
The Activity of Calcium Glycerophosphate and Fluoride against Cariogenic Biofilms of Streptococcus mutans and Candida albicans Formed In Vitro

 , and
, and 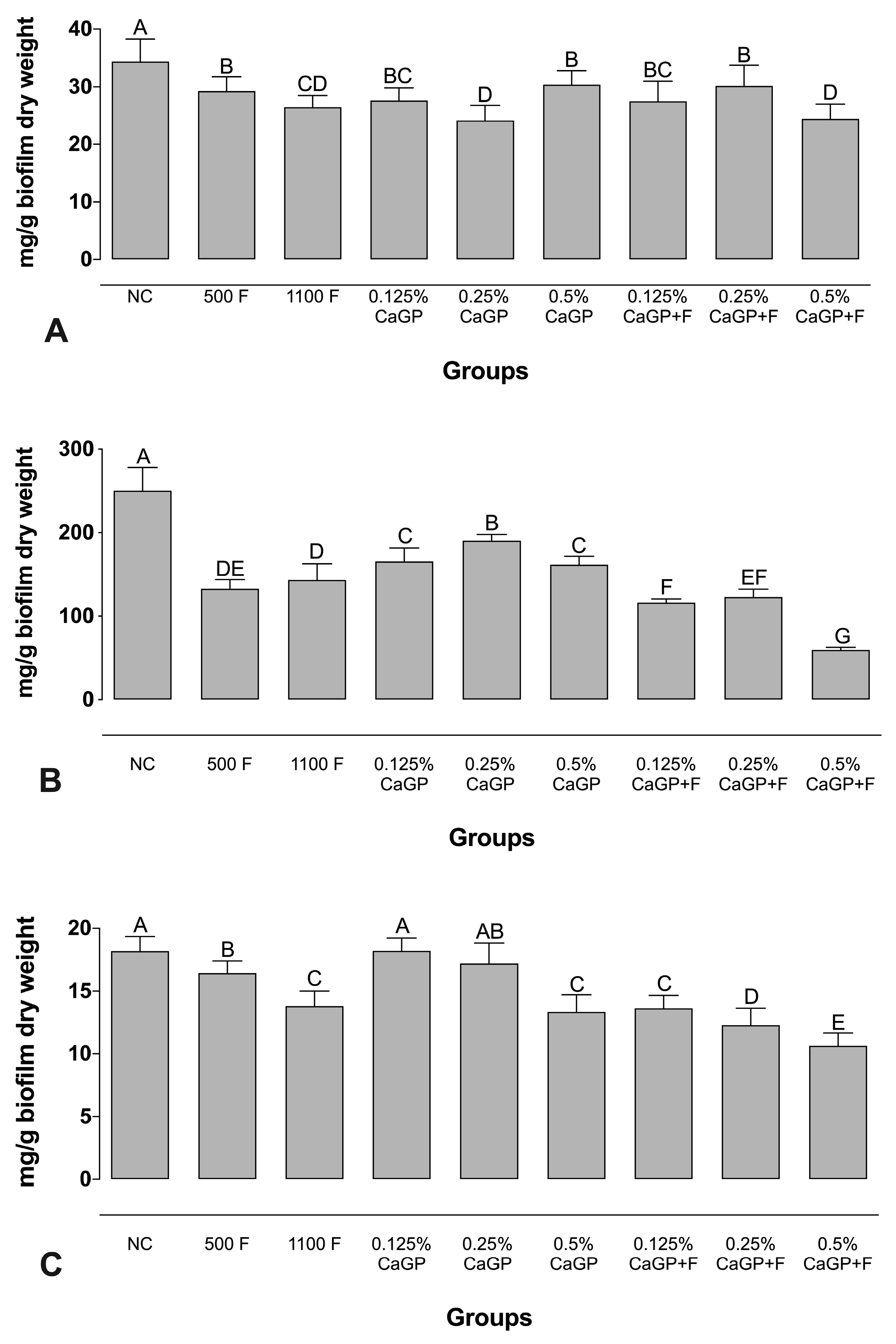{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Evaluation of Extracellular Matrix Composition
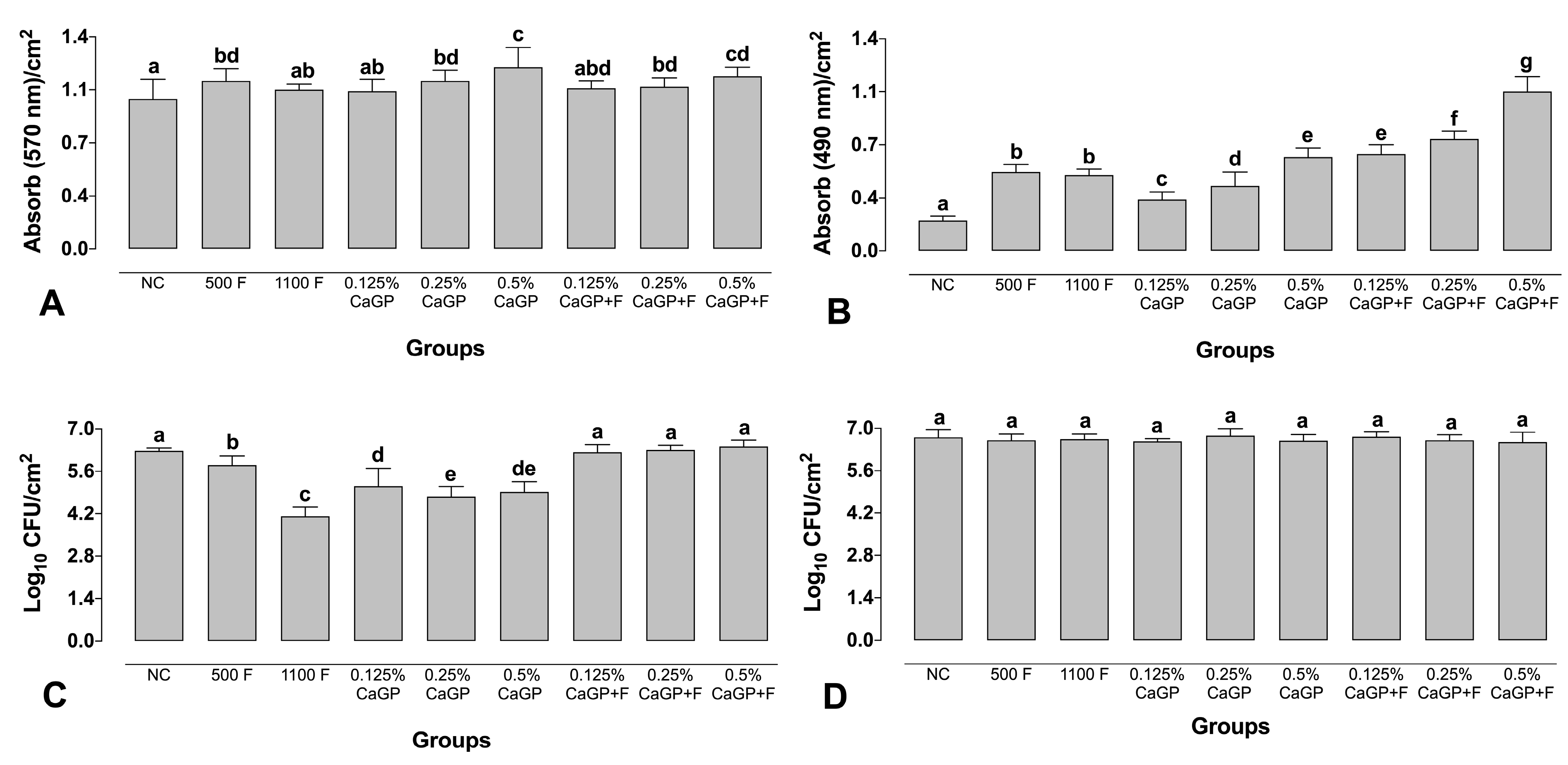
2.2. Quantification Assays
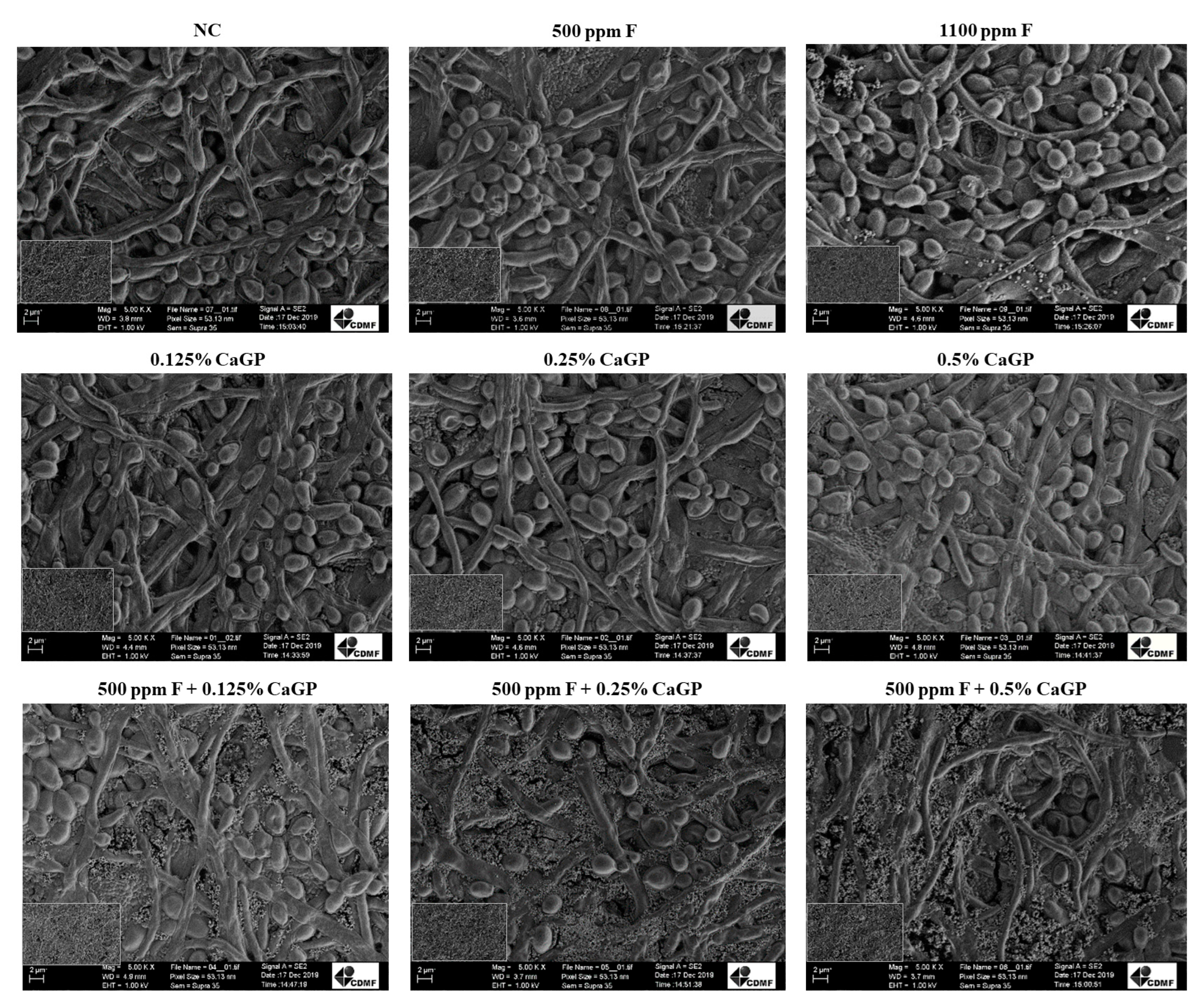
2.3. Structural Analysis of Biofilms
3. Discussion
4. Materials and Methods
4.1. Biofilm Formation
4.2. Treatment of the Biofilms
4.3. Evaluation of Extracellular Matrix Composition
4.4. Quantification Assay
4.5. Structural Analysis of the Biofilm
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pitts, N.B.; Zero, D.T.; Marsh, P.D.; Ekstrand, K.; Weintraub, J.A.; Ramos-Gomez, F.; Tagami, J.; Twetman, S.; Tsakos, G.; Ismail, A. Dental Caries. Nat. Rev. Dis. Prim. 2017, 3, 17030. [Google Scholar] [CrossRef] [PubMed]
- Sheiham, A.; James, W.P. Diet and Dental Caries: The pivotal role of free sugars reemphasized. J. Dent. Res. 2015, 94, 1341–1347. [Google Scholar] [CrossRef] [PubMed]
- Machiulskiene, V.; Campus, G.; Carvalho, J.C.; Dige, I.; Ekstrand, K.R.; Jablonski-Momeni, A.; Maltz, M.; Manton, D.J.; Martignon, S.; Martinez-Mier, E.A.; et al. Terminology of Dental Caries and Dental Caries Management: Consensus Report of a Workshop Organized by ORCA and Cariology Research Group of IADR. Caries Res. 2020, 54, 7–14. [Google Scholar] [CrossRef]
- Kanasi, E.; Johansson, I.; Lu, S.; Kressin, N.; Nunn, M.; Kent, R., Jr.; Tanner, A. Microbial Risk Markers for Childhood Caries in Pediatricians’ Offices. J. Dent. Res. 2010, 89, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Sengupta, A.; Niepa, T.H.R.; Lee, B.-H.; Weljie, A.; Freitas-Blanco, V.S.; Murata, R.M.; Stebe, K.J.; Lee, D.; Koo, H. Candida albicans stimulates Streptococcus mutans microcolony development via cross-kingdom biofilm-derived metabolites. Sci. Rep. 2017, 7, srep41332. [Google Scholar] [CrossRef]
- Xiao, J.; Grier, A.; Faustoferri, R.; Alzoubi, S.; Gill, A.; Feng, C.; Liu, Y.; Quivey, R.; Kopycka-Kedzierawski, D.; Koo, H.; et al. Association between Oral Candida and Bacteriome in Children with Severe ECC. J. Dent. Res. 2018, 97, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Toumba, K.J.; Twetman, S.; Splieth, C.; Parnell, C.; van Loveren, C.; Lygidakis, N. Guidelines on the use of fluoride for caries prevention in children: An updated EAPD policy document. Eur. Arch. Paediatr. Dent. 2019, 20, 507–516. [Google Scholar] [CrossRef]
- Walsh, T.; Worthington, H.V.; Glenny, A.M.; Marinho, V.C.C.; Jeroncic, A. Fluoride toothpastes of different concentrations for preventing dental caries. Cochrane Database Syst. Rev. 2019, 3, CD007868. [Google Scholar] [CrossRef]
- Mejàre, I. Current guidance for fluoride intake: Is it appropriate? Adv. Dent. Res. 2018, 29, 167–176. [Google Scholar] [CrossRef]
- Amaral, J.G.D.; Sassaki, K.T.; Martinhon, C.C.R.; Delbem, A.C.B. Effect of low-fluoride dentifrices supplemented with calcium glycerophosphate on enamel demineralization in situ. Am. J. Dent. 2013, 26, 75–80. [Google Scholar]
- Zaze, A.C.S.F.; Dias, A.P.; Sassaki, K.T.; Delbem, A.C.B. The effects of low-fluoride toothpaste supplemented with calcium glycerophosphate on enamel demineralization. Clin. Oral Investig. 2013, 18, 1619–1624. [Google Scholar] [CrossRef] [PubMed]
- Lynch, R.J.M. Calcium glycerophosphate and caries: A review of the literature. Int. Dent. J. 2004, 54, 310–314. [Google Scholar] [CrossRef]
- Cavazana, T.P.; Hosida, T.Y.; Sampaio, C.; de Morais, L.A.; Monteiro, D.R.; Pessan, J.P.; Delbem, A.C.B. Calcium glycerophosphate and fluoride affect the pH and inorganic composition of dual-species biofilms of Streptococcus mutans and Candida albicans. J. Dent. 2021, 115, 103844. [Google Scholar] [CrossRef] [PubMed]
- Cavazana, T.P.; Hosida, T.Y.; Pessan, J.P.; Sampaio, C.; Monteiro, D.R.; Delbem, A.C.B. Activity of sodium trimetaphosphate, associated or not with fluoride, on dual-species biofilms. Biofouling 2019, 27, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Hosida, T.Y.; Pessan, J.P.; Cavazana, T.P.; Sampaio, C.; Monteiro, D.R.; Delbem, A.C.B. Effect of sodium hexametaphosphate and fluoride on dual-species biofilms of Candida albicans and Streptococcus mutans. Biofouling 2021, 37, 939–948. [Google Scholar] [CrossRef]
- Frencken, J.E.; Sharma, P.; Stenhouse, L.; Green, D.; Laverty, D.; Dietrich, T. Global epidemiology of dental caries and severe periodontitis—A comprehensive review. J. Clin. Periodontol. 2017, 44 (Suppl. S18), S94–S105. [Google Scholar] [CrossRef]
- Islam, M.S. Calcium Signaling: From Basic to Bedside. Adv. Exp. Med. Biol. 2020, 1131, 1–6. [Google Scholar]
- Astasov-Frauenhoffer, M.; Varenganayil, M.M.; Decho, A.W.; Waltimo, T.; Braissant, O. Exopolysaccharides regulate calcium flow in cariogenic biofilms. PLoS ONE 2017, 12, e0186256. [Google Scholar] [CrossRef]
- Rosch, J.W.; Sublett, J.; Gao, G.; Wang, Y.-D.; Tuomanen, E.I. Calcium efflux is essential for bacterial survival in the eukaryotic host. Mol. Microbiol. 2008, 70, 435–444. [Google Scholar] [CrossRef]
- Ripoll, C.; Norris, V.; Thellier, M. Ion condensation and signal transduction. Bioessays 2004, 26, 549–557. [Google Scholar] [CrossRef]
- Dominguez, D.C. Calcium signalling in bacteria. Mol. Microbiol. 2004, 54, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Trombe, M.-C.; Clave, C.; Manias, J.-M. Calcium regulation of growth and differentiation in Streptococcus pneumoniae. J. Gen. Microbiol. 1992, 138, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zhang, B.; Yang, B.; Chen, J.; Wang, H.; Jia, C.; Ding, X.; Xu, N.; Dong, Y.; Zhang, B.; et al. Interaction among the vacuole, the mitochondria, and the oxidative stress response is governed by the transient receptor potential channel in Candida albicans. Free. Radic. Biol. Med. 2014, 77, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Peiter, E. Calcium Transport Proteins in Fungi: The Phylogenetic Diversity of Their Relevance for Growth, Virulence, and Stress Resistance. Front. Microbiol. 2020, 10, 3100. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Shibuya, K.; Lida, H.; Anraku, Y.; Suzuki, T. Roles of Ca2+ in hyphal and yeast-form growth in Candida albicans. Growth regulation by altered extracellular and intracellular free Ca2+ concentrations. Mycoscience 1997, 38, 215–225. [Google Scholar] [CrossRef]
- Nassar, H.M.; Gregory, R.L. Biofilm sensitivity of seven Streptococcus mutans strains to different fluoride levels. J. Oral Microbiol. 2017, 9, 1328265. [Google Scholar] [CrossRef]
- Leitao, T.; Cury, J.A.; Tenuta, L.M.A. Kinetics of calcium binding to dental biofilm bacteria. PLoS ONE 2018, 13, e0191284. [Google Scholar] [CrossRef]
- Rose, R. The role of calcium in oral streptococcal aggregation and the implications for biofilm formation and retention. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2000, 1475, 76–82. [Google Scholar] [CrossRef]
- Körstgens, V.; Flemming, H.-C.; Wingender, J.; Borchard, W. Influence of calcium ions on the mechanical properties of a model biofilm of mucoid Pseudomonas aeruginosa. Water Sci. Technol. 2001, 43, 49–57. [Google Scholar] [CrossRef]
- Hobley, L.; Harkins, C.; MacPhee, C.; Stanley-Wall, N.R. Giving structure to the biofilm matrix: An overview of individual strategies and emerging common themes. FEMS Microbiol. Rev. 2015, 39, 649–669. [Google Scholar] [CrossRef]
- Tomé, F.M.; Paula Ramos, L.; Freire, F.; Pereira, C.A.; de Oliveira, I.C.B.; Junqueira, J.C.; Jorge, A.O.C.; Oliveira, L.D. Influence of sucrose on growth and sensitivity of Candida albicans alone and in combination with Enterococcus faecalis and Streptococcus mutans to photodynamic therapy. Lasers Med. Sci. 2017, 32, 1237–1243. [Google Scholar] [CrossRef]
- Koo, H.; Falsetta, M.L.; Klein, M.I. The exopolysaccharide matrix: A virulence determinant of cariogenic biofilm. J. Dent. Res. 2013, 92, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Klein, M.I.; Falsetta, M.L.; Lu, B.; Delahunty, C.M.; Yates, J.R.; Heydorn, A.; Koo, H. The Exopolysaccharide Matrix Modulates the Interaction between 3D Architecture and Virulence of a Mixed-Species Oral Biofilm. PLOS Pathog. 2012, 8, e1002623. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Cavazana, T.P.; Pessan, J.P.; Hosida, T.Y.; Monteiro, D.R.; Delbem, A.C.B. pH changes of mixed biofilms of Streptococcus mutans and Candida albicans after exposure to sucrose solutions in vitro. Arch. Oral Biol. 2018, 90, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Schobel, B.; Scott-Anne, K.; Watson, G.; Bowen, W.H.; Cury, J.A.; Rosalen, P.L.; Park, Y.K. Apigenin and tt-farnesol with fluoride effects on S. mutans biofilms and dental caries. J. Dent. Res. 2005, 84, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of non-Candida albicans Candida species: Quantification, structure and matrix composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Monteiro, D.R.; Silva, S.; Negri, M.; Gorup, L.F.; de Camargo, E.R.; Oliveira, R.; Barbosa, D.B.; Henriques, M. Silver colloidal nanoparticles: Effect on matrix composition and structure of Candida albicans and Candida glabrata biofilms. J. Appl. Microbiol. 2013, 114, 1175–1183. [Google Scholar] [CrossRef]
- Monteiro, D.R.; Gorup, L.F.; Silva, S.; Negri, M.; de Camargo, E.R.; Oliveira, R.; Barbosa, D.B.; Henriques, M. Silver colloidal nanoparticles: Antifungal effect against adhered cells and biofilms of Candida albicans and Candida glabrata. Biofouling 2011, 7, 711–717. [Google Scholar] [CrossRef]
- Silva, S.; Pires, P.; Monteiro, D.R.; Negri, M.; Gorup, L.F.; Camargo, E.R.; Barbosa, D.B.; Oliveira, R.; Williams, D.W.; Henriques, M.; et al. The effect of silver nanoparticles and nystatin on mixed biofilms of Candida glabrata and Candida albicans on acrylic. Med. Mycol. 2013, 51, 178–184. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavazana, T.P.; Hosida, T.Y.; Sampaio, C.; de Morais, L.A.; Monteiro, D.R.; Pessan, J.P.; Delbem, A.C.B. The Activity of Calcium Glycerophosphate and Fluoride against Cariogenic Biofilms of Streptococcus mutans and Candida albicans Formed In Vitro. Antibiotics 2023, 12, 422. https://doi.org/10.3390/antibiotics12020422
Cavazana TP, Hosida TY, Sampaio C, de Morais LA, Monteiro DR, Pessan JP, Delbem ACB. The Activity of Calcium Glycerophosphate and Fluoride against Cariogenic Biofilms of Streptococcus mutans and Candida albicans Formed In Vitro. Antibiotics. 2023; 12(2):422. https://doi.org/10.3390/antibiotics12020422
Chicago/Turabian StyleCavazana, Thamires Priscila, Thayse Yumi Hosida, Caio Sampaio, Leonardo Antônio de Morais, Douglas Roberto Monteiro, Juliano Pelim Pessan, and Alberto Carlos Botazzo Delbem. 2023. "The Activity of Calcium Glycerophosphate and Fluoride against Cariogenic Biofilms of Streptococcus mutans and Candida albicans Formed In Vitro" Antibiotics 12, no. 2: 422. https://doi.org/10.3390/antibiotics12020422
APA StyleCavazana, T. P., Hosida, T. Y., Sampaio, C., de Morais, L. A., Monteiro, D. R., Pessan, J. P., & Delbem, A. C. B. (2023). The Activity of Calcium Glycerophosphate and Fluoride against Cariogenic Biofilms of Streptococcus mutans and Candida albicans Formed In Vitro. Antibiotics, 12(2), 422. https://doi.org/10.3390/antibiotics12020422