
Termitophily Documented in Earwigs (Dermaptera)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Material Collection and Identification
2.2. Taxonomy
2.3. Behavioral Observations
2.4. Gastrointestinal Tract Analysis
2.5. Molecular Methods
3. Results
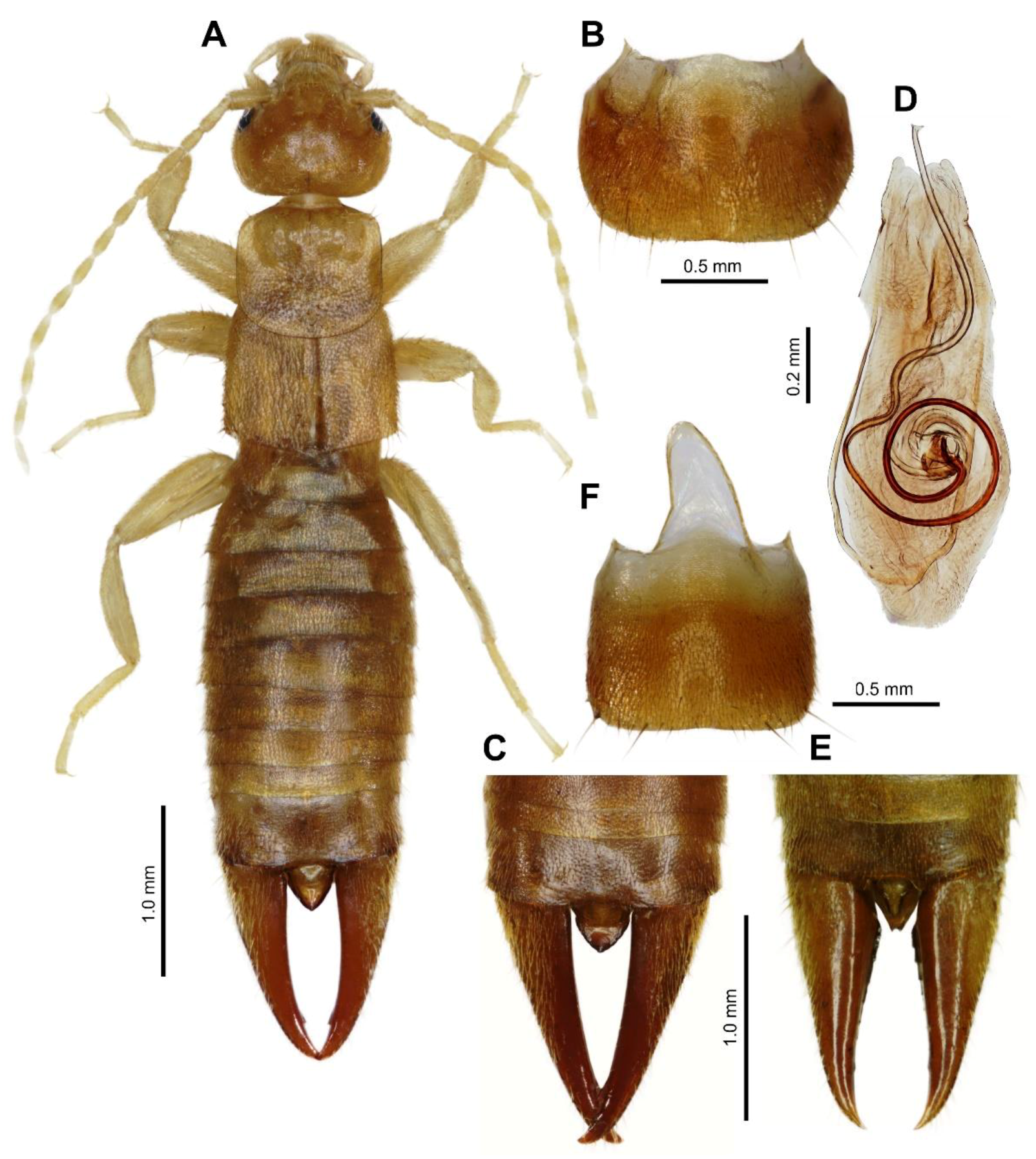
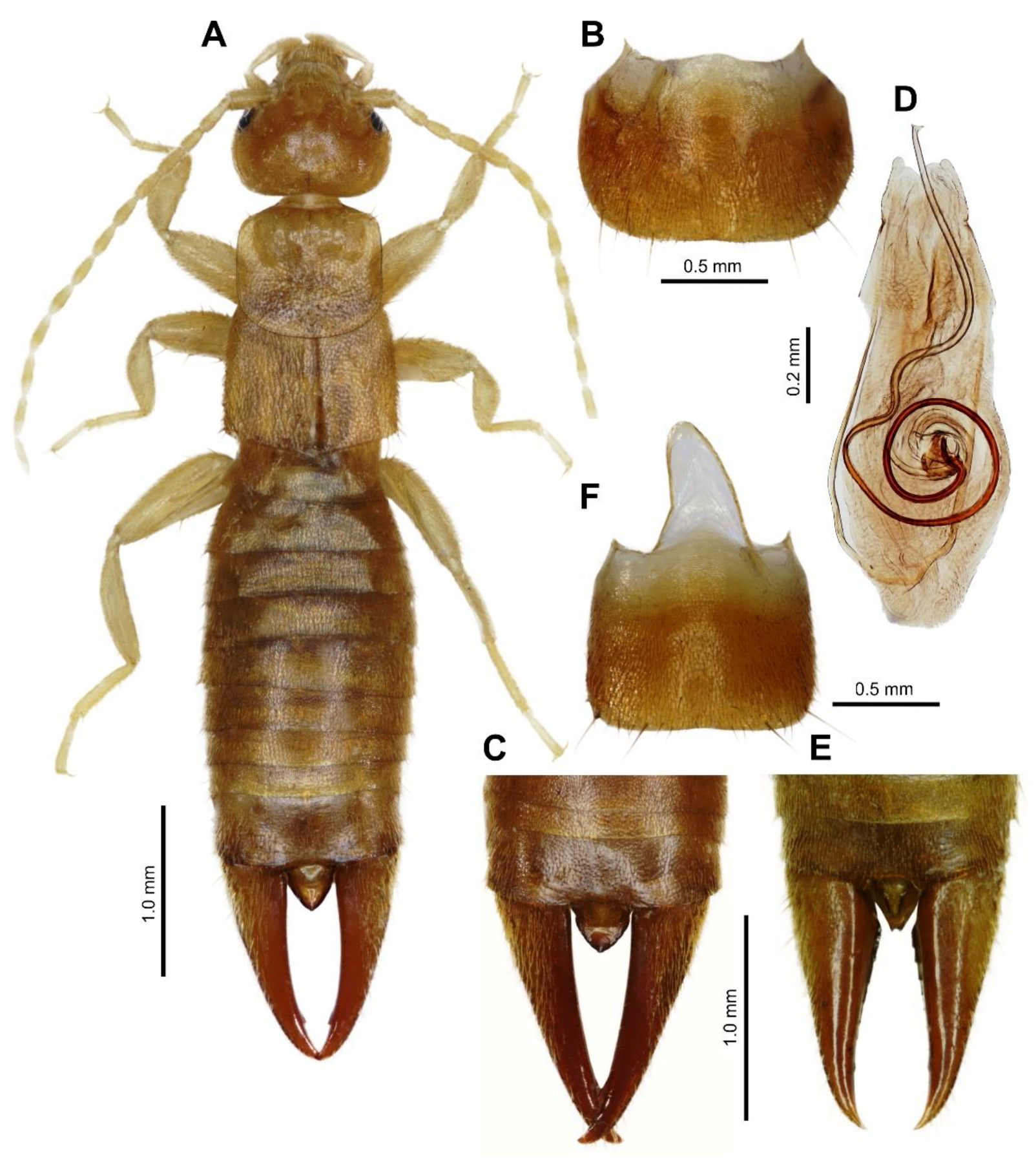
3.1. Taxonomy
3.2. Earwig–Termite Interactions
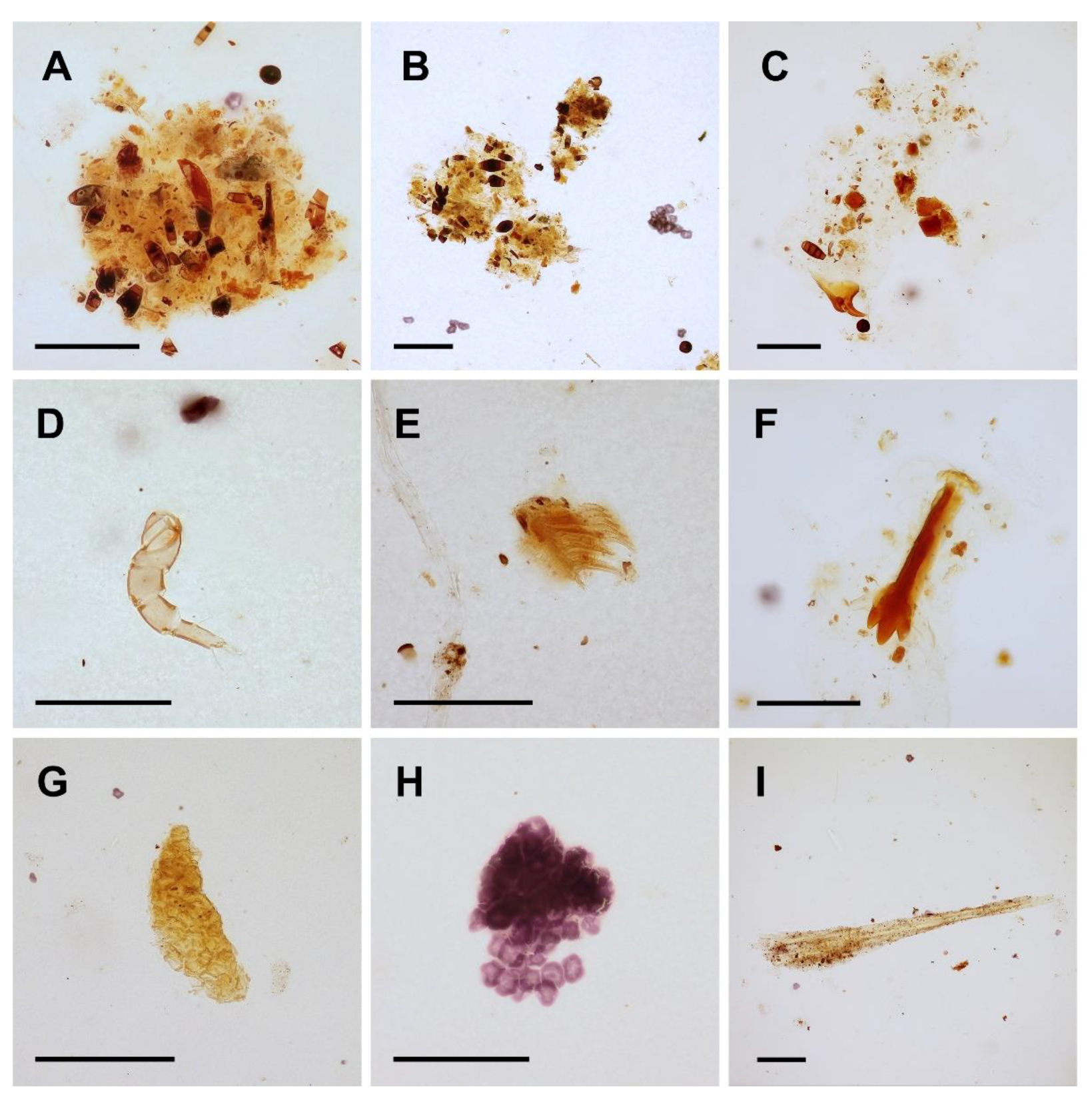
3.3. Food Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fewel, J.; Abbot, P. Sociality. In Insect Behavior: From Mechanisms to Ecological and Evolutionary Consequences; Córdoba-Aguilar, A., González-Tokman, D., González-Santoyo, I., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 219–235. [Google Scholar]
- Parmentier, T. Guests of social insects. In Encyclopedia of Social Insects; Starr, C., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 1–15. [Google Scholar]
- Korb, J. Termite Mound Architecture, from Function to Construction. In Biology of Termites: A Modern Synthesis; Bignell, D., Roisin, Y., Lo, N., Eds.; Springer: London, UK, 2010; pp. 349–373. [Google Scholar]
- Wilson, E.O. The Insect Societies; Harvard University Press: Cambridge, MA, USA, 1971; pp. 1–548. [Google Scholar]
- Kistner, D.H. Social and evolutionary significance of social insect symbionts. In Social Insects; Herman, H.R., Ed.; Academic: London, UK, 1979; Volume I, pp. 339–413. [Google Scholar]
- Kistner, D.H. The integration of foreign insects into termite societies or why do termites tolerate foreign insects in their societies? Sociobiology 1990, 17, 191–215. [Google Scholar]
- Howard, R.W.; McDaniel, C.A.; Blomquist, G.J. Chemical Mimicry as an Integrating Mechanism: Cuticular Hydrocarbons of a Termitophile and Its Host. Science 1980, 210, 431–433. [Google Scholar] [CrossRef] [PubMed]
- Costa, D.A.; de Carvalho, R.A.; de Lima Filho, G.F.; Brandao, D. Inquilines and invertebrate fauna associated with termite nests of Cornitermes cumulans (Isoptera, Termitidae) in the Emas National Park, Mineiros, Goiás, Brazil. Sociobiology 2009, 53, 443–453. [Google Scholar]
- Haas, F. Biodiversity of Dermaptera. In Insect Biodiversity: Science and Society, 1st ed.; Foottit, G.R., Adler, P.H., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2018; Volume II, pp. 315–334. [Google Scholar]
- Brindle, A. The Dermaptera of the Solomon Islands. Pac. Insects 1970, 12, 641–700. [Google Scholar]
- Steinmann, H. Dermaptera: Eudermaptera I. Tierreich 106; Gruyter: Berlin, Germany, 1990; pp. 1–558. [Google Scholar]
- Sakai, S. Dermapterorum Catalogus XXV: Iconographia IX. Explicatio Series VIIIb: Spongiphoridae Verhoeff (1902) et Kevan (1980). III; Daito Bunka University: Tokyo, Japan, 1993; pp. 4691–4820. [Google Scholar]
- Maiti, P.K. A taxonomic monograph on the world species of termites of the family Rhinotermitidae (Isoptera: Insecta). Mem. Zool. Surv. India 2006, 20, 1–272. [Google Scholar]
- Kamimura, Y. Pre- and postcopulatory sexual selection and the evolution of sexually dimorphic traits in earwigs (Dermaptera). Èntomol. Sci. 2013, 17, 139–166. [Google Scholar] [CrossRef] [Green Version]
- Kocarek, P.; Dvorak, L.; Kirstova, M. Euborellia annulipes (Dermaptera: Anisolabididae), a new alien earwig in Central Eu-ropean greenhouses: Potential pest or beneficial inhabitant? Appl. Entomol. Zool. 2015, 50, 201–206. [Google Scholar] [CrossRef]
- Anderson, L.E. Hoyer’s solution as a rapid permanent mounting medium for bryophytes. Bryologist 1954, 57, 242–244. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Srivastava, G.K. Catalogue of Oriental Dermaptera. Rec. Zool. Surv. India Occas. Pap. 1976, 2, 1–94. [Google Scholar]
- Srivastava, G.K. On the classification of Spongiphoridae (=Labiidae) with a list of Dermaptera. Rec. Zool. Surv. India 1995, 95, 71–105. [Google Scholar]
- Steinmann, H. World catalogue of Dermaptera. Ser. Entomol. 1989, 43, 1–934. [Google Scholar]
- Steinmann, H. Two new genera and species for the subfamily Labiidae (Dermaptera: Labiidae). Acta. Zool. Hung. 1987, 33, 177–186. [Google Scholar]
- Kevan, D.K.M.E.; Vickery, V.R. An annotated provisional list of non-saltatorial orthopteroid insects of Micronesia, compiled mainly from the literature. Micronesia 1997, 30, 269–353. [Google Scholar]
- Kamimura, Y.; Nishikawa, M.; Lee, C.-Y. The earwig fauna (Insecta: Dermaptera) of Penang Island, Malaysia, with descriptions of two new species. Zootaxa 2016, 4084, 233–257. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, H.; Maehr, M.D.; Haas, F.; Deem, L.S. Dermaptera Species File. Version 5.0/5.0. Available online: http://Dermaptera.SpeciesFile.org (accessed on 30 October 2021).
- Shellman-Reeve, J. The spectrum of eusociality in termites. In The Evolution of Social Behaviour in Insects and Arachnids; Choe, J.C., Crespi, B.C., Fraser, S., Eds.; Cambridge University Press: Cambridge, UK, 1997; Volume 1, pp. 52–93. [Google Scholar]
- Hugo, H.; Cristaldo, P.F.; DeSouza, O. Nonaggressive behavior: A strategy employed by an obligate nest invader to avoid conflict with its host species. Ecol. Evol. 2020, 10, 8741–8754. [Google Scholar] [CrossRef] [PubMed]
- Cristaldo, P.F.; Rodrigues, V.B.; Elliot, S.L.; Araújo, A.P.; DeSouza, O. Heterospecific detection of host alarm cues by an in-quiline termite species (Blattodea: Isoptera: Termitidae). Anim. Behav. 2016, 120, 43–49. [Google Scholar] [CrossRef]
- Noirot, C. The nests of termites. In Biology of Termites; Krishna, K., Weesner, F.M., Eds.; Academic Press: New York, NY, USA, 1970; Volume 2, pp. 311–350. [Google Scholar]
- Sherratt, T.N.; Kang, C. Anti-predator behavior. In Insect Behavior: From Mechanisms to Ecological and Evolutionary Consequences; Córdoba-Aguilar, A., González-Tokman, D., González-Santoyo, I., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 130–144. [Google Scholar]
- Prestwich, G.D. Defense mechanisms of termites. Annu. Rev. Entomol. 1984, 29, 201–232. [Google Scholar] [CrossRef]
- Šobotník, J.; Bourguignon, T.; Hanus, R.; Weyda, F.; Roisin, Y. Structure and function of defensive glands in soldiers of Glos-sotermes oculatus (Isoptera: Serritermitidae). Biol. J. Linn. Soc. 2010, 99, 839–848. [Google Scholar] [CrossRef] [Green Version]
- Kaib, M. Disruption of Ant Recruitment by the Frontal Gland Secretion of a Termite: A Chemical Defense Strategy. In The Biology of Social Insects; Breed, M.D., Michener, C.D., Evans, H.E., Eds.; CFC Press: New York, NY, USA, 2019; pp. 385–389. [Google Scholar] [CrossRef]
- Matthews, R.W.; Matthews, J.R. Insect Behavior; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1–514. [Google Scholar]
- Bignell, D.E.; Roisin, Y.; Lo, N. (Eds.) Biology of Termites: A Modern Synthesis; Springer: Berlin/Heidelberg, Germany, 2011; pp. 19–576. [Google Scholar]
- Abe, T.; Bignell, D.E.; Higashi, M.; Higashi, T. (Eds.) Termites: Evolution, Sociality, Symbioses, Ecology; Springer: Berlin/Heidelberg, Germany, 2000; pp. 1–466. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kočárek, P.; Wahab, R.A. Termitophily Documented in Earwigs (Dermaptera). Biology 2021, 10, 1243. https://doi.org/10.3390/biology10121243
Kočárek P, Wahab RA. Termitophily Documented in Earwigs (Dermaptera). Biology. 2021; 10(12):1243. https://doi.org/10.3390/biology10121243
Chicago/Turabian StyleKočárek, Petr, and Rodzay Abdul Wahab. 2021. "Termitophily Documented in Earwigs (Dermaptera)" Biology 10, no. 12: 1243. https://doi.org/10.3390/biology10121243
APA StyleKočárek, P., & Wahab, R. A. (2021). Termitophily Documented in Earwigs (Dermaptera). Biology, 10(12), 1243. https://doi.org/10.3390/biology10121243