Genome-Wide Identification of the Defensin Gene Family in Triticum durum and Assessment of Its Response to Environmental Stresses

 , , , , , , ,
, , , , , , ,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Stress Treatments
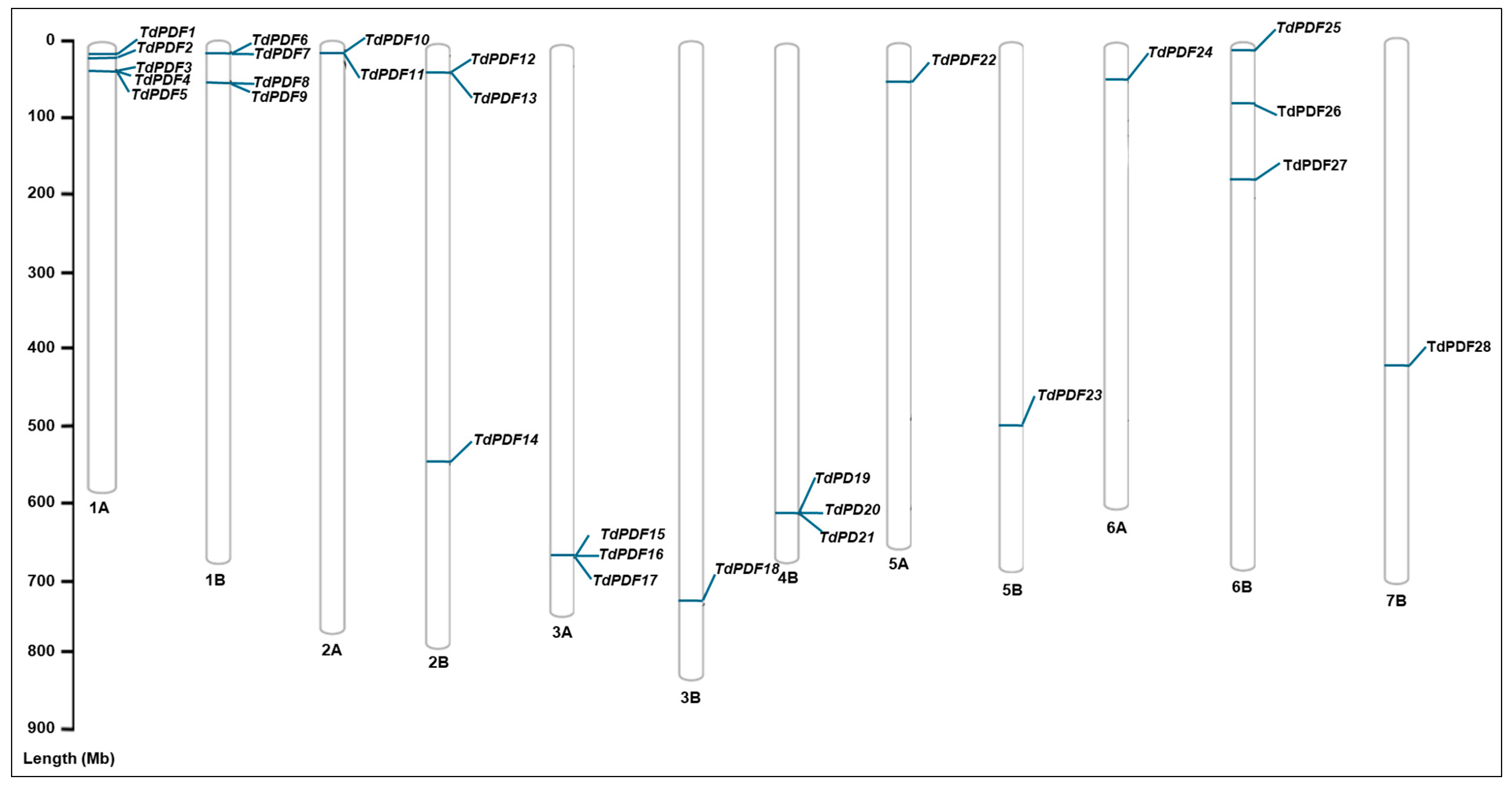
2.2. Identification and Chromosomal Mapping of TdPDF Genes in Durum Wheat
2.3. Characterization of TdPDF Proteins
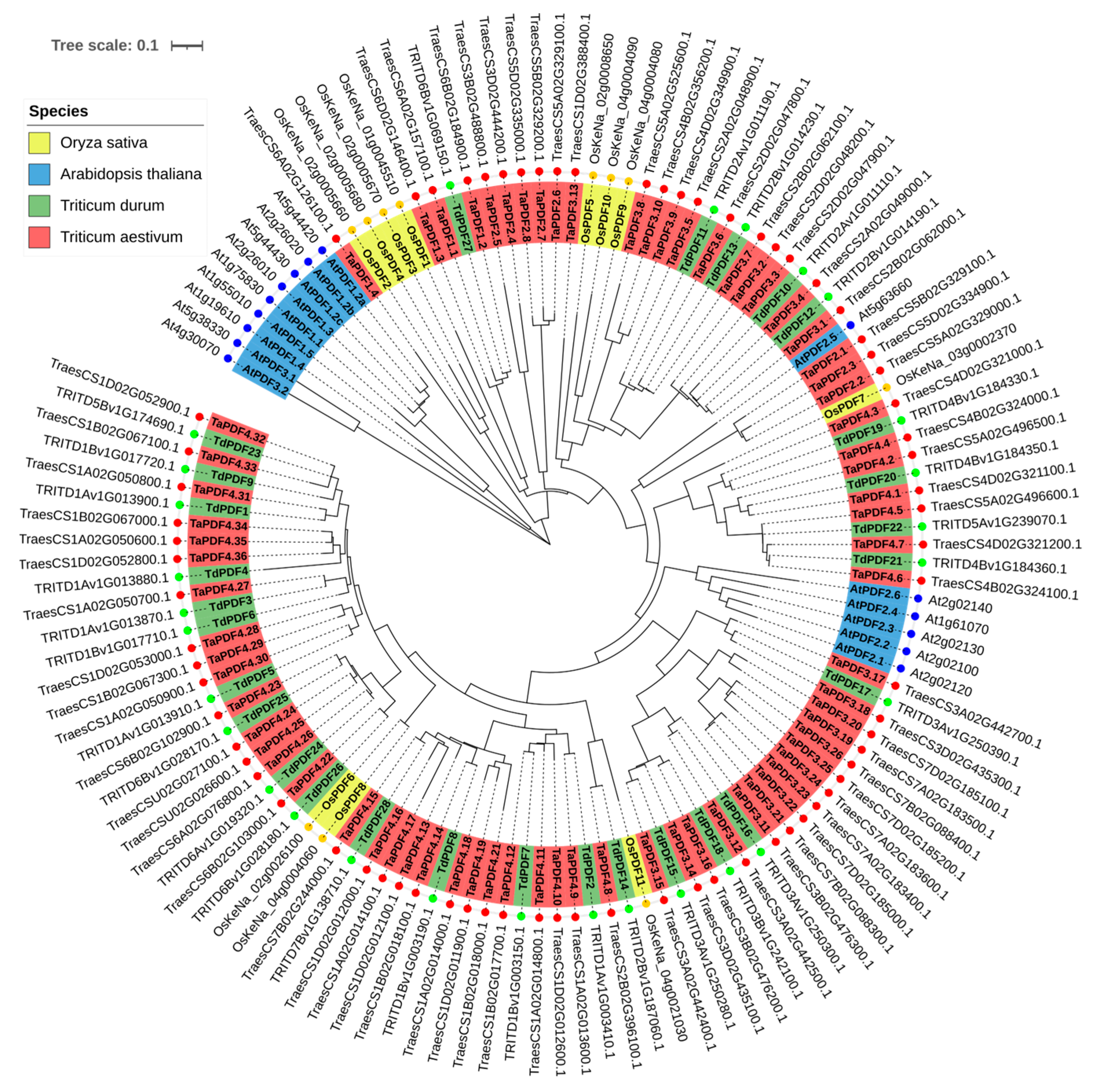
2.4. Phylogenetic Analysis
2.5. Analyses of TdPDF Genes and Promoter Regions
2.6. Quantitative RT PCR Analyses
2.7. Docking Studies and Structural Modeling
2.8. Assessments of Protein-Protein Interaction Network
3. Results
3.1. Screening and Identification of TdPDF Genes in Durum Wheat
3.2. TdPDF Protein Characterization and Subcellular Localization Prediction
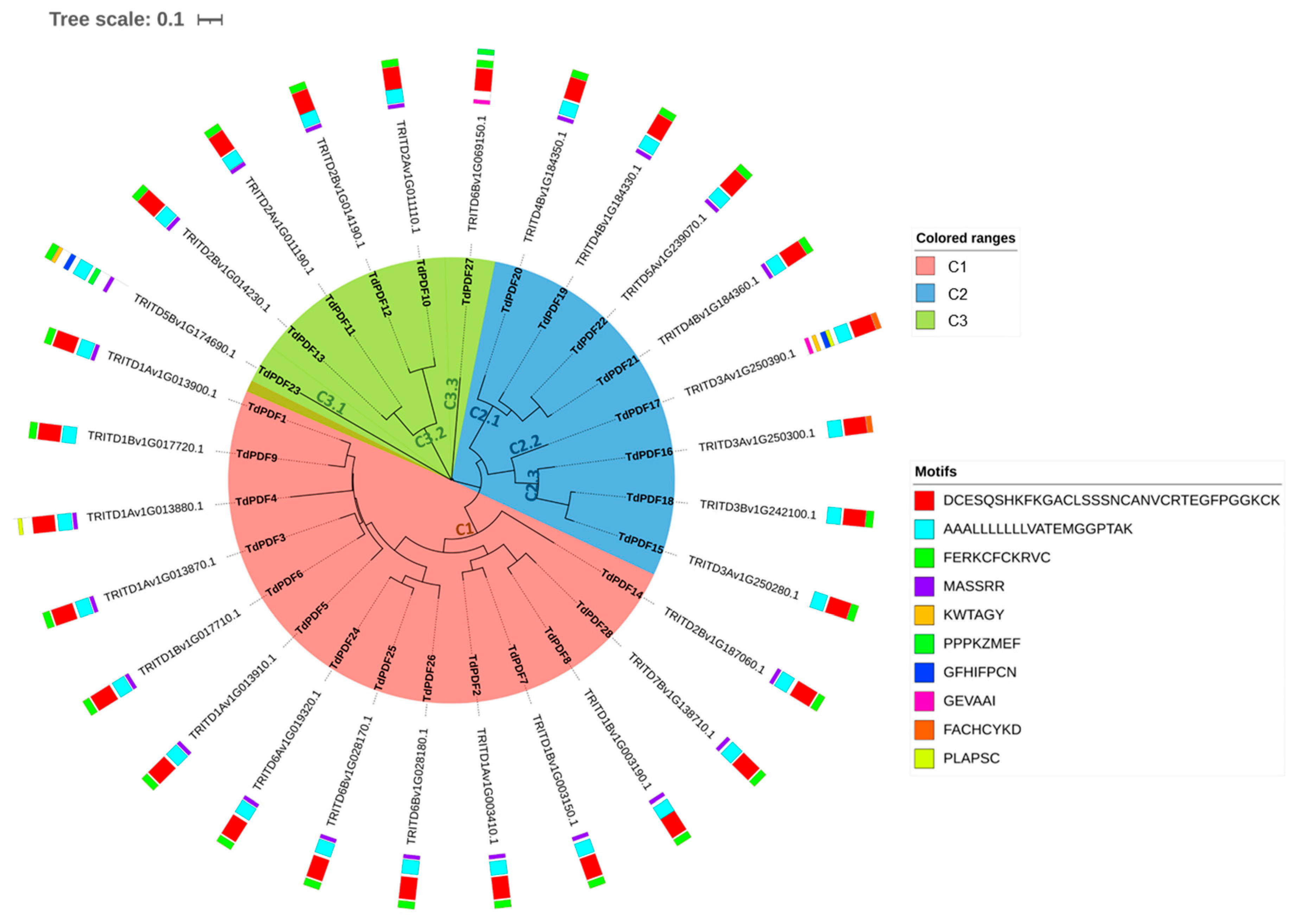
3.3. Conserved Motif and Phylogenetic Analysis of TdPDF Proteins
3.4. TdPDF Gene Duplication and Ka/Ks Analysis
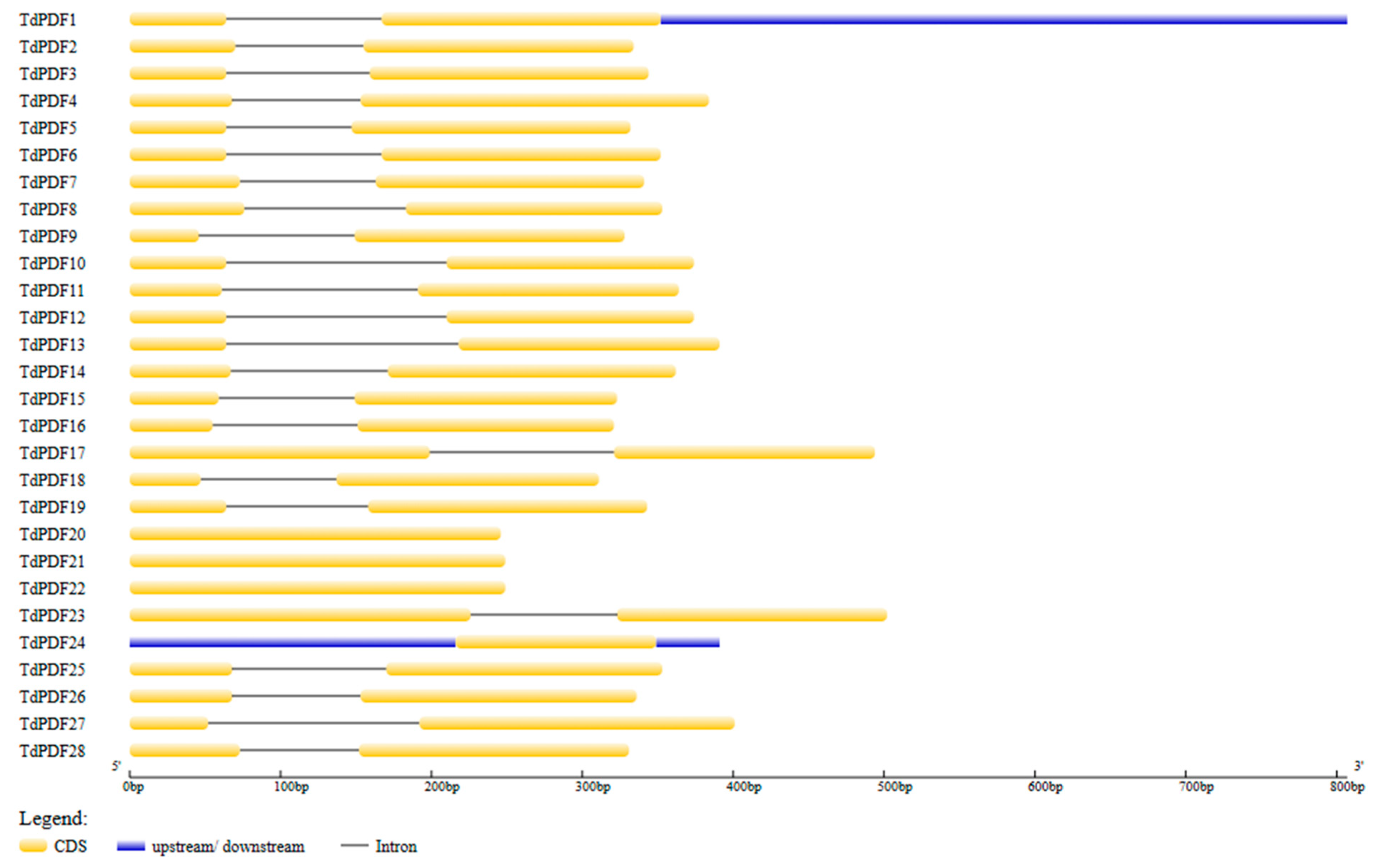
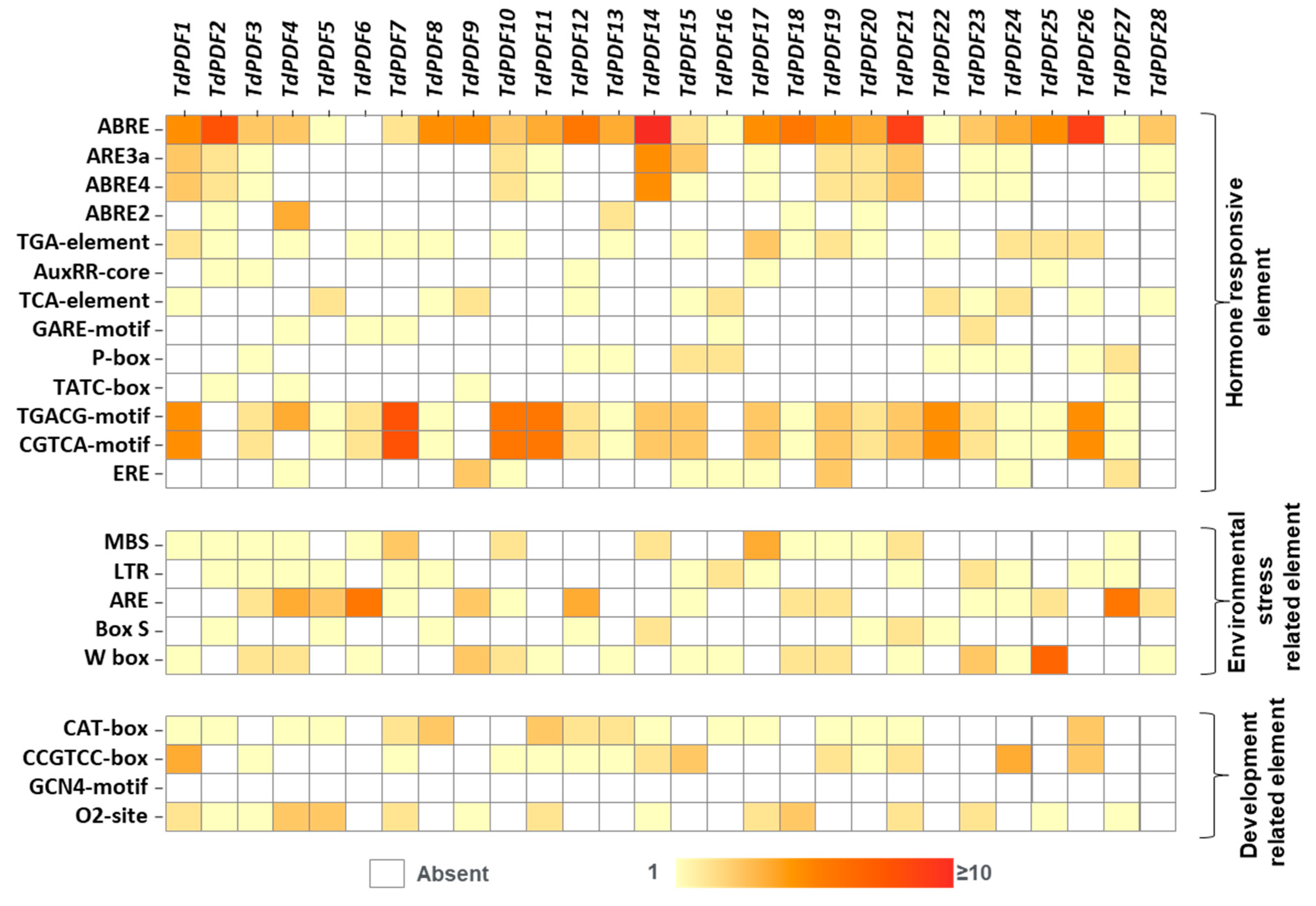
3.5. Gene Structure and Cis-Regulatory Element Analyses
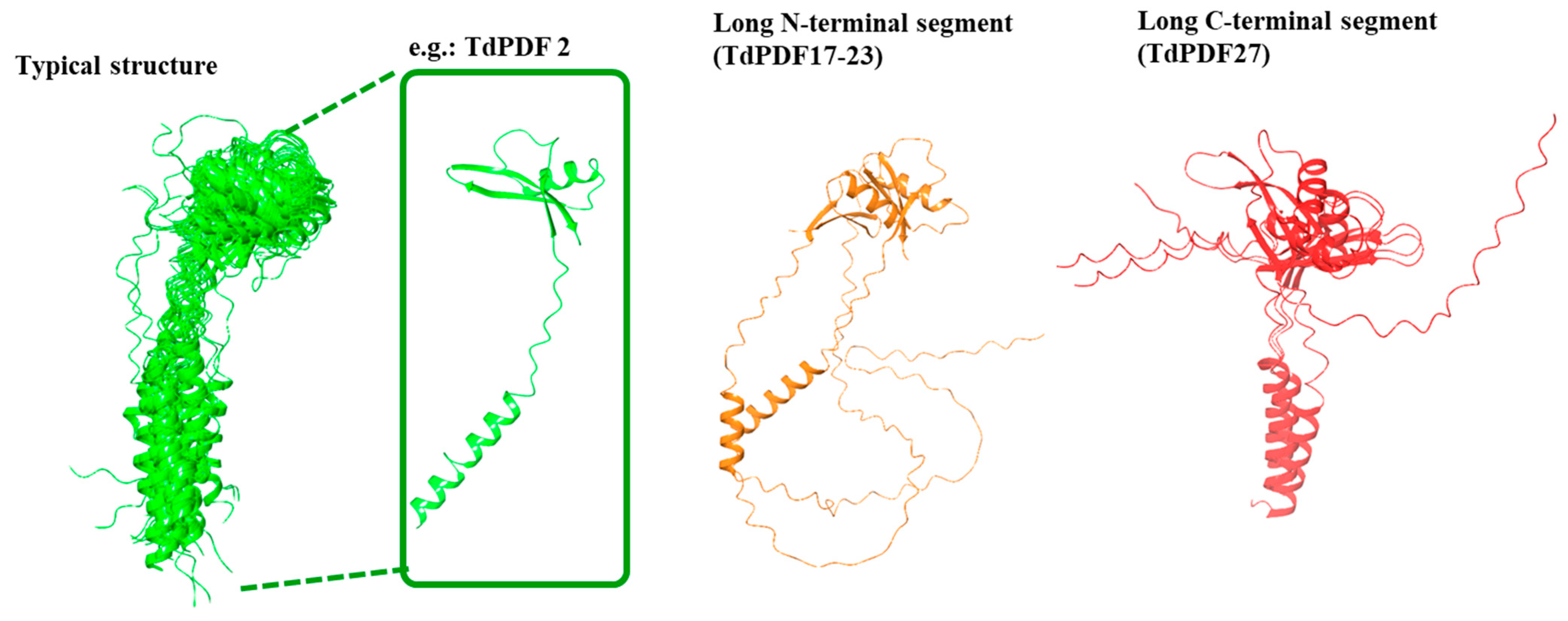
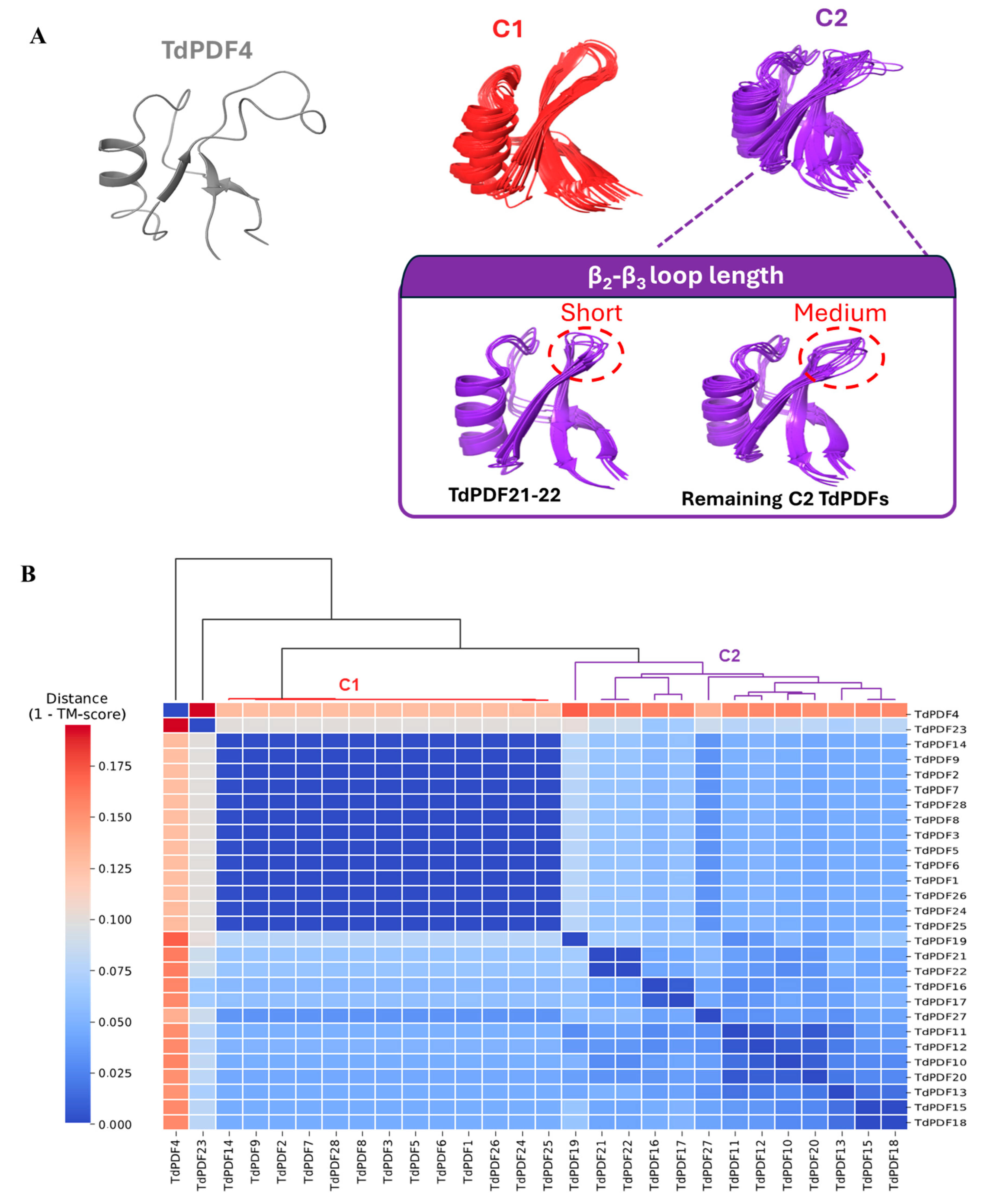
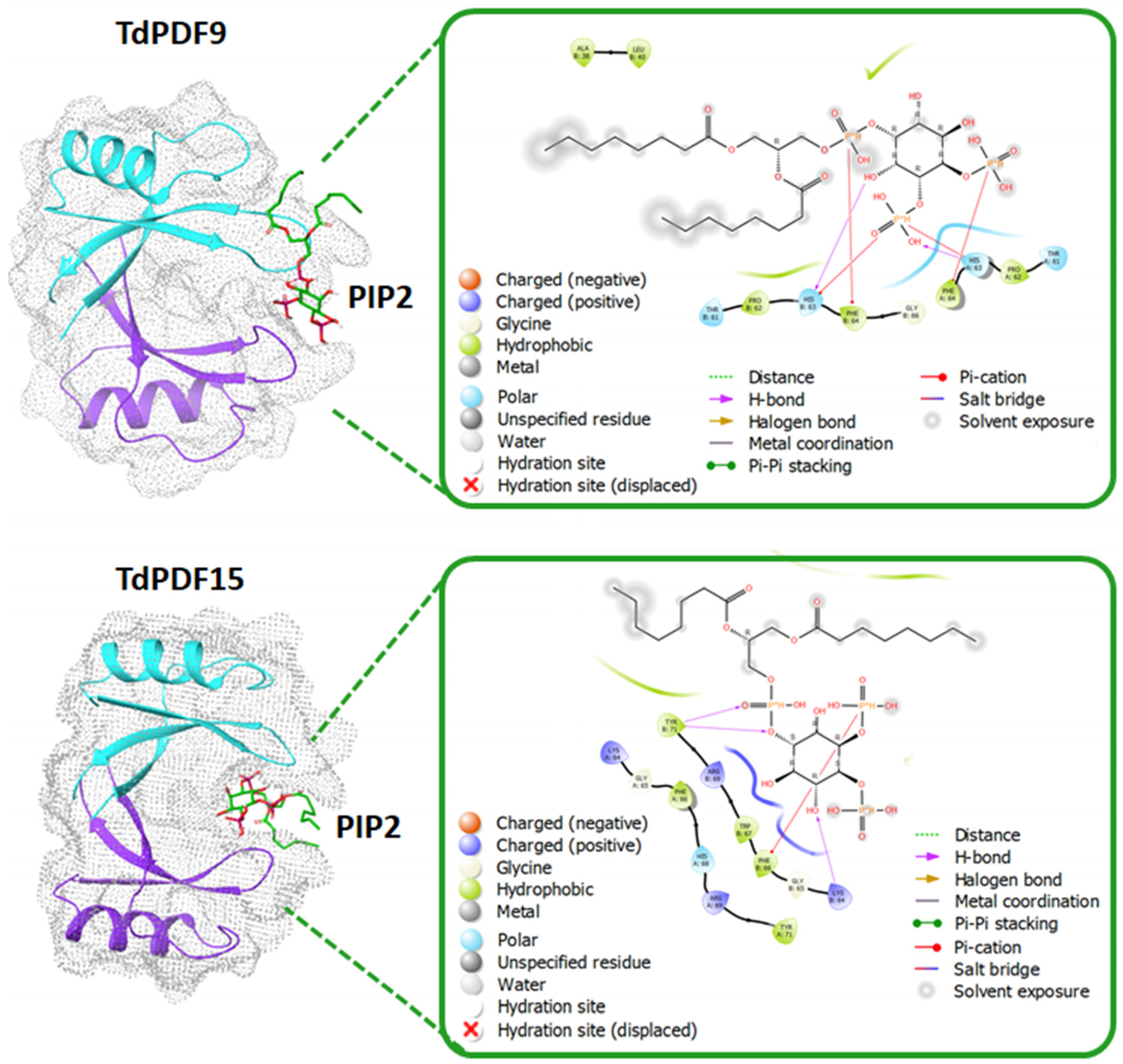
3.6. Docking Analysis and Structural Modeling
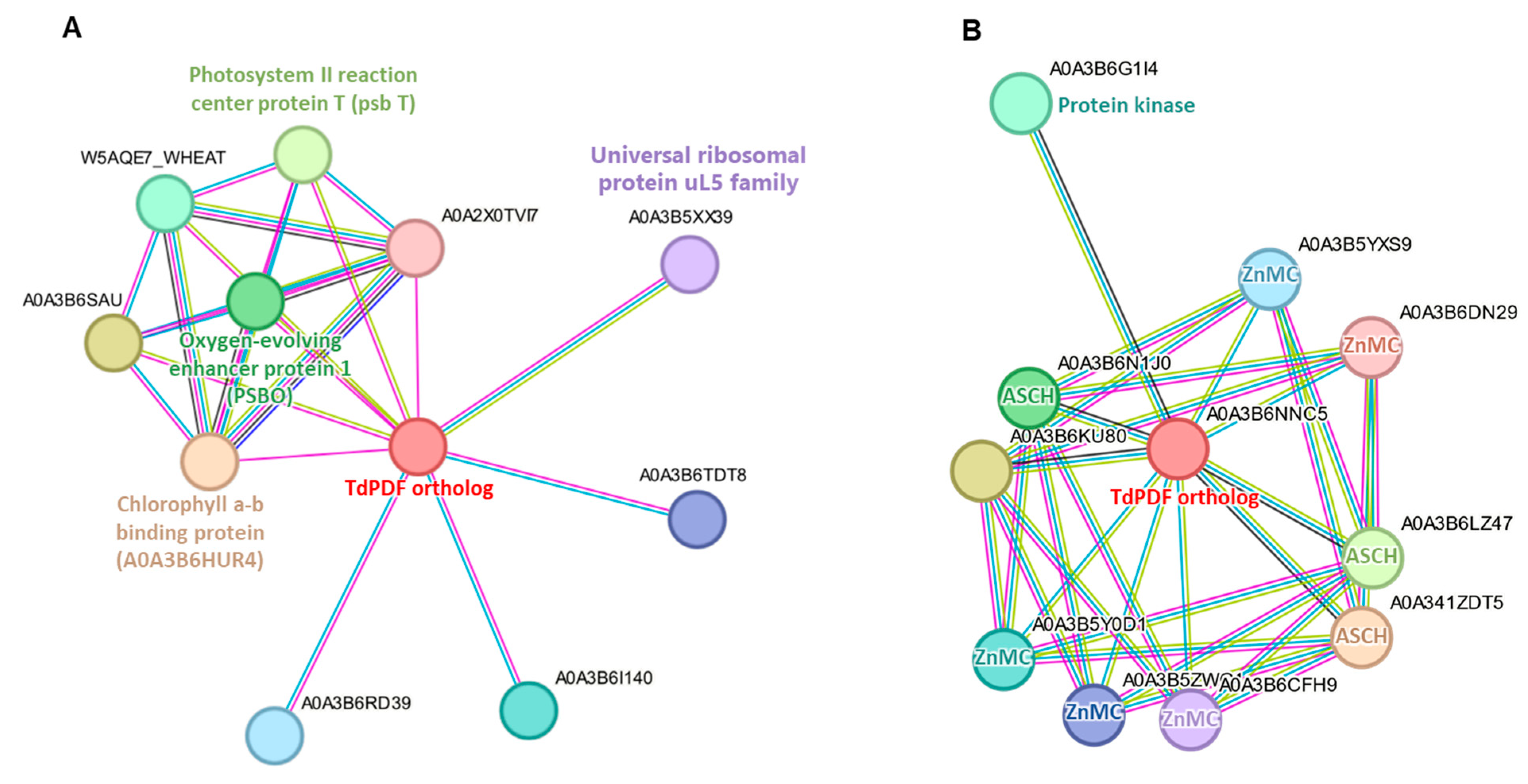
3.7. Protein-Protein Interaction Network Analysis
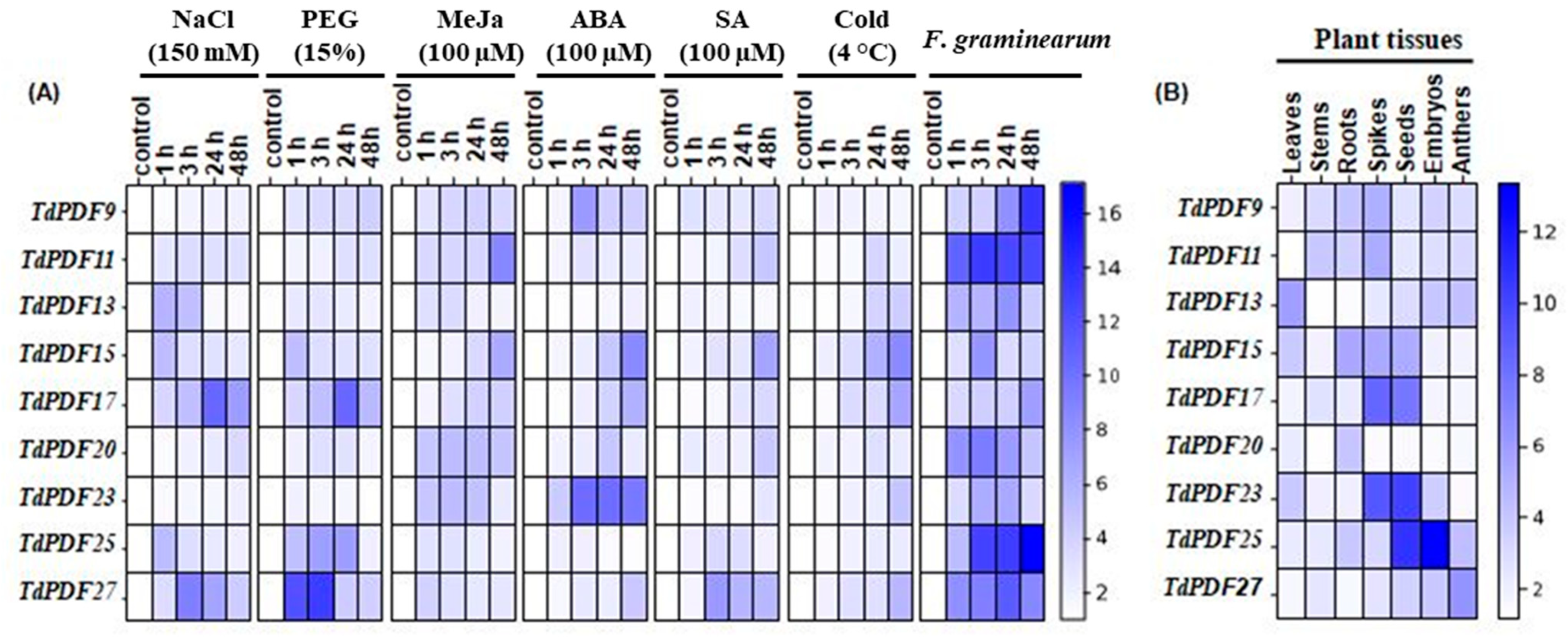
3.8. TdPDF Genes Are Highly Expressed in Developing Seeds and Induced by Multiple Stresses and Phytohormones Treatments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vendramin, V.; Ormanbekova, D.; Scalabrin, S.; Scaglione, D.; Maccaferri, M.; Martelli, P.; Salvi, S.; Jurman, I.; Casadio, R.; Cattonaro, F.; et al. Genomic Tools for Durum Wheat Breeding: De Novo Assembly of Svevo Transcriptome and SNP Discovery in Elite Germplasm. BMC Genom. 2019, 20, 278. [Google Scholar] [CrossRef]
- Marcotuli, I.; Colasuonno, P.; Hsieh, Y.S.Y.; Fincher, G.B.; Gadaleta, A. Non-Starch Polysaccharides in Durum Wheat: A Review. Int. J. Mol. Sci. 2020, 21, 2933. [Google Scholar] [CrossRef]
- Maggio, A.; De Pascale, S.; Fagnano, M.; Barbieri, G. Saline Agriculture in Mediterranean Environments. Ital. J. Agron. 2011, 6, e7. [Google Scholar] [CrossRef]
- Avery, S.V.; Singleton, I.; Magan, N.; Goldman, G.H. The Fungal Threat to Global Food Security. Fungal Biol. 2019, 123, 555–557. [Google Scholar] [CrossRef]
- Ntui, V.O.; Azadi, P.; Thirukkumaran, G.; Khan, R.S.; Chin, D.P.; Nakamura, I.; Mii, M. Increased Resistance to Fusarium Wilt in Transgenic Tobacco Lines Co-Expressing Chitinase and Wasabi Defensin Genes. Plant Pathol. 2011, 60, 221–231. [Google Scholar] [CrossRef]
- Pappas, M.; Baptista, P.; Broufas, G.; Dalakouras, A.; Djobbi, W.; Flors, V.; Msaad Guerfali, M.; Khayi, S.; Mentag, R.; Pastor, V.; et al. Biological and Molecular Control Tools in Plant Defense; Springer: Berlin/Heidelberg, Germany, 2020; pp. 3–43. ISBN 978-3-030-51034-3. [Google Scholar]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by Small-Molecule Hormones in Plant Immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef]
- Wu, J.; Baldwin, I.T. New Insights into Plant Responses to the Attack from Insect Herbivores. Annu. Rev. Genet. 2010, 44, 1–24. [Google Scholar] [CrossRef]
- Thaler, J.S.; Farag, M.A.; Paré, P.W.; Dicke, M. Jasmonate-Deficient Plants Have Reduced Direct and Indirect Defences against Herbivores. Ecol. Lett. 2002, 5, 764–774. [Google Scholar] [CrossRef]
- Bodenhausen, N.; Reymond, P. Signaling Pathways Controlling Induced Resistance to Insect Herbivores in Arabidopsis. Mol. Plant-Microbe Interact. 2007, 20, 1406–1420. [Google Scholar] [CrossRef]
- Nawrot, R.; Barylski, J.; Nowicki, G.; Broniarczyk, J.; Buchwald, W.; Goździcka-Józefiak, A. Plant Antimicrobial Peptides. Folia Microbiol. 2014, 59, 181–196. [Google Scholar] [CrossRef]
- Caldwell, J.E.; Abildgaard, F.; Dzakula, Z.; Ming, D.; Hellekant, G.; Markley, J.L. Solution Structure of the Thermostable Sweet-Tasting Protein Brazzein. Nat. Struct. Biol. 1998, 5, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Gracy, J.; Le-Nguyen, D.; Gelly, J.-C.; Kaas, Q.; Heitz, A.; Chiche, L. KNOTTIN: The Knottin or Inhibitor Cystine Knot Scaffold in 2007. Nucleic Acids Res. 2008, 36, D314–D319. [Google Scholar] [CrossRef]
- Swathi Anuradha, T.; Divya, K.; Jami, S.K.; Kirti, P.B. Transgenic Tobacco and Peanut Plants Expressing a Mustard Defensin Show Resistance to Fungal Pathogens. Plant Cell Rep. 2008, 27, 1777–1786. [Google Scholar] [CrossRef]
- Terras, F.R.; Eggermont, K.; Kovaleva, V.; Raikhel, N.V.; Osborn, R.W.; Kester, A.; Rees, S.B.; Torrekens, S.; Van Leuven, F.; Vanderleyden, J. Small Cysteine-Rich Antifungal Proteins from Radish: Their Role in Host Defense. Plant Cell 1995, 7, 573–588. [Google Scholar]
- Jha, S.; Tank, H.; Prasad, B.; Chattoo, B. Expression of Dm-AMP1 in Rice Confers Resistance to Magnaporthe oryzae and Rhizoctonia Solani. Transgenic Res. 2008, 18, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wang, Y.; Tang, M.; Chen, W.; Chai, Y.; Wang, W. Bioinformatic Analysis of Wheat Defensin Gene Family and Function Verification of Candidate Genes. Front. Plant Sci. 2023, 14, 1279502. [Google Scholar] [CrossRef]
- Franco, O.L. Peptide Promiscuity: An Evolutionary Concept for Plant Defense. FEBS Lett. 2011, 585, 995–1000. [Google Scholar] [CrossRef]
- Stotz, H.U.; Thomson, J.; Wang, Y. Plant Defensins: Defense, Development and Application. Plant Signal. Behav. 2009, 4, 1010–1012. [Google Scholar] [CrossRef]
- van der Weerden, N.; Anderson, M. Plant Defensins: Common Fold, Multiple Functions. Fungal Biol. Rev. 2013, 26, 121–131. [Google Scholar] [CrossRef]
- Khan, R.S.; Darwish, N.A.; Khattak, B.; Ntui, V.O.; Kong, K.; Shimomae, K.; Nakamura, I.; Mii, M. Retransformation of Marker-Free Potato for Enhanced Resistance against Fungal Pathogens by Pyramiding Chitinase and Wasabi Defensin Genes. Mol. Biotechnol. 2014, 56, 814–823. [Google Scholar] [CrossRef]
- Velivelli, S.L.S.; Islam, K.T.; Hobson, E.; Shah, D.M. Modes of Action of a Bi-Domain Plant Defensin MtDef5 Against a Bacterial Pathogen Xanthomonas campestris. Front. Microbiol. 2018, 9, 934. [Google Scholar] [CrossRef] [PubMed]
- Mirouze, M.; Sels, J.; Richard, O.; Czernic, P.; Loubet, S.; Jacquier, A.; François, I.E.J.A.; Cammue, B.P.A.; Lebrun, M.; Berthomieu, P.; et al. A Putative Novel Role for Plant Defensins: A Defensin from the Zinc Hyper-Accumulating Plant, Arabidopsis halleri, Confers Zinc Tolerance. Plant J. 2006, 47, 329–342. [Google Scholar] [CrossRef]
- Lin, K.-F.; Lee, T.-R.; Tsai, P.-H.; Hsu, M.-P.; Chen, C.-S.; Lyu, P.-C. Structure-Based Protein Engineering for Alpha-Amylase Inhibitory Activity of Plant Defensin. Proteins 2007, 68, 530–540. [Google Scholar] [CrossRef]
- Kaewklom, S.; Wongchai, M.; Petvises, S.; Hanpithakphong, W.; Aunpad, R. Structural and Biological Features of a Novel Plant Defensin from Brugmansia x Candida. PLoS ONE 2018, 13, e0201668. [Google Scholar] [CrossRef]
- Lay, F.T.; Anderson, M.A. Defensins--Components of the Innate Immune System in Plants. Curr. Protein Pept. Sci. 2005, 6, 85–101. [Google Scholar] [CrossRef]
- Parisi, K.; Doyle, S.R.; Lee, E.; Lowe, R.G.T.; van der Weerden, N.L.; Anderson, M.A.; Bleackley, M.R. Screening the Saccharomyces cerevisiae Nonessential Gene Deletion Library Reveals Diverse Mechanisms of Action for Antifungal Plant Defensins. Antimicrob. Agents Chemother. 2019, 63, e01097-19. [Google Scholar] [CrossRef] [PubMed]
- Farvardin, A.; González-Hernández, A.I.; Llorens, E.; Camañes, G.; Scalschi, L.; Vicedo, B. The Dual Role of Antimicrobial Proteins and Peptides: Exploring Their Direct Impact and Plant Defense-Enhancing Abilities. Plants 2024, 13, 2059. [Google Scholar] [CrossRef]
- Nitnavare, R.B.; Pothana, A.; Yeshvekar, R.K.; Bhattacharya, J.; Sapara, V.; Reddy, P.S.; Ramtirtha, Y.; Tarafdar, A.; Sharma, M.; Bhatnagar-Mathur, P. Chickpea Defensin Gene Family: Promising Candidates for Resistance Against Soil-Borne Chickpea Fungal Pathogens. J. Plant Growth Regul. 2022, 42, 6244–6260. [Google Scholar] [CrossRef]
- Kerenga, B.K.; McKenna, J.A.; Harvey, P.J.; Quimbar, P.; Garcia-Ceron, D.; Lay, F.T.; Phan, T.K.; Veneer, P.K.; Vasa, S.; Parisi, K.; et al. Salt-Tolerant Antifungal and Antibacterial Activities of the Corn Defensin ZmD32. Front. Microbiol. 2019, 10, 795. [Google Scholar] [CrossRef]
- Kant, P.; Liu, W.-Z.; Pauls, K.P. PDC1, a Corn Defensin Peptide Expressed in Escherichia coli and Pichia pastoris Inhibits Growth of Fusarium Graminearum. Peptides 2009, 30, 1593–1599. [Google Scholar] [CrossRef]
- Kushmerick, C.; de Souza Castro, M.; Santos Cruz, J.; Bloch, C.; Beirão, P.S. Functional and Structural Features of Gamma-Zeathionins, a New Class of Sodium Channel Blockers. FEBS Lett. 1998, 440, 302–306. [Google Scholar] [CrossRef]
- Parashina, E.; Shadenkov, A.; Lavrova, N.; Avetisov, V. The Use of the Defensive Peptide (Defensin) Gene from Radish Seeds in Order to Improve the Resistance of Tomatoes towards Diseases Caused by Fungi. Biotekhnologiya 1999, 15, 35–41. [Google Scholar]
- de Beer, A.; Vivier, M.A. Four Plant Defensins from an Indigenous South African Brassicaceae Species Display Divergent Activities against Two Test Pathogens despite High Sequence Similarity in the Encoding Genes. BMC Res. Notes 2011, 4, 459. [Google Scholar] [CrossRef]
- Lin, P.; Wong, J.H.; Ng, T.B. A Defensin with Highly Potent Antipathogenic Activities from the Seeds of Purple Pole Bean. Biosci. Rep. 2009, 30, 101–109. [Google Scholar] [CrossRef]
- Bouteraa, M.T.; Garzoli, S.; Romdhane, W.B.; Baazaoui, N.; Chouaibi, Y.; Hsouna, A.B.; Saad, R.B. Foliar Application of Lobularia Maritima-Derived Polysaccharides Modulates Chemical Composition and Enhances Salt Tolerance in Greenhouse-Cultivated Durum Wheat. J. Soil. Sci. Plant Nutr. 2025, 25, 589–602. [Google Scholar] [CrossRef]
- Ben Romdhane, W.; Ben Saad, R.; Meynard, D.; Zouari, N.; Tarroum, M.; Ali, A.; Droc, G.; Périn, C.; Morel, J.-B.; Fki, L.; et al. Expression of an A20/AN1 Stress-Associated Protein from Aeluropus littoralis in Rice Deregulates Stress-Related Genes. J. Plant Growth Regul. 2022, 41, 848–862. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Springer Protocols Handbooks; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. ISBN 978-1-59259-890-8. [Google Scholar]
- Savojardo, C.; Martelli, P.L.; Fariselli, P.; Profiti, G.; Casadio, R. BUSCA: An Integrative Web Server to Predict Subcellular Localization of Proteins. Nucleic Acids Res. 2018, 46, W459–W466. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v6: Recent Updates to the Phylogenetic Tree Display and Annotation Tool. Nucleic Acids Res. 2024, 52, W78–W82. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Ben Saad, R.; Safi, H.; Ben Hsouna, A.; Brini, F.; Ben Romdhane, W. Functional Domain Analysis of LmSAP Protein Reveals the Crucial Role of the Zinc-Finger A20 Domain in Abiotic Stress Tolerance. Protoplasma 2019, 256, 1333–1344. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Giménez, M.J.; Pistón, F.; Atienza, S.G. Identification of Suitable Reference Genes for Normalization of qPCR Data in Comparative Transcriptomics Analyses in the Triticeae. Planta 2011, 233, 163–173. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology Modelling of Protein Structures and Complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Zhang, Y.; Skolnick, J. TM-Align: A Protein Structure Alignment Algorithm Based on the TM-Score. Nucleic Acids Res. 2005, 33, 2302–2309. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization and Multithreading. J. Comput. Chem. 2010, 31, 455. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING Database in 2023: Protein-Protein Association Networks and Functional Enrichment Analyses for Any Sequenced Genome of Interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The Roles of Segmental and Tandem Gene Duplication in the Evolution of Large Gene Families in Arabidopsis Thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Lay, F.T.; Brugliera, F.; Anderson, M.A. Isolation and Properties of Floral Defensins from Ornamental Tobacco and Petunia. Plant Physiol. 2003, 131, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Oomen, R.J.F.J.; Séveno-Carpentier, E.; Ricodeau, N.; Bournaud, C.; Conéjéro, G.; Paris, N.; Berthomieu, P.; Marquès, L. Plant Defensin AhPDF1.1 Is Not Secreted in Leaves but It Accumulates in Intracellular Compartments. New Phytol. 2011, 192, 140–150. [Google Scholar] [CrossRef]
- Cools, T.L.; Struyfs, C.; Cammue, B.P.; Thevissen, K. Antifungal Plant Defensins: Increased Insight in Their Mode of Action as a Basis for Their Use to Combat Fungal Infections. Future Microbiol. 2017, 12, 441–454. [Google Scholar] [CrossRef]
- Weerawanich, K.; Webster, G.; Ma, J.K.-C.; Phoolcharoen, W.; Sirikantaramas, S. Gene Expression Analysis, Subcellular Localization, and in Planta Antimicrobial Activity of Rice (Oryza sativa L.) Defensin 7 and 8. Plant Physiol. Biochem. 2018, 124, 160–166. [Google Scholar] [CrossRef]
- Xu, M.; Wang, Q.; Wang, G.; Zhang, X.; Liu, H.; Jiang, C. Combatting Fusarium Head Blight: Advances in Molecular Interactions between Fusarium graminearum and Wheat. Phytopathol. Res. 2022, 4, 37. [Google Scholar] [CrossRef]
- Odintsova, T.I.; Slezina, M.P.; Istomina, E.A. Defensins of Grasses: A Systematic Review. Biomolecules 2020, 10, 1029. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, M.; Zhang, Z.; Ren, L.; Du, L.; Zhang, B.; Xu, H.; Xin, Z. Expression of a Radish Defensin in Transgenic Wheat Confers Increased Resistance to Fusarium graminearum and Rhizoctonia cerealis. Funct. Integr. Genom. 2011, 11, 63–70. [Google Scholar] [CrossRef]
- Odintsova, T.I.; Rogozhin, E.A.; Baranov, Y.; Musolyamov, A.K.; Yalpani, N.; Egorov, T.A.; Grishin, E.V. Seed Defensins of Barnyard Grass Echinochloa crusgalli (L.) Beauv. Biochimie 2008, 90, 1667–1673. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Jiang, N.; Meng, J.; Hou, X.; Yang, G.; Luan, Y. Identification and Characterization of Defensin Genes Conferring Phytophthora infestans Resistance in Tomato. Physiol. Mol. Plant Pathol. 2018, 103, 28–35. [Google Scholar] [CrossRef]
- Graham, M.; Silverstein, K.; Vandenbosch, K. Defensin-like Genes: Genomic Perspectives on a Diverse Superfamily in Plants. Crop Sci. 2008, 48, S-3–S-11. [Google Scholar] [CrossRef]
- Nanni, V.; Schumacher, J.; Giacomelli, L.; Brazzale, D.; Sbolci, L.; Moser, C.; Tudzynski, P.; Baraldi, E. VvAMP2, a Grapevine Flower-Specific Defensin Capable of Inhibiting Otrytis Cinerea Growth: Insights into Its Mode of Action. Plant Pathol. 2014, 63, 899–910. [Google Scholar] [CrossRef]
- De-Paula, V.S.; Razzera, G.; Medeiros, L.; Miyamoto, C.A.; Almeida, M.S.; Kurtenbach, E.; Almeida, F.C.L.; Valente, A.P. Evolutionary Relationship between Defensins in the Poaceae Family Strengthened by the Characterization of New Sugarcane Defensins. Plant Mol. Biol. 2008, 68, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Mendez, E.; Moreno, A.; Colilla, F.; Pelaez, F.; Limas, G.G.; Mendez, R.; Soriano, F.; Salinas, M.; de Haro, C. Primary Structure and Inhibition of Protein Synthesis in Eukaryotic Cell-Free System of a Novel Thionin, Gamma-Hordothionin, from Barley Endosperm. Eur. J. Biochem. 1990, 194, 533–539. [Google Scholar] [CrossRef]
- Liu, Y.; Hua, Y.-P.; Chen, H.; Zhou, T.; Yue, C.-P.; Huang, J.-Y. Genome-Scale Identification of Plant Defensin (PDF) Family Genes and Molecular Characterization of Their Responses to Diverse Nutrient Stresses in Allotetraploid Rapeseed. PeerJ 2021, 9, e12007. [Google Scholar] [CrossRef]
- Giacomelli, L.; Nanni, V.; Lenzi, L.; Zhuang, J.; Dalla Serra, M.; Banfield, M.J.; Town, C.D.; Silverstein, K.A.T.; Baraldi, E.; Moser, C. Identification and Characterization of the Defensin-like Gene Family of Grapevine. Mol. Plant Microbe Interact. 2012, 25, 1118–1131. [Google Scholar] [CrossRef]
- Padovan, L.; Segat, L.; Tossi, A.; Calsa, T.; Ederson, A.K.; Brandao, L.; Guimarães, R.L.; Pandolfi, V.; Pestana-Calsa, M.C.; Belarmino, L.C.; et al. Characterization of a New Defensin from Cowpea (Vigna unguiculata (L.) Walp.). Protein Pept. Lett. 2010, 17, 297–304. [Google Scholar] [CrossRef]
- Clavijo, B.J.; Venturini, L.; Schudoma, C.; Accinelli, G.G.; Kaithakottil, G.; Wright, J.; Borrill, P.; Kettleborough, G.; Heavens, D.; Chapman, H.; et al. An Improved Assembly and Annotation of the Allohexaploid Wheat Genome Identifies Complete Families of Agronomic Genes and Provides Genomic Evidence for Chromosomal Translocations. Genome Res. 2017, 27, 885–896. [Google Scholar] [CrossRef]
- Paterson, A.H.; Freeling, M.; Tang, H.; Wang, X. Insights from the Comparison of Plant Genome Sequences. Annu. Rev. Plant Biol. 2010, 61, 349–372. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Park, C. Adaptive evolution of β-defensin gene clusters by gene duplication and loss during environmental changes in mammals. J. Anim. Breed. Genom. 2022, 6, 143–154. [Google Scholar]
- Du, L.; Ma, Z.; Mao, H. Duplicate Genes Contribute to Variability in Abiotic Stress Resistance in Allopolyploid Wheat. Plants 2023, 12, 2465. [Google Scholar] [CrossRef]
- Kondrashov, F.A. Gene Duplication as a Mechanism of Genomic Adaptation to a Changing Environment. Proc. Biol. Sci. 2012, 279, 5048–5057. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.S.M.; Pires, Á.S.; Damaceno, N.B.; Rigueiras, P.O.; Maximiano, M.R.; Franco, O.L.; Porto, W.F. In Silico Characterization of Class II Plant Defensins from Arabidopsis thaliana. Phytochemistry 2020, 179, 112511. [Google Scholar] [CrossRef]
- Zhao, K.; Ren, R.; Ma, X.; Zhao, K.; Qu, C.; Cao, D.; Ma, Q.; Ma, Y.; Gong, F.; Li, Z.; et al. Genome-Wide Investigation of Defensin Genes in Peanut (Arachis hypogaea L.) Reveals AhDef2.2 Conferring Resistance to Bacterial Wilt. Crop J. 2022, 10, 809–819. [Google Scholar] [CrossRef]
- Mir Drikvand, R.; Sohrabi, S.M.; Samiei, K. Molecular Cloning and Characterization of Six Defensin Genes from Lentil Plant (Lens culinaris L.). 3 Biotech. 2019, 9, 104. [Google Scholar] [CrossRef]
- Houlné, G.; Meyer, B.; Schantz, R. Alteration of the Expression of a Plant Defensin Gene by Exon Shuffling in Bell Pepper (Capsicum annuum L.). Mol. Gen. Genet. 1998, 259, 504–510. [Google Scholar] [CrossRef]
- Spelbrink, R.G.; Dilmac, N.; Allen, A.; Smith, T.J.; Shah, D.M.; Hockerman, G.H. Differential Antifungal and Calcium Channel-Blocking Activity among Structurally Related Plant Defensins. Plant Physiol. 2004, 135, 2055–2067. [Google Scholar] [CrossRef]
- Hanks, J.N.; Snyder, A.K.; Graham, M.A.; Shah, R.K.; Blaylock, L.A.; Harrison, M.J.; Shah, D.M. Defensin Gene Family in Medicago Truncatula: Structure, Expression and Induction by Signal Molecules. Plant Mol. Biol. 2005, 58, 385–399. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J.; Cammue, B.P.A.; Thevissen, K. Plant Defensins. Planta 2002, 216, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, K.A.T.; Graham, M.A.; Paape, T.D.; VandenBosch, K.A. Genome Organization of More than 300 Defensin-like Genes in Arabidopsis. Plant Physiol. 2005, 138, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Ben Saad, R.; Ben Romdhane, W.; Zouari, N.; Ben Hsouna, A.; Harbaoui, M.; Brini, F.; Ghneim-Herrera, T. Characterization of a Novel LmSAP Gene Promoter from Lobularia maritima: Tissue Specificity and Environmental Stress Responsiveness. PLoS ONE 2020, 15, e0236943. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Yusuf, M.A.; Yadav, P.; Narayan, S.; Kumar, M. Overexpression of Chickpea Defensin Gene Confers Tolerance to Water-Deficit Stress in Arabidopsis thaliana. Front. Plant Sci. 2019, 10, 290. [Google Scholar] [CrossRef]
- Wei, H.; Movahedi, A.; Xu, C.; Sun, W.; Wang, X.; Li, D.; Zhuge, Q. Overexpression of PtDefensin Enhances Resistance to Septotis populiperda in Transgenic Poplar. Plant Sci. 2020, 292, 110379. [Google Scholar] [CrossRef]
- Gao, L.; Kantar, M.B.; Moxley, D.; Ortiz-Barrientos, D.; Rieseberg, L.H. Crop Adaptation to Climate Change: An Evolutionary Perspective. Mol. Plant 2023, 16, 1518–1546. [Google Scholar] [CrossRef]
- Sathoff, A.E.; Velivelli, S.; Shah, D.M.; Samac, D.A. Plant Defensin Peptides Have Antifungal and Antibacterial Activity Against Human and Plant Pathogens. Phytopathology 2019, 109, 402–408. [Google Scholar] [CrossRef]
- Poon, I.K.; Baxter, A.A.; Lay, F.T.; Mills, G.D.; Adda, C.G.; Payne, J.A.; Phan, T.K.; Ryan, G.F.; White, J.A.; Veneer, P.K.; et al. Phosphoinositide-Mediated Oligomerization of a Defensin Induces Cell Lysis. Elife 2014, 3, e01808. [Google Scholar] [CrossRef]
- Vriens, K.; Cammue, B.P.A.; Thevissen, K. Antifungal Plant Defensins: Mechanisms of Action and Production. Molecules 2014, 19, 12280–12303. [Google Scholar] [CrossRef]
- Oeemig, J.S.; Lynggaard, C.; Knudsen, D.H.; Hansen, F.T.; Nørgaard, K.D.; Schneider, T.; Vad, B.S.; Sandvang, D.H.; Nielsen, L.A.; Neve, S.; et al. Eurocin, a New Fungal Defensin: Structure, Lipid Binding, and Its Mode of Action. J. Biol. Chem. 2012, 287, 42361–42372. [Google Scholar] [CrossRef]
- Tanvir, A. Antimicrobial Peptides and Their Interaction Cell Membrane and Microbial Surface Interactions. J. Med. Org. Chem. 2022, 5, 86–92. [Google Scholar]
- Bakhat, N.; Vielba-Fernández, A.; Padilla-Roji, I.; Martínez-Cruz, J.; Polonio, Á.; Fernández-Ortuño, D.; Pérez-García, A. Suppression of Chitin-Triggered Immunity by Plant Fungal Pathogens: A Case Study of the Cucurbit Powdery Mildew Fungus Podosphaera xanthii. J. Fungi 2023, 9, 771. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, T.I.; Slezina, M.P.; Istomina, E.A. Plant thionins: Structure, biological functions and potential use in biotechnology. Vavilov J. Genet. Breed. 2018, 22, 667–675. [Google Scholar] [CrossRef]
- Wei, H.; Movahedi, A.; Xu, C.; Sun, W.; Wang, P.; Li, D.; Yin, T.; Zhuge, Q. Characterization, Expression Profiling, and Functional Analysis of PtDef, a Defensin-Encoding Gene from Populus trichocarpa. Front. Microbiol. 2020, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.P.H.J.; Broekaert, W.F. Tissue-Specific Expression of Plant Defensin Genes PDF2.1 and PDF2.2 in Arabidopsis thaliana. Plant Physiol. Biochem. 1998, 36, 533–537. [Google Scholar] [CrossRef]
- Carvalho, A.d.O.; Gomes, V.M. Plant Defensins--Prospects for the Biological Functions and Biotechnological Properties. Peptides 2009, 30, 1007–1020. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, L.; Yang, C.-L.; Han, S.; Yang, S.; Liu, G.; Zeng, Q.; Liu, Y. Molecular Identification and Antifungal Activity of a Defensin (PaDef) from Spruce. J. Plant Growth Regul. 2021, 41, 494–506. [Google Scholar] [CrossRef]
- Domingo, G.; Locato, V.; Cimini, S.; Ciceri, L.; Marsoni, M.; De Gara, L.; Bracale, M.; Vannini, C. A Comprehensive Characterization and Expression Profiling of Defensin Family Peptides in Arabidopsis thaliana with a Focus on Their Abiotic Stress-Specific Transcriptional Modulation. Curr. Plant Biol. 2024, 39, 100376. [Google Scholar] [CrossRef]
- Osborn, R.W.; De Samblanx, G.W.; Thevissen, K.; Goderis, I.; Torrekens, S.; Van Leuven, F.; Attenborough, S.; Rees, S.B.; Broekaert, W.F. Isolation and Characterisation of Plant Defensins from Seeds of Asteraceae, Fabaceae, Hippocastanaceae and Saxifragaceae. FEBS Lett. 1995, 368, 257–262. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Modulation of Antioxidant Defense System Is Associated with Combined Drought and Heat Stress Tolerance in Citrus. Front. Plant Sci. 2017, 8, 953. [Google Scholar] [CrossRef]
- Kulaeva, O.; Kliukova, M.; Afonin, A.; Sulima, A.; Zhukov, V.; Tikhonovich, I. The Role of Plant Antimicrobial Peptides (AMPs) in Response to Biotic and Abiotic Environmental Factors. Biol. Commun. 2020, 65, 187–199. [Google Scholar] [CrossRef]
- Shahzad, Z.; Ranwez, V.; Fizames, C.; Marquès, L.; Le Martret, B.; Alassimone, J.; Godé, C.; Lacombe, E.; Castillo, T.; Saumitou-Laprade, P.; et al. Plant Defensin Type 1 (PDF1): Protein Promiscuity and Expression Variation within the Arabidopsis Genus Shed Light on Zinc Tolerance Acquisition in Arabidopsis Halleri. New Phytol. 2013, 200, 820–833. [Google Scholar] [CrossRef] [PubMed]
- Kovaleva, V. Recombinant Expression, Affinity Purification and Functional Characterization of Scots Pine Defensin 1. Appl. Microbiol. Biotechnol. 2011, 89, 1093–1101. [Google Scholar] [CrossRef]
- Thevissen, K.; de Mello Tavares, P.; Xu, D.; Blankenship, J.; Vandenbosch, D.; Idkowiak-Baldys, J.; Govaert, G.; Bink, A.; Rozental, S.; de Groot, P.W.J.; et al. The Plant Defensin RsAFP2 Induces Cell Wall Stress, Septin Mislocalization and Accumulation of Ceramides in Candida albicans. Mol. Microbiol. 2012, 84, 166–180. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chr | Start Site | End Site | Length (bp) | CDS | Exon Number |
|---|---|---|---|---|---|---|---|
| TdPDF1 | TRITD1Av1G013900.1 | 1A | 29,234,920 | 29,235,726 | 352 | 249 | 2 |
| TdPDF2 | TRITD1Av1G003410.1 | 1A | 7,557,945 | 7,558,278 | 334 | 249 | 2 |
| TdPDF3 | TRITD1Av1G013870.1 | 1A | 29,069,143 | 29,069,486 | 344 | 249 | 2 |
| TdPDF4 | TRITD1Av1G013880.1 | 1A | 29,172,467 | 29,172,850 | 384 | 297 | 2 |
| TdPDF5 | TRITD1Av1G013910.1 | 1A | 29,237,984 | 29,238,315 | 332 | 249 | 2 |
| TdPDF6 | TRITD1Bv1G017710.1 | 1B | 45,983,093 | 45,983,444 | 352 | 249 | 2 |
| TdPDF7 | TRITD1Bv1G003150.1 | 1B | 7,441,922 | 7,442,263 | 342 | 252 | 2 |
| TdPDF8 | TRITD1Bv1G003190.1 | 1B | 7,501,776 | 7,502,128 | 353 | 246 | 2 |
| TdPDF9 | TRITD1Bv1G017720.1 | 1B | 46,017,987 | 46,018,314 | 328 | 225 | 2 |
| TdPDF10 | TRITD2Av1G011110.1 | 2A | 19,957,063 | 19,957,436 | 374 | 228 | 2 |
| TdPDF11 | TRITD2Av1G011190.1 | 2A | 20,339,771 | 20,340,134 | 364 | 234 | 2 |
| TdPDF12 | TRITD2Bv1G014190.1 | 2B | 29,124,967 | 29,125,340 | 374 | 228 | 2 |
| TdPDF13 | TRITD2Bv1G014230.1 | 2B | 29,274,981 | 29,275,371 | 391 | 237 | 2 |
| TdPDF14 | TRITD2Bv1G187060.1 | 2B | 553,467,008 | 553,467,369 | 362 | 258 | 2 |
| TdPDF15 | TRITD3Av1G250280.1 | 3A | 675,334,189 | 675,334,511 | 323 | 231 | 2 |
| TdPDF16 | TRITD3Av1G250300.1 | 3A | 675,335,903 | 675,336,223 | 321 | 225 | 2 |
| TdPDF17 | TRITD3Av1G250390.1 | 3A | 675,594,401 | 675,594,894 | 494 | 372 | 2 |
| TdPDF18 | TRITD3Bv1G242100.1 | 3B | 736,591,016 | 736,591,326 | 311 | 219 | 2 |
| TdPDF19 | TRITD4Bv1G184330.1 | 4B | 618,745,628 | 618,745,970 | 343 | 249 | 2 |
| TdPDF20 | TRITD4Bv1G184350.1 | 4B | 618,748,241 | 618,748,486 | 246 | 246 | 1 |
| TdPDF21 | TRITD4Bv1G184360.1 | 4B | 618,758,106 | 618,758,354 | 249 | 249 | 1 |
| TdPDF22 | TRITD5Av1G239070.1 | 5A | 622,552,360 | 622,552,608 | 249 | 249 | 1 |
| TdPDF23 | TRITD5Bv1G174690.1 | 5B | 511,145,927 | 511,146,428 | 502 | 405 | 2 |
| TdPDF24 | TRITD6Av1G019320.1 | 6A | 45,820,912 | 45,821,245 | 334 | 249 | 2 |
| TdPDF25 | TRITD6Bv1G028170.1 | 6B | 77,285,794 | 77,286,146 | 353 | 249 | 2 |
| TdPDF26 | TRITD6Bv1G028180.1 | 6B | 77,294,347 | 77,294,682 | 336 | 249 | 2 |
| TdPDF27 | TRITD6Bv1G069150.1 | 6B | 202,420,199 | 202,420,599 | 401 | 261 | 2 |
| TdPDF28 | TRITD7Bv1G138710.1 | 7B | 436,196,050 | 436,196,380 | 331 | 252 | 2 |
| Gene Name | Gene ID | Protein Length | Pl | MW (KDa) | Ii | GRAVY | Subcellular Localization | Signal Peptide Position |
|---|---|---|---|---|---|---|---|---|
| TdPDF1 | TRITD1Av1G013900.1 | 82 | 8.92 | 8.945 | 58.10 | 0.029 | Extracellular | 1–33 |
| TdPDF2 | TRITD1Av1G003410.1 | 82 | 8.90 | 8.972 | 46.25 | −0.017 | Extracellular | 1–33 |
| TdPDF3 | TRITD1Av1G013870.1 | 82 | 8.50 | 8.823 | 66.51 | 0.054 | Extracellular | 1–33 |
| TdPDF4 | TRITD1Av1G013880.1 | 98 | 9.79 | 10.612 | 72.63 | 0.265 | Extracellular | 1–33 |
| TdPDF5 | TRITD1Av1G013910.1 | 82 | 8.92 | 8.837 | 60.72 | 0.178 | Extracellular | 1–33 |
| TdPDF6 | TRITD1Bv1G017710.1 | 82 | 8.72 | 8.973 | 55.86 | −0.060 | Extracellular | 1–33 |
| TdPDF7 | TRITD1Bv1G003150.1 | 83 | 9.10 | 9.080 | 51.22 | −0.060 | Extracellular | 1–34 |
| TdPDF8 | TRITD1Bv1G003190.1 | 81 | 8.50 | 8.977 | 58.07 | −0.265 | Extracellular | 1–32 |
| TdPDF9 | TRITD1Bv1G017720.1 | 74 | 8.17 | 7.942 | 42.43 | 0.177 | Extracellular | 1–18 |
| TdPDF10 | TRITD2Av1G011110.1 | 75 | 5.72 | 7.949 | 64.70 | 0.313 | Extracellular | 1–28 |
| TdPDF11 | TRITD2Av1G011190.1 | 77 | 8.47 | 8.295 | 46.32 | 0.386 | Extracellular | 1–27 |
| TdPDF12 | TRITD2Bv1G014190.1 | 75 | 5.72 | 7.947 | 70.96 | 0.299 | Extracellular | 1–28 |
| TdPDF13 | TRITD2Bv1G014230.1 | 78 | 8.48 | 8.359 | 42.82 | 0.373 | Extracellular | 1–28 |
| TdPDF14 | TRITD2Bv1G187060.1 | 85 | 8.91 | 9.857 | 33.90 | −0.455 | Extracellular | 1–32 |
| TdPDF15 | TRITD3Av1G250280.1 | 76 | 8.14 | 8.424 | 20.38 | 0.242 | Extracellular | 1–20 |
| TdPDF16 | TRITD3Av1G250300.1 | 74 | 8.46 | 8.132 | 38.15 | 0.264 | Extracellular | 1–29 |
| TdPDF17 | TRITD3Av1G250390.1 | 123 | 6.63 | 13.426 | 51.05 | −0.145 | Mitochondrion | - |
| TdPDF18 | TRITD3Bv1G242100.1 | 72 | 8.47 | 8.003 | 22.45 | 0.215 | Extracellular | 1–25 |
| TdPDF19 | TRITD4Bv1G184330.1 | 82 | 9.60 | 9.076 | 63.91 | −0.160 | Extracellular | 1–33 |
| TdPDF20 | TRITD4Bv1G184350.1 | 81 | 9.54 | 8.644 | 42.15 | 0.086 | Extracellular | 1–33 |
| TdPDF21 | TRITD4Bv1G184360.1 | 82 | 9.49 | 8.912 | 26.87 | −0.144 | Extracellular | 1–33 |
| TdPDF22 | TRITD5Av1G239070.1 | 82 | 9.69 | 8.945 | 26.52 | −0.133 | Extracellular | 1–33 |
| TdPDF23 | TRITD5Bv1G174690.1 | 80 | 6.50 | 8.666 | 34.92 | 0.216 | Extracellular | - |
| TdPDF24 | TRITD6Av1G019320.1 | 82 | 9.22 | 8.761 | 43.00 | 0.195 | Extracellular | 1–26 |
| TdPDF25 | TRITD6Bv1G028170.1 | 82 | 9.22 | 8.819 | 40.65 | 0.155 | Extracellular | 1–33 |
| TdPDF26 | TRITD6Bv1G028180.1 | 82 | 9.08 | 8.885 | 58.38 | 0.113 | Extracellular | 1–33 |
| TdPDF27 | TRITD6Bv1G069150.1 | 86 | 5.55 | 9.498 | 61.52 | −0.117 | Extracellular | 1–21 |
| TdPDF28 | TRITD7Bv1G138710.1 | 83 | 8.16 | 9.170 | 37.04 | −0.212 | Extracellular | 1–34 |
| T. durum | T. aestivum | |||||
|---|---|---|---|---|---|---|
| Exon Number | Gene | Duplicate Gene | Homologs/ Paralogs | Duplication Type | Orthologs | Exon Number |
| 2 | TdPDF3 | TdPDF4 | Paralogs | Segmental duplication | TaPDF1 | 2 |
| 2 | TdPDF1 | - | - | - | - | - |
| 2 | TdPDF7 | TdPDF2 | Homologs | Whole-genome duplication | TaPDF2 | 2 |
| 2 | TdPDF7 | TdPDF28 | Paralogs | Segmental duplication | TaPDF2 | 2 |
| 2 | TdPDF8 | TdPDF28 | Paralogs | Segmental duplication | TaPDF3 | 2 |
| 2 | TdPDF6 | TdPDF3 | Homologs | Whole-genome duplication | TaPDF1 | 2 |
| 2 | TdPDF6 | TdPDF9 | Paralogs | Segmental duplication | TaPDF1 | 2 |
| 2 | TdPDF6 | TdPDF4 | Homologs | Whole-genome duplication | TaPDF1 | 2 |
| 2 | TdPDF12 | TdPDF10 | Homologs | Whole-genome duplication | TaPDF5 | 2 |
| 2 | TdPDF13 | TdPDF11 | Homologs | Whole-genome duplication | TaPDF6 | 2 |
| 2 | TdPDF13 | TdPDF12 | Paralogs | Segmental duplication | TaPDF6 | 2 |
| 2 | TdPDF15 | TdPDF16 | Paralogs | Segmental duplication | TaPDF10 | 3 |
| 2 | TdPDF17 | TdPDF16 | Paralogs | Segmental duplication | TaPDF13 | 2 |
| 2 | TdPDF18 | TdPDF15 | Homologs | Whole-genome duplication | TaPDF11 | 2 |
| 1 | TdPDF20 | TdPDF21 | Paralogs | Segmental duplication | TaPDF15 | 1 |
| 1 | TdPDF21 | TdPDF19 | Paralogs | Segmental duplication | TaPDF14 | 4 |
| 1 | TdPDF22 | TdPDF21 | Homologs | Whole-genome duplication | TaPDF20 | 1 |
| 2 | TdPDF24 | TdPDF25 | Homologs | Whole-genome duplication | TaPDF21 | 2 |
| 2 | TdPDF24 | TdPDF5 | Paralogs | Segmental duplication | TaPDF21 | 2 |
| 2 | TdPDF26 | TdPDF14 | Paralogs | Segmental duplication | TaPDF25 | 2 |
| 2 | TdPDF26 | TdPDF11 | Paralogs | Segmental duplication | TaPDF25 | 2 |
| Gene | Duplicate | Ka | Ks | Ka/Ks |
|---|---|---|---|---|
| TdPDF 3 | TdPDF4 | 0.2146 | 0.3813 | 0.5629 |
| TdPDF7 | TdPDF2 | 0.0328 | 0.1481 | 0.2219 |
| TdPDF7 | TdPDF28 | 0.1199 | 0.0891 | 1.3454 |
| TdPDF8 | TdPDF28 | 0.0508 | 0.0736 | 0.6903 |
| TdPDF6 | TdPDF3 | 0.0551 | 0.1738 | 0.3173 |
| TdPDF6 | TdPDF9 | 0.2252 | 0.3708 | 0.6073 |
| TdPDF6 | TdPDF4 | 0.2422 | 0.5648 | 0.4287 |
| TdPDF12 | TdPDF10 | 0.0298 | 0.0577 | 0.5167 |
| TdPDF13 | TdPDF11 | 0.0172 | 0.1178 | 0.1462 |
| TdPDF13 | TdPDF12 | 0.2303 | 0.5163 | 0.4461 |
| TdPDF15 | TdPDF16 | 0.0333 | 0.0555 | 0.5998 |
| TdPDF17 | TdPDF16 | 0.2330 | 1.0568 | 0.2204 |
| TdPDF18 | TdPDF15 | 0.0243 | 0.1092 | 0.2229 |
| TdPDF20 | TdPDF21 | 0.1077 | 0.1375 | 0.7835 |
| TdPDF22 | TdPDF21 | 0.0109 | 0.0665 | 0.1653 |
| TdPDF24 | TdPDF25 | 0.0161 | 0.0531 | 0.3038 |
| TdPDF24 | TdPDF5 | 0.1132 | 0.1997 | 0.5669 |
| TdPDF26 | TdPDF14 | 0.3258 | 0.4370 | 0.7454 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gamas, N.; Smaoui, F.; Ben Romdhane, W.; Wiszniewska, A.; Baazaoui, N.; Bouteraa, M.T.; Chouaibi, Y.; Ben Hsouna, A.; Kačániová, M.; Kluz, M.I.; et al. Genome-Wide Identification of the Defensin Gene Family in Triticum durum and Assessment of Its Response to Environmental Stresses. Biology 2025, 14, 404. https://doi.org/10.3390/biology14040404
Gamas N, Smaoui F, Ben Romdhane W, Wiszniewska A, Baazaoui N, Bouteraa MT, Chouaibi Y, Ben Hsouna A, Kačániová M, Kluz MI, et al. Genome-Wide Identification of the Defensin Gene Family in Triticum durum and Assessment of Its Response to Environmental Stresses. Biology. 2025; 14(4):404. https://doi.org/10.3390/biology14040404
Chicago/Turabian StyleGamas, Nawress, Fahmi Smaoui, Walid Ben Romdhane, Alina Wiszniewska, Narjes Baazaoui, Mohamed Taieb Bouteraa, Yosra Chouaibi, Anis Ben Hsouna, Miroslava Kačániová, Maciej Ireneusz Kluz, and et al. 2025. "Genome-Wide Identification of the Defensin Gene Family in Triticum durum and Assessment of Its Response to Environmental Stresses" Biology 14, no. 4: 404. https://doi.org/10.3390/biology14040404
APA StyleGamas, N., Smaoui, F., Ben Romdhane, W., Wiszniewska, A., Baazaoui, N., Bouteraa, M. T., Chouaibi, Y., Ben Hsouna, A., Kačániová, M., Kluz, M. I., Garzoli, S., & Ben Saad, R. (2025). Genome-Wide Identification of the Defensin Gene Family in Triticum durum and Assessment of Its Response to Environmental Stresses. Biology, 14(4), 404. https://doi.org/10.3390/biology14040404