Phytochemical Composition and In Vitro Biological Activity of Iris spp. (Iridaceae): A New Source of Bioactive Constituents for the Inhibition of Oral Bacterial Biofilms

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Preparation of Extracts
2.2. Phytochemical Analysis: Ultra-High-Performance Liquid Chromatography Coupled with High-Resolution Tandem Mass Spectrometry (UHPLC–HRMS/MS)
2.3. Antimicrobial Activity
2.4. Anti-Biofilm Activity
2.5. Anti-Quorum Sensing Activity
2.6. Cytotoxicity Assay
2.7. Data Processing and Statistical Analysis
3. Results
3.1. Phytochemical Analysis of Plant Extracts
3.2. Antimicrobial Activity
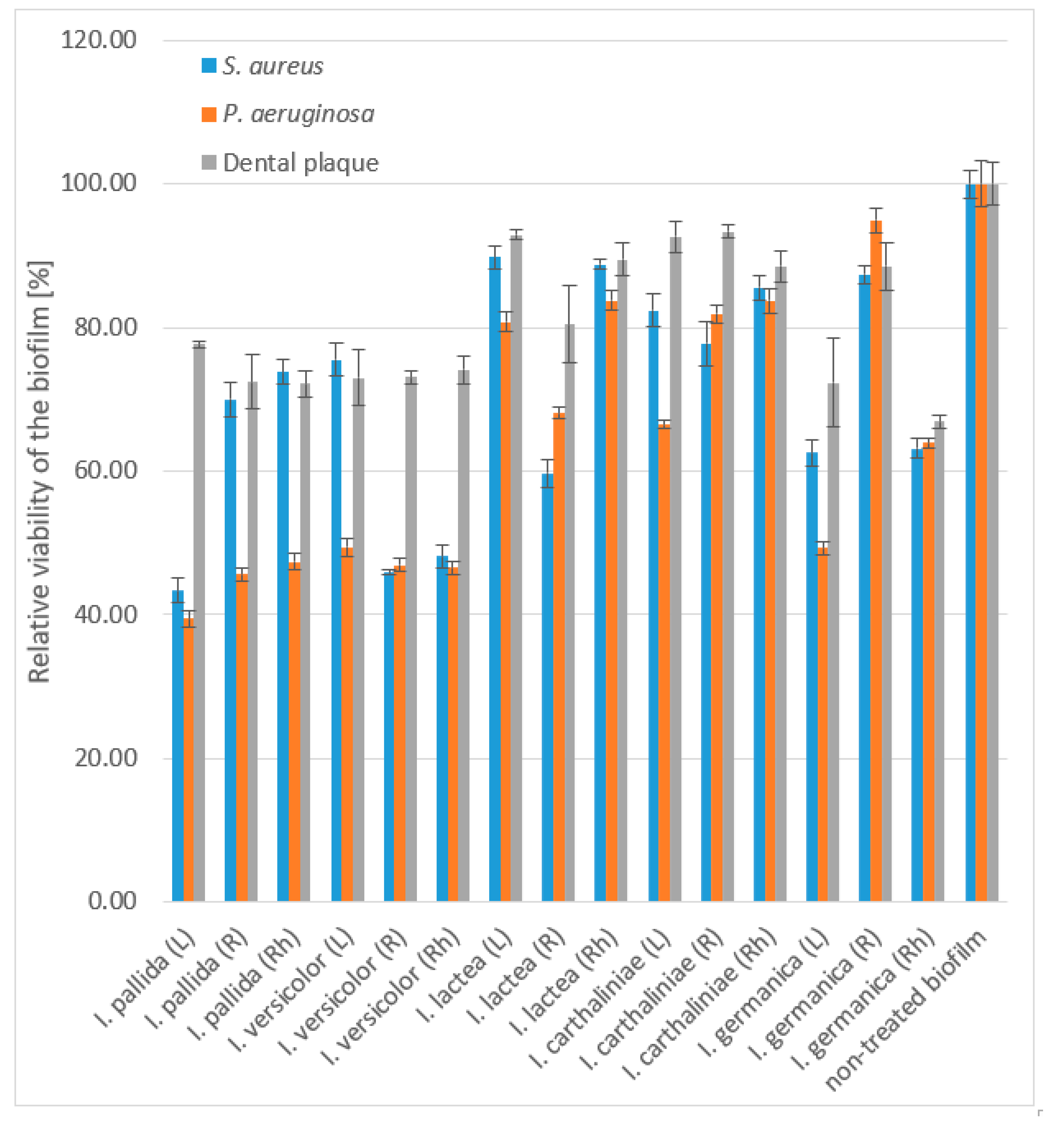
3.3. Anti-Biofilm Activity
3.4. Cell-To-Cell Communication Inhibition Assay in V. campbellii
3.5. In Vitro Cytotoxicity
3.6. Correlation of Biological Activities and Extract Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
| No. | Compound Name | Molecular Formula | Neutral Mass * | tR (min) | I. pallida. | I. versicolor | I. lactea | I. carthaliniae | I. germanica | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L | R | Rh | L | R | Rh | L | R | Rh | L | R | Rh | L | Rh | R | |||||
| Xanthones | |||||||||||||||||||
| 1 | Isomangiferin/Mangiferin/Nigricanside | C19H18O11 | 422.0844 | 1.03 | - | - | - | - | + | - | - | - | - | - | - | - | - | + | + |
| 2 | Iriflophenone | C13H10O5 | 246.0523 | 2.08 | - | - | + | + | + | + | + | + | - | + | + | - | - | - | - |
| 3 | Iriflophenone 2-O-hexoside/Iriflophenone 4-O-hexoside | C19H20O10 | 408.1039 | 2.5 | - | - | + | - | - | + | + | + | - | - | - | - | - | - | - |
| 4 | Iriflophenone | C13H10O5 | 246.0521 | 2.69 | - | - | + | + | + | + | - | + | - | + | + | - | - | - | - |
| 5 | Iriflophenone | C13H10O5 | 246.0529 | 2.87 | - | - | - | + | - | - | - | - | - | - | - | - | + | + | + |
| 6 | Iriflophenone 2-O-hexoside/Iriflophenone 4-O-hexoside | C19H20O10 | 408.1053 | 2.88 | + | + | + | + | - | + | + | - | + | + | - | - | + | + | + |
| 7 | Isomangiferin/Mangiferin/Nigricanside | C19H18O11 | 422.0845 | 2.96 | + | + | + | + | + | + | + | + | + | - | - | - | + | + | + |
| 8 | 4-O-methyliriflophenone | C14H12O5 | 260.0685 | 3.09 | - | + | - | - | - | - | - | - | + | - | - | - | - | - | + |
| 9 | 7-O-methylisomangiferin/7-O-methylmangiferin | C20H20O11 | 436.0995 | 3.24 | + | + | + | + | - | - | - | - | + | - | - | - | + | + | + |
| 10 | Iriflophenone | C13H10O5 | 246.0525 | 3.37 | - | + | + | - | + | + | - | - | + | - | - | - | - | + | + |
| 11 | Isomangiferin/Mangiferin/Nigricanside | C19H18O11 | 422.0844 | 3.39 | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - |
| 12 | Bellidifolin | C14H10O6 | 274.047 | 3.45 | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - |
| 13 | 4-O-methyliriflophenone | C14H12O5 | 260.0685 | 3.61 | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 14 | Bellidifolin | C14H10O6 | 274.047 | 3.62 | - | - | - | + | + | + | + | - | - | - | - | + | - | - | - |
| 15 | 7-O-methylisomangiferin/7-O-methylmangiferin | C20H20O11 | 436.0998 | 3.64 | - | - | - | + | + | + | - | - | - | - | - | - | - | - | - |
| 16 | 4-O-methyliriflophenone | C14H12O5 | 260.0674 | 3.91 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 17 | Isomangiferin/Mangiferin/Nigricanside | C19H18O11 | 422.0836 | 6.52 | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - |
| 18 | Bellidifolin | C14H10O6 | 274.047 | 4.21 | + | + | + | + | + | + | + | + | + | + | + | - | + | + | + |
| Phenols | |||||||||||||||||||
| 19 | Vanillic acid | C8H8O4 | 168.042 | 1.68 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 20 | trans-Cinnamic acid | C9H8O2 | 148.0526 | 2.06 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 21 | Salicylic acid/p-hydroxybenzoic acid | C7H6O3 | 138.0319 | 2.08 | - | - | + | - | + | + | + | + | - | - | - | - | - | - | - |
| 22 | Protocatechuic acid | C7H6O4 | 154.0263 | 2.17 | + | + | + | + | + | + | + | + | - | + | - | + | + | - | + |
| 23 | Ferulic acid | C10H10O4 | 194.0574 | 2.61 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 24 | Salicylic acid/p-hydroxybenzoic acid | C7H6O3 | 138.0319 | 2.69 | - | - | + | - | + | + | - | + | - | + | + | - | - | - | - |
| 25 | 3-Hydroxy-5-acetophenone/3-Hydroxy-5-methoxyacetonphenoone/Apocynin | C9H10O3 | 166.0625 | 2.91 | - | + | + | - | - | - | - | - | + | - | - | - | - | + | + |
| 26 | Salicylic acid/p-hydroxybenzoic acid | C7H6O3 | 138.0316 | 2.96 | - | - | + | - | + | + | - | + | + | + | + | + | - | - | - |
| 27 | Caffeic acid | C9H8O4 | 180.042 | 2.96 | + | - | - | + | + | - | + | + | - | - | + | + | - | - | - |
| 28 | Vanillic acid | C8H8O4 | 168.0419 | 2.98 | - | + | + | + | + | + | + | + | + | + | + | + | - | + | + |
| 29 | Vanillic acid | C8H8O4 | 168.0419 | 3.5 | - | - | + | + | - | + | + | + | - | - | - | - | - | - | - |
| 30 | Vanillic acid | C8H8O4 | 168.0419 | 3.75 | - | + | + | - | + | + | + | + | - | + | + | - | + | - | + |
| Flavonoids and isoflavonoids | |||||||||||||||||||
| 31 | 4’,7-di-O-methyldihydroquercetin-5,3,3’-trihydroxy-7,4’-dimethoxyflavanone | C17H16O7 | 332.0911 | 1.09 | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - |
| 32 | Luteolin 7-O-(2“-p- umaroyl) rhamnoside | C30H26O12 | 578.1409 | 2.71 | - | - | - | - | + | + | + | + | - | + | + | - | - | - | - |
| 33 | Swertiajaponin/Tectoridin | C22H22O11 | 462.116 | 2.85 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| 34 | Apigenin 8-C-(2“-hexosyl) hexoside/Kaempferol 7-O-(6“-rhamnosyl) hexoside | C27H30O15 | 594.1577 | 2.85 | + | - | - | - | - | - | + | - | - | - | - | + | + | - | - |
| 35 | Isorhamnetin 3-O-(2“-hamnosyl) hexoside/Isorhamnetin 3-O-(6“-rhamnosyl) hexoside/Tectorigenin-7-O-glucosyl-4′-O-glucoside | C28H32O16 | 624.169 | 2.85 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| 36 | Luteolin 7-O-(2“-p-oumaroyl)rhamnoside | C30H26O12 | 578.1413 | 2.88 | - | - | - | - | - | - | - | - | - | + | + | - | - | - | - |
| 37 | Iristectorigenin A 7-O-hexuronide/Irisdichotin A/Iristectoridin B/Iristectorin A/Iristectorin B | C23H24O12 | 492.1255 | 2.9 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| 38 | Dichotomitin 3′-O-hexoside | C24H24O12 | 520.1204 | 2.92 | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - |
| 39 | Tectorigenin-4′-O-diglucoside/Tectorigenin-7-O-diglucoside | C28H32O17 | 640.1639 | 2.97 | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - |
| 40 | Iriflophenone 4-O-(6’’-acetyl) hexoside | C21H22O11 | 450.1154 | 3.03 | - | - | + | - | - | + | - | + | - | - | - | + | - | - | - |
| 41 | Apigenin 8-C-(2“-pentosyl) hexoside/Isoschaftoside/Schaftoside | C26H28O14 | 564.1477 | 3.05 | - | + | - | + | - | - | - | - | - | - | + | - | + | + | + |
| 42 | Kaempferol 3-O-galactoside/Isoorientin/Kaempferol 3-O-glucoside/Luteolin 6-C-glucoside/Luteolin 8-C-hexoside | C21H20O11 | 448.1 | 3.1 | + | - | - | + | - | - | + | - | - | + | - | - | + | - | |
| 43 | Swertiajaponin/Tectoridin | C22H22O11 | 462.115 | 3.14 | + | - | - | - | - | - | - | - | - | + | - | - | + | - | |
| 44 | 7,4’-dimethoxy-8,3’,5’-trihydroxy-6-O-β-D-glucopyranosylisoflavone | C23H24O13 | 508.1207 | 3.16 | + | + | - | - | - | - | - | - | + | - | - | - | - | - | + |
| 45 | Iridin | C24H26O13 | 522.1371 | 3.16 | + | + | - | - | - | - | - | - | + | - | - | - | - | - | + |
| 46 | Tectorigenin-4′-O-diglucoside/Tectorigenin-7-O-diglucoside | C28H32O17 | 640.1629 | 3.17 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| 47 | Quercetin 3-O-galactoside/Quercetin 3-O-glucoside | C21H20O12 | 464.0946 | 3.18 | - | + | - | - | - | - | - | - | - | - | - | - | + | - | + |
| 48 | Apigenin 6-C-hexoside/Apigenin 8-C-glucoside/Isovitexin | C21H20O10 | 432.1055 | 3.19 | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - |
| 49 | Apigenin 8-C-(2“-pentosyl) hexoside/Isoschaftoside/Schaftoside | C26H28O14 | 564.1475 | 3.19 | - | - | - | - | - | - | - | - | - | - | + | - | - | - | - |
| 50 | Germanaism A/5,4′-Methoxy-6,7-methylenedioxy-isoflavone-3′-O-β-d-glucopyranoside | C24H24O12 | 504.1253 | 3.2 | - | - | - | - | - | - | - | - | - | - | + | + | + | - | - |
| 51 | Isorhamnetin 3-O-(2“-hamnosyl) hexoside/Isorhamnetin 3-O-(6“-rhamnosyl) hexoside/Tectorigenin-7-O-glucosyl-4′-O-glucoside | C28H32O16 | 624.1692 | 3.2 | - | - | - | - | - | - | - | - | - | - | + | + | - | + | + |
| 52 | Genistein | C15H10O5 | 270.0521 | 3.24 | - | - | - | - | - | - | - | - | - | - | + | + | - | + | + |
| 53 | Swertiajaponin/Tectoridin | C22H22O11 | 462.1159 | 3.24 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| 54 | Iristectorigenin A 7-O-hexuronide/Irisdichotin A/Iristectoridin B/Iristectorin A/Iristectorin B | C23H24O12 | 492.1268 | 3.27 | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + |
| 55 | Apigenin 8-C-(2“-hexosyl) hexoside/Kaempferol 7-O-(6“-rhamnosyl) hexoside | C27H30O15 | 594.1579 | 3.27 | - | - | - | - | - | - | - | - | - | - | - | + | - | + | + |
| 56 | Irilin D | C16H12O7 | 316.0571 | 3.29 | - | - | + | - | + | - | + | + | - | - | + | + | - | - | - |
| 57 | Iristectorigenin B/Iristectorigenin A | C17H14O7 | 330.0739 | 3.29 | - | + | - | - | - | - | + | + | + | - | - | + | - | + | + |
| 58 | 6,3’,4’-trimethoxy-7,8,5’-trihydroxyisoflavone/Irigenin | C18H16O8 | 360.0834 | 3.32 | + | + | - | - | - | - | - | - | + | - | - | - | - | - | - |
| 59 | Apigenin 6-C-hexoside/Apigenin 8-C-glucoside/Isovitexin | C21H20O10 | 432.1043 | 3.32 | - | - | - | - | - | - | + | - | - | - | - | - | + | - | - |
| 60 | Genistein | C15H10O5 | 270.0533 | 3.33 | + | + | - | - | - | - | - | - | + | - | + | + | - | + | + |
| 61 | 4’,7-di-O-methyldihydroquercetin-5,3,3’-trihydroxy-7,4’-dimethoxyflavanone | C17H16O7 | 332.0895 | 3.35 | - | - | - | - | - | - | - | - | - | - | - | - | - | + | + |
| 62 | Quercetin 3-O-galactoside/Quercetin 3-O-glucoside | C21H20O12 | 464.0948 | 3.35 | - | + | - | - | - | - | - | - | - | - | - | - | + | - | + |
| 63 | Apigenin 8-C-(2“-pentosyl) hexoside/Isoschaftoside/Schaftoside | C26H28O14 | 564.1467 | 3.35 | + | - | - | - | - | - | - | - | + | - | - | + | - | - | - |
| 64 | Irilin A/Tectorigenin | C16H12O6 | 300.0633 | 3.37 | + | + | - | - | - | - | - | - | + | - | + | + | - | + | + |
| 65 | 4′-O-Methylapigenin 6-C-hexoside/4′-O-Methylapigenin 8-C-hexoside/Swertisin | C22H22O10 | 446.1214 | 3.37 | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - |
| 66 | 3′-Hydroxyltectoridin/Isorhamnetin 3-O-galactoside/Isorhamnetin 3-O-glucoside | C22H22O12 | 478.11 | 3.43 | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - |
| 67 | Iristectorigenin B/Iristectorigenin A | C17H14O7 | 330.0731 | 3.44 | + | + | - | - | - | - | - | - | + | - | - | + | - | + | + |
| 68 | Iristectorigenin A 7-O-hexuronide/Irisdichotin A/Iristectoridin B/Iristectorin A/Iristectorin B | C23H24O12 | 492.1257 | 3.45 | - | + | - | - | - | - | - | - | + | - | - | + | - | + | + |
| 69 | Germanaism B | C23H22O11 | 474.1161 | 3.47 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 70 | Swertiajaponin/Tectoridin | C22H22O11 | 462.1155 | 3.48 | + | + | + | - | - | - | - | - | + | - | - | - | - | + | + |
| 71 | Apigenin-6,8-di-C-arabinoside | C25H26O13 | 534.1363 | 3.48 | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - |
| 72 | Apigenin 8-C-(2“-hexosyl)hexoside/Kaempferol 7-O-(6“-rhamnosyl)hexoside | C27H30O15 | 594.1575 | 3.48 | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - |
| 73 | 6,3’,4’-trimethoxy-7,8,5’-trihydroxyisoflavone/Irigenin | C18H16O8 | 360.0839 | 3.49 | + | + | + | - | - | - | - | - | + | - | - | - | + | + | + |
| 74 | Iridin | C24H26O13 | 522.1378 | 3.5 | + | + | - | - | - | - | - | - | + | - | - | - | + | + | + |
| 75 | Irilin D | C16H12O7 | 316.0583 | 3.56 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| 76 | Irilin A/Tectorigenin | C16H12O6 | 300.0634 | 3.58 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| 77 | Irilone A | C16H10O6 | 298.0478 | 3.6 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 78 | Irilone 4′-O-hexoside/Irilone 4′-O-β-d-glucopyranoside/Irilone B | C22H20O11 | 460.1007 | 3.6 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 79 | 5,3’,4’,5’-tetramethoxy-6,7-methylenedioxyisoflavone | C20H18O9 | 402.0943 | 3.62 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| 80 | Swertiajaponin/Tectoridin | C22H22O11 | 462.1161 | 3.62 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| 81 | Irilone 4′-O-(6“-hexosyl)hexoside/Irilone 4′-O-β-d-glucopyranoside-(2 → 1)-l-rhamnoside | C28H30O16 | 622.1529 | 3.62 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 82 | Germanaism B | C23H22O11 | 474.1167 | 3.67 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 83 | Germanaism A/5,4′-Methoxy-6,7-methylenedioxy-isoflavone-3′-O-β-d-glucopyranoside | C24H24O12 | 504.1251 | 3.68 | + | + | - | - | - | - | - | - | - | - | - | - | - | + | + |
| 84 | Apigenin-6,8-di-C-arabinoside | C25H26O13 | 534.1365 | 3.68 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 85 | Apigenin 8-C-(2“-pentosyl) hexoside/Isoschaftoside/Schaftoside | C26H28O14 | 564.1444 | 3.68 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 86 | 4′-O-Methylapigenin 6-C-hexoside/4′-O-Methylapigenin 8-C-hexoside/Swertisin | C22H22O10 | 446.12 | 3.74 | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - |
| 87 | 4’,7-di-O-methyldihydroquercetin-5,3,3’-trihydroxy-7,4’-dimethoxyflavanone | C17H16O7 | 332.0894 | 3.77 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 88 | Irigenin S | C19H18O8 | 374.0989 | 3.78 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 89 | Irilin B/Irisolidone | C17H14O6 | 314.0782 | 3.8 | - | - | - | - | - | - | - | - | + | - | - | - | - | + | - |
| 90 | Embinin | C29H34O14 | 606.1955 | 3.81 | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 91 | 4’-methyltectorigenin-7-glucoside/6,4’-dimethoxy-5-hydroxyflavone-7-glucoside/Irisdichotin C/Irisolidone 7-O-hexoside/Irisolidone-7-O-α-d-glucoside | C23H24O11 | 476.1302 | 3.82 | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - |
| 92 | Irilone 4′-O-[6“-(3-hydroxy-3-methylglutaryl)] hexoside | C28H28O15 | 604.1424 | 3.82 | - | + | - | - | - | - | - | - | + | - | - | - | - | - | + |
| 93 | 7-O-Methyltectorigenin 4′-O-(6“-hexosyl) hexoside | C29H34O16 | 638.1854 | 3.84 | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - |
| 94 | Irilone A | C16H10O6 | 298.0476 | 3.87 | + | + | - | - | - | - | - | - | + | - | - | - | + | + | + |
| 95 | Irifloside/Irisflogenin-4′-O-[ß-d-glucopyranosyl(1 → 6)ß-d-glucopyranoside] | C23H22O12 | 490.1096 | 3.87 | + | - | - | - | - | - | - | - | + | - | - | - | + | + | + |
| 96 | Irilone 4′-O-hexoside/Irilone 4′-O-β-d-glucopyranoside/Irilone B | C22H20O11 | 460.1006 | 3.89 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 97 | 4’-methyltectorigenin-7-glucoside/6,4’-dimethoxy-5-hydroxyflavone-7-glucoside/Irisdichotin C/Irisolidone 7-O-hexoside/Irisolidone-7-O-α-d-glucoside | C23H24O11 | 476.1315 | 4.02 | - | - | + | - | - | - | - | + | - | - | - | - | - | + | - |
| 98 | Irilin D | C16H12O7 | 316.0577 | 4.07 | - | - | + | - | + | - | + | + | - | + | + | + | - | - | - |
| 99 | 4’-methyltectorigenin-7-glucoside/6,4’-dimethoxy-5-hydroxyflavone-7-glucoside/Irisdichotin C/Irisolidone 7-O-hexoside/Irisolidone-7-O-α-d-glucoside | C23H24O11 | 476.1307 | 4.13 | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - |
| 100 | Irilin B/Irisolidone | C17H14O6 | 314.0783 | 4.15 | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - |
| 101 | 6,3’,4’-trimethoxy-7,8,5’-trihydroxyisoflavone/Irigenin | C18H16O8 | 360.084 | 4.15 | + | + | + | - | - | - | - | - | + | - | - | - | + | + | + |
| 102 | Iristectorigenin B/Iristectorigenin A | C17H14O7 | 330.0731 | 4.18 | + | + | + | - | + | - | - | + | + | - | - | + | + | + | + |
| 103 | Irisoid B/Irisoid C/Iriskashmirianin/4’,5-dimethoxy-3-hydroxy-6,7-methylenedioxyisoflavone/Nigricanin | C18H14O7 | 342.0737 | 4.25 | - | + | - | - | - | - | - | - | - | - | + | - | - | + | + |
| 104 | Iristectorigenin B/Iristectorigenin A | C17H14O7 | 330.0728 | 4.27 | - | - | - | - | - | - | - | - | - | + | - | - | - | - | - |
| 105 | Genistein | C15H10O5 | 270.0531 | 4.28 | + | + | - | - | + | - | - | - | + | - | + | - | - | + | + |
| 106 | 5-methoxy-4’-hydroxy-6,7-methylenedioxyisoflavone/Dichotomin/Nigricin | C17H12O6 | 312.0637 | 4.28 | + | + | - | - | - | - | - | - | + | - | + | - | - | + | + |
| 107 | Irilin D | C16H12O7 | 316.0582 | 4.3 | - | - | - | - | - | - | - | - | - | - | + | + | - | - | - |
| 108 | Iristectorigenin B/Iristectorigenin A | C17H14O7 | 330.0733 | 4.4 | - | - | - | - | - | - | - | - | - | - | + | + | - | - | - |
| 109 | Irilin A/Tectorigenin | C16H12O6 | 300.0627 | 4.53 | - | - | + | + | + | - | + | + | - | + | + | + | - | - | - |
| 110 | Irisflorentin a/Iriskumaonin methyl ether | C19H16O7 | 356.0893 | 4.53 | - | + | + | - | - | - | - | - | + | - | - | - | + | - | + |
| 111 | Irigenin S | C19H18O8 | 374.099 | 4.54 | + | + | - | - | - | - | - | - | + | - | - | - | - | + | + |
| 112 | Irisflorentin b | C20H18O8 | 386.1001 | 4.55 | - | + | + | - | - | - | - | - | + | - | - | - | + | + | + |
| 113 | Irilin B/Irisolidone | C17H14O6 | 314.0778 | 4.57 | + | + | - | - | - | + | - | - | + | - | - | - | - | + | + |
| 114 | dichotomitin | C18H14O8 | 358.0694 | 4.58 | + | + | - | - | - | - | - | - | + | - | - | - | + | + | + |
| 115 | Irilone A | C16H10O6 | 298.0468 | 4.71 | + | + | + | + | + | + | - | - | + | + | + | + | + | + | + |
| 116 | Irilin B/Irisolidone | C17H14O6 | 314.079 | 4.71 | - | - | - | - | + | - | - | - | - | + | - | - | - | - | - |
| 117 | Irilin B/Irisolidone | C17H14O6 | 314.0792 | 4.86 | - | - | - | - | - | - | - | - | - | - | + | + | - | - | - |
| 118 | Iristectorigenin B/Iristectorigenin A | C17H14O7 | 330.0732 | 4.88 | - | - | - | - | - | - | - | - | - | - | + | + | - | - | - |
| 119 | Irilin B/Irisolidone | C17H14O6 | 314.078 | 5.01 | - | - | + | - | - | - | + | + | - | - | + | + | - | - | - |
| 120 | Irilin B/Irisolidone | C17H14O6 | 314.0784 | 5.15 | - | - | - | - | + | - | - | - | - | - | - | - | - | + | - |
| 121 | Irilin B/Irisolidone | C17H14O6 | 314.0782 | 5.41 | - | - | + | - | - | - | - | + | - | - | + | - | - | - | - |
| 122 | Genistein | C15H10O5 | 270.0519 | 5.52 | - | - | - | - | + | - | - | + | - | - | - | - | - | - | - |
| 123 | Irilone A | C16H10O6 | 298.0471 | 5.98 | - | - | + | + | + | + | + | + | + | - | - | - | - | - | - |
| Terpenoids | |||||||||||||||||||
| 124 | Camphor | C10H16O | 152.1202 | 3.87 | - | + | - | - | + | - | - | - | - | - | + | - | - | - | + |
| 125 | 22,23-epoxy-21-hydroxyiridal/Iritectol A/Iritectol B | C30H50O5 | 490.3656 | 5.59 | - | - | - | + | - | + | - | - | - | - | - | + | + | + | - |
| 126 | 22,23-epoxy-10-deoxy-21-hydroxyiridal/22,23-epoxyiridal/Isoiridogermanal | C30H50O4 | 474.3694 | 6.28 | - | - | - | + | + | + | - | - | - | - | - | - | + | + | - |
| 127 | α-Dehydroirigermanal | C30H48O3 | 456.3604 | 6.29 | - | - | - | - | + | + | - | - | - | - | - | - | + | + | - |
| 128 | α-Dehydroirigermanal | C30H48O3 | 456.3606 | 6.51 | + | + | + | + | + | + | + | + | + | + | + | + | + | - | + |
| 129 | 22,23-epoxy-10-deoxy-21-hydroxyiridal/22,23-epoxyiridal/Isoiridogermanal | C30H50O4 | 474.3692 | 6.51 | + | + | + | + | + | + | + | + | + | + | + | + | + | - | + |
| 130 | spirobicyclic triterpenoid | C30H48O5 | 488.3487 | 6.54 | - | - | - | - | - | - | - | - | - | - | - | + | - | - | - |
| 131 | 22,23-epoxy-10-deoxy-21-hydroxyiridal/22,23-epoxyiridal/Isoiridogermanal | C30H50O4 | 474.3702 | 6.65 | - | - | - | + | - | + | - | - | - | - | - | - | + | + | - |
| 132 | Belamcandal | C32H48O6 | 528.3444 | 6.72 | + | + | - | - | + | - | + | + | - | + | + | - | + | - | - |
| 133 | α-Dehydroirigermanal | C30H48O3 | 456.3604 | 6.91 | - | - | - | + | - | + | - | - | - | - | - | - | - | - | - |
| 134 | 22,23-epoxy-10-deoxy-21-hydroxyiridal/22,23-epoxyiridal/Isoiridogermanal | C30H50O4 | 474.3706 | 6.92 | - | - | - | + | - | + | - | - | - | - | - | - | - | - | - |
| 135 | Iridotectoral A/Iridotectoral B | C30H46O5 | 486.3333 | 6.96 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | - |
| 136 | Belamcandal | C32H48O6 | 528.3434 | 6.98 | + | + | - | - | + | - | + | + | + | + | + | - | + | - | - |
| 137 | Belamcandal | C32H48O6 | 528.344 | 7.27 | + | + | + | + | + | + | + | + | + | + | + | + | + | - | + |
| 138 | α-Dehydroirigermanal | C30H48O3 | 456.3601 | 7.34 | - | + | - | - | - | + | - | - | + | - | - | - | - | - | + |
| 139 | spirobicyclic triterpenoid | C30H48O5 | 488.3486 | 7.51 | - | - | + | + | + | + | + | - | - | - | - | - | - | - | - |
| 140 | Iridal/α-irigermanal | C30H50O4 | 458.3739 | 7.52 | - | + | - | + | + | + | - | - | + | - | - | - | - | + | + |
| 141 | Iridotectoral A/Iridotectoral B | C30H46O5 | 486.3332 | 7.56 | - | - | - | + | - | + | - | - | - | + | + | + | - | - | - |
| 142 | Iridotectoral A/Iridotectoral B | C30H46O5 | 486.3336 | 7.94 | - | - | + | + | + | + | + | - | - | - | - | - | - | - | - |
| 143 | 22,23-epoxy-10-deoxy-21-hydroxyiridal/22,23-epoxyiridal/Isoiridogermanal | C30H46O5 | 474.3696 | 7.97 | - | - | - | + | - | + | - | - | - | - | - | - | - | + | - |
| 144 | Amorphene/α-Muurolene/β-Gurjuenene/γ-Elemene | C15H24 | 204.1866 | 8.57 | + | + | - | + | + | - | + | - | + | + | - | - | + | + | + |
| 145 | α-Dehydroirigermanal | C30H48O3 | 456.36 | 8.86 | - | - | - | + | - | + | - | - | - | - | - | - | - | + | - |
| 146 | Amorphene/α-Muurolene/β-Gurjuenene/γ-Elemene | C15H24 | 204.1877 | 9.26 | - | - | - | + | - | + | - | - | - | - | - | + | - | - | - |
| 147 | Iridal/α-irigermanal | C30H50O3 | 458.3756 | 9.26 | + | - | + | + | + | + | + | + | + | + | + | + | - | - | + |
| 148 | iriversical | C31H52O3 | 472.3899 | 9.75 | + | - | - | - | - | - | - | - | - | - | + | + | + | - | - |
| 149 | iriversical | C31H52O3 | 472.3898 | 10.03 | - | - | - | - | - | - | - | - | + | - | - | - | - | + | - |
| 150 | Palmitic acid | C16H32O2 | 256.2397 | 10.7 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 151 | Iridotectoral A/Iridotectoral B | C30H46O5 | 486.3347 | 10.9 | - | + | - | - | + | - | - | + | - | - | + | - | - | - | - |
| 152 | Aristolone | C15H22O | 218.1669 | 11.02 | + | - | - | - | - | - | - | - | - | - | - | + | + | + | - |
| 153 | α-Dehydroirigermanal | C30H48O3 | 456.359 | 11.29 | - | - | - | - | + | + | - | - | - | - | - | - | - | - | - |
| 154 | α-Dehydroirigermanal | C30H48O3 | 456.3594 | 11.49 | - | + | - | - | - | + | - | + | - | - | + | + | - | + | + |
| 155 | Iridial | C29H48O3 | 444.3602 | 12.19 | + | - | - | + | - | - | + | - | - | + | - | - | + | - | - |
| 156 | iriversical | C31H52O3 | 472.3921 | 13.5 | - | - | - | - | - | + | - | - | - | - | - | - | - | - | - |
| 157 | Irisgermanic C | C28H48O3 | 432.36 | 13.55 | + | - | - | - | - | - | + | - | - | - | - | - | + | - | - |
| 158 | Iridial | C29H48O3 | 444.3599 | 13.55 | + | - | - | - | - | - | - | - | - | + | - | - | + | - | - |
| 159 | 22,23-epoxy-10-deoxy-21-hydroxyiridal/22,23-epoxyiridal/Isoiridogermanal | C30H50O4 | 474.3693 | 13.46 | - | + | - | - | + | - | - | + | - | - | - | - | - | + | + |
| 160 | Iridal/α-irigermanal | C30H50O3 | 458.3731 | 13.62 | + | - | + | + | - | - | + | - | - | + | - | - | + | - | - |
| 161 | Amorphene/α-Muurolene/β-Gurjuenene/γ-Elemene | C15H24 | 204.19 | 14.01 | - | - | - | - | - | - | - | - | + | - | - | - | - | + | - |
| Fatty acids | |||||||||||||||||||
| 162 | Myristic acid | C14H28O2 | 228.2078 | 4.98 | + | - | - | - | - | - | + | - | - | + | - | - | + | - | - |
| 163 | Lauric acid | C12H24O2 | 200.1766 | 6.23 | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - |
| 164 | Myristic acid | C14H28O2 | 228.2079 | 7.92 | + | + | - | - | - | - | - | - | + | - | - | - | + | + | + |
| 165 | Lauric acid | C12H24O2 | 200.1771 | 8.08 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 166 | Myristic acid | C14H28O2 | 228.2084 | 9.36 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 167 | Lauric acid | C12H24O2 | 200.1769 | 11.28 | - | - | - | + | + | + | - | + | + | - | - | - | + | + | - |
| 168 | Stearic acid | C18H36O2 | 284.2706 | 11.87 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 169 | Myristic acid | C14H28O2 | 228.2081 | 12.13 | + | - | - | - | - | - | - | - | - | - | - | - | + | + | - |
| 170 | Myristic acid | C14H28O2 | 228.208 | 12.38 | + | - | - | - | - | - | - | - | + | - | - | + | + | + | - |
| 171 | Palmitic acid | C16H32O2 | 256.2392 | 13.46 | - | - | + | + | + | + | + | - | - | - | - | + | - | - | - |
| 172 | Stearic acid | C18H36O2 | 284.2704 | 13.76 | - | - | + | + | + | + | + | - | - | + | - | + | - | - | - |
| Steroids | |||||||||||||||||||
| 173 | Stigmasterol | C29H48O | 412.3688 | 9.73 | - | - | - | - | - | - | - | - | - | - | + | + | - | - | - |
| 174 | Stigmasterol-3-O-b-D-glucopyranoside | C35H58O6 | 574.422 | 11.91 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 175 | Stigmasterol-3-O-b-D-glucopyranoside | C35H58O6 | 574.4219 | 12.32 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 176 | 7-β-Hydroxystigmast-4-en-3-one | C29H48O2 | 428.3655 | 12.65 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + |
| 177 | Stigmasterol | C35H58O6 | 412.3701 | 13.48 | + | + | - | - | + | - | - | + | + | + | + | - | + | - | + |
| Quinones | |||||||||||||||||||
| 178 | Irisoquin B | C24H40O4 | 392.294 | 8.4 | - | - | - | - | - | - | + | + | - | + | - | - | + | - | - |
| 179 | Irisquinone A | C24H38O3 | 374.282 | 11.32 | + | - | - | - | - | - | + | - | - | - | - | - | + | - | - |
| 180 | Pallasone B Dihydroirisquinone | C24H40O3 | 376.2976 | 12.26 | + | - | - | - | - | - | + | - | - | - | - | - | + | - | - |
References
- Lee, D.; Seo, Y.; Khan, M.S.; Hwang, J.; Jo, Y.; Son, J.; Lee, K.; Park, C.; Chavan, S.; Gilad, A.A.; et al. Use of nanoscale materials for the effective prevention and extermination of bacterial biofilms. Biotechnol. Bioprocess Eng. 2018, 23, 1–10. [Google Scholar] [CrossRef]
- Yu, O.Y.; Zhao, I.S.; Mei, M.L.; Lo, E.C.; Chu, C.H. Dental Biofilm and Laboratory Microbial Culture Models for Cariology Research. Dent. J. 2017, 5, 21. [Google Scholar] [CrossRef]
- Basavaraju, M.; Sisnity, V.S.; Palaparthy, R.; Addanki, P.K. Quorum quenching: signal jamming in dental plaque biofilms. J. Dent. Sci. 2016, 11, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Kolenbrander, P.E.; Palmer, R.J.; Periasamy, S.; Jakubovics, N.S. Oral multispecies biofilm development and the key role of cell-cell distance. Nat. Rev. Microbiol. 2010, 8, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Zuanazzi, D.; Souto, R.; Mattos, M.B.A.; Zuanazzi, M.R.; Tura, B.R.; Sansone, C.; Colombo, A.P.V. Prevalence of potential bacterial respiratory pathogens in the oral cavity of hospitalised individuals. Arch. Oral Biol. 2010, 55, 21–28. [Google Scholar] [CrossRef]
- Da Silva-Boghossian, C.M.I.; Do Souto, R.M.; Luiz, R.R.; Colombo, A.P.V. Association of red complex, A. Actinomycetemcomitans and non-oral bacteria with periodontal diseases. Arch. Oral Biol. 2011, 56, 899–906. [Google Scholar] [CrossRef]
- Al-Jumaili, A.; Kumar, A.; Bazaka, K.; Jacob, M.V. Plant secondary metabolite-derived polymers: a potential approach to develop antimicrobial films. Polymers 2018, 10, 515. [Google Scholar] [CrossRef]
- Furiga, A.; Roques, C.; Badet, C. Preventive effects of an original combination of grape seed polyphenols with amine fluoride on dental biofilm formation and oxidative damage by oral bacteria. J. Appl. Microbiol. 2014, 116, 761–771. [Google Scholar] [CrossRef]
- Furiga, A.; Lonvaud-Funel, A.; Dorignac, G.; Badet, C. In vitro anti-bacterial and anti-adherence effects of natural polyphenolic compounds on oral bacteria. J. Appl. Microbiol. 2008, 105, 1470–1476. [Google Scholar] [CrossRef]
- Antonio, A.G.; Iorio, N.L.P.; Pierro, V.S.S.; Candreva, M.S.; Farah, A.; Dos Santos, K.R.N.; Maia, L.C. Inhibitory properties of Coffea canephora extract against oral bacteria and its effect on demineralisation of deciduous teeth. Arch. Oral Biol. 2011, 56, 556–564. [Google Scholar] [CrossRef]
- Antonio, A.; Iorio, N.P.; Farah, A.; Dos Santos, K.N.; Maia, L. Effect of Coffea canephora aqueous extract on microbial counts in ex vivo oral biofilms: a case study. Planta Med. 2012, 78, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Gartenmann, S.J.; Steppacher, S.L.; Von Weydlich, Y.; Heumann, C.; Attin, T.; Schmidlin, P.R. The effect of green tea on plaque and gingival inflammation: a systematic review. J. Herb. Med. 2020, 21, 100337. [Google Scholar] [CrossRef]
- Moon, K.H.; Lee, Y.; Kim, J.N. Effects of foreign plant extracts on cell growth and biofilm formation of Streptococcus mutans. J. Life Sci. 2019, 29, 712–723. [Google Scholar] [CrossRef]
- Burcu, B.; Aysel, U.; Nurdan, S. Antimicrobial, antioxidant, antimutagenic activities, and phenolic compounds of Iris germanica. Ind. Crop. Prod. 2014, 61, 526–530. [Google Scholar] [CrossRef]
- Fang, R.; Houghton, P.J.; Hylands, P.J. Cytotoxic effects of compounds from Iris tectorum on human cancer cell lines. J. Ethnopharmacol. 2008, 118, 257–263. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, G.A.; Al-Musayeib, N.M. New constituents from the rhizomes of Egyptian Iris germanica L. Molecules 2012, 17, 2587–2598. [Google Scholar] [CrossRef]
- Kostić, A.; Gašić, U.M.; Pešić, M.B.; Stanojević, S.P.; Barać, M.B.; Mačukanović-Jocić, M.P.; Avramov, S.N.; Tešić, Ž.L. Phytochemical analysis and total antioxidant capacity of rhizome, above-ground vegetative parts and flower of three Iris species. Chem. Biodivers. 2019, 16, e1800565. [Google Scholar] [CrossRef]
- Moaket, S.; Oguzkan, S.B.; Kilic, I.H.; Selvi, B.; Karagoz, I.D.; Erdem, M.; Erdoğan, N.; Tekin, H.; Ozaslan, M. Biological activity of Iris sari Schott ex Baker in Turkey. J. Biol. Sci. 2017, 17, 136–141. [Google Scholar] [CrossRef][Green Version]
- Mocan, A.; Zengin, G.; Mollica, A.; Uysal, A.; Gunes, E.; Crişan, G.; Aktumsek, A. Biological effects and chemical characterization of Iris schachtii Markgr extracts: a new source of bioactive constituents. Food Chem. Toxicol. 2018, 112, 448–457. [Google Scholar] [CrossRef]
- Nadaroğlu, H.; Demir, Y.; Demir, N. Antioxidant and radical scavenging properties of Iris germanica. Pharm. Chem. J. 2007, 41, 409–415. [Google Scholar] [CrossRef]
- Singab, A.N.B.; Ayoub, I.M.; El-Shazly, M.; Korinek, M.; Wu, T.Y.; Cheng, Y.B.; Chang, F.R.; Wu, Y.C. Shedding the light on iridaceae: ethnobotany, phytochemistry and biological activity. Ind. Crop. Prod. 2016, 92, 308–335. [Google Scholar] [CrossRef]
- Xie, G.Y.; Qin, X.Y.; Liu, R.; Wang, Q.; Lin, B.B.; Wang, G.K.; Xu, G.K.; Wen, R.; Qin, M.J. New isoflavones with cytotoxic activity from the rhizomes of Iris germanica L. Nat. Prod. Res. 2013, 27, 2173–2177. [Google Scholar] [CrossRef] [PubMed]
- Kaššák, P. Secondary metabolites of the chosen genus Iris species. Acta Univ. Agric. Silvic. Mendel. Brun. 2013, 60, 269–280. [Google Scholar] [CrossRef]
- Viktorova, J.; Stranska-Zachariasova, M.; Fenclova, M.; Vitek, L.; Hajslova, J.; Kren, V.; Ruml, T. Complex evaluation of antioxidant capacity of milk thistle dietary supplements. Antioxidants 2019, 8, 317. [Google Scholar] [CrossRef] [PubMed]
- Pogačnik, L.; Bergant, T.; Skrt, M.; Ulrih, N.P.; Viktorová, J.; Ruml, T. In Vitro Comparison of the Bioactivities of Japanese and Bohemian Knotweed Ethanol Extracts. Foods 2020, 9, 544. [Google Scholar] [CrossRef] [PubMed]
- Viktorová, J.; Stupák, M.; Řehořová, K.; Dobiasová, S.; Hoang, L.; Hajšlová, J.; Van Thanh, T.; Van Tri, L.; Van Tuan, N.; Ruml, T. Lemon grass essential oil does not modulate cancer cells multidrug resistance by citral—its dominant and strongly antimicrobial compound. Foods 2020, 9, 585. [Google Scholar] [CrossRef]
- Sandberg, M.E.; Schellmann, D.; Brunhofer, G.; Erker, T.; Busygin, I.; Leino, R.; Vuorela, P.M.; Fallarero, A. Pros and cons of using resazurin staining for quantification of viable Staphylococcus aureus biofilms in a screening assay. J. Microbiol. Methods 2009, 78, 104–106. [Google Scholar] [CrossRef]
- Tran, V.N.; Viktorova, J.; Augustynkova, K.; Jelenova, N.; Dobiasova, S.; Rehorova, K.; Fenclova, M.; Stranska-Zachariasova, M.; Vitek, L.; Hajslova, J.; et al. In silico and in vitro studies of mycotoxins and their cocktails; Their toxicity and its mitigation by silibinin pre-treatment. Toxins 2020, 12, 148. [Google Scholar] [CrossRef]
- Kukula-Koch, W.; Sieniawska, E.; Widelski, J.; Urjin, O.; Głowniak, P.; Skalicka-Woźniak, K. Major secondary metabolites of Iris spp. Phytochem. Rev. 2015, 14, 51–80. [Google Scholar] [CrossRef]
- Roger, B.; Jeannot, V.; Fernandez, X.; Cerantola, S.; Chahboun, J. Characterisation and quantification of flavonoids in Iris germanica L. and Iris pallida Lam. resinoids from Morocco. Phytochem. Anal. 2012, 23, 450–455. [Google Scholar] [CrossRef]
- Nasim, S.; Baig, I.; Jalil, S.; Orhan, I.; Sener, B.; Choudhary, M.I. Anti-inflammatory isoflavonoids from the rhizomes of Iris germanica. J. Ethnopharmacol. 2003, 86, 177–180. [Google Scholar] [CrossRef]
- Nasim, S.; Baig, I.; Jalil, S.; Orhan, I.; Sener, B.; Choudhary, M.I. Isoflavonoid glycosides from the rhizomes of Iris germanica. Chem. Pharm. Bull. 2002, 50, 1100–1102. [Google Scholar] [CrossRef]
- Rigano, D.; Formisano, C.; Grassia, A.; Grassia, G.; Perrone, A.; Piacente, S.; Vuotto, M.L.; Senatore, F. Antioxidant flavonoids and isoflavonoids from rhizomes of Iris pseudopumila. Planta Med. 2007, 73, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Krick, W.; Marner, F.-J.; Jaenicke, L. Isolation and structural determination of a new methylated triterpenoid from rhizomes of Iris versicolor L. Zeitschrift fur Naturforsch C J. Biosci. 1983, 38, 689–692. [Google Scholar] [CrossRef]
- Marner, F.J.; Longerich, I. Isolation and structure determination of new iridals from Iris sibirica and Iris versicolor. Liebigs Ann. Chem. 1992, 269–272. [Google Scholar] [CrossRef]
- Ouyang, J.; Sun, F.; Feng, W.; Sun, Y.; Qiu, X.; Xiong, L.; Liu, Y.; Chen, Y. Quercetin is an effective inhibitor of quorum sensing, biofilm formation and virulence factors in Pseudomonas aeruginosa. J. Appl. Microbiol. 2016, 120, 966–974. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, J.H.; Cho, H.S.; Joo, S.W.; Cho, M.H.; Lee, J. Anti-biofilm activities of quercetin and tannic acid against Staphylococcus aureus. Biofouling 2013, 29, 491–499. [Google Scholar] [CrossRef]
- Wang, J.; Song, M.; Pan, J.; Shen, X.; Liu, W.; Zhang, X.; Li, H.; Deng, X. Quercetin impairs Streptococcus pneumoniae biofilm formation by inhibiting sortase A activity. J. Cell. Mol. Med. 2018, 22, 6228–6237. [Google Scholar] [CrossRef]
- Zeng, Y.; Nikitkova, A.; Abdelsalam, H.; Li, J.; Xiao, J. Activity of quercetin and kaemferol against Streptococcus mutans biofilm. Arch. Oral Biol. 2019, 98, 9–16. [Google Scholar] [CrossRef]
- Yang, W.Y.; Kim, C.K.; Ahn, C.H.; Kim, H.; Shin, J.; Oh, K.B. Flavonoid glycosides inhibit sortase A and sortase A-mediated aggregation of Streptococcus mutans, an oral bacterium responsible for human dental caries. J. Microbiol. Biotechnol. 2016, 26, 1557–1565. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, X.; Wang, L.; Chen, S.; Shi, D.; Wang, H. Molecular mechanism of the flavonoid natural product dryocrassin ABBA against Staphylococcus aureus sortase A. Molecules 2016, 21, 1428. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.; Xiang, H.; Dong, H.; Wang, D.; Wang, T. Isovitexin, a potential candidate inhibitor of sortase A of Staphylococcus aureus USA300. J. Microbiol. Biotechnol. 2018, 28, 1426–1432. [Google Scholar] [CrossRef] [PubMed]
- Slobodníková, L.; Fialová, S.; Rendeková, K.; Kováč, J.; Mučaji, P. Antibiofilm activity of plant polyphenols. Molecules 2016, 21, 1717. [Google Scholar] [CrossRef] [PubMed]
- Pande, G.S.J.; Natrah, F.M.I.; Sorgeloos, P.; Bossier, P.; Defoirdt, T. The Vibrio campbellii quorum sensing signals have a different impact on virulence of the bacterium towards different crustacean hosts. Vet. Microbiol. 2013, 167, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Asfour, H. Anti-quorum sensing natural compounds. J. Microsc. Ultrastruct. 2018, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gomes, L.C.; Moreira, J.M.; Araújo, J.D.; Mergulhão, F.J. Surface conditioning with Escherichia coli cell wall components can reduce biofilm formation by decreasing initial adhesion. AIMS Microbiol. 2017, 3, 613–628. [Google Scholar] [CrossRef]
- Abd-Alla, M.H.; Bashandy, S.R. Production of quorum sensing inhibitors in growing onion bulbs infected with Pseudomonas aeruginosa E (HQ324110). ISRN Microbiol. 2012, 2012, 161890. [Google Scholar] [CrossRef][Green Version]
- Prasath, K.G.; Sethupathy, S.; Pandian, S.K. Proteomic analysis uncovers the modulation of ergosterol, sphingolipid and oxidative stress pathway by myristic acid impeding biofilm and virulence in Candida albicans. J. Proteom. 2019, 208, 103503. [Google Scholar] [CrossRef]
- Duckworth, R.M.; Maguire, A.; Omid, N.; Steen, I.N.; McCracken, G.I.; Zohoori, F.V. Effect of rinsing with mouthwashes after brushing with a fluoridated toothpaste on salivary fluoride concentration. Caries Res. 2009, 43, 391–396. [Google Scholar] [CrossRef]

| Species | Number of Detected Compounds (Tentative Identity) | |||||||
|---|---|---|---|---|---|---|---|---|
| (Iso)flavonoids | Phenols | Fatty Acids | Terpenoids | Steroids | Xanthones | Quinones | All | |
| I. pallida (L) | 35 | 5 | 8 | 14 | 4 | 5 | 2 | 73 |
| I. pallida (R) | 38 | 7 | 6 | 13 | 3 | 8 | - | 75 |
| I. pallida (Rh) | 15 | 11 | 6 | 8 | 3 | 9 | - | 52 |
| I. versicolor (L) | 8 | 7 | 7 | 20 | 3 | 13 | - | 58 |
| I. versicolor (R) | 11 | 10 | 7 | 17 | 4 | 9 | - | 58 |
| I. versicolor (Rh) | 5 | 10 | 7 | 22 | 3 | 10 | - | 57 |
| I. lactea (L) | 10 | 9 | 7 | 13 | 3 | 7 | 3 | 52 |
| I. lactea (R) | 12 | 11 | 5 | 10 | 4 | 7 | 1 | 50 |
| I. lactea (Rh) | 37 | 6 | 7 | 12 | 4 | 7 | - | 73 |
| I. carthaliniae (L) | 11 | 8 | 6 | 12 | 4 | 5 | 1 | 47 |
| I. carthaliniae (R) | 23 | 8 | 4 | 13 | 5 | 4 | - | 57 |
| I. carthaliniae (Rh) | 31 | 7 | 7 | 12 | 4 | 2 | - | 63 |
| I. germanica (L) | 19 | 5 | 9 | 17 | 4 | 6 | 3 | 63 |
| I. germanica (R) | 46 | 5 | 8 | 15 | 3 | 8 | - | 85 |
| I. germanica (Rh) | 43 | 7 | 5 | 10 | 4 | 9 | - | 78 |
| species | Anti-AdhesionIC50 [mg/L] | AntibiofilmIC50 [mg/L] | ||||
|---|---|---|---|---|---|---|
| S. aureus | P. aeruginosa | Dental Plaque | S. aureus | P. aeruginosa | Dental Plaque | |
| I. pallida (L) | 139.4 ± 2.0 | 88.5 ± 1.6 | 163.5 ± 6.5 | 406.3 ± 20.4 | 287.0 ± 10.3 | >666.7 |
| I. pallida (R) | 177.2 ± 13.0 | 122.7 ± 4.0 | 326.9 ± 13.5 | >666.7 | 447.6 ± 45.9 | >666.7 |
| I. pallida (Rh) | 334.5 ± 8.1 | 297.2 ± 23.1 | 556.6 ± 48.3 | >666.7 | 628.2 ± 13.9 | >666.7 |
| I. versicolor (L) | 169.0 ± 6.1 | 132.2 ± 14.8 | 357.0 ± 13.0 | >666.7 | 497.2 ± 22.2 | >666.7 |
| I. versicolor (R) | 161.0 ± 5.5 | 177.0 ± 4.3 | 315.3 ± 38.6 | 549.6 ± 8.3 | 526.1 ± 15.9 | >666.7 |
| I. versicolor (Rh) | 98.5 ± 9.5 | 112.4 ± 5.2 | 201.6 ± 12.4 | 615.6 ± 28.9 | 500.4 ± 16.4 | >666.7 |
| I. lactea (L) | >666.7 | >666.7 | >666.7 | >666.7 | >666.7 | >666.7 |
| I. lactea (R) | 255.3 ± 26.0 | 393.4 ± 17.0 | 542.8 ± 46.3 | >666.7 | >666.7 | >666.7 |
| I. lactea (Rh) | 370.5 ± 22.6 | >666.7 | >666.7 | >666.7 | >666.7 | >666.7 |
| I. carthaliniae (L) | 494.5 ± 69.1 | 512.8 ± 14.6 | >666.7 | >666.7 | >666.7 | >666.7 |
| I. carthaliniae (R) | 427.8 ± 19.1 | >666.7 | >666.7 | >666.7 | >666.7 | >666.7 |
| I. carthaliniae (Rh) | >666.7 | >666.7 | >666.7 | >666.7 | >666.7 | >666.7 |
| I. germanica (L) | 178.1 ± 23.2 | 267.5 ± 8.6 | 357.4 ± 16.3 | >666.7 | 513.0 ± 56.1 | >666.7 |
| I. germanica (R) | 334.6 ± 8.4 | >666.7 | >666.7 | >666.7 | >666.7 | >666.7 |
| I. germanica (Rh) | 258.1 ± 10.0 | 630.7 ± 19.0 | 542.0 ± 23.6 | >666.7 | >666.7 | >666.7 |
| V. campbellii BAA1118 | V. campbellii BAA1119 | |
|---|---|---|
| species | QS IC50 [mg/L] | QS IC50 [mg/L] |
| I. pallida (L) | 533.8 ± 3.2 | 605.0 ± 3.0 |
| I. pallida (R) | 560.6 ± 10.4 | >666.7 |
| I. pallida (Rh) | >666.7 | >666.7 |
| I. versicolor (L) | 543.4 ± 59.5 | >666.7 |
| I. versicolor (R) | 542.0 ± 1.8 | 644.0 ± 10.0 |
| I. versicolor (Rh) | 638.5 ± 16.2 | 597.2 ± 33.2 |
| I. lactea (L) | >666.7 | >666.7 |
| I. lactea (R) | >666.7 | >666.7 |
| I. lactea (Rh) | >666.7 | >666.7 |
| I. carthaliniae (L) | >666.7 | >666.7 |
| I. carthaliniae (R) | >666.7 | >666.7 |
| I. carthaliniae (Rh) | >666.7 | >666.7 |
| I. germanica (L) | 297.1 ± 10.3 | >666.7 |
| I. germanica (R) | >666.7 | >666.7 |
| I. germanica (Rh) | >666.7 | >666.7 |
| Erythromycin | 20.7 ± 1.0 | 2.6 ± 0.1 |
| Iris part | IC50 [mg/L] |
|---|---|
| I. pallida (L) | >666.7 |
| I. pallida (R) | >666.7 |
| I. pallida (Rh) | >666.7 |
| I. versicolor (L) | >666.7 |
| I. versicolor (R) | >666.7 |
| I. versicolor (Rh) | 96.0 ± 6.7 |
| I. lacteal (L) | >666.7 |
| I. lacteal (R) | >666.7 |
| I. lacteal (Rh) | >666.7 |
| I. carthaliniae (L) | >666.7 |
| I. carthaliniae (R) | >666.7 |
| I. carthaliniae (Rh) | 317.3 ± 40.0 |
| I. germanica (L) | >666.7 |
| I. germanica (R) | >666.7 |
| I. germanica (Rh) | >666.7 |
| Doxorubicin | 0.4 ± 0.007 |
| Compound | Inhibition of Dental Biofilm Adhesion | Inhibition of BA1118 Communication | ||||
|---|---|---|---|---|---|---|
| df | Rcrit | R2 | df | Rcrit | R2 | |
| Myristic acid | 7 | 0.666 | 0.753 | |||
| Germanaism B | 1 | 0.997 | 0.999 | |||
| 7-β-hydroxystigmast-4-en-3-one | 4 | 0.811 | 0.933 | |||
| Amorphene/α-muurolene/β-gurjuenene/γ-elemene | 3 | 0.878 | 0.932 | |||
| Isomangiferin/Mangiferin/Nigricanside | 4 | 0.811 | 0.827 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoang, L.; Beneš, F.; Fenclová, M.; Kronusová, O.; Švarcová, V.; Řehořová, K.; Baldassarre Švecová, E.; Vosátka, M.; Hajšlová, J.; Kaštánek, P.; et al. Phytochemical Composition and In Vitro Biological Activity of Iris spp. (Iridaceae): A New Source of Bioactive Constituents for the Inhibition of Oral Bacterial Biofilms. Antibiotics 2020, 9, 403. https://doi.org/10.3390/antibiotics9070403
Hoang L, Beneš F, Fenclová M, Kronusová O, Švarcová V, Řehořová K, Baldassarre Švecová E, Vosátka M, Hajšlová J, Kaštánek P, et al. Phytochemical Composition and In Vitro Biological Activity of Iris spp. (Iridaceae): A New Source of Bioactive Constituents for the Inhibition of Oral Bacterial Biofilms. Antibiotics. 2020; 9(7):403. https://doi.org/10.3390/antibiotics9070403
Chicago/Turabian StyleHoang, Lan, František Beneš, Marie Fenclová, Olga Kronusová, Viviana Švarcová, Kateřina Řehořová, Eva Baldassarre Švecová, Miroslav Vosátka, Jana Hajšlová, Petr Kaštánek, and et al. 2020. "Phytochemical Composition and In Vitro Biological Activity of Iris spp. (Iridaceae): A New Source of Bioactive Constituents for the Inhibition of Oral Bacterial Biofilms" Antibiotics 9, no. 7: 403. https://doi.org/10.3390/antibiotics9070403
APA StyleHoang, L., Beneš, F., Fenclová, M., Kronusová, O., Švarcová, V., Řehořová, K., Baldassarre Švecová, E., Vosátka, M., Hajšlová, J., Kaštánek, P., Viktorová, J., & Ruml, T. (2020). Phytochemical Composition and In Vitro Biological Activity of Iris spp. (Iridaceae): A New Source of Bioactive Constituents for the Inhibition of Oral Bacterial Biofilms. Antibiotics, 9(7), 403. https://doi.org/10.3390/antibiotics9070403