
Differential Effects of Rutin and Its Aglycone Quercetin on Cytotoxicity and Chemosensitization of HCT 116 Colon Cancer Cells to Anticancer Drugs 5-Fluorouracil and Doxorubicin



Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Chemicals and Antibodies
2.2. Cell Treatments
2.3. Cell Viability Assay
2.4. Western Blot
2.5. Immunofluorescence
2.6. Statistical Analysis
3. Results
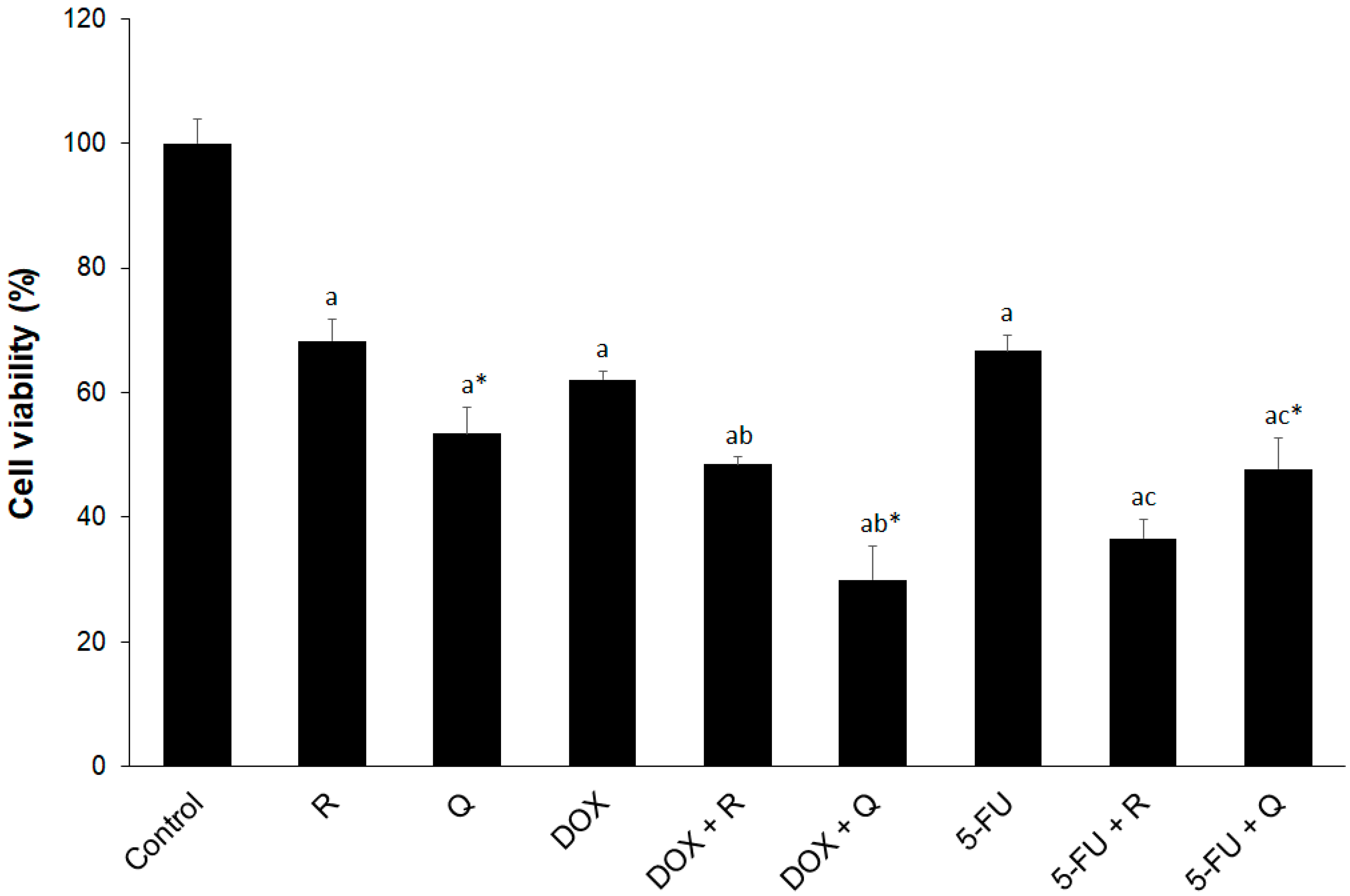
3.1. The Cytotoxicity of Rutin, Quercetin, DOX, and 5-FU
3.2. The Expression of Antioxidant Enzymes in the Treatments
3.3. The Effect of the Treatments on Cell Cycle Proteins
3.4. The Activation of Apoptotic Cell Death and Autophagy
3.5. The Modulation of Key Signaling Pathways
3.6. Autophagy Played a Dominantly Protective Role in the Treatments
3.7. HCT116 Treatments Induced FOXO3a Nuclear Accumulation
4. Discussion
Immediate Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Vodenkova, S.; Buchler, T.; Cervena, K.; Veskrnova, V.; Vodicka, P.; Vymetalkova, V. 5-fluorouracil and other fluoropyrimidines in colorectal cancer: Past, present and future. Pharmacol. Ther. 2020, 206, 107447. [Google Scholar] [CrossRef] [PubMed]
- Khaleel, S.A.; Al-Abd, A.M.; Ali, A.A.; Abdel-Naim, A.B. Didox and resveratrol sensitize colorectal cancer cells to doxorubicin via activating apoptosis and ameliorating P-glycoprotein activity. Sci. Rep. 2016, 6, 36855. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, R.; Ilari, A.; Colotti, B.; Mosca, L.; Fazi, F.; Colotti, G. Doxorubicin and other anthracyclines in cancers: Activity, chemoresistance and its overcoming. Mol. Asp. Med. 2003, 93, 101205. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- de Oliveira Júnior, R.G.; Adrielly, A.F.C.; da Silva Almeida, J.R.G.; Grougnet, R.; Thiéry, V.; Picot, L. Sensitization of tumor cells to chemotherapy by natural products: A systematic review of preclinical data and molecular mechanisms. Fitoterapia 2018, 129, 383–400. [Google Scholar] [CrossRef]
- Widyananda, M.H.; Pratama, S.K.; Ansori, A.N.M.; Antonius, Y.; Kharisma, V.D.; Murtadlo, A.A.A.; Jakhmola, V.; Rebezov, M.; Khayrullin, M.; Derkho, M.; et al. Quercetin as an anticancer candidate for glioblastoma multiforme by targeting AKT1, MMP9, ABCB1, and VEGFA: An in silico study. Karbala Int. J. Mod. Sci. 2023, 9, 10. [Google Scholar] [CrossRef]
- Magar, R.T.; Sohng, J.K. A review on structure, modifications and structure-activity relation of quercetin and its derivatives. J. Microbiol. Biotechnol. 2020, 30, 11–20. [Google Scholar] [CrossRef]
- Domitrović, R.; Jakovac, H.; Marchesi, V.V.; Vladimir-Knežević, S.; Cvijanović, O.; Tadić, Ž.; Romić, Ž.; Rahelić, D. Differential hepatoprotective mechanisms of rutin and quercetin in CCl4-intoxicated BALB/cN mice. Acta Pharmacol. Sin. 2012, 33, 1260–1270. [Google Scholar] [CrossRef]
- Ghanbari-Movahed, M.; Mondal, A.; Farzaei, M.H.; Bishayee, A. Quercetin- and rutin-based nano-formulations for cancer treatment: A systematic review of improved efficacy and molecular mechanisms. Phytomedicine 2022, 97, 153909. [Google Scholar] [CrossRef]
- Tang, Z.; Wang, L.; Chen, Y.; Zheng, X.; Wang, R.; Liu, B.; Zhang, S.; Wang, H. Quercetin reverses 5-fluorouracil resistance in colon cancer cells by modulating the NRF2/HO-1 pathway. Eur. J. Histochem. 2023, 67, 3719. [Google Scholar] [CrossRef] [PubMed]
- Roman, A.; Smeu, A.; Lascu, A.; Dehelean, C.A.; Predescu, I.A.; Motoc, A.; Borza, C.; Draghici, G.A.; Trandafirescu, C.M.; Anton, A.; et al. Quercetin enhances 5-fluorouracil-driven cytotoxicity dose-dependently in A375 Human melanoma cells. Life 2024, 14, 1685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Gao, M.; Wang, Z.; Zhang, J.; Cui, W.; Li, J.; Zhu, X.; Zhang, H.; Yang, D.H.; Xu, X. Curcumin reverses doxorubicin resistance in colon cancer cells at the metabolic level. J. Pharm. Biomed. Anal. 2021, 201, 114129. [Google Scholar] [CrossRef] [PubMed]
- Pawar, C.S.; Prasad, N.R.; Yadav, P.; Enoch, I.V.M.V.; Manikantan, V.; Dey, B.; Baruah, P. Enhanced delivery of quercetin and doxorubicin using β-cyclodextrin polymer to overcome P-glycoprotein mediated multidrug resistance. Int. J. Pharm. 2023, 635, 122763. [Google Scholar] [CrossRef]
- Potočnjak, I.; Šimić, L.; Vukelić, I.; Batičić, L.; Domitrović, R. Oleanolic acid induces HCT116 colon cancer cell death through the p38/FOXO3a/Sirt6 pathway. Chem. Biol. Interact. 2022, 363, 110010. [Google Scholar] [CrossRef]
- Chicote, J.; Yuste, V.J.; Boix, J.; Ribas, J. Cell death triggered by the autophagy inhibitory drug 3-methyladenine in growing conditions proceeds with DNA damage. Front. Pharmacol. 2020, 11, 580343. [Google Scholar] [CrossRef]
- Cheng, Y.P.; Li, S.; Chuang, W.L.; Li, C.H.; Chen, G.J.; Chang, C.C.; Or, C.R.; Lin, P.Y.; Chang, C.C. Blockade of STAT3 Signaling contributes to anticancer effect of 5-acetyloxy-6,7,8,4′-tetra-methoxyflavone, a tangeretin derivative, on human glioblastoma multiforme cells. Int. J. Mol. Sci. 2019, 20, 3366. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Catalano, T.; Selvaggi, F.; Cotellese, R.; Aceto, G.M. The role of reactive oxygen species in colorectal cancer initiation and progression: Perspectives on theranostic approaches. Cancers 2025, 17, 752. [Google Scholar] [CrossRef]
- Visconti, R.; Della Monica, R.; Grieco, D. Cell cycle checkpoint in cancer: A therapeutically targetable double-edged sword. J. Exp. Clin. Cancer Res. 2016, 35, 153. [Google Scholar] [CrossRef]
- Liu, Y.; Ao, X.; Ding, W.; Ponnusamy, M.; Wu, W.; Hao, X.; Yu, W.; Wang, Y.; Li, P.; Wang, J. Critical role of FOXO3a in carcinogenesis. Mol. Cancer 2018, 17, 104. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Bao, Q.; Yang, T.; Yang, M.; Mao, C. Precision treatment of colon cancer using doxorubicin-loaded metal-organic-framework-coated magnetic nanoparticles. ACS Appl. Mater. Interfaces 2024, 16, 49003–49012. [Google Scholar] [CrossRef]
- Markandeywar, T.S.; Narang, R.K.; Singh, D.; Rai, V.K. Targeted delivery of doxorubicin as a potential chemotherapeutic agent. Curr. Drug Deliv. 2023, 20, 904–918. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhu, L. IKKα kinase silencing increases doxorubicin-induced apoptosis through regulation of oxidative DNA damage response in colon cancer cells. Chem. Biol. Drug Des. 2023, 101, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Xiong, S.; Xiao, G.W. Reverting doxorubicin resistance in colon cancer by targeting a key signaling protein, steroid receptor coactivator. Exp. Ther. Med. 2018, 15, 3751–3758. [Google Scholar] [CrossRef]
- Bardelčíková, A.; Šoltys, J.; Mojžiš, J. Oxidative stress, inflammation and colorectal cancer: An overview. Antioxidants 2023, 12, 901. [Google Scholar] [CrossRef]
- Xie, Y.; Kang, R.; Klionsky, D.J.; Tang, D. GPX4 in cell death, autophagy, and disease. Autophagy 2023, 19, 2621–2638. [Google Scholar] [CrossRef]
- Meng, Q.; Xu, J.; Liang, C.; Liu, J.; Hua, J.; Zhang, Y.; Ni, Q.; Shi, S.; Yu, X. GPx1 is involved in the induction of protective autophagy in pancreatic cancer cells in response to glucose deprivation. Cell Death Dis. 2018, 9, 1187. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, W.; Razak, S.R.A.; Han, T.; Ahmad, N.H.; Li, X. Ferroptosis as a potential target for cancer therapy. Cell Death Dis. 2023, 14, 460. [Google Scholar] [CrossRef]
- Chiang, S.-K.; Chen, S.-E.; Chang, L.-C. A Dual role of heme oxygenase-1 in cancer cells. Int. J. Mol. Sci. 2018, 20, 39. [Google Scholar] [CrossRef]
- Yun, H.R.; Jo, Y.H.; Kim, J.; Shin, Y.; Kim, S.S.; Choi, T.G. Roles of autophagy in oxidative stress. Int. J. Mol. Sci. 2020, 21, 3289. [Google Scholar] [CrossRef] [PubMed]
- Filomeni, G.; De Zio, D.; Cecconi, F. Oxidative stress and autophagy: The clash between damage and metabolic needs. Cell Death Differ. 2015, 22, 377–388. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.-Y.; Kim, J.; Yoon, N.; Maeng, L.-S.; Byun, J.H. Prognostic potential of cyclin d1 expression in colorectal cancer. J. Clin. Med. 2023, 12, 572. [Google Scholar] [CrossRef] [PubMed]
- Ravizza, R.; Gariboldi, M.B.; Passarelli, L.; Monti, E. Role of the p53/p21 system in the response of human colon carcinoma cells to doxorubicin. BMC Cancer 2004, 4, 92. [Google Scholar] [CrossRef]
- Mansilla, S.F.; De La Vega, M.B.; Calzetta, N.L.; Siri, S.O.; Gottifredi, V. CDK-independent and PCNA-dependent functions of p21 in DNA replication. Genes 2020, 11, 593. [Google Scholar] [CrossRef]
- Salanci, Š.; Vilková, M.; Martinez, L.; Mirossay, L.; Michalková, R.; Mojžiš, J. The induction of G2/M phase cell cycle arrest and apoptosis by the chalcone derivative 1C in sensitive and resistant ovarian cancer cells is associated with ROS generation. Int. J. Mol. Sci. 2024, 25, 7541. [Google Scholar] [CrossRef]
- Schimek, V.; Strasser, K.; Beer, A.; Göber, S.; Walterskirchen, N.; Brostjan, C.; Müller, C.; Bachleitner-Hofmann, T.; Bergmann, M.; Dolznig, H.; et al. Tumour cell apoptosis modulates the colorectal cancer immune microenvironment via interleukin-8-dependent neutrophil recruitment. Cell Death Dis. 2022, 13, 113. [Google Scholar] [CrossRef]
- Julien, O.; Wells, J.A. Caspases and their substrates. Cell Death Differ. 2017, 24, 1380–1389. [Google Scholar] [CrossRef]
- Chaitanya, G.V.; Alexander, J.S.; Babu, P.P. PARP-1 cleavage fragments: Signatures of cell-death proteases in neurodegeneration. Cell Commun. Signal. 2010, 8, 31. [Google Scholar] [CrossRef]
- Dent, P. Crosstalk between ERK, AKT, and cell survival. Cancer Biol. Ther. 2014, 15, 245–246. [Google Scholar] [CrossRef]
- Villanueva-Paz, M.; Cotán, D.; Garrido-Maraver, J.; Oropesa-Ávila, M.; de la Mata, M.; Delgado-Pavón, A.; de Lavera, I.; Alcocer-Gómez, E.; Álvarez-Córdoba, M.; Sánchez-Alcázar, J.A. AMPK regulation of cell growth, apoptosis, autophagy, and bioenergetics. Exp. Suppl. 2016, 107, 45–71. [Google Scholar] [CrossRef] [PubMed]
- Folkerts, H.; Hilgendorf, S.; Vellenga, E.; Bremer, E.; Wiersma, V.R. The multifaceted role of autophagy in cancer and the microenvironment. Med. Res. Rev. 2019, 39, 517–560. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Gao, H.; Su, X. Autophagy-related signaling pathways are involved in cancer (Review). Exp. Ther. Med. 2021, 22, 710. [Google Scholar] [CrossRef] [PubMed]
- Koukourakis, M.I.; Kalamida, D.; Giatromanolaki, A.; Zois, C.E.; Sivridis, E.; Pouliliou, S.; Mitrakas, A.; Gatter, K.C.; Harris, A.L. Autophagosome proteins LC3A, LC3B and LC3C have distinct subcellular distribution kinetics and expression in cancer cell lines. PLoS ONE 2015, 10, e0137675. [Google Scholar] [CrossRef]
- Kumar, A.V.; Mills, J.; Lapierre, L.R. Selective autophagy receptor p62/SQSTM1, a pivotal player in stress and aging. Front. Cell Dev. Biol. 2022, 10, 793328. [Google Scholar] [CrossRef]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in mammalian autophagy research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef]
- de Mattos, S.F.; Villalonga, P.; Clardy, J.; Lam, E.W.-F. FOXO3a mediates the cytotoxic effects of cisplatin in colon cancer cells. Mol. Cancer Ther. 2008, 7, 3237–3246. [Google Scholar] [CrossRef]
- Gao, Z.; Li, Z.; Liu, Y.; Liu, Z. Forkhead box O3 promotes colon cancer proliferation and drug resistance by activating MDR1 expression. Mol. Genet. Genom. Med. 2019, 7, e554. [Google Scholar] [CrossRef]
- Hornsveld, M.; Dansen, T.B.; Derksen, P.W.; Burgering, B.M.T. Re-evaluating the role of FOXOs in cancer. Semin. Cancer Biol. 2018, 50, 90–100. [Google Scholar] [CrossRef]
- Gómez-Puerto, M.C.; Verhagen, L.P.; Braat, A.K.; Lam, E.W.F.; Coffer, P.J.; Lorenowicz, M.J. Activation of autophagy by FOXO3 regulates redox homeostasis during osteogenic differentiation. Autophagy 2016, 12, 1804–1816. [Google Scholar] [CrossRef]
- Wang, X.; Hu, S.; Liu, L. Phosphorylation and acetylation modifications of FOXO3a: Independently or synergistically? Oncol. Lett. 2017, 13, 2867–2872. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cellular Event | Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | R | Q | DOX | DOX + R | DOX + Q | 5-FU | 5-FU + R | 5-FU + Q | |
| Apoptosis | − | + | +++ | +++ | +++ | ++ | ++ | +++ | ++ |
| Autophagy | − | + | +++ | + | + | + | ++ | +++ | +++ |
| Akt activation | − | ++ | ++ | − | + | − | ++ | ++ | +++ |
| ERK activation | − | −− | + | ++ | + | +++ | −− | −− | +++ |
| JNK activation | − | + | + | −− | −− | − | + | 0 | ++ |
| P38 activation | − | 0 | + | + | ++ | ++ | + | +++ | +++ |
| AMPK activation | − | − | + | −−− | −−− | 0 | −−− | −− | 0 |
| Nuclear FOXO3a | − | ++ | ++ | ++ | ++ | +++ | ++ | +++ | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suman, I.; Jezidžić, A.; Dobrić, D.; Domitrović, R. Differential Effects of Rutin and Its Aglycone Quercetin on Cytotoxicity and Chemosensitization of HCT 116 Colon Cancer Cells to Anticancer Drugs 5-Fluorouracil and Doxorubicin. Biology 2025, 14, 527. https://doi.org/10.3390/biology14050527
Suman I, Jezidžić A, Dobrić D, Domitrović R. Differential Effects of Rutin and Its Aglycone Quercetin on Cytotoxicity and Chemosensitization of HCT 116 Colon Cancer Cells to Anticancer Drugs 5-Fluorouracil and Doxorubicin. Biology. 2025; 14(5):527. https://doi.org/10.3390/biology14050527
Chicago/Turabian StyleSuman, Iva, Alberta Jezidžić, Dorotea Dobrić, and Robert Domitrović. 2025. "Differential Effects of Rutin and Its Aglycone Quercetin on Cytotoxicity and Chemosensitization of HCT 116 Colon Cancer Cells to Anticancer Drugs 5-Fluorouracil and Doxorubicin" Biology 14, no. 5: 527. https://doi.org/10.3390/biology14050527
APA StyleSuman, I., Jezidžić, A., Dobrić, D., & Domitrović, R. (2025). Differential Effects of Rutin and Its Aglycone Quercetin on Cytotoxicity and Chemosensitization of HCT 116 Colon Cancer Cells to Anticancer Drugs 5-Fluorouracil and Doxorubicin. Biology, 14(5), 527. https://doi.org/10.3390/biology14050527