
Development of an SNP Assay for Marker-Assisted Selection of Soil-Borne Rhizoctonia solani AG-2-2-IIIB Resistance in Sugar Beet

 , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.1.1. Germplasm for Rhizoctonia Phenotyping and SNP Discovery
2.1.2. Germplasm for SNP Validation
2.2. Phenotyping for Rhizoctonia Resistance
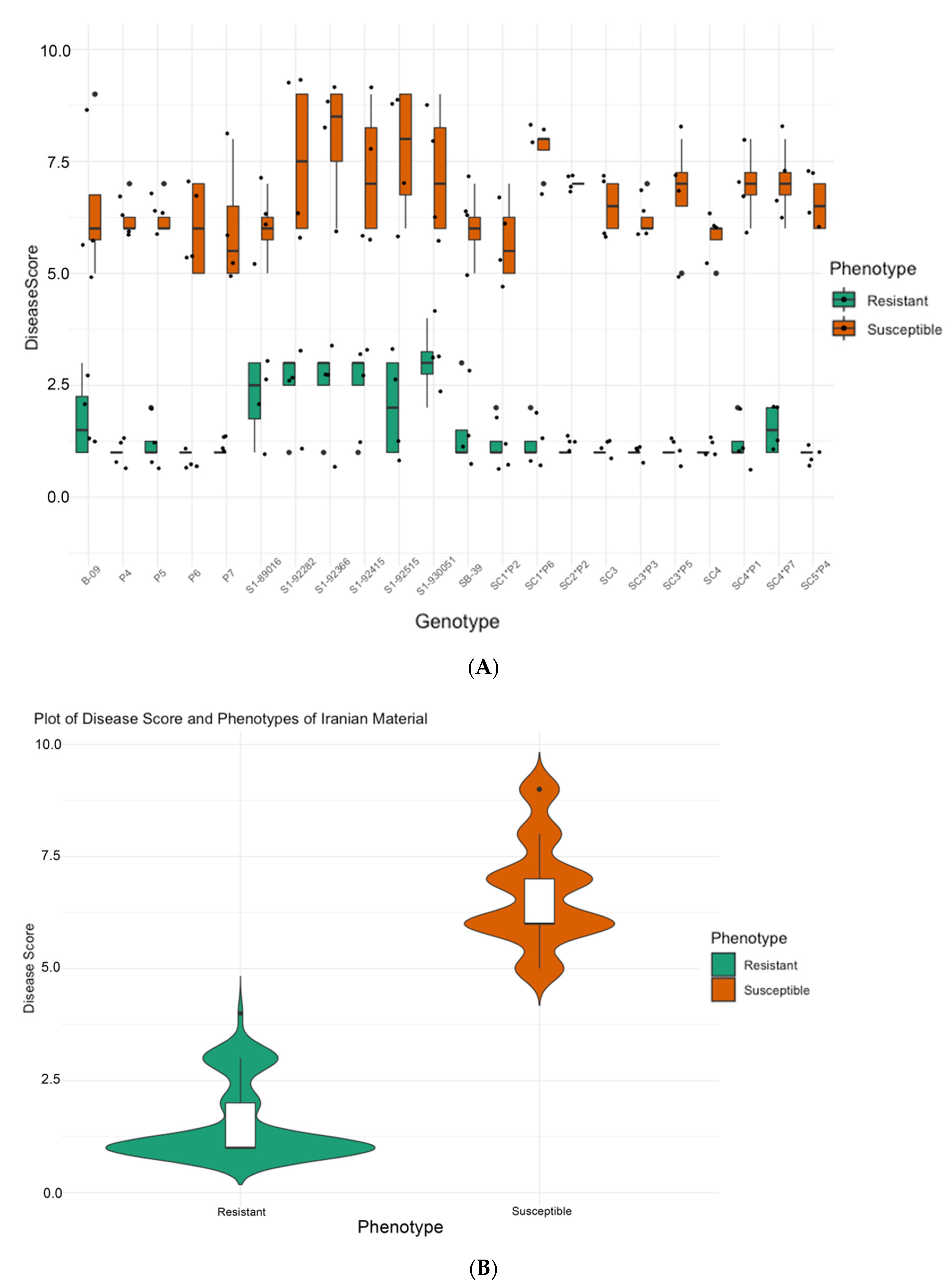
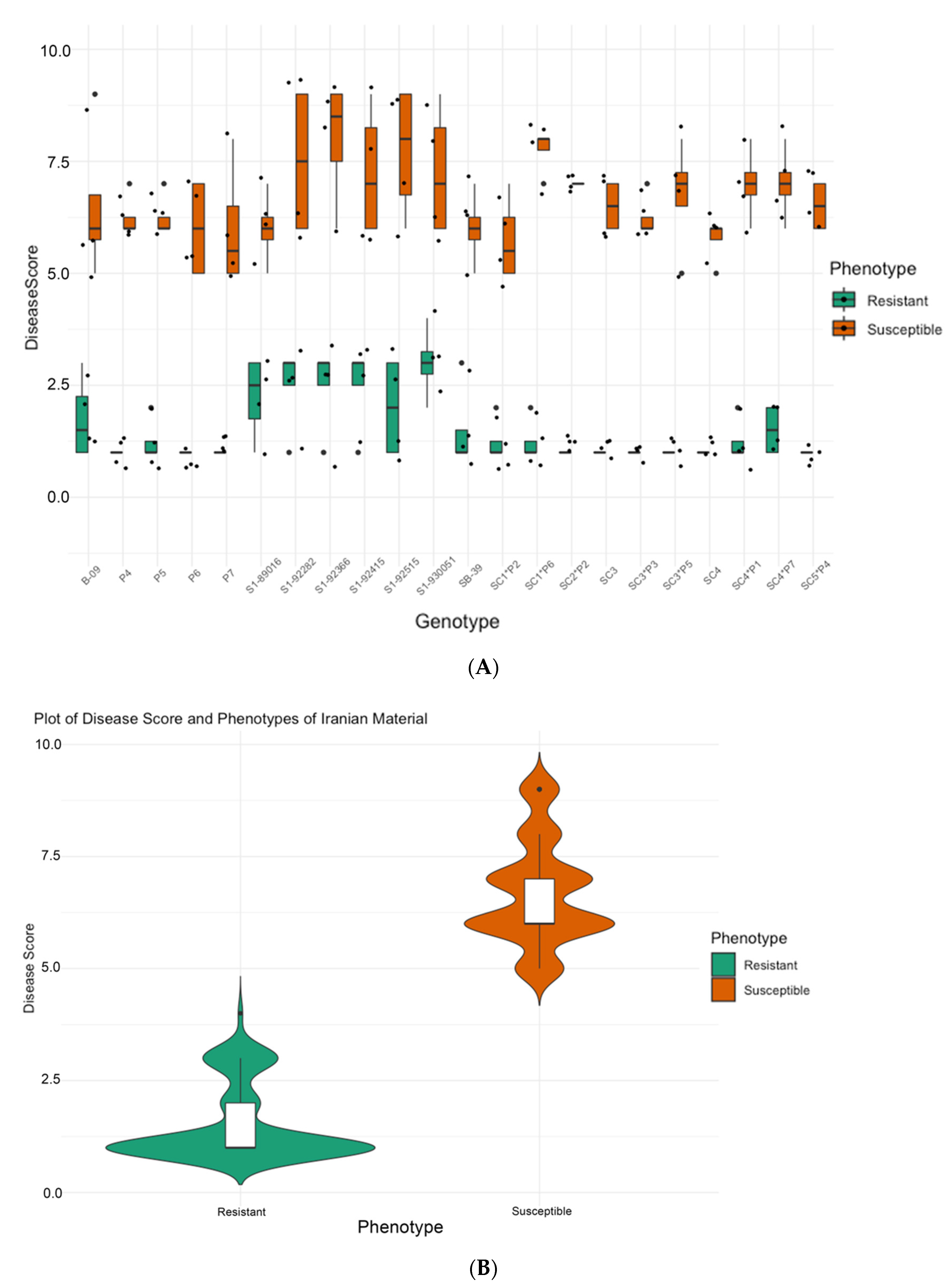
2.2.1. Phenotyping of SBSI Germplasm, Iran
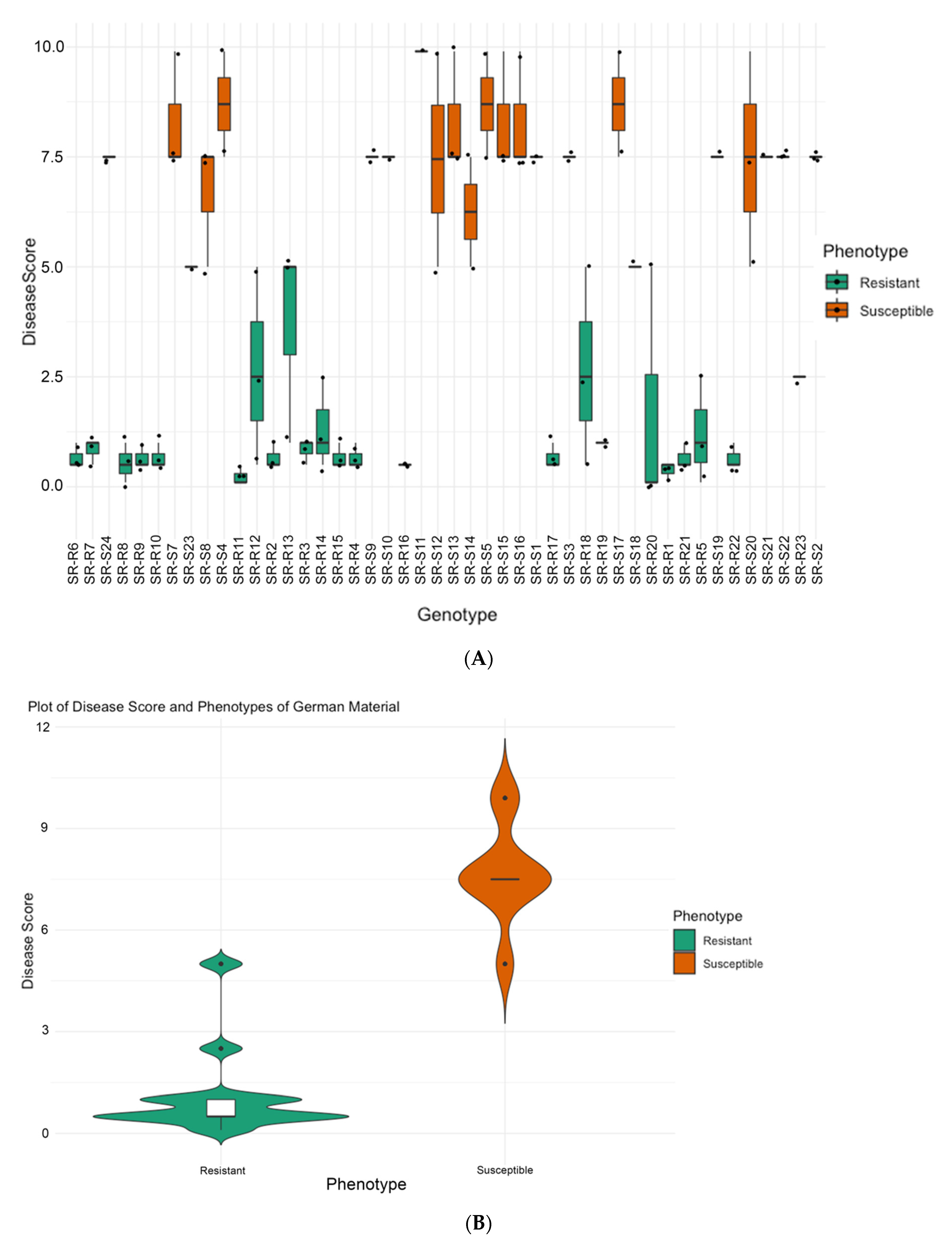
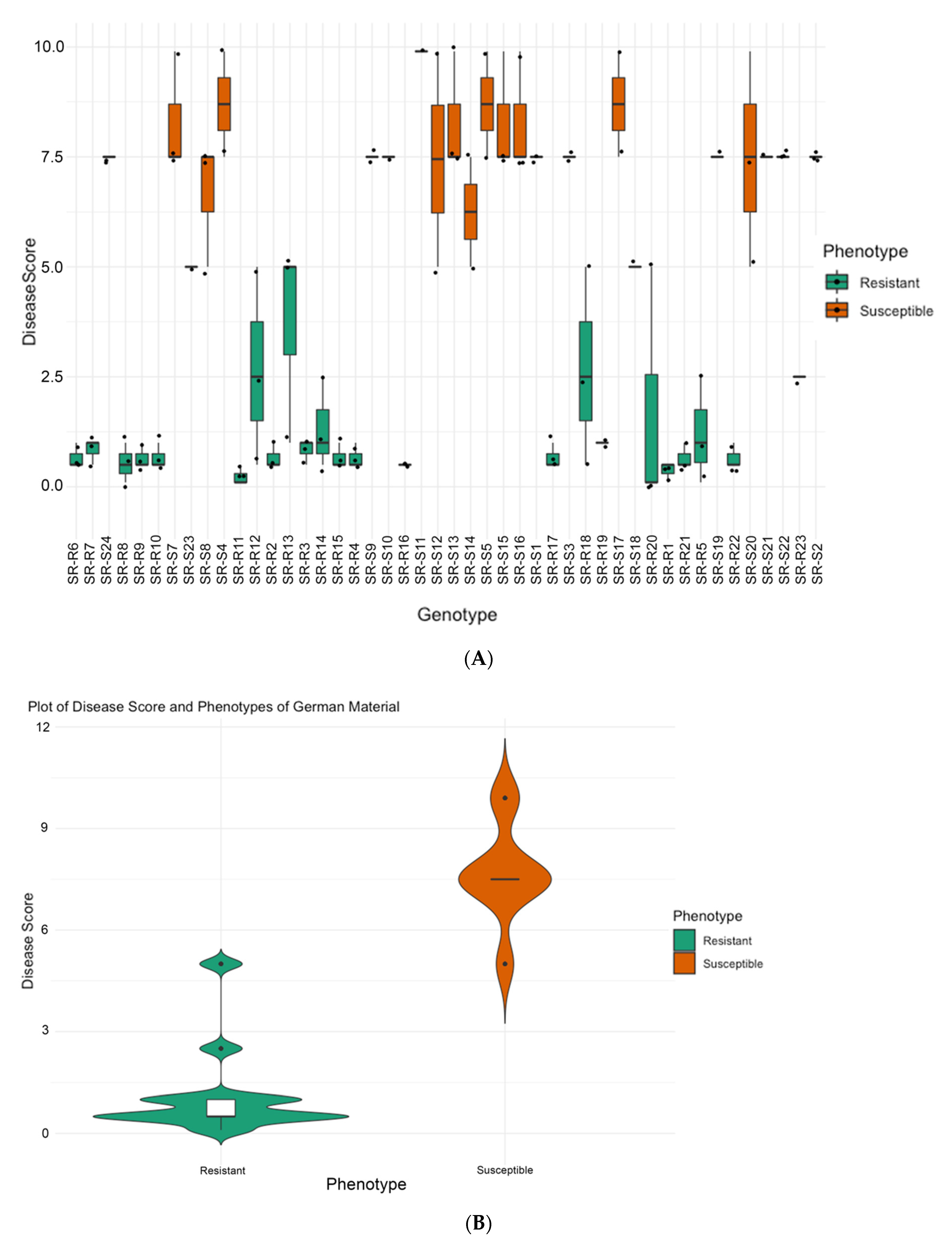
2.2.2. Phenotyping at Strube Research, Germany
2.3. DNA Isolation and RAD Sequencing
2.4. Data Analysis for SNP Discovery
2.5. Molecular Validation
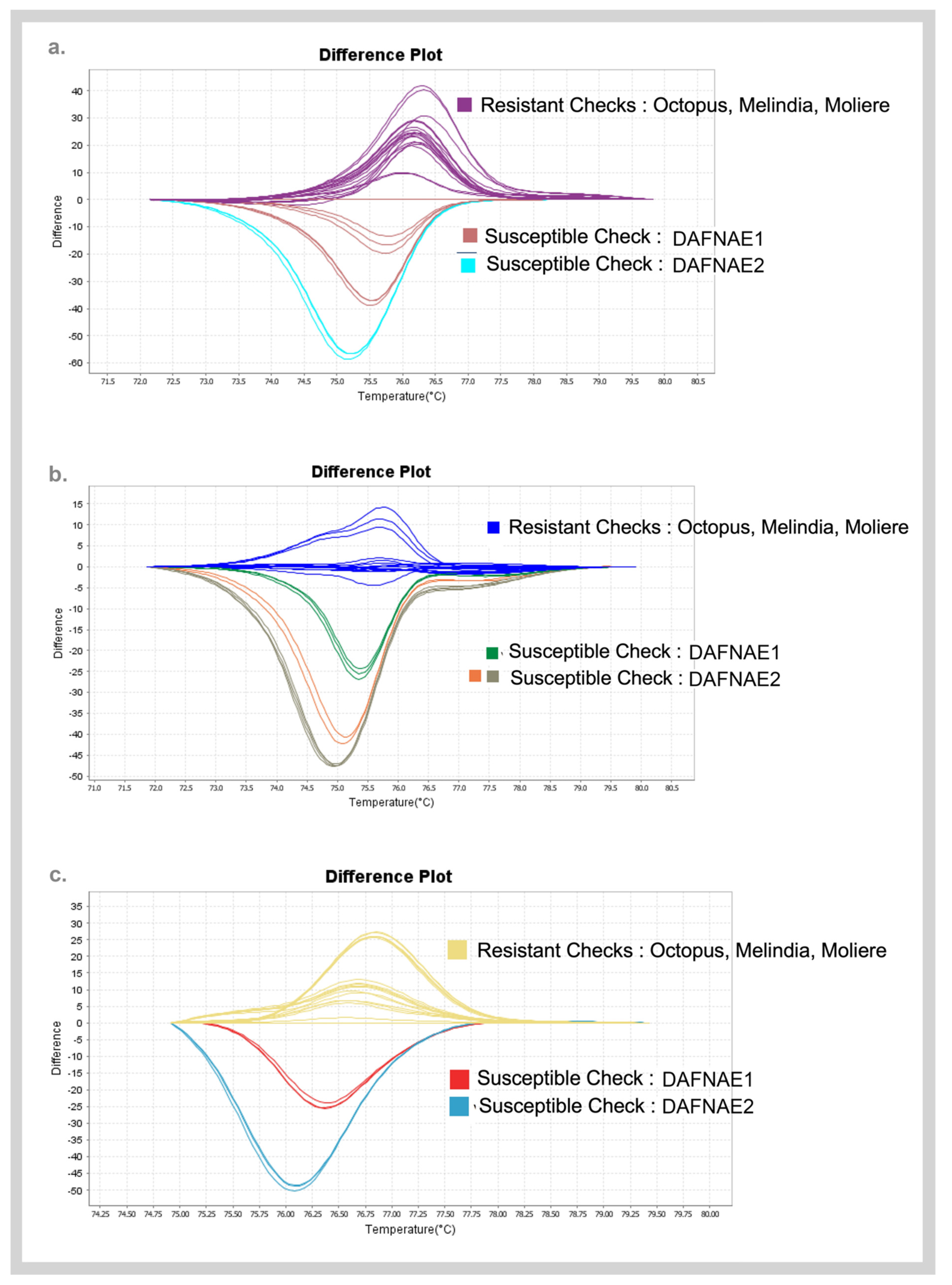
2.5.1. High-Resolution Melting (HRM) Analysis
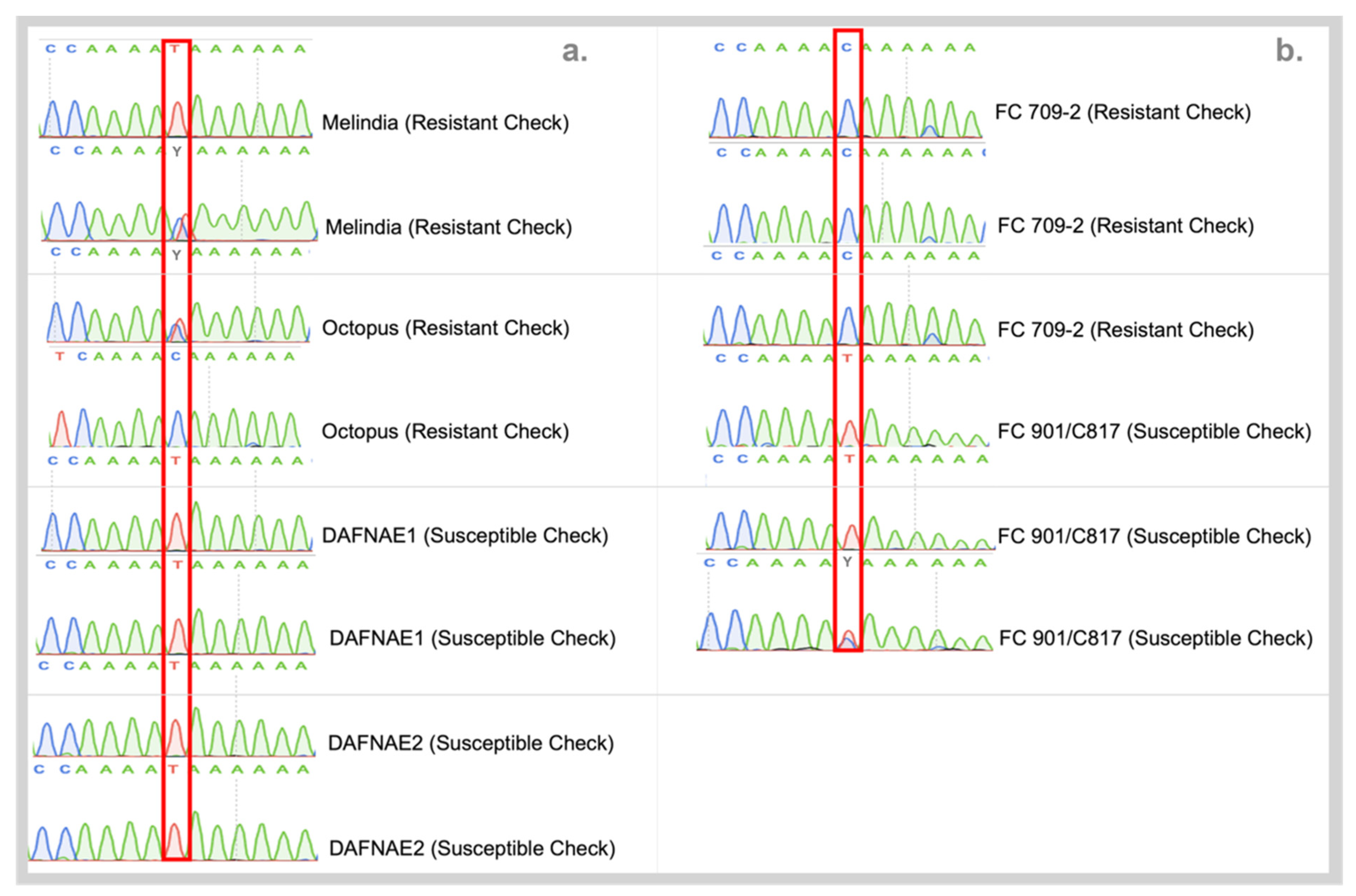
2.5.2. Sanger Sequencing
2.5.3. Genotyping
3. Results
3.1. Phenotyping
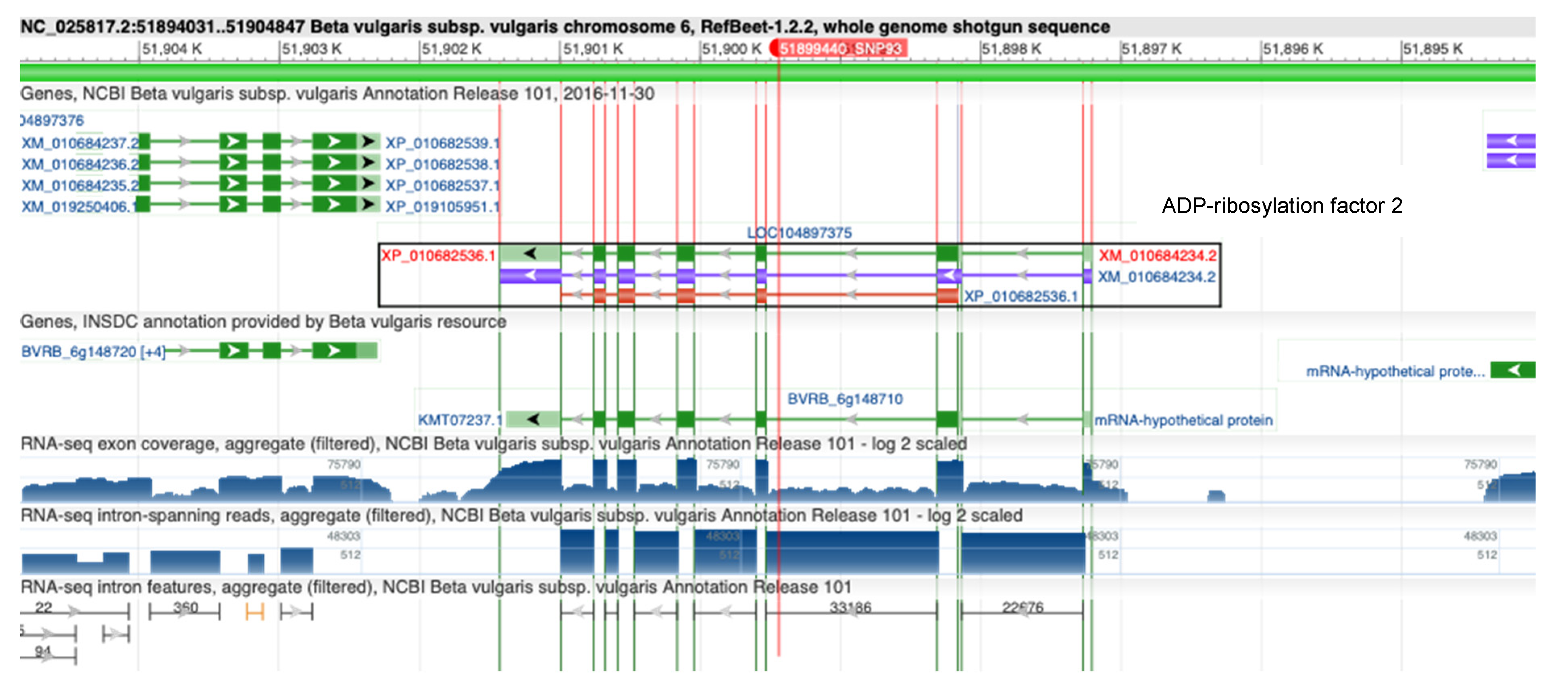
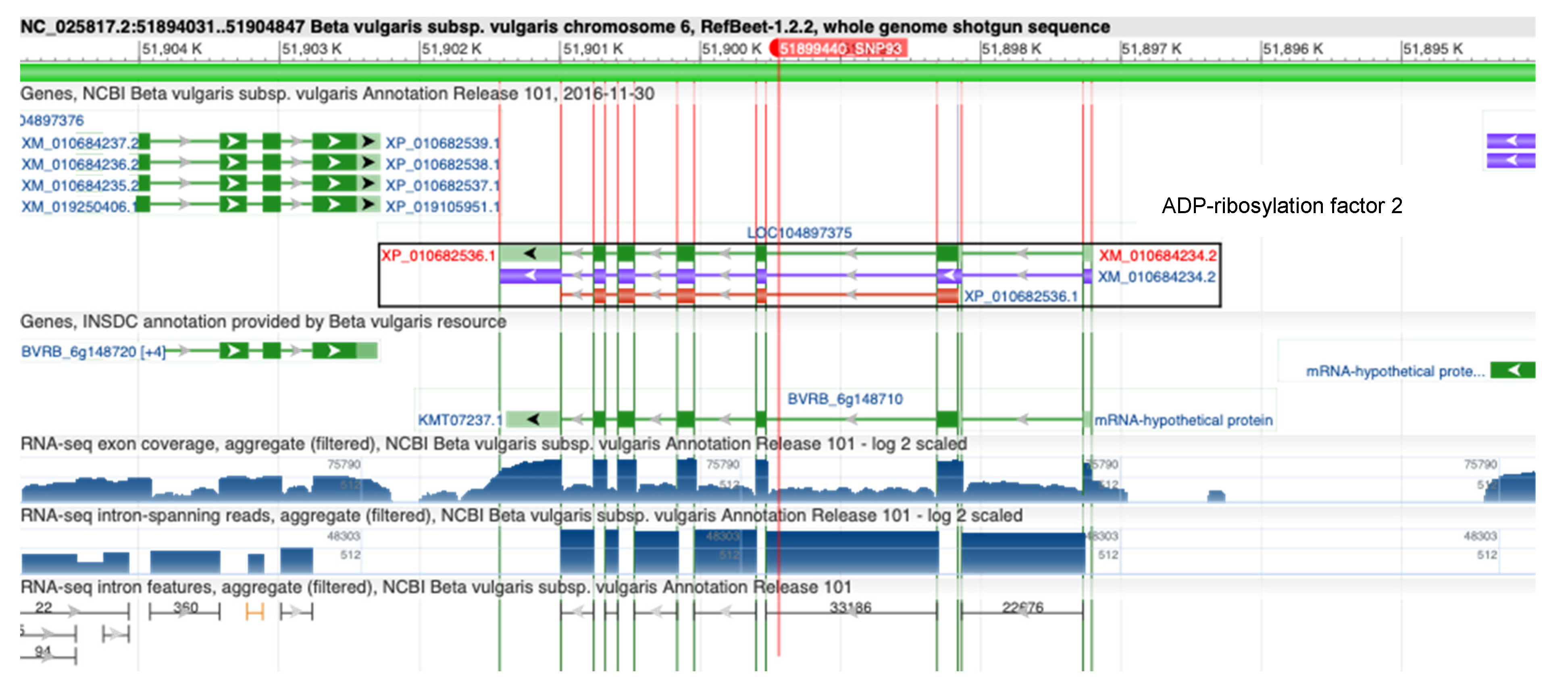
3.2. SNP Discovery for Rhizoctonia Resistance
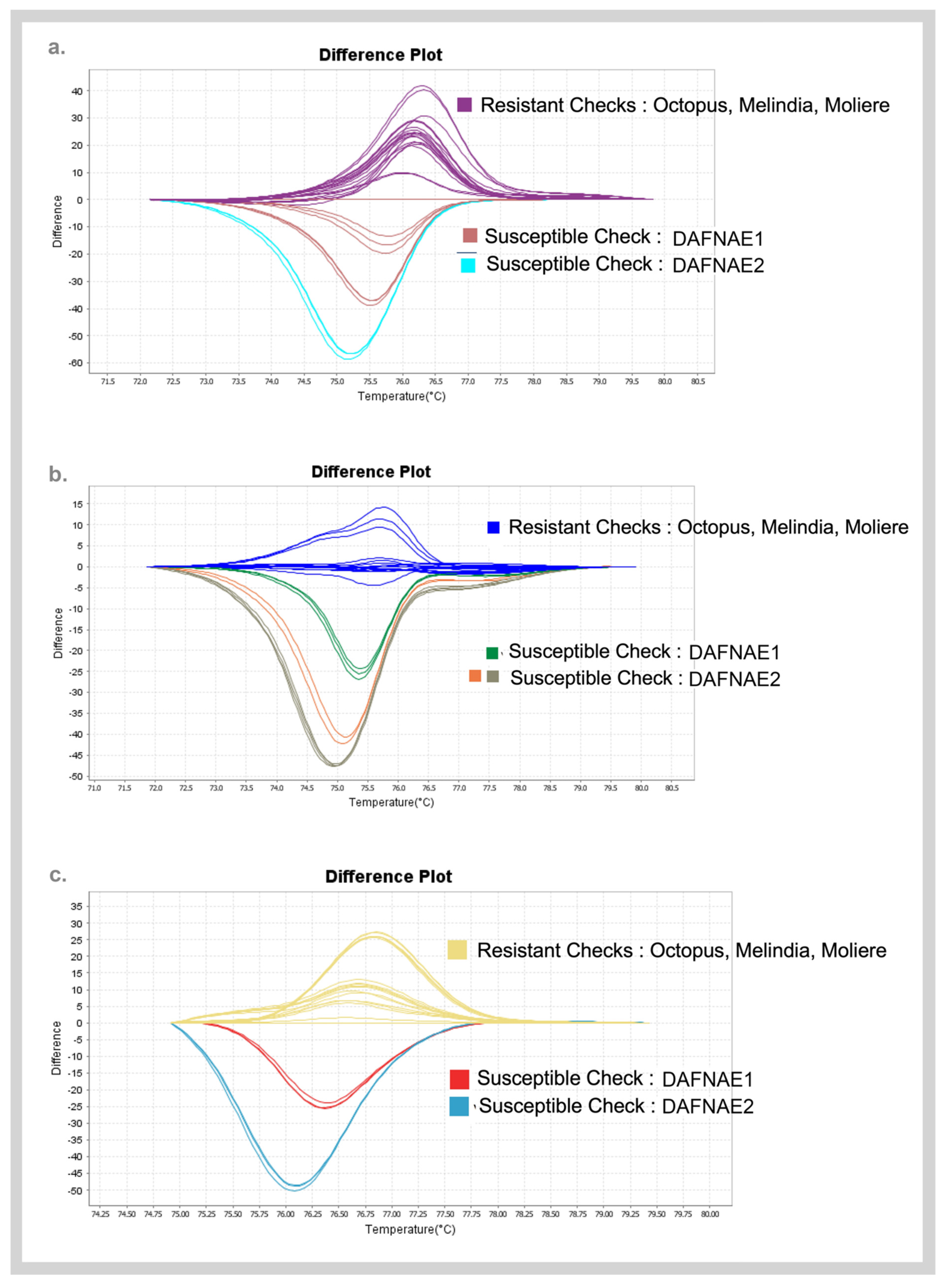
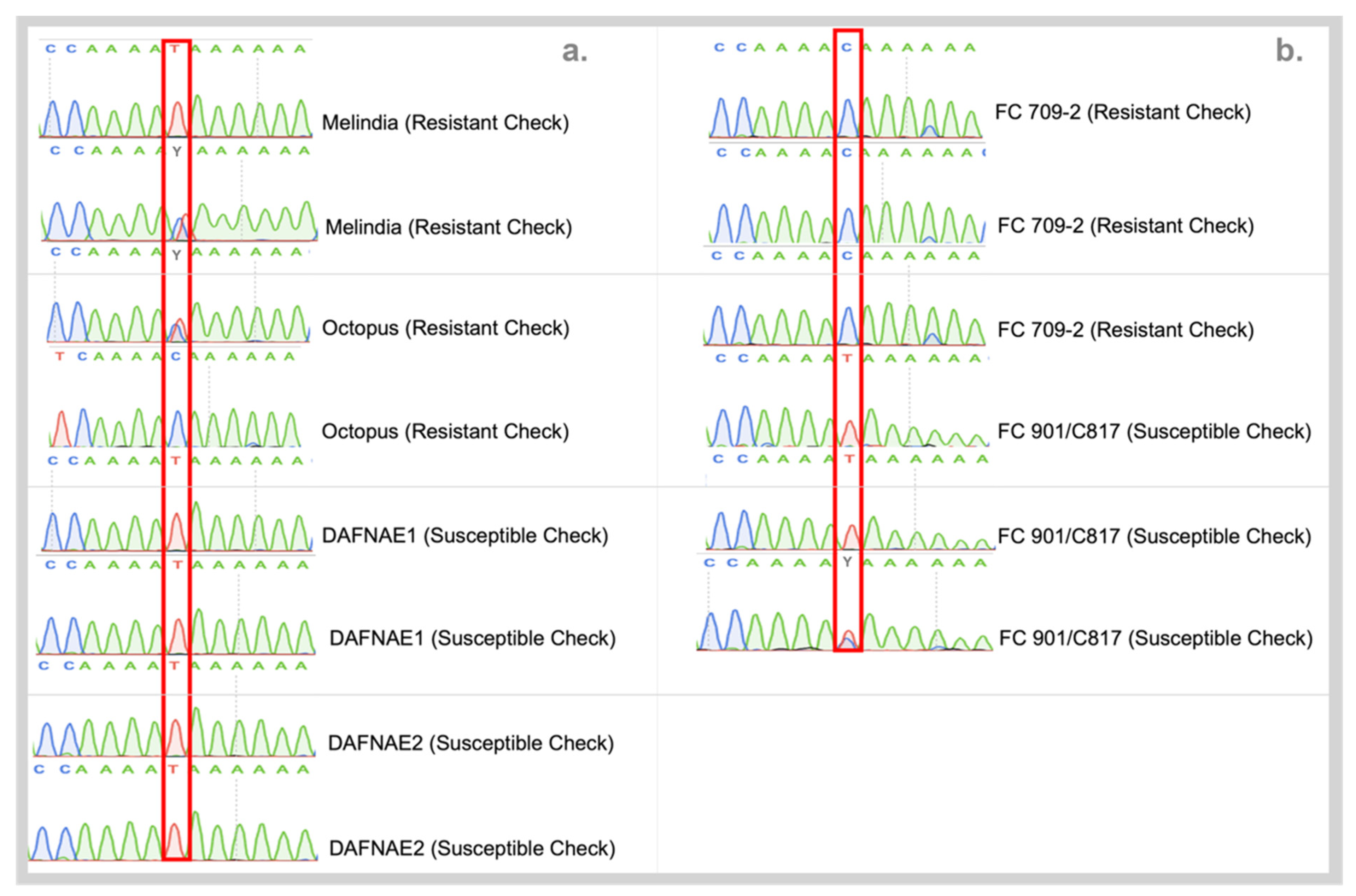
3.3. Validation of SNPs Using High-Resolution Melting (HRM) Analysis and Sanger Sequencing
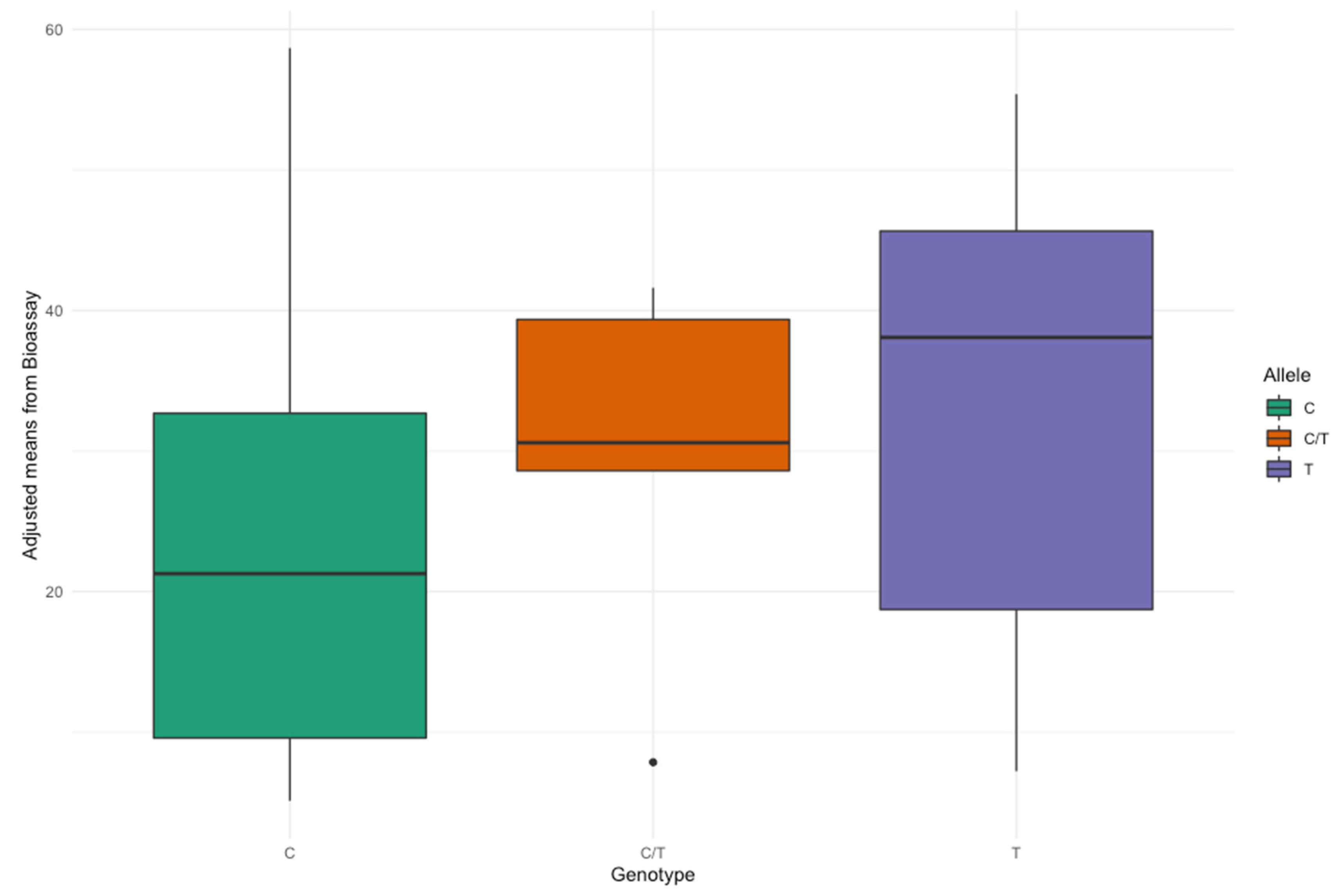
3.4. Association between RsBv1 Marker Genotype and Rhizoctonia Resistance in Strube Research Material
3.5. Further Validation of RsBv1 Based on Genotyping the USDA and SBSI Materials
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ithurrart, M.E.F.; Büttner, G.; Petersen, J. Rhizoctonia Root Rot in Sugar Beet (Beta Vulgaris Ssp. Altissima)—Epidemiological Aspects in Relation to Maize (Zea Mays) as a Host Plant/Rhizoctonia-Rübenfäule in Zuckerrüben (Beta Vulgaris Ssp. Altissima)—Epidemiologische Aspekte in Zusammenhang Mit Mais (Zea Mays) Als Wirtspflanze. Z. Für Pflanzenkrankh. Und Pflanzenschutz/J. Plant Dis. Prot. 2004, 111, 302–312. [Google Scholar]
- Buttner, G.; Pfahler, B.; Marlander, B. Greenhouse and Field Techniques for Testing Sugar Beet for Resistance to Rhizoctonia Root and Crown Rot. Plant Breed. Z. Für Pflanzenzucht. 2004, 123, 158–166. [Google Scholar] [CrossRef]
- Buhre, C.; Kluth, C.; Bürcky, K.; Märländer, B.; Varrelmann, M. Integrated Control of Root and Crown Rot in Sugar Beet: Combined Effects of Cultivar, Crop Rotation, and Soil Tillage. Plant Dis. 2009, 93, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Strausbaugh, C.A.; Eujayl, I.A.; Panella, L.W.; Hanson, L.E. Virulence, Distribution and Diversity of Rhizoctonia Solani from Sugar Beet in Idaho and Oregon. Can. J. Plant Pathol. 2011, 33, 210–226. [Google Scholar] [CrossRef]
- Kluth, C.; Buhre, C.; Varrelmann, M. Susceptibility of Intercrops to Infection with Rhizoctonia Solani AG 2-2 IIIB and Influence on Subsequently Cultivated Sugar Beet. Plant Pathol. 2010, 59, 683–692. [Google Scholar] [CrossRef]
- Gaskill, J.O. Breeding for Rhizoctonia Resistance in Sugarbeet. J. Am. Soc. Sugar Beet Technol. 1968, 15, 105–119. [Google Scholar] [CrossRef]
- Hecker, R.J.; Ruppel, E.G. Rhizoctonia Root-Rot Resistance in Sugarbeet: Breeding and Related Research. J. Am. Soc. Sugar Beet Technol. 1977, 19, 246–256. [Google Scholar] [CrossRef]
- Ruppel, E.G.; Hecker, R.J. Rhizoctonia Root Rot on Sugarbeet Cultivars Having Varied Degrees of Resistance. J. Sugar Beet Res. 1994, 31, 135–142. [Google Scholar] [CrossRef]
- Panella, L. Registration of FC709-2 and FC727 Sugarbeet Germplasm Resistant to Rhizoctonia Root Rot and Cercospora Leaf Spot. Crop Sci. 1999, 39, 298–299. [Google Scholar] [CrossRef]
- Hecker, R.J.; Ruppel, E.G. Registration of Rhizoctonia Root Rot Resistant Sugarbeet Germplasm FC712. Crop Sci. 1986, 26, 213–214. [Google Scholar] [CrossRef]
- Hecker, R.J.; Ruppel, E.G. Polyploid and Maternal Effects on Rhizoctonia Root Rot Resistance in Sugarbeet. Euphytica 1976, 25, 419–423. [Google Scholar] [CrossRef]
- Halloin, J.M. Registration of EL51 Sugarbeet Germplasm with Resistance to Rhizoctonia Crown and Root Rot. Crop Sci. 2000, 40, 586. [Google Scholar] [CrossRef]
- Hecker, R.J.; Gaskill, J.O. Registration of FC 701 and FC 702 Sugarbeet Germplasm1 (Reg. No. GP 1 and 2). Crop Sci. 1972, 12, 400. [Google Scholar] [CrossRef]
- Hecker, J.; Ruppel, E.G. Registration of Diploid and Tetraploid FC 701/4 and FC 703 Sugarbeet Germplasm1 (Reg. Nos. GP 11 to 14). Crop Sci. 1977, 17, 678. [Google Scholar] [CrossRef]
- Hecker, R.J.; Ruppel, E.G. Registration of Rhizoctonia Root Rot Resistant Sugarbeet Germplasms FC 701/6, FC 702/7, and FC 705/1. Crop Sci. 1985, 25, 374. [Google Scholar] [CrossRef] [Green Version]
- Hecker, R.J.; Ruppel, E.G. Registration of Rhizoctonia Root Rot Resistant Sugarbeet Germplasm FC 707(4x). Crop Sci. 1987, 27, 822. [Google Scholar] [CrossRef]
- Hecker, R.J.; Ruppel, E.G. Registration of Rhizoctonia Root Rot Resistant Sugarbeet Germplasm FC 710. Crop Sci. 1991, 31, 494. [Google Scholar] [CrossRef]
- Panella, L.; Campbell, L.G.; Eujayl, I.A.; Lewellen, R.T.; McGrath, J.M. USDA-ARS Sugarbeet Releases and Breeding Over the Past 20 Years. J. Sugar Beet Res. 2016, 52, 40–85. [Google Scholar] [CrossRef]
- Jacobsen, B. Effect of Fungicide And Fungicide Application Timing On Reducing Yield Loss To Rhizoctonia Crown And Root Rot. Sugarbeet Res. Ext. Rep. 2004, 35, 224. [Google Scholar]
- Strausbaugh, C.A.; Eujayl, I.A.; Panella, L.W. Interaction of Sugar Beet Host Resistance and Rhizoctonia Solani AG-2-2 IIIB Strains. Plant Dis. 2013, 97, 1175–1180. [Google Scholar] [CrossRef] [Green Version]
- Bayer, P.E.; Hu, H.; Petereit, J.; Derbyshire, M.C.; Varshney, R.K.; Valliyodan, B.; Nguyen, H.T.; Batley, J.; Edwards, D. Yield Is Negatively Correlated with Nucleotide-Binding Leucine-Rich Repeat Gene Content in Soybean. aRxiv 2021, arXiv:2021.12.12.472330. [Google Scholar]
- Lein, J.; Sagstetter, C.; Scheuermann, D.; Thurau, T.; Varrelmann, M.; Saal, B.; Koch, G.; Borchardt, D.; Jung, C. Mapping of Rhizoctonia Root Rot Resistance Genes in Sugar Beet Using Pathogen Response-related Sequences as Molecular Markers. Plant Breed. 2008, 127, 602–611. [Google Scholar] [CrossRef]
- Holmquist, L.; Dölfors, F.; Fogelqvist, J.; Cohn, J.; Kraft, T.; Dixelius, C. Major Latex Protein-like Encoding Genes Contribute to Rhizoctonia Solani Defense Responses in Sugar Beet. Mol. Genet. Genom. 2021, 296, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Stevanato, P.; Trebbi, D.; Panella, L.; Richardson, K.; Broccanello, C.; Pakish, L.; Fenwick, A.; Saccomani, M. Identification and Validation of a SNP Marker Linked to the Gene HsBvm-1 for Nematode Resistance in Sugar Beet. Plant Mol. Biol. Rep. 2014, 33, 474–479. [Google Scholar] [CrossRef]
- Taranto, F.; Nicolia, A.; Pavan, S.; De Vita, P.; D’Agostino, N. Biotechnological and Digital Revolution for Climate-Smart Plant Breeding. Agronomy 2018, 8, 277. [Google Scholar] [CrossRef] [Green Version]
- Stevanato, P.; Broccanello, C.; Pajola, L.; Biscarini, F.; Richards, C.; Panella, L.; Hassani, M.; Formentin, E.; Chiodi, C.; Concheri, G.; et al. Targeted Next-Generation Sequencing Identification of Mutations in Disease Resistance Gene Analogs (RGAs) in Wild and Cultivated Beets. Genes 2017, 8, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trebbi, D.; Ravi, S.; Broccanello, C.; Chiodi, C.; Francis, G.; Oliver, J.; Mulpuri, S.; Srinivasan, S.; Stevanato, P. Identification and Validation of SNP Markers Linked to Seed Toxicity in Jatropha curcas L. Sci. Rep. 2019, 9, 10220. [Google Scholar] [CrossRef] [Green Version]
- Blary, A.; Jenczewski, E. Manipulation of Crossover Frequency and Distribution for Plant Breeding. Theor. Appl. Genet. 2019, 132, 575–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dohm, J.C.; Minoche, A.E.; Holtgräwe, D.; Capella-Gutiérrez, S.; Zakrzewski, F.; Tafer, H.; Rupp, O.; Sörensen, T.R.; Stracke, R.; Reinhardt, R.; et al. The Genome of the Recently Domesticated Crop Plant Sugar Beet (Beta Vulgaris). Nature 2014, 505, 546–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funk, A.; Galewski, P.; McGrath, J.M. Nucleotide-Binding Resistance Gene Signatures in Sugar Beet, Insights from a New Reference Genome. Plant J. 2018, 95, 659–671. [Google Scholar] [CrossRef] [PubMed]
- Engelkes, C.A.; Windels, C.E. Susceptibility of Sugar Beet and Beans to Rhizoctonia Solani AG-2-2 IIIB and AG-2-2 IV. Plant Dis. 1996, 80, 1413–1417. [Google Scholar] [CrossRef]
- Barreto, L.C.; Martínez-Arias, R.; Schechert, A. Field Detection of Rhizoctonia Root Rot in Sugar Beet by Near Infrared Spectrometry. Sensors 2021, 21, 8068. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J.; Doyle, J.L. Isolation of Plant DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Andrews, K.R.; Good, J.M.; Miller, M.R.; Luikart, G.; Hohenlohe, P.A. Harnessing the Power of RADseq for Ecological and Evolutionary Genomics. Nat. Rev. Genet. 2016, 17, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Ravi, S.; Campagna, G.; Della Lucia, M.C.; Broccanello, C.; Bertoldo, G.; Chiodi, C.; Maretto, L.; Moro, M.; Eslami, A.S.; Srinivasan, S.; et al. SNP Alleles Associated with Low Bolting Tendency in Sugar Beet. Front. Plant Sci. 2021, 12, 693285. [Google Scholar] [CrossRef] [PubMed]
- Leisch, F.; Man, M. Population Genetics. R Package Version 1.3.8.1.; R Foundation: Vienna, Austria, 2021; Available online: Https://CRAN.R-Project.Org/Package=genetics (accessed on 29 November 2021).
- Li, L.-Y. CMplot: Circle Manhattan Plot. R Package Version 3.6.2; R Foundation: Vienna, Austria, 2020; Available online: https://CRAN.R-Project.Org/Package=CMplot (accessed on 29 November 2021).
- Le, S.; Joss, J.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Gauch, H.G., Jr.; Qian, S.; Piepho, H.P.; Zhou, L.; Chen, R. Consequences of PCA Graphs, SNP Codings, and PCA Variants for Elucidating Population Structure. PLoS ONE 2019, 14, e0218306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Ablett, G.R.; Anderson, T.R.; Rajcan, I.; Schaafsma, A.W. Inheritance and Genetic Mapping of Resistance to Rhizoctonia Root and Hypocotyl Rot in Soybean. Crop Sci. 2005, 45, 1441–1447. [Google Scholar] [CrossRef]
- Kaiser, N.; Douches, D.; Dhingra, A.; Glenn, K.C.; Herzig, P.R.; Stowe, E.C.; Swarup, S. The Role of Conventional Plant Breeding in Ensuring Safe Levels of Naturally Occurring Toxins in Food Crops. Trends Food Sci. Technol. 2020, 100, 51–66. [Google Scholar] [CrossRef]
- Bourne, H.R.; Sanders, D.A.; McCormick, F. The GTPase Superfamily: Conserved Structure and Molecular Mechanism. Nature 1991, 349, 117–127. [Google Scholar] [CrossRef]
- Boman, A.L.; Zhang, C.; Zhu, X.; Kahn, R.A. A Family of ADP-Ribosylation Factor Effectors That Can Alter Membrane Transport through the Trans-Golgi. MBoC 2000, 11, 1241–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.Y.; Hong, J.K.; Kim, C.Y.; Chun, H.J.; Park, H.C.; Kim, J.C.; Yun, D.-J.; Chung, W.S.; Lee, S.-H.; Lee, S.Y.; et al. Over-Expressed Rice ADP-Ribosylation Factor 1 (RARF1) Induces Pathogenesis-Related Genes and Pathogen Resistance in Tobacco Plants. Physiol. Plant. 2003, 119, 573–581. [Google Scholar] [CrossRef]
- Meng, H.; Wang, S.; Yang, W.; Ding, X.; Li, N.; Chu, Z.; Li, X. Identification of Virulence Associated MilRNAs and Their Bidirectional Targets in Rhizoctonia Solani and Maize during Infection. BMC Plant Biol. 2021, 21, 155. [Google Scholar] [CrossRef]
- Muthamilarasan, M.; Mangu, V.R.; Zandkarimi, H.; Prasad, M.; Baisakh, N. Structure, Organization and Evolution of ADP-Ribosylation Factors in Rice and Foxtail Millet, and Their Expression in Rice. Sci. Rep. 2016, 6, 24008. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Description | No. of Samples | Replicate | Year |
|---|---|---|---|---|
| S1-930051 | Self-pollinated | 4 | 2 | 2018 |
| S1-89016 | Self-pollinated | 6 | 2 | 2018 |
| S1-92282 | Self-pollinated | 5 | 2 | 2018 |
| S1-92366 | Self-pollinated | 7 | 2 | 2018 |
| S1-92415 | Self-pollinated | 4 | 2 | 2018 |
| S1-92515 | Self-pollinated | 4 | 2 | 2018 |
| SB-39 | Self-pollinated | 5 | 2 | 2018 |
| B-09 | Self-pollinated | 3 | 2 | 2018 |
| SC4*P1 (SC MH41 * SHR01-P.12) | Hybrid | 4 | 2 | 2018 |
| SC1*P2 (SC MH070 * SHR02-P.4) | Hybrid | 5 | 2 | 2018 |
| SC2*P2 (SC MH076 * SHR02-P.4) | Hybrid | 1 | 2 | 2018 |
| SC5*P4 ((7112 * SB36) * S1-88605) | Hybrid | 2 | 2 | 2018 |
| SC3*P5 (SC MH7 * F-8726) | Hybrid | 4 | 2 | 2018 |
| SC1*P6 (SC MH070 * F-8738) | Hybrid | 4 | 2 | 2018 |
| SC4*P7 (SC MH41 * SB27) | Hybrid | 2 | 2 | 2018 |
| P4 (S1-88605) | Pollinator | 5 | 2 | 2018 |
| P5 (F-8726) | Pollinator | 5 | 2 | 2018 |
| P6 (F-8738) | Pollinator | 8 | 2 | 2018 |
| P7 (SB27) | Pollinator | 2 | 2 | 2018 |
| SC3 (SC MH7) | Single crosses (MS) | 7 | 2 | 2018 |
| SC4 (SC MH41) | Single crosses (MS) | 5 | 2 | 2018 |
| Source | C | T | Chi-Square | p-Value | |
|---|---|---|---|---|---|
| USDA | Resistant | 38 | 2 | 51.58 | <0.00001 |
| (n = 20) | 95% | 5% | |||
| Moderately resistant | 32 | 0 | |||
| (n = 16) | 100% | 0% | |||
| Susceptible | 11 | 23 | |||
| (n = 17) | 32.4% | 67.6% | |||
| SBSI | Resistant | 63 | 45 | 18.11 | <0.0001 |
| (n = 54) | 58.3% | 41.7% | |||
| Susceptible | 29 | 71 | |||
| (n = 50) | 29.0% | 71.0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravi, S.; Hassani, M.; Heidari, B.; Deb, S.; Orsini, E.; Li, J.; Richards, C.M.; Panella, L.W.; Srinivasan, S.; Campagna, G.; et al. Development of an SNP Assay for Marker-Assisted Selection of Soil-Borne Rhizoctonia solani AG-2-2-IIIB Resistance in Sugar Beet. Biology 2022, 11, 49. https://doi.org/10.3390/biology11010049
Ravi S, Hassani M, Heidari B, Deb S, Orsini E, Li J, Richards CM, Panella LW, Srinivasan S, Campagna G, et al. Development of an SNP Assay for Marker-Assisted Selection of Soil-Borne Rhizoctonia solani AG-2-2-IIIB Resistance in Sugar Beet. Biology. 2022; 11(1):49. https://doi.org/10.3390/biology11010049
Chicago/Turabian StyleRavi, Samathmika, Mahdi Hassani, Bahram Heidari, Saptarathi Deb, Elena Orsini, Jinquan Li, Christopher M. Richards, Lee W. Panella, Subhashini Srinivasan, Giovanni Campagna, and et al. 2022. "Development of an SNP Assay for Marker-Assisted Selection of Soil-Borne Rhizoctonia solani AG-2-2-IIIB Resistance in Sugar Beet" Biology 11, no. 1: 49. https://doi.org/10.3390/biology11010049
APA StyleRavi, S., Hassani, M., Heidari, B., Deb, S., Orsini, E., Li, J., Richards, C. M., Panella, L. W., Srinivasan, S., Campagna, G., Concheri, G., Squartini, A., & Stevanato, P. (2022). Development of an SNP Assay for Marker-Assisted Selection of Soil-Borne Rhizoctonia solani AG-2-2-IIIB Resistance in Sugar Beet. Biology, 11(1), 49. https://doi.org/10.3390/biology11010049