
A New Fossil Species of Nothotsuga from the Mula Basin, Litang County, Sichuan Province and Its Paleoclimate and Paleoecology Significance


Abstract
Simple Summary
Abstract
1. Introduction
2. Geological Backgrounds
3. Materials and Methods
4. Systematic Paleontology
- Order: Pinales Dumortier
- Family: Pinaceae Spreng. ex F. Rudolphi
- Genus: Nothotsuga Hu ex C. N. Page
- Species: Nothotsuga mulaensis Z. Li & J. L. Dong, sp. nov.
- Holotype: LTML001 (Figure 2A,B)
- Type locality: Xiamula village, Litang County, Sichuan Province, southwestern China.
- Type horizon: Changtai Formation, Miocene.
- Repository: Nature Museum of the Chengdu University of Technology.
- Etymology: The specific epithet “mula” refers to the fossil site, Mula region, Litang County, the eastern Tibetan Plateau.

4.1. Diagnosis
4.2. Description
5. Discussion
5.1. Comparisons with Extant Taxa Pinaceae
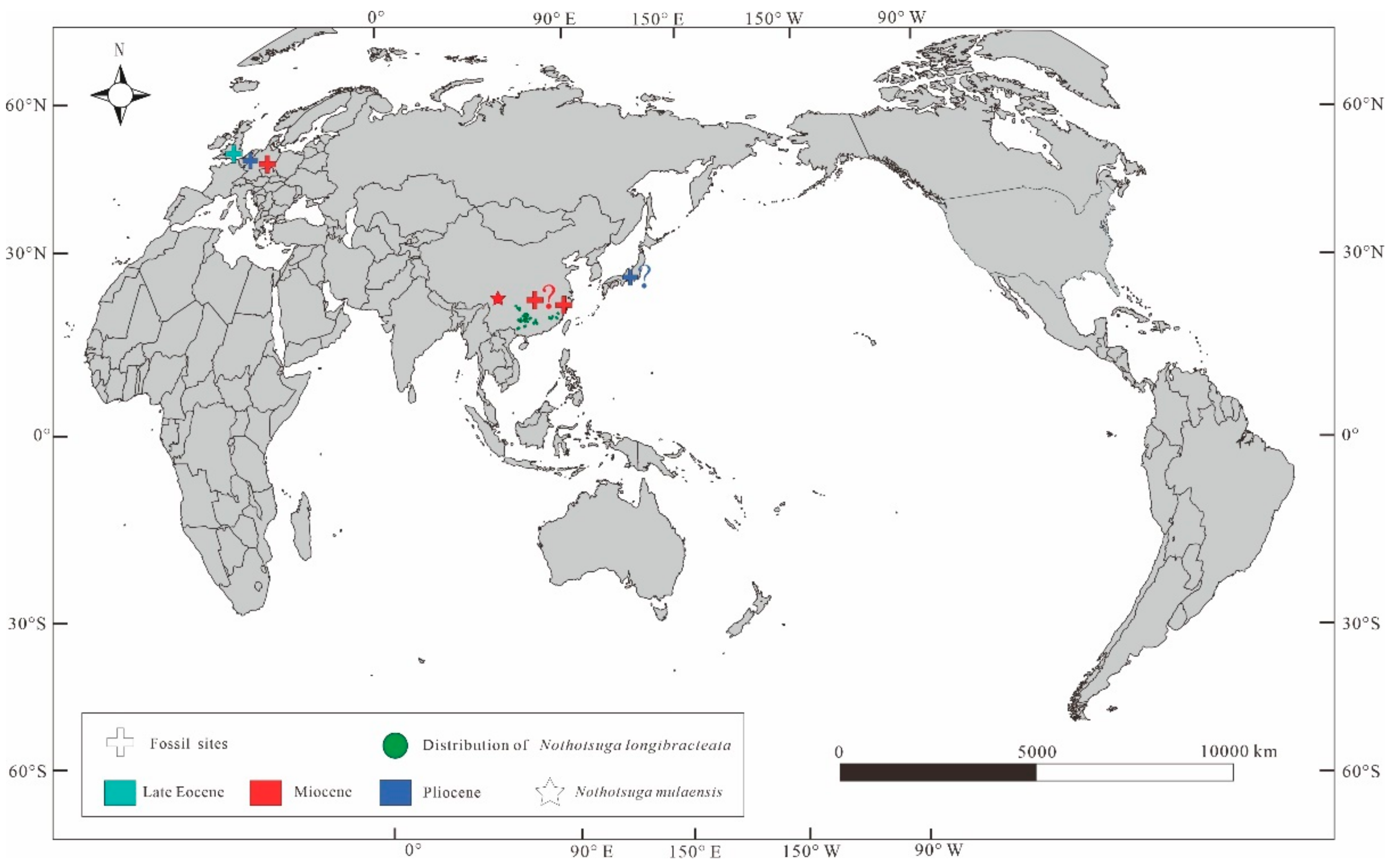
5.2. Implications for Biogeography of Nothotsuga
5.3. Implications for the Paleoclimate and Paleoecology of Nothotsuga
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, Y.J.; Su, T.; Zhou, Z.K. Late Pliocene diversity and distribution of Drynaria (Polypodiaceae) in western Yunnan explained by forest vegetation and humid climates. Plant Divers 2016, 38, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.L.; Su, T.; Huang, J. Occurrence of Christella (Thelypteridaceae) in Southwest China and its indications of the paleoenvironment of the Qinghai–Tibetan Plateau and adjacent areas. J. Syst. Evol. 2019, 57, 169–179. [Google Scholar] [CrossRef]
- Boufford, D.E. Biodiversity Hotspot: China’s Hengduan Mountains. Arnoldia 2014, 72, 24–35. [Google Scholar]
- Xing, Y.W.; Ree, R.H. Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot. Proc. Natl. Acad. Sci. USA 2017, 114, E3444–E3451. [Google Scholar] [CrossRef]
- Ding, W.N.; Ree, R.H.; Spicer, R.A.; Xing, Y.W. Ancient orogenic and monsoon-driven assembly of the world’s richest temperate alpine flora. Science 2020, 369, 578–581. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Qian, L.S.; Spalink, D.; Sun, L.; Chen, J.G.; Sun, H. Spatial phylogenetics of two topographic extremes of the Hengduan Mountains in southwestern China and its implications for biodiversity conservation. Plant Divers 2021, 43, 181–191. [Google Scholar] [CrossRef]
- Qiu, Y.J.; Yi, G.M.; Ning, Z.L.; Huang, H.W. Geographic distribution, current status of resources and endangered factors of endangered plant Nothotsuga Longibracteata. J. Plant Resour. Environ. 2011, 20, 53–59, (In Chinese, with English abstract). [Google Scholar]
- Page, N.A. New and maintained genera in the conifer families Podocarpaceae and Pinaceae. Notes Roy. Bot. Gard. Edinburgh 1988, 45, 377–395. [Google Scholar]
- Zheng, W.J.; Fu, L.G. Flora reipublicae popularis sinicae. In Tomus 7: Gymnospermae; Science Press: Beijing, China, 1978. (In Chinese) [Google Scholar]
- Farjon, A. A Handbook of the World’s Conifers; Revised and Updated Edition; Brill Academic Publishers: Leiden, The Netherlands, 2017. [Google Scholar]
- Tan, X.; Zhang, L.; Zhang, A.P.; Wang, Y.; Huang, D.; Wu, X.G.; Sun, X.M.; Xiong, Q.L.; Pan, K.W. The suitable distribution area of Tsuga longibracteata revealed by a climate and spatial constraint model under future climate change scenarios. Acta Ecol. Sin. 2018, 38, 8934–8945, (In Chinese, with English abstract). [Google Scholar]
- Lin, J.X.; Hu, Y.S.; Wang, X.T. The biology and conservation of Tsuga longibracteata. Chin. Biodivers. 1995, 3, 147–152, (In Chinese, with English abstract). [Google Scholar]
- Zhang, B.; Yao, Y.; Zhao, C.; Wang, J.; Yu, F. Conifers in mountains of China. In Conifers; Gonçalves, A.C., Ed.; IntechOpen: London, UK, 2018; pp. 91–111. [Google Scholar]
- Sadowski, E.M.; Schmidt, A.R.; Seyfullah, L.J.; Kunzmann, L. Conifers of the “Baltic Amber Forest” and their palaeoecological significance. Land Oberösterreich Oberösterreichisches Landesmus. 2017, 106, 1–73. [Google Scholar]
- Kunzmann, L.; Mai, D.H. Conifers of the Mastixioideae-flora from Wiesa near Kamenz (Saxony, Miocene) with special consideration of leaves. Palaeontogr. Abt. B. 2005, 272, 67–135. [Google Scholar] [CrossRef]
- Kunzmann, L. On the fossil history of Pseudotsuga Carr. (Pinaceae) in Europe. Palaeobio. Paleoenv. 2014, 94, 393–409. [Google Scholar] [CrossRef]
- Ding, S.T.; Chen, S.Y.; Ruan, S.C.; Yang, M.; Han, Y.; Wang, X.H.; Zhang, T.H.; Sun, B.N. First fossil record of Nothotsuga (Pinaceae) in China: Implications for palaeobiogeography and palaeoecology. Hist. Biol. 2021, 33, 3617–3624. [Google Scholar] [CrossRef]
- Mai, D.H. Fossile Koniferenreste in der meridionalen Zone Europas. Feddes Repert. 1994, 105, 207–227. [Google Scholar] [CrossRef]
- Fu, L.G. The Red Book of Chinese Plants; Sciences Press: New York, NY, USA, 1991. [Google Scholar]
- Wang, S.; Xie, Y. The Red List of Chinese Species; Higher Education Press: Beijing, China, 2004. [Google Scholar]
- Jackson, J.W.T.; Robinson, D.M.; Weislogel, A.L.; Shang, F.; Jian, X. Mesozoic development of nonmarine basins in the northern Yidun terrane: Deposition and deformation in the eastern Tibetan Plateau prior to the India–Asia collision. Tectonics 2018, 37, 2466–2485. [Google Scholar] [CrossRef]
- Tian, Y.; Kohn, B.P.; Gleadow, A.J.W.; Hu, S.A. Thermochronological perspective on the morphotectonic evolution of the southeastern Tibetan Plateau. J. Geophys. Res. Solid Earth 2014, 119, 676–698. [Google Scholar] [CrossRef]
- Zheng, J.J. Stratigraphical Lexicon of China-Tertiary. In Editorial Committee of Stratigraphical Lexicon of China; Cheng, Y.Q., Ed.; Geology Press: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Liu, G.W.; Li, W.T. Upper cretaceous and tertiary of western Sichuan plateau. J. Stratigr. 2002, 26, 161–169. [Google Scholar]
- Bureau of Geology and Mineral Resources of Sichuan province (BGMRS). Regional Geology of Sichuan Province; Geological Publishing House: Beijing, China, 1991; pp. 1–735. (In Chinese) [Google Scholar]
- Chen, M.H.; Kong, S.C.; Chen, H. Plant Fossils from Mula Formation in Litang Country and Their Significance in Palaeogeography and Palaeoclimate. In The Qinghai-Tibet Plateau Research Hengduan Mountain Investigation Collection (2); Science Press: Beijing, China, 1986; pp. 71–79, (In Chinese, with English abstract). [Google Scholar]
- Liu, G.W.; Li, J.G. Pollen and spore assemblages from the Miocene Maladun Formation at Hongtupo, Songpan Country, western Sichuan Plateau and their bearing on palaeoenvironments. Acta Palaeontol. Sin. 2019, 58, 402–414. [Google Scholar]
- Fu, L.K.; Li, N.; Elias, T.S.; Mill, R.R. Pinaceae. In Flora of China; Wu, Z.Y., Ed.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 1999; Volume 4, pp. 11–52. [Google Scholar]
- Earle, C.J. Gymnosperm database 2020. Available online: http://www.conifers.org/ (accessed on 6 May 2022).
- Wu, M.X.; Huang, J.; Su, T.; Leng, Q.; Zhou, Z.K. Tsuga seed cones from the late Paleogene of southwestern China and their biogeographical and paleoenvironmental implications. Palaeoworld 2020, 29, 617–628. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Yang, L. Cathaya vanderburghii, a misnomer for European Neogene fossil cones. Taxon 2022, 71, 1107–1111. [Google Scholar] [CrossRef]
- Winterscheid, H.; Gossmann, R. Validation of Cathaya vanderburghii (Pinaceae) from European Neogene. Phytotaxa 2017, 302, 188–192. [Google Scholar] [CrossRef]
- Frankis, M.P. Generic inter-relationships in Pinaceae. Notes Roy. Bot. Gard. Edinburgh 1988, 45, 527–548. [Google Scholar]
- Farjon, A. Pinaceae. Drawings and Descriptions of the Genera Abies, Cedrus, Pseudolarix, Keteleeria, Nothotsuga, Tsuga, Cathaya, Pseudotsuga, Larix and Picea; Koeltz Scientific Books: Königstein, Germany, 1990; pp. 1–330. [Google Scholar]
- Xing, Y.W.; Liu, Y.C.; Su, T.; Jacques, F.M.B.; Zhou, Z. A new Tsuga species from the upper Miocene of Yunnan, southwestern China and its palaeogeographic significance. Palaeoworld 2013, 22, 159–167. [Google Scholar] [CrossRef]
- Miki, S. The occurrence of the remain of Taiwania and Palaeotsuga (n. subg.) from Pliocene beds in Japan. P. Jpn. Acad. 1954, 30, 976–981. [Google Scholar] [CrossRef][Green Version]
- Chen, X.J.; Xu, Y.J.; Li, Y.F.; Qi, G.F.; Xu, R.H. Studies on some fossil wood in Yangluo Town, Xinzhou County, Hubei Province. Liaoning Geol. 1992, 9, 368–373, (In Chinese, with English abstract). [Google Scholar]
- Yang, T.N.; Hou, Z.Q.; Wang, Y.; Zhang, H.R.; Wang, Z.L. Late Paleozoic to Early Mesozoic tectonic evolution of northeast Tibet: Evidence from the Triassic composite western Jinsha-Garzê-Litang suture. Tectonics 2012, 31, TC4004. [Google Scholar] [CrossRef]
- Zetter, R.; Ferguson, D.K. Trapaceae pollen in the Cenozoic. Acta Palaeobot. 2001, 41, 321–339. [Google Scholar]
- Ding, L.; Spicer, R.A.; Yang, J.; Xu, Q.; Cai, F.L.; Li, S.; Lai, Q.Z.; Wang, H.Q.; Spicer, T.; Yue, Y.H. Quantifying the rise of the Himalaya orogen and implications for the South Asian monsoon. Geology 2017, 45, 215–218. [Google Scholar] [CrossRef]
- Su, T.; Spicer, R.A.; Wu, F.X.; Farnsworth, A.; Huang, J.; Rio, C.D.; Deng, T.; Ding, L.; Deng, W.Y.D.; Huang, Y.J.; et al. A Middle Eocene lowland humid subtropical “Shangri-La” ecosystem in central Tibet. Proc. Natl. Acad. Sci. USA 2020, 117, 32989–32995. [Google Scholar] [CrossRef]
- Puri, G.S. Fossil plants and the Himalayan uplift. Indian Batanical Soc. 1946, 25, 157–184. [Google Scholar]
- Su, T.; Spicer, R.A.; Li, S.H.; Xu, H.; Huang, J.; Sherlock, S.; Huang, Y.J.; Li, S.F.; Wang, L.; Jia, L.B.; et al. Uplift, climate and biotic changes at the Eocene–Oligocene transition in south-eastern Tibet. Natl. Sci. Rev. 2019, 6, 495–504. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Ding, L.; Xiong, Z.; Spicer, R.A.; Farnsworth, A.; Valdes, P.J.; Wang, C.; Cai, F.; Wang, H.; Sun, Y.; et al. A distinctive Eocene Asian monsoon and modern biodiversity resulted from the rise of eastern Tibet. Sci. Bull. 2022, 67, 2245–2258. [Google Scholar] [CrossRef] [PubMed]
- Karavayev, M.N. Tsuga longibracteata Cheng, first found in a fossil condition on the territory of the USSR. Bull. De La Société Des Nat. De Moscou Sect. Biol. 1958, 63, 73–76. [Google Scholar]
- Martinetto, E. East Asian elements in the Plio-Pleistocene floras of Italy. In Proceedings of the First International Symposium on Floristic Characteristics and Diversity of East Asian Plants, Kunming, China, 25–27 July 1996; Zhang, A.L., Wu, S.G., Eds.; Springer: Berlin, Germany, 1998; pp. 71–87. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site/Species | Mean Annual Temperature MAT (°C) | Coldest Monthly Mean Temperature (CMMT) (°C) | Mean Temperature of the Warmest Month (MTWM) (°C) | Annual Range of Monthly Mean Temperature (ART) (°C) | Mean Annual Precipitation (MAP) (mm) | Altitude (m) | Reference |
|---|---|---|---|---|---|---|---|
| Nothotsuga longibracteata | 13.0–18.5 | 2.0–9.0 | 21.0–28.5 | 18.0–20.0 | 1100–2000 | 300–2300 | [8,11,12,38] |
| Modern Litang Country | 5.7 | −5.35 | 12.1 | 18.0 | 741.6 | 3700 | This paper |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, J.; Li, Z.; Gao, J.; Wang, Q.; Sun, B. A New Fossil Species of Nothotsuga from the Mula Basin, Litang County, Sichuan Province and Its Paleoclimate and Paleoecology Significance. Biology 2023, 12, 46. https://doi.org/10.3390/biology12010046
Dong J, Li Z, Gao J, Wang Q, Sun B. A New Fossil Species of Nothotsuga from the Mula Basin, Litang County, Sichuan Province and Its Paleoclimate and Paleoecology Significance. Biology. 2023; 12(1):46. https://doi.org/10.3390/biology12010046
Chicago/Turabian StyleDong, Junling, Zhe Li, Jingxin Gao, Qian Wang, and Bainian Sun. 2023. "A New Fossil Species of Nothotsuga from the Mula Basin, Litang County, Sichuan Province and Its Paleoclimate and Paleoecology Significance" Biology 12, no. 1: 46. https://doi.org/10.3390/biology12010046
APA StyleDong, J., Li, Z., Gao, J., Wang, Q., & Sun, B. (2023). A New Fossil Species of Nothotsuga from the Mula Basin, Litang County, Sichuan Province and Its Paleoclimate and Paleoecology Significance. Biology, 12(1), 46. https://doi.org/10.3390/biology12010046