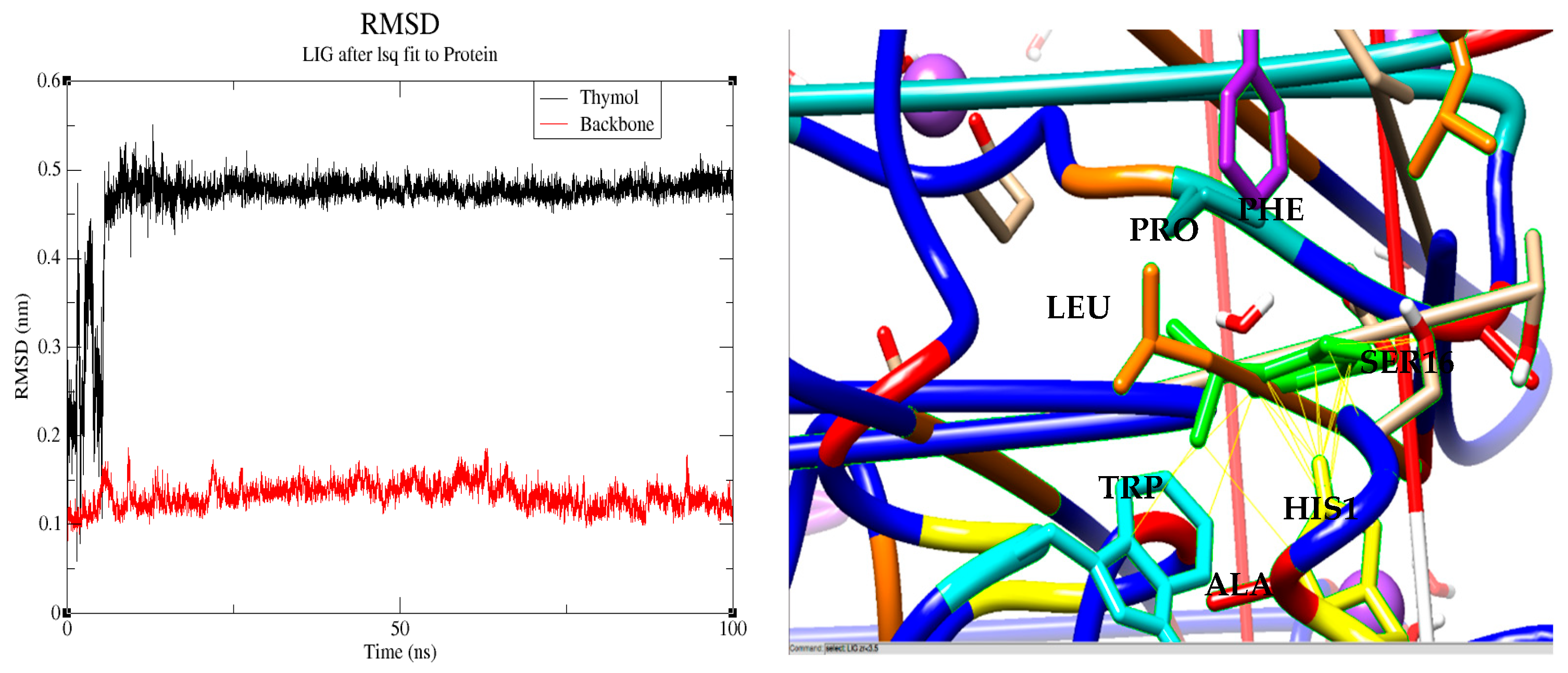
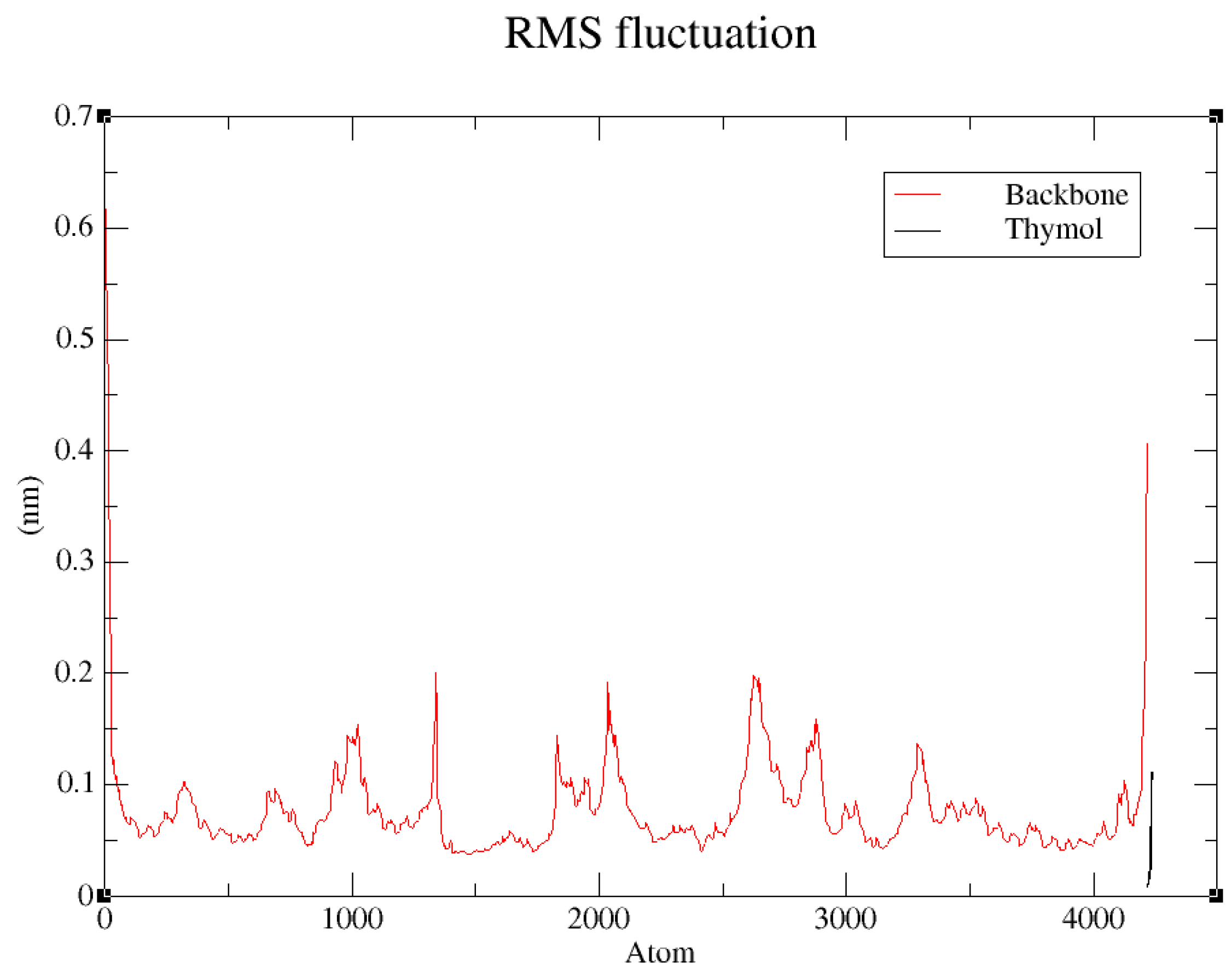
2.9. Implementation of Molecular Dynamics Simulations Using GROMACS
2.10. Statistical Analysis
3. Results
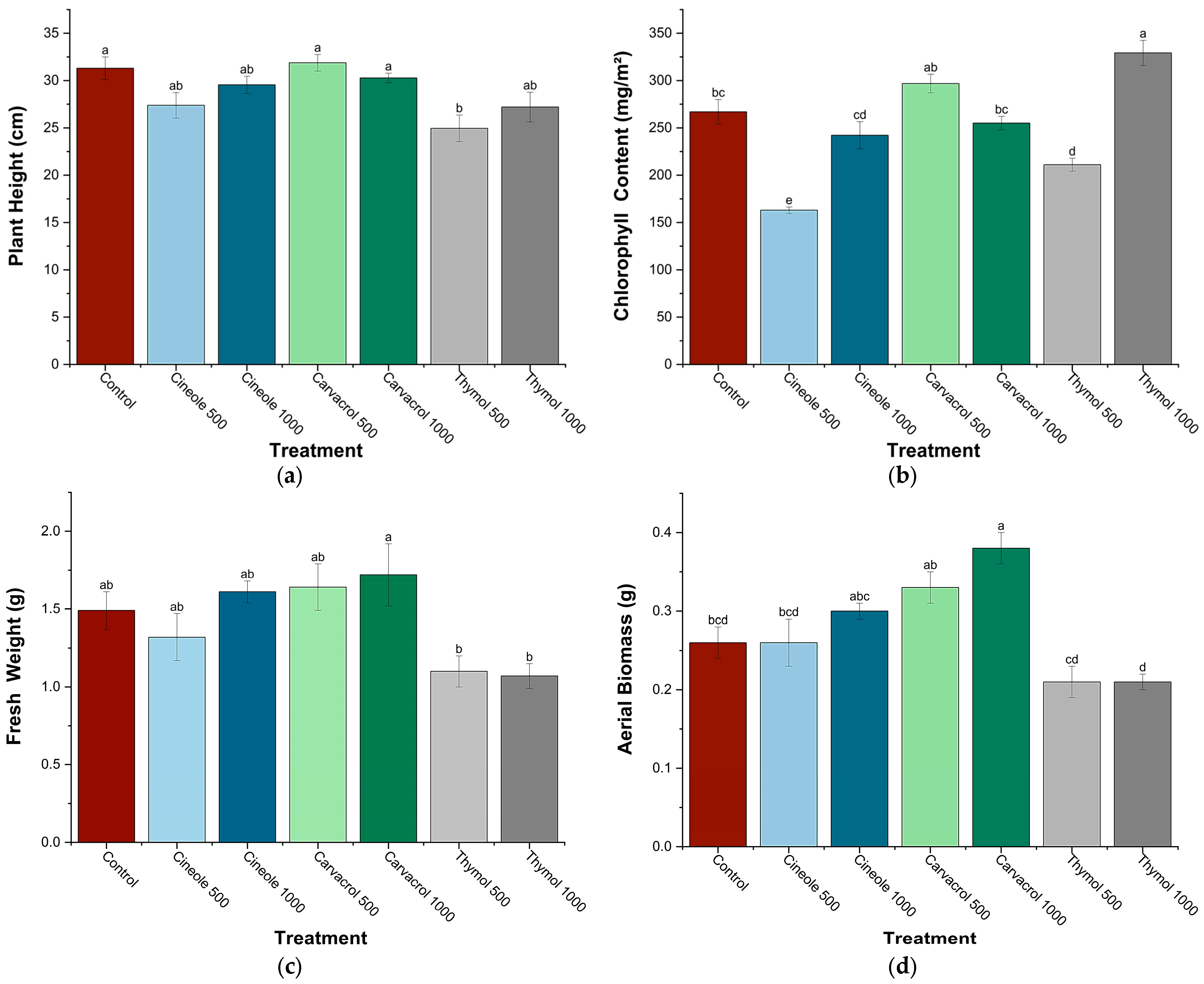
3.1. Plant Growth Parameters
3.2. Mineral Element Concentrations
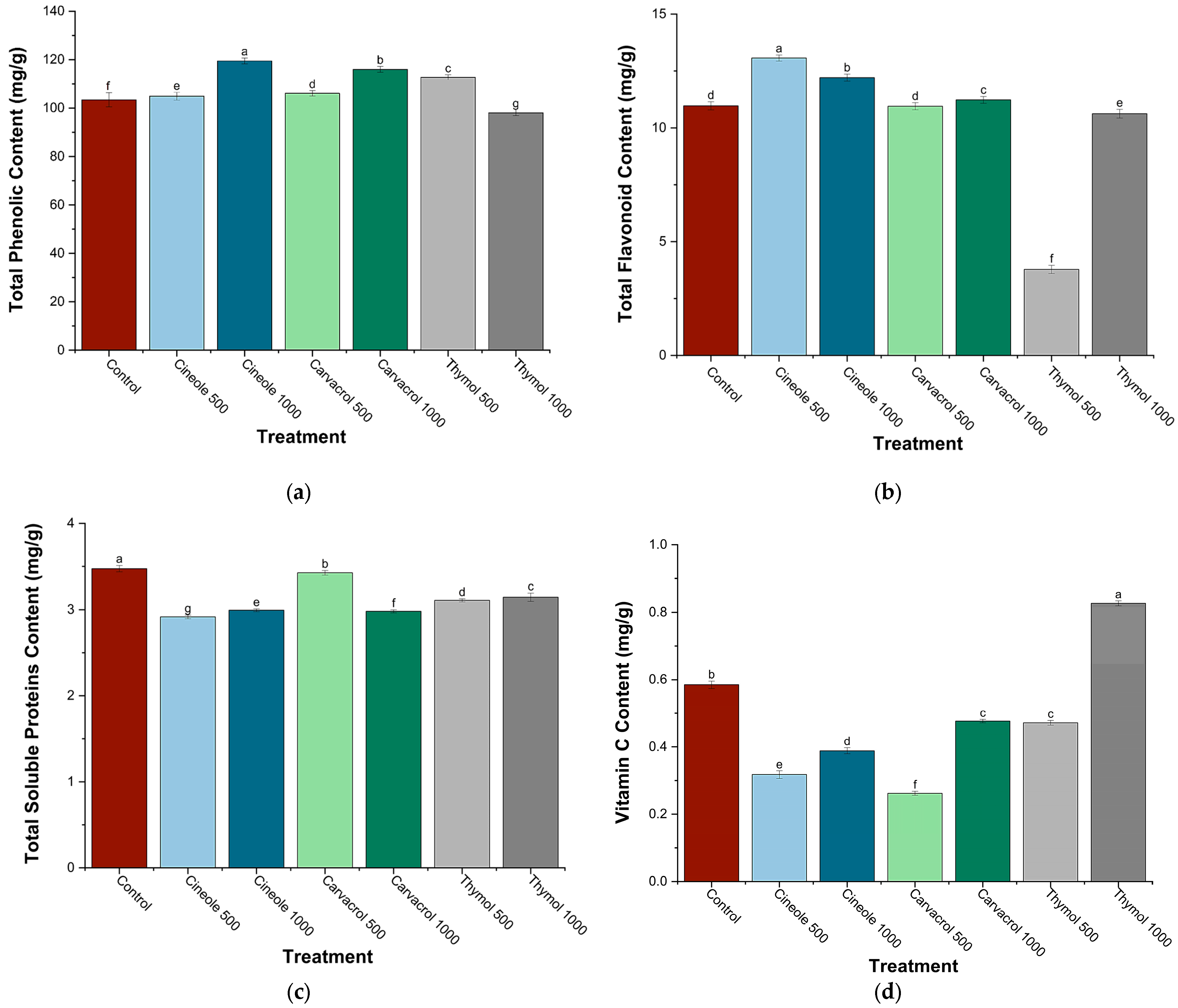
3.3. Phytochemical and Biochemical Parameters
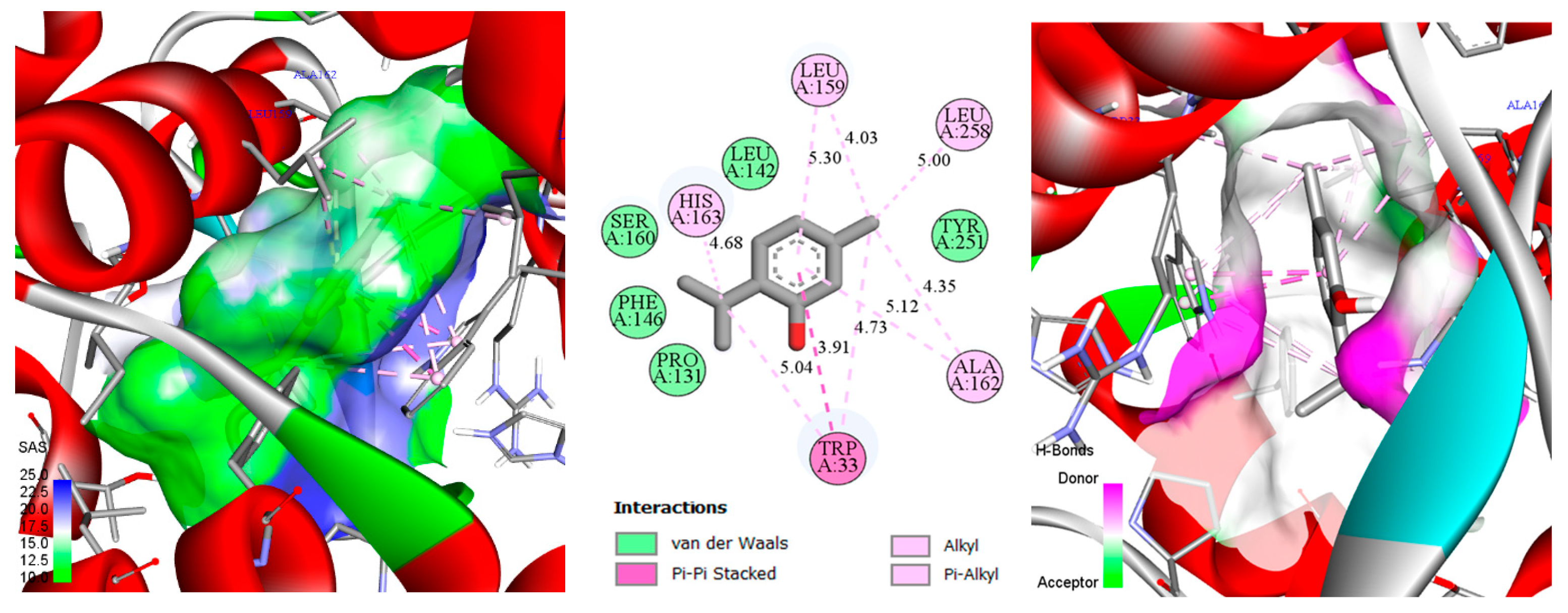
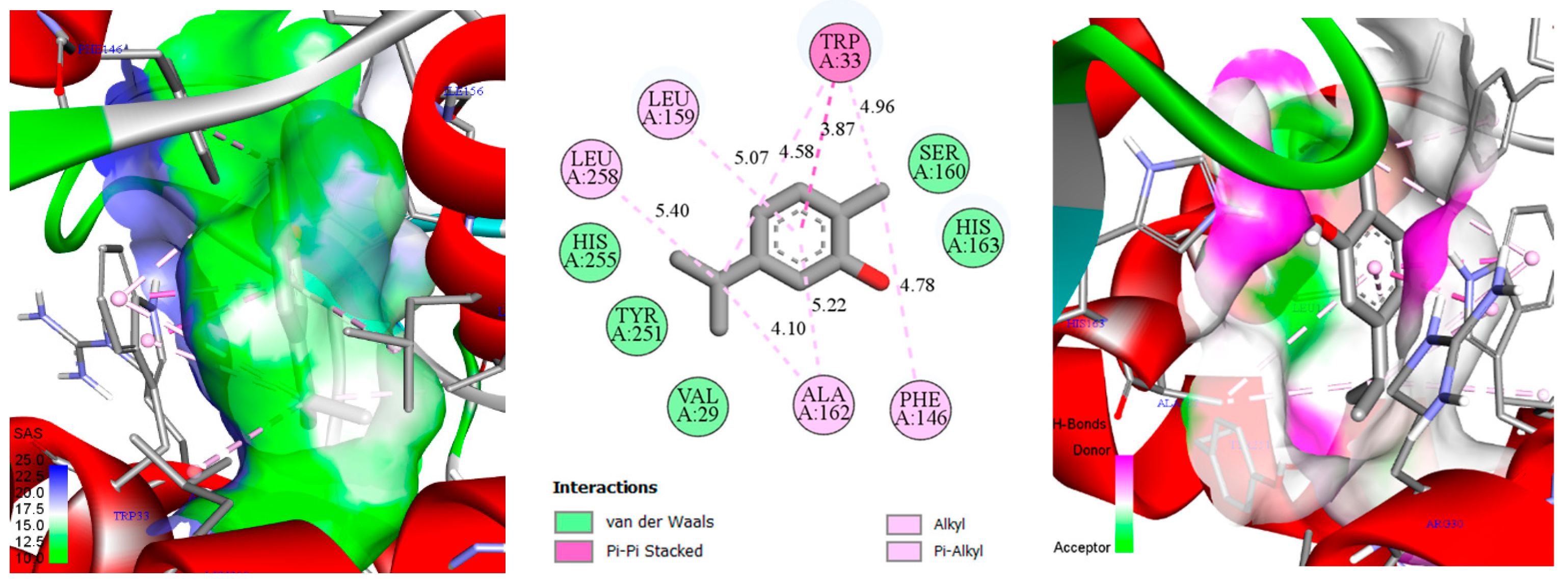
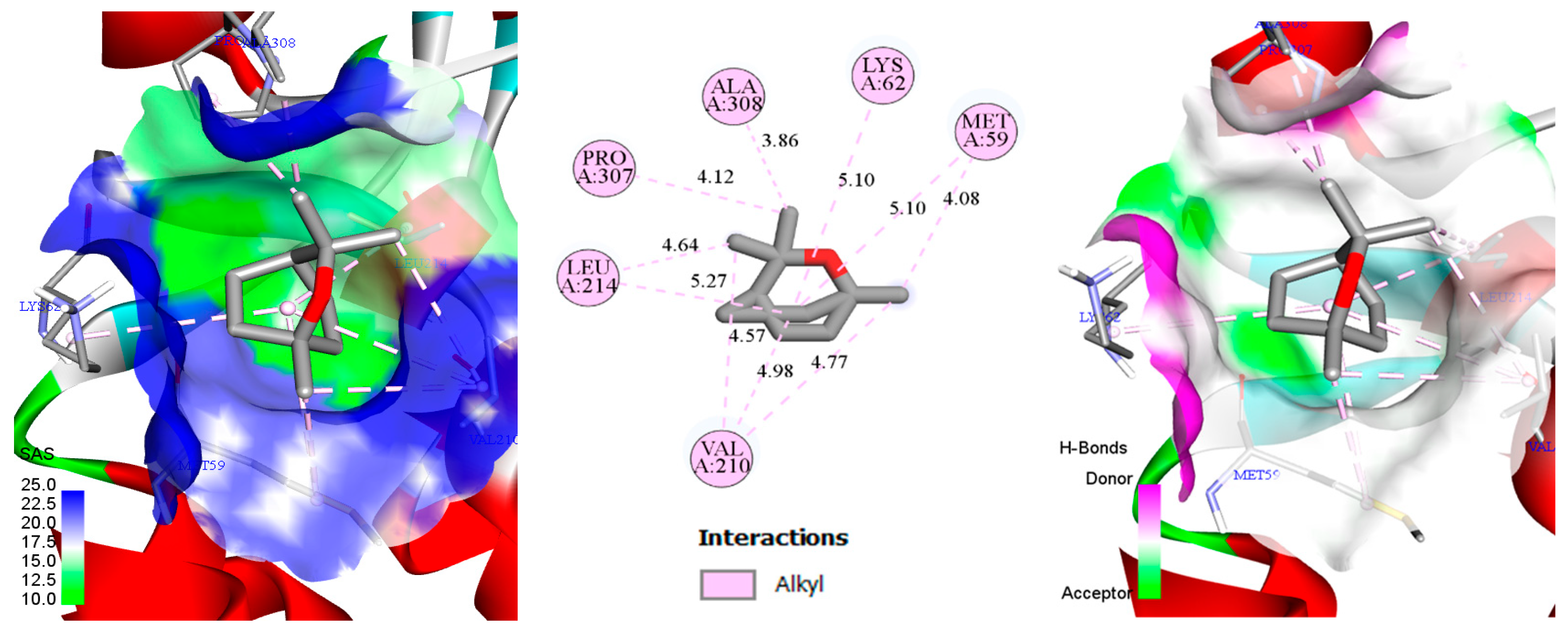
3.4. Docking of Monoterpenes with Chickpea Enzymes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Varshney, R.K.; Mohan, S.M.; Gaur, P.M.; Gangarao, N.V.P.R.; Pandey, M.K.; Bohra, A.; Sawargaonkar, S.L.; Chitikineni, A.; Kimurto, P.K.; Janila, P.; et al. Achievements and Prospects of Genomics-Assisted Breeding in Three Legume Crops of the Semi-Arid Tropics. Biotechnol. Adv. 2013, 31, 1120–1134. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Chávez, E.; Carrasco, B.; Cabeza, R.; Schwember, A.R. A Comprehensive Review on Chickpea (Cicer arietinum L.): Breeding for Abiotic Stress Tolerance and Climate Change Resilience. Agronomy 2022, 12, 1393. [Google Scholar] [CrossRef]
- Jukanti, A.K.; Gaur, P.M.; Gowda, C.L.; Chibbar, R.N. Nutritional Quality and Health Benefits of Chickpea (Cicer arietinum L.): A Review. Br. J. Nutr. 2012, 108, S11–S26. [Google Scholar] [CrossRef] [PubMed]
- Asati, R.; Tripathi, M.K.; Tiwari, S.; Yadav, R.K.; Tripathi, N. Molecular Breeding and Drought Tolerance in Chickpea. Life 2022, 12, 1846. [Google Scholar] [CrossRef]
- Houasli, C.; Idrissi, O.; Nsarellah, N. Chickpea Genetic Improvement in Morocco: State of the Art, Progress, and Prospects. Moroc. J. Agric. Sci. 2020, 1, 5–8. [Google Scholar]
- Houasli, C.; Sahri, A.; Nsarellah, N.; Idrissi, O. Chickpea (Cicer arietinum L.) Breeding in Morocco: Genetic Gain and Stability of Grain Yield and Seed Size under Winter Planting Conditions. Euphytica 2021, 217, 159. [Google Scholar] [CrossRef]
- Vaccino, P.; Antonetti, M.; Balconi, C.; Brandolini, A.; Cappellozza, S.; Caputo, A.R.; Carboni, A.; Caruso, M.; Copetta, A.; De Dato, G.; et al. Plant Genetic Resources for Food and Agriculture: The Role and Contribution of CREA (Italy) within the National Program RGV-FAO. Agronomy 2024, 14, 1263. [Google Scholar] [CrossRef]
- Abou Chehade, L.; Tavarini, S.; Bozzini, M.F.; Koskey, G.; Caturegli, L.; Antichi, D.; Angelini, L.G. Agronomic and Phytochemical Characterization of Chickpea Local Genetic Resources for the Agroecological Transition and Sustainable Food Systems. Agronomy 2024, 14, 2229. [Google Scholar] [CrossRef]
- Pacucci, G.; Troccoli, C.; Leoni, B. Effects of Supplementary Irrigation on Yield of Chickpea Genotypes in a Mediterranean Climate. Agric. Eng. Int. CIGR J. 2006, 8, 1–9. [Google Scholar]
- Landi, S.; Polito, F.; Esposito, S.; Sorbo, S.; Cianciullo, P.; Postiglione, A.; De Feo, V.; Basile, A.; Maresca, V. Protective Effect of Ocimum basilicum L. Essential Oil on Lactuca sativa L. Treated with Cadmium. Plant Physiol. Biochem. 2025, 219, 109437. [Google Scholar] [CrossRef]
- Oğuz, M.Ç.; Oğuz, E.; Güler, M. Seed Priming with Essential Oils for Sustainable Wheat Agriculture in Semi-Arid Region. PeerJ 2023, 11, e15126. [Google Scholar] [CrossRef] [PubMed]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial Activity of Some Essential Oils—Present Status and Future Perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, D.P.; Damasceno, R.O.S.; Amorati, R.; Elshabrawy, H.A.; De Castro, R.D.; Bezerra, D.P.; Nunes, V.R.V.; Gomes, R.C.; Lima, T.C. Essential Oils: Chemistry and Pharmacological Activities. Biomolecules 2023, 13, 1144. [Google Scholar] [CrossRef] [PubMed]
- Pezantes-Orellana, C.; Bermúdez, F.A.G.; De la Cruz, C.M.; Montalvo, J.L.; Orellana-Manzano, A. Essential Oils: A Systematic Review on Revolutionizing Health, Nutrition, and Omics for Optimal Well-Being. Front. Med. 2024, 11, 1337785. [Google Scholar] [CrossRef]
- Mandal, S.; Anand, U.; López-Bucio, J.; Kumar, M.; Lal, M.K.; Tiwari, R.K. Biostimulants and Environmental Stress Mitigation in Crops: A Novel and Emerging Approach for Agricultural Sustainability under Climate Change. Environ. Res. 2023, 233, 116357. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Y.; Liu, H.; Li, J.; Chen, X.; Li, Y. Dynamic Physiological Responses of Cinnamomum camphora with Different Chemotypes to High Temperature Stress. Forests 2023, 14, 2005. [Google Scholar] [CrossRef]
- Brunetti, C.; Fini, A.; Loreto, F.; Ferrini, F.; Tattini, M. Isoprenoids and Plant Resistance to Abiotic Stress: A Critical Review. Plant Cell Environ. 2013, 36, 1957–1969. [Google Scholar] [CrossRef]
- Zhou, H.; Ashworth, K.; Dodd, I.C. Exogenous Monoterpenes Mitigate H2O2-Induced Lipid Damage but Do Not Attenuate Photosynthetic Decline during Water Deficit in Tomato. J. Exp. Bot. 2023, 74, 5327–5340. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Martel, A.B.; Strugnell, C.A. Environmental Factors Regulate Plant Secondary Metabolites. Plants 2023, 12, 447. [Google Scholar] [CrossRef]
- Tian, J.; Zhang, Y.; Zuo, Z.; Wang, Y.; Wang, Y.; Liu, Y.; Zhang, L.; Wang, Y. The Uppermost Monoterpenes Improving Cinnamomum camphora Thermotolerance by Regulating Gene Expression in ROS Metabolism and Photosynthetic Capacity. Front. Plant Sci. 2022, 13, 1072931. [Google Scholar] [CrossRef]
- Zielińska-Błajet, M.; Feder-Kubis, J. Monoterpenes and Their Derivatives Recent Development in Biological and Medical Applications. Int. J. Mol. Sci. 2020, 21, 7078. [Google Scholar] [CrossRef] [PubMed]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant Biostimulants: Importance of the Quality and Yield of Horticultural Crops and the Improvement of Plant Tolerance to Abiotic Stress—A Review. Agronomy 2019, 9, 335. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Kaur, S. Eucalyptus Essential Oil as a Natural Pesticide. For. Ecol. Manag. 2008, 256, 2166–2174. [Google Scholar] [CrossRef]
- Kim, H.-M.; Kwon, H.; Kim, K.; Lee, S.-E. Antifungal and Antiaflatoxigenic Activities of 1,8-Cineole and t-Cinnamaldehyde on Aspergillus flavus. Appl. Sci. 2018, 8, 1655. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Varoni, E.M.; Iriti, M.; Martorell, M.; Setzer, W.N.; Del Mar Contreras, M.; Salehi, B.; Soltani-Nejad, A.; Rajabi, S.; Tajbakhsh, M.; et al. Carvacrol and Human Health: A Comprehensive Review. Phytother. Res. 2018, 32, 1675–1687. [Google Scholar] [CrossRef]
- Saghrouchni, H.; Barnossi, A.E.; Mssillou, I.; Lavkor, I.; Ay, T.; Kara, M.; Var, I. Potential of Carvacrol as Plant Growth-Promotor and Green Fungicide against Fusarium Wilt Disease of Perennial Ryegrass. Front. Plant Sci. 2023, 14, 973207. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Shukla, I.; Sharifi-Rad, M.; Del Mar Contreras, M.; Segura-Carretero, A.; Fathi, H.; Nasrabadi, N.N.; Kobarfard, F.; Sharifi-Rad, J. Thymol, Thyme, and Other Plant Sources: Health and Potential Uses. Phytother. Res. 2018, 32, 1688–1706. [Google Scholar] [CrossRef]
- Li, J.; Xu, W.; Zhao, Y.; Ji, X.; Peng, L.; Zhang, L.; Wang, Y.; Guo, J. Thymol Stimulates Lateral Root Formation via Regulating Endogenous Reactive Oxygen Species. Agronomy 2025, 15, 784. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef]
- Radkowski, A.; Radkowska, I.; Bocianowski, J.; Sladkovska, T.; Wolski, K. The Effect of Foliar Application of an Amino Acid-Based Biostimulant on Lawn Functional Value. Agronomy 2020, 10, 1656. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, B.; Wang, D.; Jia, X.; Zhang, Q.; Liu, W.; Zhou, H. Investigation into the Binding Site of (-)-Spirobrassinin for Herbicidal Activity Using Molecular Docking and Molecular Dynamics Simulations. Appl. Sci. 2023, 13, 7287. [Google Scholar] [CrossRef]
- Maffei, M.E.; Gertsch, J.; Appendino, G. Plant Volatiles: Production, Function and Pharmacology. Nat. Prod. Rep. 2011, 28, 1359–1380. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, J.K.; Gershenzon, J. Multiple Stress Factors and the Emission of Plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Zunino, M.P.; Zygadlo, J.A. Changes in the Composition of Phospholipid Fatty Acids and Sterols of Maize Root in Response to Monoterpenes. J. Chem. Ecol. 2005, 31, 1269–1283. [Google Scholar] [CrossRef]
- Caetano-Anollés, G.; Favelukes, G.; Bauer, W.D. Optimization of Surface Sterilization for Legume Seed. Crop Sci. 1990, 30, 708–712. [Google Scholar] [CrossRef]
- Maresca, V.; Fedeli, R.; Vannini, A.; Munzi, S.; Corrêa, A.; Cruz, C.; Loppi, S. Wood Distillate Enhances Seed Germination of Chickpea, Lettuce, and Basil. Appl. Sci. 2024, 14, 631. [Google Scholar] [CrossRef]
- Frailey, D.C.; Zhang, Q.; Wood, D.J.; Davis, T.M. Defining the Mutation Sites in Chickpea Nodulation Mutants PM233 and PM405. BMC Plant Biol. 2022, 22, 66. [Google Scholar] [CrossRef]
- Kosgei, A.J.; Kimurto, P.K.; Gaur, P.M.; Yeboah, M.A.; Offei, S.K.; Danquah, E.Y. Introgression of Drought Tolerance Root Traits into Kenyan Commercial Chickpea Varieties Using Marker-Assisted Backcrossing. Afr. Crop Sci. J. 2022, 30, 31–50. [Google Scholar] [CrossRef]
- Souri, M.K.; Bakhtiarizade, M. Biostimulation Effects of Rosemary Essential Oil on Growth and Nutrient Uptake of Tomato Seedlings. Sci. Hortic. 2019, 243, 472–476. [Google Scholar] [CrossRef]
- Dias, B.L.; Sarmento, R.A.; Venzon, M.; Jumbo, L.O.V.; Dos Santos, L.S.S.; De Souza Moura, W.; Mourão, D.d.S.C.; Fernandes, P.R.d.S.; Neitzke, T.R.; Oliveira, J.V.d.A.; et al. Morinda citrifolia Essential Oil: A Plant Resistance Biostimulant and a Sustainable Alternative for Controlling Phytopathogens and Insect Pests. Biology 2024, 13, 479. [Google Scholar] [CrossRef]
- Fedeli, R.; Vannini, A.; Celletti, S.; Maresca, V.; Munzi, S.; Cruz, C.; Alexandrov, D.; Guarnieri, M.; Loppi, S. Foliar Application of Wood Distillate Boosts Plant Yield and Nutritional Parameters of Chickpea. Ann. Appl. Biol. 2023, 182, 57–64. [Google Scholar] [CrossRef]
- Negusse, H.; Haileselassie, T.; Geleta, M.; Tesfaye, K. Genetic Variability of Ethiopian Chickpea (Cicer arietinum L.) Landraces for Acid Soil Tolerance. Plants 2025, 14, 311. [Google Scholar] [CrossRef]
- Xu, T.; Vo, Q.A.T.; Barnett, S.J.; Ballard, R.A.; Zhu, Y.; Franco, C.M.M. Revealing the Underlying Mechanisms Mediated by Endophytic Actinobacteria to Enhance the Rhizobia–Chickpea (Cicer arietinum L.) Symbiosis. Plant Soil 2022, 474, 299–318. [Google Scholar] [CrossRef]
- Fedeli, R.; Zhatkanbayeva, Z.; Loppi, S. Wood Distillate as a Solution for Growing Crops under Water Deficiency. Crops 2025, 5, 22. [Google Scholar] [CrossRef]
- Fedeli, R.; Zhatkanbayeva, Z.; Loppi, S. Soil Amendment with Biochar from Slaughterhouse Waste Bones Enhances Soil Quality and Promotes the Growth of Crop Plants. Plant Biosyst. 2025, 159, 378–386. [Google Scholar] [CrossRef]
- Fedeli, R.; Di Lella, L.A.; Loppi, S. Suitability of XRF for Routine Analysis of Multi-Elemental Composition: A Multi-Standard Verification. Methods Protoc. 2024, 7, 53. [Google Scholar] [CrossRef]
- Azarnejad, N.; Celletti, S.; Ghorbani, M.; Fedeli, R.; Loppi, S. Dose-Dependent Effects of a Corn Starch-Based Bioplastic on Basil (Ocimum basilicum L.): Implications for Growth, Biochemical Parameters, and Nutrient Content. Toxics 2024, 12, 80. [Google Scholar] [CrossRef]
- Lamaro, G.P.; Tsehaye, Y.; Girma, A.; Vannini, A.; Fedeli, R.; Loppi, S. Evaluation of Yield and Nutraceutical Traits of Orange-Fleshed Sweet Potato Storage Roots in Two Agro-Climatic Zones of Northern Ethiopia. Plants 2023, 12, 1319. [Google Scholar] [CrossRef]
- Fedeli, R.; Cruz, C.; Loppi, S.; Munzi, S. Hormetic Effect of Wood Distillate on Hydroponically Grown Lettuce. Plants 2024, 13, 447. [Google Scholar] [CrossRef]
- Fedeli, R.; Mazza, I.; Perini, C.; Salerni, E.; Loppi, S. New Frontiers in the Cultivation of Edible Fungi: The Application of Biostimulants Enhances the Nutritional Characteristics of Pleurotus eryngii (DC.) Quél. Agriculture 2024, 14, 1012. [Google Scholar] [CrossRef]
- Celletti, S.; Fedeli, R.; Ghorbani, M.; Loppi, S. Impact of Starch-Based Bioplastic on Growth and Biochemical Parameters of Basil Plants. Sci. Total Environ. 2023, 856, 159163. [Google Scholar] [CrossRef] [PubMed]
- Reinbothe, C.; El Bakkouri, M.; Buhr, F.; Muraki, N.; Nomata, J.; Kurisu, G.; Fujita, Y.; Reinbothe, S. Chlorophyll biosynthesis: Spotlight on protochlorophyllide reduction. Trends Plant Sci. 2010, 15, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.T.H.; Linthorst, H.J.M.; Verpoorte, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, C.-J. Multifaceted Regulations of Gateway Enzyme Phenylalanine Ammonia-Lyase in the Biosynthesis of Phenylpropanoids. Mol. Plant 2015, 8, 17–27. [Google Scholar] [CrossRef]
- Patel, P.K.; Hemantaranjan, A. Antioxidant Defence System in Chickpea (Cicer arietinum L.): Influence by Drought Stress Implemented at Pre- and Post-anthesis Stage. Am. J. Plant Physiol. 2012, 7, 164–173. [Google Scholar] [CrossRef]
- Parveen, S.; Gupta, D.B.; Dass, S.; Kumar, A.; Pandey, A.; Chakraborty, S.; Chakraborty, N. Chickpea Ferritin CaFer1 Participates in Oxidative Stress Response, and Promotes Growth and Development. Sci. Rep. 2016, 6, 31218. [Google Scholar] [CrossRef]
- DeLano, W.L. PyMOL: An Open-Source Molecular Graphics Tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]
- Huey, R.; Morris, G.M.; Forli, S. Using AutoDock 4 and AutoDock Vina with AutoDockTools: A Tutorial; The Scripps Research Institute Molecular Graphics Laboratory: La Jolla, CA, USA, 2012. [Google Scholar]
- Al-Dhahli, A.S.; Al-Hassani, F.A.; Alarjani, M. Essential Oil from the Rhizomes of the Saudi and Chinese Zingiber officinale Cultivars: Comparison of Chemical Composition, Antibacterial and Molecular Docking Studies. J. King Saud Univ. Sci. 2020, 32, 3343–3350. [Google Scholar] [CrossRef]
- Jejurikar, B.L.; Rohane, S.H. Drug Designing in Discovery Studio. Asian J. Res. Chem. 2021, 14, 135–138. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High Performance Molecular Simulations through Multi-Level Parallelism from Laptops to Supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Khush, G.S. Green Revolution: The Way Forward. Nat. Rev. Genet. 2001, 2, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Rademacher, W. Plant Growth Regulators: Backgrounds and Uses in Plant Production. J. Plant Growth Regul. 2015, 34, 845–872. [Google Scholar] [CrossRef]
- Erofeeva, E.A. A Method for Assessing the Frequency of Hormetic Trade-Offs in Plants. MethodsX 2021, 8, 101400. [Google Scholar] [CrossRef]
- Heumann, S.; Fier, A.; Haßdenteufel, M.; Höper, H.; Schäfer, M. Inhibition of Nitrification by Essential Oils and Their Components. J. Plant Nutr. Soil. Sci. 2013, 176, 387–396. [Google Scholar] [CrossRef]
- Araniti, F.; Miras-Moreno, B.; Lucini, L.; Landi, M.; Abenavoli, M.R. Metabolomic, Proteomic and Physiological Insights into the Potential Mode of Action of Thymol, a Phytotoxic Natural Monoterpenoid Phenol. Plant Physiol. Biochem. 2020, 153, 141–153. [Google Scholar] [CrossRef]
- Kordali, S.; Cakir, A.; Mavi, A.; Kilic, H.; Yildirim, A. Antifungal, Phytotoxic and Insecticidal Properties of Essential Oil Isolated from Turkish Origanum acutidens and Its Three Components, Carvacrol, Thymol and p-Cymene. Bioresour. Technol. 2008, 99, 8788–8795. [Google Scholar] [CrossRef]
- Santos, C.V.; Rao, I.M. Calcium’s Role as a Second Messenger in Stress Responses. Plant Physiol. Biochem. 2000, 38, 403–414. [Google Scholar] [CrossRef]
- Muñoz, E.; García, M.; Domínguez-Valencia, A.; González-Coloma, A. Phytotoxic Effects of Three Natural Compounds: Pelargonic Acid, Carvacrol, and Cinnamic Aldehyde against Problematic Weeds in Mediterranean Crops. Agronomy 2020, 10, 791. [Google Scholar] [CrossRef]
- Kaur, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Essential Oil of Spearmint (Mentha spicata L.) Exhibits Allelopathic Activity and Antioxidant Potential. J. Plant Interact. 2020, 15, 313–319. [Google Scholar] [CrossRef]
- Miguel, M.G. Antioxidant Activity of Medicinal and Aromatic Plants: A Review. Flavour. Fragr. J. 2010, 25, 291–312. [Google Scholar] [CrossRef]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of Phenolics in the Resistance Mechanisms of Plants against Fungal Pathogens and Insects. In Phytochemistry: Advances in Research; Research Signpost: Trivandrum, India, 2006; pp. 23–67. [Google Scholar]
- Gholami, H.; Saharkhiz, M.J.; Fard, F.R.; Ghani, A.; Nadaf, F. Effect of Thymol on Antioxidant Activity, Phenolic and Flavonoid Content in Dracocephalum moldavica under Saline Conditions. J. Essent. Oil Bear. Plants 2021, 24, 756–768. [Google Scholar] [CrossRef]
- Kleinwächter, M.; Selmar, D. New Insights Explain That Drought Stress Enhances the Quality of Spice and Medicinal Plants: Potential Applications. Agron. Sustain. Dev. 2015, 35, 121–131. [Google Scholar] [CrossRef]
- Mahmood, T.; Rana, R.M.; Ahmar, S.; Saeed, S.; Gulzar, A.; Khan, M.A. Effect of Drought Stress on Phenolic Accumulation and Antioxidant Activity in Chickpea (Cicer arietinum L.) Genotypes. Acta Physiol. Plant. 2018, 40, 1–12. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanisms in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox Regulation in Photosynthetic Organisms: Signaling, Acclimation, and Practical Implications. Antioxid. Redox Signal. 2009, 11, 861–905. [Google Scholar] [CrossRef]
- Chavan, P.D.; Nikam, T.D. Stress-Induced Changes in Protein Metabolism in Chickpea (Cicer arietinum L.) Cultivars Differing in Drought Tolerance. Acta Physiol. Plant. 2017, 39, 120. [Google Scholar] [CrossRef]
- Hossain, M.A.; Wani, S.H.; Bhattacharjee, S.; Burritt, D.J.; Tran, L.-S.P. Drought Stress Tolerance in Plants, Vol. 1: Physiology and Biochemistry; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Patil, G.; Mian, R.; Vuong, T.; Nguyen, H.T. Molecular and Functional Characterization of Drought Tolerance in Soybean. Genom. Insights 2012, 5, 1–11. [Google Scholar] [CrossRef]
- Smirnoff, N. Ascorbic Acid Metabolism and Functions: A Comparison of Plants and Mammals. Free Radic. Biol. Med. 2018, 122, 116–129. [Google Scholar] [CrossRef]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic Acid—A Potential Oxidant Scavenger and Its Role in Plant Development and Abiotic Stress Tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef]
- Gallie, D.R. The Role of L-Ascorbic Acid Recycling in Responding to Environmental Stress and in Promoting Plant Growth. J. Exp. Bot. 2013, 64, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Araniti, F.; Sorgonà, A.; Lupini, A.; Mauceri, A.; Abenavoli, M.R. Thymol, Carvacrol, and Eugenol Induce Differential Oxidative Stress and Antioxidant Responses in Basil (Ocimum basilicum L.) and Peppermint (Mentha piperita L.). Plant Physiol. Biochem. 2018, 127, 229–239. [Google Scholar] [CrossRef]
- Chroho, M.; Rouphael, Y.; Petropoulos, S.A.; Bouissane, L. Carvacrol and Thymol Content Affects the Antioxidant and Antibacterial Activities of Oregano (Origanum compactum) and Thyme (Thymus zygis) Essential Oils. Antibiotics 2024, 13, 139. [Google Scholar] [CrossRef] [PubMed]
- Natal, C.M.; Fernandes, M.J.G.; Pinto, N.F.S.; Pereira, R.B.; Vieira, T.F.; Rodrigues, A.R.O.; Pereira, D.M.; Sousa, S.F.; Fortes, A.G.; Castanheira, E.M.S.; et al. New Carvacrol and Thymol Derivatives as Potential Insecticides: Synthesis, Biological Activity, Computational Studies and Nanoencapsulation. RSC Adv. 2021, 11, 34024–34036. [Google Scholar] [CrossRef] [PubMed]
- Spyrou, S.; Bellou, M.G.; Papanikolaou, A.; Nakou, K.; Kontogianni, V.G.; Chatzikonstantinou, A.V.; Stamatis, H. Evaluation of Antioxidant, Antibacterial and Enzyme-Inhibitory Properties of Dittany and Thyme Extracts and Their Application in Hydrogel Preparation. BioChem 2024, 4, 166–188. [Google Scholar] [CrossRef]
- Dong, C.S.; Zhang, W.L.; Wang, Q.; Li, Y.S.; Wang, X.; Zhang, M.; Liu, L. Crystal Structures of Cyanobacterial Light-Dependent Protochlorophyllide Oxidoreductase. Proc. Natl. Acad. Sci. USA 2020, 117, 8455–8461. [Google Scholar] [CrossRef]
- Jez, J.M.; Bowman, M.E.; Noel, J.P. Structure-Guided Programming of Polyketide Chain-Length Determination in Chalcone Synthase. Biochemistry 2001, 40, 14829–14838. [Google Scholar] [CrossRef]
- Jun, S.Y.; Sattler, S.A.; Cortez, G.S.; Vermerris, W.; Sattler, S.E.; Kang, C. Biochemical and Structural Analysis of Substrate Specificity of a Phenylalanine Ammonia-Lyase. Plant Physiol. 2018, 176, 1452–1468. [Google Scholar] [CrossRef]
- Wada, K.; Tada, T.; Nakamura, Y.; Ishikawa, T.; Yabuta, Y.; Yoshimura, K.; Nishimura, K. Crystal Structure of Chloroplastic Ascorbate Peroxidase from Tobacco Plants and Structural Insights into Its Instability. J. Biochem. 2003, 134, 239–244. [Google Scholar] [CrossRef]
- Arosio, P.; Elia, L.; Poli, M. Ferritin, Cellular Iron Storage and Regulation. IUBMB Life 2017, 69, 414–422. [Google Scholar] [CrossRef]
- Muhammad, S.A.; Fatima, N. In Silico Analysis and Molecular Docking Studies of Potential Angiotensin-Converting Enzyme Inhibitor Using Quercetin Glycosides. Pharmacogn. Mag. 2015, 11 (Suppl. S1), S123. [Google Scholar] [CrossRef] [PubMed]
- Bouammali, H.; Zraibi, L.; Ziani, I.; Merzouki, M.; Bourassi, L.; Fraj, E.; Touzani, R. Rosemary as a Potential Source of Natural Antioxidants and Anticancer Agents: A Molecular Docking Study. Plants 2023, 13, 89. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Alshrari, A.S.; Hafiz, M.N.; Jawad, M.M.; Khan, A.; Alanazi, F.J.; Asdaq, S.M.B. Exploring Therapeutic Paradigm Focusing on Genes, Proteins, and Pathways to Combat Leprosy and Tuberculosis: A Network Medicine and Drug Repurposing Approach. J. Infect. Public Health 2025, 18, 102763. [Google Scholar] [CrossRef] [PubMed]
- Camargo, P.G.; da Silva, R.B.; Zuma, A.A.; Garden, S.J.; Albuquerque, M.G.; Rodrigues, C.R.; da Silva Lima, C.H. In Silico Evaluation of N-Aryl-1,10-Phenanthroline-2-Amines as Potential Inhibitors of T. cruzi GP63 Zinc-Metalloprotease by Docking and Molecular Dynamics Simulations. Sci. Rep. 2025, 15, 6036. [Google Scholar] [CrossRef]
- Lazzarotto, F.; Menguer, P.K.; Del-Bem, L.E.; Zámocký, M.; Margis-Pinheiro, M. Ascorbate Peroxidase Neofunctionalization at the Origin of APX-R and APX-L: Evidence from Basal Archaeplastida. Antioxidants 2021, 10, 597. [Google Scholar] [CrossRef]
- Gu, S.; Shen, C.; Yu, J.; Zhao, H.; Liu, H.; Liu, L.; Kang, Y. Can Molecular Dynamics Simulations Improve Predictions of Protein–Ligand Binding Affinity with Machine Learning? Brief Bioinform. 2023, 24, bbad008. [Google Scholar] [CrossRef]
- Murphy, K.M.; Reeves, P.G.; Jones, S.S. Relationship between Yield and Mineral Nutrient Concentrations in Historical and Modern Spring Wheat Cultivars. Euphytica 2008, 163, 381–390. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mineral Elements | Control | Cineole | Carvacrol | Thymol | |||
|---|---|---|---|---|---|---|---|
| 500 ppm | 1000 ppm | 500 ppm | 1000 ppm | 500 ppm | 1000 ppm | ||
| P | 8716 ± 68 ab | 8313 ± 56 c | 8870 ± 61 a | 7471 ± 49 d | 6797 ± 28 e | 8553 ± 27 b | 7000 ± 17 e |
| S | 3920 ± 21 a | 3376 ± 43 b | 2704 ± 33 e | 2854 ± 21 d | 3243 ± 16 c | 3196 ± 25 c | 2967 ± 27 d |
| Cl | 1769 ± 20 d | 2093 ± 12 b | 2352 ± 17 a | 2153 ± 17 b | 1923 ± 11 c | 1872 ± 16 c | 1411 ± 13 e |
| K | 27,445 ± 130 a | 26,356 ± 163 b | 23,495 ± 93 d | 24,634 ± 60 c | 23,051 ± 91 d | 26,252 ± 81 b | 21,531 ± 113 e |
| Ca | 7649 ± 39 c | 8303 ± 45 b | 9984 ± 39 a | 10,096 ± 39 a | 6789 ± 19 d | 9944 ± 36 a | 6758 ± 27 d |
| Mn | 45.1 ± 0.5 b | 36.7 ± 0.7 c | 39.7 ± 0.8 c | 48.4 ± 0.3 a | 29.9 ± 0.6 d | 45.0 ± 1.1 b | 22.9 ± 0.7 e |
| Fe | 60.7 ± 2.3 a | 38.4 ± 0.9 cd | 44.0 ± 1.3 bc | 49.9 ± 1.5 b | 36.5 ± 1.7 d | 48.7 ± 1.5 b | 29.6 ± 0.2 e |
| Cu | 3.6 ± 0.4 a | 3.2 ± 0.2 ab | 2.4 ± 0.2 ab | 3.2 ± 0.3 ab | 3.0 ± 0.2 ab | 3.5 ± 0.2 ab | 2.3 ± 0.1 b |
| Zn | 50.7 ± 0.4 c | 48.2 ± 0.5 c | 53.8 ± 0.3 b | 57.8 ± 1.0 a | 41.4 ± 1.0 d | 58.9 ± 0.6 a | 29.5 ± 0.6 e |
| Molecules | Docking Score (kcal/mol) | ||||
|---|---|---|---|---|---|
| 6L1H | 1I89 | 6AT7 | 1IYN | 3OGH | |
| Thymol | −6.4 | −6.1 | −6.7 | −6.7 | −5.3 |
| Carvacrol | −6.2 | −5.1 | −6.5 | −6.5 | −5.3 |
| Cineole | −6.1 | −5.2 | −5.4 | −5.7 | −4.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Et-Tazy, L.; Fedeli, R.; Khibech, O.; Lamiri, A.; Challioui, A.; Loppi, S. Effects of Monoterpene-Based Biostimulants on Chickpea (Cicer arietinum L.) Plants: Functional and Molecular Insights. Biology 2025, 14, 657. https://doi.org/10.3390/biology14060657
Et-Tazy L, Fedeli R, Khibech O, Lamiri A, Challioui A, Loppi S. Effects of Monoterpene-Based Biostimulants on Chickpea (Cicer arietinum L.) Plants: Functional and Molecular Insights. Biology. 2025; 14(6):657. https://doi.org/10.3390/biology14060657
Chicago/Turabian StyleEt-Tazy, Lamyae, Riccardo Fedeli, Oussama Khibech, Abdeslam Lamiri, Allal Challioui, and Stefano Loppi. 2025. "Effects of Monoterpene-Based Biostimulants on Chickpea (Cicer arietinum L.) Plants: Functional and Molecular Insights" Biology 14, no. 6: 657. https://doi.org/10.3390/biology14060657
APA StyleEt-Tazy, L., Fedeli, R., Khibech, O., Lamiri, A., Challioui, A., & Loppi, S. (2025). Effects of Monoterpene-Based Biostimulants on Chickpea (Cicer arietinum L.) Plants: Functional and Molecular Insights. Biology, 14(6), 657. https://doi.org/10.3390/biology14060657