Abstract
The urgent need to combat antibiotic resistance and develop novel antimicrobial therapies has triggered studies on novel metal-based formulations. N-heterocyclic carbene (NHC) complexes coordinate transition metals to generate a broad range of anticancer and/or antimicrobial agents, with ongoing efforts being made to enhance the lipophilicity and drug stability. The lead silver(I) acetate complex, 1,3-dibenzyl-4,5-diphenylimidazol-2-ylidene (NHC*) (SBC3), has previously demonstrated promising growth and biofilm-inhibiting properties. In this work, the responses of two structurally different bacteria to SBC3 using label-free quantitative proteomics were characterised. Multidrug-resistant Pseudomonas aeruginosa (Gram-negative) and Staphylococcus aureus (Gram-positive) are associated with cystic fibrosis lung colonisation and chronic wound infections, respectively. SBC3 increased the abundance of alginate biosynthesis, the secretion system and drug detoxification proteins in P. aeruginosa, whilst a variety of pathways, including anaerobic respiration, twitching motility and ABC transport, were decreased in abundance. This contrasted the affected pathways in S. aureus, where increased DNA replication/repair and cell redox homeostasis and decreased protein synthesis, lipoylation and glucose metabolism were observed. Increased abundance of cell wall/membrane proteins was indicative of the structural damage induced by SBC3 in both bacteria. These findings show the potential broad applications of SBC3 in treating Gram-positive and Gram-negative bacteria.
1. Introduction
The emergence of drug-resistant pathogens necessitates the development of novel antimicrobial agents with alternative modes of action to conventional antimicrobial agents [1]. The rise in drug resistance has been caused partly by the incorrect use of treatments, and this has led to reduced efficacy against Gram-positive and Gram-negative bacteria [2]. β-lactam derivatives such as cephalosporins, monobactams and carbapenems show enhanced broad-spectrum activity and have tackled the growing rates of drug resistance to a certain extent [3]. β-lactam antibiotics target the synthesis of peptidoglycan—a meshwork of polysaccharide strands and peptides that maintain cell structure and rigidity [4]. Gram-positive bacteria (such as Staphylococcus aureus) possess an inner cytoplasmic membrane and thick peptidoglycan layer functionalised with teichoic acids. These anionic glycopolymers play roles in regulating cell morphology and division, cell adhesion and defence against temperature, osmotic and toxic stresses [5]. Gram-negative bacteria (such as Pseudomonas aeruginosa) have a thin peptidoglycan layer sandwiched between an inner cytoplasmic membrane and outer lipopolysaccharide membrane. The latter component offers osmo-protection, regulating the permeability of the cell and the influx and efflux of nutrients and toxins, whilst the presence of outer membrane proteins, or porins, creates channels that permit the passage of molecules in and out of the cell [6]. Decreased production and/or mutations of porins reduce permeability and prevent the influx of drugs, which can confer resistance. Mutations in the expression of the substrate-specific porin, OprD, contribute to carbapenem resistance, which is enhanced when combined with increased efflux system expression [7]. In contrast, Gram-positive bacteria have two main resistance strategies through β-lactamase enzyme production to degrade the β-lactam ring and/or mutations of the drug target site—penicillin-binding proteins (PBPs) [8].
A decrease in antibiotic discovery since the 1970s has encouraged the search for novel therapeutic candidates [9]. N-heterocyclic carbene (NHC) complexes possess broad-spectrum activity against a range of bacterial, fungal and viral pathogens, including SARS-CoV-2 [10,11]. These versatile ligands offer stability, enhance bioavailability and are compatible with a variety of metals to create agents with antimicrobial and/or anticancer properties [12]. The antimicrobial activity of the NHC* metal complex 1,3-dibenzyl-4,5-diphenylimidazol-2-ylidine silver(I) acetate (SBC3) synthesised by the Tacke group has previously demonstrated in vitro and in vivo [13]. Escherichia coli thioredoxin reductase is an SBC3 target and contributor in deregulating redox homeostasis [14]. The antimicrobial properties of silver are well recognised and have been exploited for centuries [15]. Silver sulfadiazine, silver nitrate and colloidal silver are currently in use as topical skin treatments [16].
Proteomic analysis provides the enhanced identification of novel biomarkers with improved specificity and sensitivity, increasing our understanding of disease progression and, in this context, drug discovery, development and toxicity [17]. The identification and quantification of proteins in cells, tissues or whole organisms can provide an insight into the activity of novel therapeutics at a molecular level. Label-free proteomics is an attractive technique with reduced costs, sample preparation time and variability introduced by labelling techniques. An abundance of analytical software tools allows for the extensive interpretation of raw MS data. Here, liquid chromatography tandem mass spectrometry (LC MS/MS) was employed to gain an insight into the mode(s) of action of SBC3 and provide a comparison of the differential responses of Gram-positive and Gram-negative bacteria to SBC3.
2. Results
2.1. The Antimicrobial Effect of SBC3
Evaluation of the effect of SBC3 on P. aeruginosa and S. aureus revealed up to 88.5% and 72.7% growth inhibition at 31.3 and 62.5 μg/mL, respectively. SBC3 demonstrated good inhibitory activity against both pathogens, with higher potency against P. aeruginosa (Figure 1). Based on the results in Figure 1, two concentrations of SBC3 that partly inhibited bacterial growth (P. aeruginosa (7.5 μg/mL) and S. aureus (12 μg/mL)) were identified and these concentrations allowed for sufficient cell density for protein extraction after six hours, and for cells to demonstrate a response to the compound.

Figure 1.
Evaluation of the effect of SBC3 on the growth of P. aeruginosa and S. aureus. All values are the mean ± S.E. of three independent experiments.
2.2. Characterisation of the Proteomic Response of Bacteria to SBC3
Label-free quantitative (LFQ) mass spectrometry was employed to characterise changes in the proteomes of SBC3-treated and untreated bacteria. P. aeruginosa and S. aureus were exposed to the relevant SBC3 concentration for six hours, after which time proteins were extracted and analysed in order to determine the effect of SBC3 on the whole-cell proteomic response.
2.3. Characterisation of the Effect of SBC3 on the Proteome of P. aeruginosa
A total of 2526 P. aeruginosa proteins were identified initially, of which 1759 remained following the filtration of contaminants. The entire data set of filtered proteins is shown on the principal component analysis plot (PCA) (Figure 2), whereby all biological replicates (n = 4) are resolved into the corresponding sample groups. Distinct differences are evident between untreated and SBC3-exposed samples, with a combined variance of 79% resulting from Components 1 and 2. Subsequent two-sample t-tests (p < 0.05) were performed on 1759 proteins post-imputation and identified 362 statistically significant (p < 0.05) and differentially abundant (SSDA) proteins with a minimum fold change of 1.5.
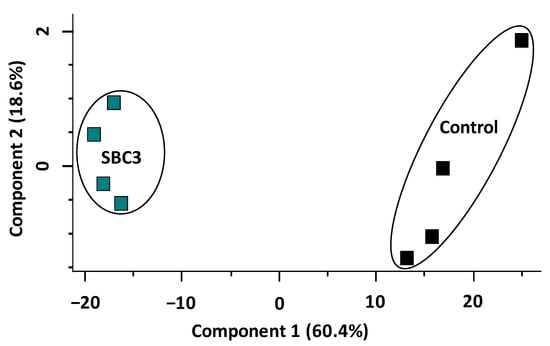
Figure 2.
Principal component analysis (PCA) of SBC3-treated P. aeruginosa versus untreated control samples.
The distribution of all filtered proteins (1631) is represented on a volcano plot (Figure 3) conducted via a pairwise Student’s t-test (p < 0.05), and the top ten most differentially abundant proteins (±fold change ≥ 1.5) are highlighted and annotated with associated gene names. The protein products are listed in Table 1. Thiol:disulfide interchange protein DsbG (+142-fold), transcription factor Amrz (+12-fold) required for environmental adaptation and probable two-component response regulator (+10-fold) for proteolysis and biofilm formation were increased in abundance. Haemagg_act domain-containing protein (−39-fold), ribosome modulation factor (−37-fold) and probable binding protein component of ATP-binding cassette (ABC) transporter (−11-fold) were decreased in abundance.

Figure 3.
Volcano plot generated from two-sample t-tests showing the distribution of all identified proteins (post-filtration of contaminants). The x-axis denotes fold change (log2 LFQ intensity difference), and the y-axis denotes the p value (−log10 p value). Statistically significant proteins (p < 0.05) lie above the horizontal, dashed line, and proteins with a fold change of ≥1.5 are to the right and left of the vertical lines. The top ten most increased and decreased proteins in abundance are coloured red and blue, respectively. The corresponding gene names are annotated, and the protein products and corresponding functions are listed in Table 1.

Table 1.
The top 20 differentially abundant proteins identified in SBC3-treated P. aeruginosa.
2.4. Characterisation of the Effect of SBC3 on the Proteome of S. aureus
Proteomic analysis of S. aureus treated with SBC3 identified 1271 proteins and 1183 filtered proteins. The PCA plot (Figure 4) summarises this entire data set, showing the contrast between untreated and SBC3-treated S. aureus proteomes. A combined variance of 65.8% arose from Components 1 and 2.
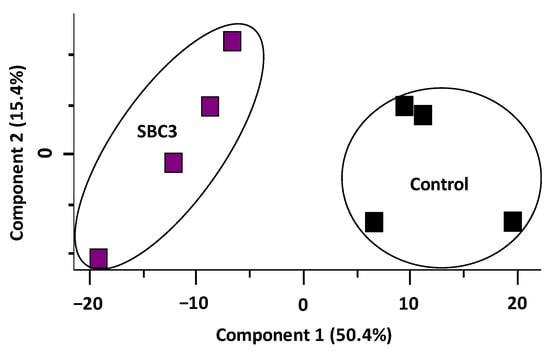
Figure 4.
Principal component analysis (PCA) of SBC3-treated S. aureus versus untreated control samples.
A volcano plot was generated representing the distribution of all filtered proteins (Figure 5). The top ten most differentially abundant proteins are highlighted and annotated by the corresponding gene names. The protein products and relative fold changes are displayed in Table 2. A metalloproteinase (+7.4-fold) and proteins associated with cell wall organisation (glycyl-glycine endopeptidase LytM; +6.2-fold, probable autolysin SsaALP; +3.3-fold), DNA replication (DNA topoisomerase 3; +5.2-fold), ABC-type transporter activity (ABC transporter permease; +4-fold) and oxidative stress defence (peroxide-responsive repressor PerR; +3.4) were increased in abundance. Proteins associated with transcriptional regulation of virulence factors (HTH-type transcriptional regulator rot; −8.8-fold), deoxyribonuclease activity (hydrolase TatD; −8.2-fold) and cell redox homeostasis (thioredoxin domain-containing protein; −4.1-fold) were decreased in abundance.
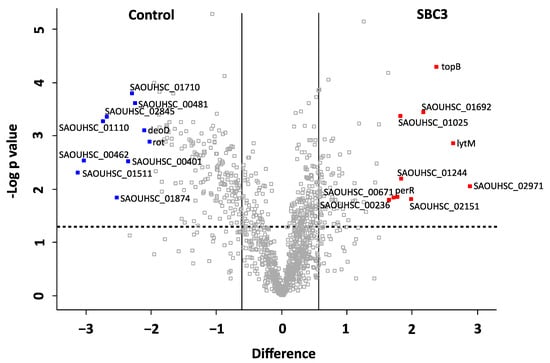
Figure 5.
Volcano plot generated from a two-sample t-test (p < 0.05) of S. aureus treated with SBC3 (12 μg/mL) versus untreated control samples. All identified and filtered proteins are displayed based on p value (-log10 p value) on the y-axis and fold change (log2 mean LFQ intensity difference) on the x-axis. Statistically significant (p < 0.05) proteins are positioned above the horizontal, dashed line, and proteins with a fold change of ≥1.5 are to the right and left of the vertical lines. The top ten most differentially abundant proteins are highlighted and annotated with gene names, which are listed in Table 2, and including the protein products, functions and fold change.
Table 2.
The top 20 most differentially abundant proteins identified in S. aureus treated with SBC3.
2.5. Interaction Network Analysis on the Response of P. aeruginosa and S. aureus to SBC3
Protein interaction networks generated via STRING incorporated SSDA proteins derived from pairwise t-tests. Individual nodes representing proteins (annotated with the corresponding gene name or STRING identifier) are connected by lines denoting an interaction and the width of the line denotes confidence i.e., the strength of data support.
Exposure of P. aeruginosa to SBC3 resulted in 366 SSDA proteins with a log2 fold difference ranging from −5.2 to 7.2. Mapping of these proteins revealed a multitude of targeted protein pathways (Figure 6). Whilst some virulence and resistance mechanisms (alginate/spermidine biosynthesis, type VI secretion and drug efflux) were elevated (Figure 6A), a large proportion of clusters were decreased and these included ABC transporters, the type III secretion system, respiration and amino acid metabolism (Figure 6B).
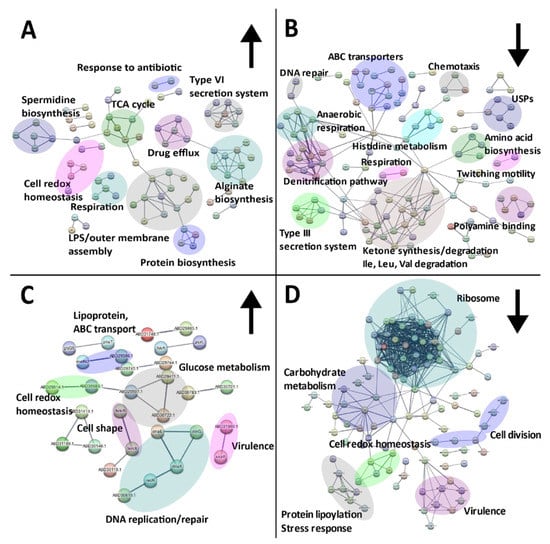
Figure 6.
STRING network analyses of SSDA proteins derived from two-sample t-tests (p < 0.05) showing the responses of P. aeruginosa (A,B) and S. aureus (C,D) to SBC3. Nodes represent individual proteins (annotated with the corresponding gene name or STRING identifier) and are connected to represent an interaction. The thickness of the line represents the strength of data support. Interaction networks with proteins increased in abundance are displayed on the left (A,C) indicated by upward arrows and those decreased are shown on the right (B,D) indicated by downward arrows. Protein genes/IDs were searched via the UniProt database in addition to functional enrichment analysis on STRING to determine KEGG and Gene Ontology (GO) terms.
S. aureus challenged with SBC3 generated 251 SSDAs with a log2 difference of −3.1 to 2.9. STRING protein clusters associated with cell wall organisation, cell redox homeostasis and DNA replication and repair were among the most elevated (Figure 6C). A substantial number of clusters, such as ribosome, lipoylation and glycolysis proteins, were decreased in treated samples (Figure 6D).
3. Discussion
P. aeruginosa and S. aureus are widespread nosocomial pathogens, displaying high levels of multidrug resistance [18]. Susceptible individuals are at risk of contracting chronic wound infections, sepsis, endocarditis and other infections introduced via indwelling medical devices (e.g., catheter-associated urinary tract infection) [19]. P. aeruginosa and S. aureus are the main causes of bacterial infection in cystic fibrosis (CF) patients. S. aureus typically precedes P. aeruginosa in the earlier stages of disease; however, incidents of co-infection enhance morbidity and mortality rates [20]. Biofilm formation is common between these species and offers high levels of protection and resistance against host immune clearance and antimicrobial agents [21].
3.1. SBC3 Compromises Cell Structural Integrity in Both S. aureus and P. aeruginosa
The onset of resistance to conventional antibiotics has prompted the development of improved silver formulations for the treatment of drug-resistant pathogens [22]. Positively charged silver (Ag+) ions bind to negatively charged cell membranes and induce cell membrane/wall leakage and/or rupture [23]. The proteomics results presented here revealed significant alterations in the abundance of proteins associated with cell wall structure in S. aureus. For example, the peptidoglycan biogenesis and degradation protein, glycyl-glycine endopeptidase LytM, was increased +6.2-fold following exposure to SBC3. Acyltransferase enzymes (endcoded by the femA and femX genes) utilised in cell wall organisation and peptidoglycan biosynthesis were also elevated [24]. Alterations in the abundance of outer membrane proteins and the lipopolysaccharide layer in P. aeruginosa provide further evidence of the roles of SBC3 in disrupting the cell’s structural integrity. The increased abundance of outer membrane protein assembly factor BamB (+2.9-fold) and outer membrane protein assembly factor BamD (+1.8-fold) is indicative of a stress response [25], whilst an array of lipopolysaccharide proteins identified within a STRING cluster could be indicative of the cell’s response in decreasing membrane permeability and limiting the entry of noxious compounds [26,27].
Elevated abundance of SpeH (+2-fold) and SpeE2 (+1.9-fold) was a further indication of the role of silver in damaging cell structural integrity. This compliments previous studies where the treatment of P. aeruginosa PAO1 with sublethal concentrations of polymyxin B (an outer-membrane-damaging agent) resulted in increased expression of SpeE [28].
3.2. The Proteomic Effects of SBC3 on P. aeruginosa
3.2.1. Translocation and Biofilm Formation
Chronic infection by P. aeruginosa in CF patients is characterised by phenotypic switching from a non-mucoidal to mucoidal phenotype and concurs with alginate over-production [29]. Proteins identified within the STRING cluster were among some of the most differentially abundant. The highly conserved transcription factor AmrZ (amrZ) and activator of alginate biosynthesis were increased +12.2-fold, whereas PA3205 (identified as LTXXQ domain protein via UniProt) was also increased +12.2-fold, and this is hypothesised to detect membrane alterations and commence adherence and biofilm formation on abiotic surfaces [30]. Other key regulators in alginate biosynthesis, such as sigma factor AlgU negative regulatory protein (mucA, +1.9-fold), sigma factor AlgU regulatory protein MucB (mucB, +1.5-fold), periplasmic serine endoprotease DegP-like (mucD, +1.6-fold), alginate biosynthesis transcriptional regulatory protein AlgB (algB, +1.6-fold) and phosphomannomutase/phosphoglucomutase (algC, +1.9-fold) are also located within this cluster.
The successful colonisation and establishment of mature biofilms depends on a chemosensory system mediated by pili for translocation towards favourable environments [31], both of which were reduced post-exposure to SBC3 (Figure 6). Detection of chemical gradients by chemotaxis initiates adherence, whilst the type IV pilus system in P. aeruginosa consists of cell surface appendages that elongate and retract to further enable adhesion, biofilm formation, twitching motility and DNA uptake [32,33]. Pil proteins can be categorised into four subcomplexes, including the outer membrane secretin pore complex (PilQ, +1.6-fold), the inner membrane alignment subcomplex (PilM, −2.3-fold) and the cytoplasmic motor subcomplex (PilU, −1.8-fold) (Figure 6/Supplementary Dataset S1B). The fourth subcomplex is the pilus itself, requiring PilA and minor pilins. The non-pilin protein PilY1, which showed a −1.8-fold reduction in abundance, has roles in pilus retraction and as a mechanosensory element, which, upon attachment, induces an acute virulence phenotype [31,34,35,36].
3.2.2. Virulence
The type III secretion system of P. aeruginosa injects effector proteins into host cell cytoplasm and exerts cytotoxicity activity and suppresses host immunity [37,38,39]. One such effector, responsible for disrupting phagocytosis, is the secreted exoenzyme S (exoS), and this was decreased in abundance by −6-fold following SBC3 treatment [40]. Translocator protein PopB (−5.8-fold) and translocator outer membrane protein PopD (−8.4-fold) comprise the pore and are linked to the needle tip type III secretion protein PcrV [41]. Transcription anti-activator ExsD (−1.7-fold) is a negative regulator of the type III secretion system regulon [42].
Upregulation of secretion systems can arise in response to antibiotic-induced stress [43]. Components of the type VI secretion system such as the type VI secretion system sheath protein TssB1 (+4.8-fold), type VI secretion system sheath protein TssC1 (+6.5-fold), type VI secretion system baseplate component TssK1 (+2.3-fold) and protein Hcp1 (+3-fold), which is an effector protein whose secretion is aided by AAA+ ATPase ClpV1 (clpV1) (+2.2-fold), were increased in abundance [44].
3.2.3. Anaerobic Respiration/Stress Response
Thick mucus in the CF lung can generate an anaerobic environment to which P. aeruginosa must adapt for survival [45]. Anaerobic growth is mediated by two main pathways, (1) arginine fermentation and (2) the denitrification pathway, whilst pyruvate fermentation is implemented in nutrient-deprived environments. Although growth and metabolism are halted, the latter sustains long-term survival [46]. Universal stress proteins (USPs), such as PA3309 (encoded by uspK) and PA4352 (uspN), have roles in pyruvate fermentation that are essential for anaerobic stationary phase survival. Both proteins were decreased in abundance in response to SBC3 (−4.5-fold and −4.8-fold, respectively). In addition, PA1789 (uspL), PA4328 (uspM) and PA5207 (uspO) proteins were decreased −4.9-, −6.3- and −5.5-fold, respectively, and all of these are induced in oxygen-depleted conditions [47]. Transcription of these genes is controlled by the global anaerobic regulator Anr (encoded by anr) and interestingly was increased +1.8-fold (gene and protein products are located in Supplementary File S1) [48].
Denitrification is a multistep process involving nitrate reduction to ultimate nitrogen gas formation [49]. Heme d1 is an isobacteriochlorin co-factor of the nitrite reductase, NirS, which catalyses the reduction of nitrite to nitric oxide [50]. Synthesis of heme d1 requires enzymes encoded by the nir genes [51,52,53], several of which (and their protein products) were decreased: nirD (siroheme decarboxylase NirD subunit, −10.1-fold), nirE (uroporphyrinogen-III C-methyltransferase, −11.7-fold), nirG (siroheme decarboxylase NirG subunit, −7.8-fold), nirL (siroheme decarboxylase NirL subunit, −3-fold), nirN (dihydro-heme d1 dehydrogenase, −4.1-fold), nirQ (denitrification regulatory protein NirQ, −13.8-fold) and nirS (nitrite reductase, −5.9-fold). Also shown are the enzymatic complex NarGHI nitrate reductase genes utilised in anaerobic respiration, narG (nitrate reductase (quinone), −2.8-fold), narH (respiratory nitrate reductase beta chain, −2.4-fold) and narJ (respiratory nitrate reductase delta chain, −2.1-fold), along with dnr (transcriptional regulator Dnr, −3.3-fold) [54]. The disruption of these growth and/or anaerobic survival strategies highlights the potential of SBC3 for the treatment of recalcitrant infections.
ABC transporters belong to a superfamily of proteins driven by ATP hydrolysis to facilitate cellular import and export. The uptake of di- and tripeptides for nitrogen sources is mediated by the DppBCDF system consisting of DppA1-5 substrate-binding proteins (in P. aeruginosa PA14), which play a role in biofilm formation and swarming ability [55]. Homologous proteins belonging to this system were decreased in abundance following treatment with SBC3 (PA4497/dppA7 and PA4496/dppA9; −10.6- and −2.9-fold, respectively). The high-affinity branched-chain amino acid transporters within the same cluster, including high-affinity branched-chain amino acid transport ATP-binding protein BraG (−1.5-fold) and leucine-, isoleucine-, valine-, threonine- and alanine-binding protein (part of the high-affinity branched amino acid transport system LIV-1; −1.8-fold), were also among the same cluster of ABC transporters. Branched-chain amino acids (Ile, Leu and Val) are fundamental nutrients for protein synthesis that not only support growth, but also environmental adaptation and virulence [56]. Interference with amino acid metabolism is also shown through the decreased abundance of proteins associated with amino acid catabolism via the liu gene cluster (Ile, Leu and Val degradation), in addition to ketone body regulation, which serves as an alternative metabolic fuel source [57].
Putrescine and spermidine are linked to a multitude of functions, such as virulence, biofilm formation and antibiotic resistance [28]. PotABCD polyamine transporters for putrescine and spermidine uptake were altered by SBC3 treatment (potA/PA0603, −1.8-fold and potD, −2.7-fold). Another ATP-binding component of the ABC transporter protein encoded by PA1807 was increased within this cluster by two-fold. The exact function of this protein in P. aeruginosa is unclear but, as part of the YejABEF ABC transporter system in Brucella melitensis, these proteins conferred resistance to polymyxin B [58].
3.2.4. Detoxification Mechanisms
Exposure to SBC3 increased the abundance of multidrug resistance proteins MexA and B, belonging to MFP and RND, respectively, by +2.2-fold and +1.9-fold. P. aeruginosa has a homologous efflux system, MexCD-OprJ, including RND multidrug efflux membrane fusion protein MexC, +1.9-fold, and efflux pump membrane transporter (encoded by mexD), +1.8-fold [59,60].
The increased abundance of cell redox homeostasis proteins such as catalase (kat B, +3.7-fold), thioredoxin reductase (trxB2, +2.3-fold), alkyl hydroperoxide reductase C (PA0848, +1.9-fold) and alkyl hydroperoxide reductase subunit F (ahpF, +1.6-fold) is an indicator of a stress response. Interestingly, the most differentially abundant P. aeruginosa protein identified here was thiol:disulfide interchange protein DsbG (+142.1-fold), which is also required for the maintenance of cell redox homeostasis. It is possible that this increase is a damage control measure against misfolded proteins, which reflects previous findings in response to copper stress [61].
3.2.5. Aerobic Respiration
The ability to adapt to hostile environments is dependent on adequate energy supplies. The tricarboxylic acid (TCA) cycle is the predominant energy source for cells as part of aerobic respiration [62]. SBC3 exposure increased the abundance of several TCA cycle proteins and decreased two cytochrome-c oxidases (ccoO2/ccOP1/N1) in P. aeruginosa (Figure 6).
3.3. The Proteomic Response of S. aureus to SBC3
3.3.1. Protein Synthesis
Inhibition of protein synthesis can have detrimental effects on cell proliferation [63]. This, along with structural differences to eukaryotic ribosomes, has been exploited in conventional antibiotics (aminoglycosides, macrolides and tetracyclines) to inhibit protein synthesis; however, increasing rates of antibiotic resistance demand alternative options [64]. Functional enrichment analysis via STRING revealed a significant reduction in S. aureus ribosomal proteins necessary for transcription and translation following SBC3 exposure. This is consistent with previous studies that examined the proteomic response of Candida parapsilosis to SBC3 [65].
3.3.2. Protein Lipoylation
The functioning of several metabolic enzymes relies on post-translation modification, namely lipoylation [66]. S. aureus can acquire lipoic acid via de novo synthesis or retrieval from their host under nutrient-deficient conditions in the environment. This important co-factor is required by enzyme complexes used for metabolism and host immune suppression [67,68]. One such complex is the glycine cleavage system (GCS), which catalyses the degradation of glycine and comprises four proteins: P, H, T and L proteins. Lipoic acid synthesis commences with the transfer of octanoic acid to GcvH via an octanoyltransferase LipM (Supplementary Dataset S2B). This mechanism is an adaptive strategy to overcome nutrient scarcity [69]. Within this downregulated cluster, glutamine synthetase (encoded by glnA) was also identified. This enzyme has multifunctional roles as a transcriptional co-regulator and chaperone in ammonium assimilation, in addition to growth and biofilm formation. This may open avenues as a novel therapeutic target [70].
3.3.3. Glucose Metabolism
The role of silver in targeting the glycolytic enzyme glyceraldehyde-3-phosphate dehydrogenase in E. coli has been characterised [71]. Silver’s disruption of glyceraldehyde-3-phosphate dehydrogenase preceded more recent studies on the inactivation of glycolytic enzymes in S. aureus in response to silver [72]. Treatment of S. aureus with SBC3 increased the abundance of some glycolytic enzymes (glucokinase and alpha amylase family and 6-phospo-beta-glucosidase proteins, putative), while a larger proportion of proteins associated with the pentose phosphate and glycolysis pathways was decreased. It is important to note that glucose-derived energy is essential for S. aureus (and other pathogens) to establish infection [73].
3.3.4. Cell Redox Homeostasis
S. aureus challenged with a higher dose of SBC3 (21.1 μM) to P. aeruginosa proved more effective in disrupting the oxidative stress response. Whilst thioredoxin reductase and YpdA family bacillithiol disulfide reductase were elevated in abundance, thioredoxin, catalase, superoxide dismutases and alkyl hydroperoxide reductase C were significantly reduced in abundance.
4. Materials and Methods
4.1. Bacterial Culture Conditions
P. aeruginosa PAO1 and S. aureus ATCC 33591 cultures were grown in nutrient broth (Oxoid, Basingstoke, UK) at 37 °C in an orbital shaker at 200 rpm. Bacterial stocks were maintained on nutrient agar at 4 °C.
4.2. Antibacterial Susceptibility Assays
Bacterial cultures were grown overnight in nutrient broth (Oxoid, Basingstoke, UK) in an orbital shaker at 200 rpm at 37 °C. SBC3 (1 mg/mL) was dissolved in sterile nutrient broth with 5% dimethylsulfoxide (DMSO; Honeywell, SLS Scientific Laboratory Supplies, Ireland Ltd). Serial dilutions of the complexes were performed in 96-well plates (Corning®, Somerville, MA, USA) containing nutrient broth (100 μL/well) to give a concentration range of 0.49–125 μg/mL. The optical density (OD) of overnight cultures was measured at 600 nm (OD600) and cell suspensions were adjusted to 0.01 in nutrient broth. Aliquots of cells (100 μL) were added to each SBC3 concentration to obtain a final cell density of approximately 1 × 107 cells/well (S. aureus) and 3 × 105 cells/well (P. aeruginosa) (DMSO was added to untreated control samples at an equivalent concentration to the highest treatment dose (0.625%)). Plates were incubated in a static incubator at 37 °C for 18 h and read at 600 nm in a plate reader (Bio-Tek Synergy HT, Mason Technology, Dublin, Ireland) to measure bacterial growth.
4.3. Proteomic Analysis of P. aeruginosa Treated with SBC3
Sterile nutrient broth was inoculated with P. aeruginosa (~9 × 106 CFUs/mL) and S. aureus (~7 × 108 CFUs/mL) and grown in the presence of SBC3 (7.5 μg/mL and 12 μg/mL, respectively) at 37 °C in an orbital shaker at 200 rpm for 6 h until the mid-exponential growth phase. Proteins were extracted, digested and purified as described [64]. Dried samples were resuspended and loaded onto a Q Exactive Mass Spectrometer (2 μL containing 750 ng of protein) using a 133 min reverse phase gradient.
Protein quantification and label-free quantification (LFQ) normalisation were processed through MaxQuant software version 1.6.3.4 using Staphylococcus aureus NCT8325 (downloaded 12 October 2021; 2889 entries) and Pseudomonas aeruginosa PAO1 (downloaded 9 July 2021; 5564 entries) databases following previously described search parameters [74]. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE [75] partner repository with the dataset identifier PXD038616.
4.4. Data Analysis
Statistical and graphical analyses were carried out by processing the resulting LFQ values generated via MaxQuant through the statistical software tool Perseus v.1.6.6.0, with all parameters set in accordance with previous methods [64].
The Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) v11.5 www.string-db.org/ (accessed on 1 November 2022) was used to map statistically significant and differentially abundant (SSDA) proteins using their corresponding gene names retrieved from UniProt gene lists for P. aeruginosa PAO1 and S. aureus NCTC 8325. A high confidence score (0.7) was used to generate protein/protein interaction networks for treatment versus control sample groups. Disconnected nodes were omitted. Functional enrichment analysis was carried out along with manual searches using the UniProt database to determine individual protein and pathway functions.
5. Conclusions
Exposure of S. aureus and P. aeruginosa to SBC3 resulted in the inhibition of growth but differential proteomic responses. Both cell types showed alterations in the abundance of proteins associated with the cell wall or envelope. However, in P. aeruginosa, a multitude of pathways were affected, including alginate biosynthesis, secretion systems, drug detoxification and anaerobic respiration. This contrasted with the response of S. aureus, where pathways such as protein synthesis, lipoylation, glucose metabolism and cell redox homeostasis were affected. The results presented here demonstrate the broad-spectrum activity of SBC3 against Gram-positive and Gram-negative bacteria, with differing proteomic responses from both cell types. SBC3 may have applications in the treatment of Gram-positive and Gram-negative mono- or dual infections.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/antibiotics12020348/s1, Supplementary dataset S1: All identified proteins in P. aeruginosa treated with SBC3.; Supplementary dataset S2: All identified proteins in S. aureus treated with SBC3.
Author Contributions
Conceptualisation, M.T. and K.K.; methodology, M.P., Z.B. and C.O.; formal analysis, M.P.; resources, K.K. and M.T.; writing—original draft preparation, M.P. and C.O.; writing—review and editing, K.K. and M.T.; supervision, K.K. and M.T.; funding acquisition, K.K. and M.T. All authors have read and agreed to the published version of the manuscript.
Funding
M.P. is funded by a grant from Science Foundation Ireland (SFI) and is co-funded under the European Regional Development Fund under grant number 12/RC/2275_P2. The Q-exactive mass spectrometer was funded under the SFI Research Infrastructure Call 2012; Grant Number: 12/RI/2346 (3).
Institutional Review Board Statement
Not applicable.
Informed Consent statement: Not applicable.
Data Availability Statement
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE [75] partner repository with the dataset identifier PXD038616.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kumar, M.; Sarma, D.K.; Shubham, S.; Kumawat, M.; Verma, V.; Nina, P.B.; JP, D.; Kumar, S.; Singh, B.; Tiwari, R.R. Futuristic Non-antibiotic Therapies to Combat Antibiotic Resistance: A Review. Front. Microbiol. 2021, 12, 609459. [Google Scholar] [CrossRef] [PubMed]
- Lee Ventola, M.C. The Antibiotics Resistance Crisis Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Worthington, R.J.; Melander, C. Overcoming resistance to β-Lactam antibiotics. J. Org. Chem. 2013, 78, 4207–4213. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Bradford, P.A. β-lactams and β-lactamase inhibitors: An overview. Cold Spring Harb. Perspect. Med. 2016, 6, a025247. [Google Scholar] [CrossRef]
- Romaniuk, J.A.H.; Cegelski, L. Peptidoglycan and Teichoic Acid Levels and Alterations in Staphylococcus aureus by Cell-Wall and Whole-Cell Nuclear Magnetic Resonance. Biochemistry 2018, 57, 3966–3975. [Google Scholar] [CrossRef]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. Biophys. Acta-Proteins Proteom. 2009, 1794, 808–816. [Google Scholar] [CrossRef]
- Li, H.; Luo, Y.F.; Williams, B.J.; Blackwell, T.S.; Xie, C.M. Structure and function of OprD protein in Pseudomonas aeruginosa: From antibiotic resistance to novel therapies. Int. J. Med. Microbiol. 2012, 302, 63–68. [Google Scholar] [CrossRef]
- Jubeh, B.; Breijyeh, Z.; Karaman, R. Resistance of gram-positive bacteria to current antibacterial agents and overcoming approaches. Molecules 2020, 25, 2888. [Google Scholar] [CrossRef]
- Aminov, R.I. A brief history of the antibiotic era: Lessons learned and challenges for the future. Front. Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef]
- Özdemir, I.; Özcan, E.Ö.; Günal, S.; Gürbüz, N. Synthesis and antimicrobial activity of novel Ag-N-Heterocyclic carbene complexes. Molecules 2010, 15, 2499–2508. [Google Scholar] [CrossRef]
- Prencipe, F.; Zanfardino, A.; Di Napoli, M.; Rossi, F.; D’errico, S.; Piccialli, G.; Mangiatordi, G.F.; Saviano, M.; Ronga, L.; Varcamonti, M.; et al. Silver (I) n-heterocyclic carbene complexes: A winning and broad spectrum of antimicrobial properties. Int. J. Mol. Sci. 2021, 22, 2497. [Google Scholar] [CrossRef]
- Hindi, K.M.; Panzner, M.J.; Tessier, C.A.; Cannon, C.L.; Youngs, W.J. The medicinal applications of imidazolium carbene-metal complexes. Chem. Rev. 2009, 109, 3859–3884. [Google Scholar] [CrossRef]
- O’Beirne, C.; Piatek, M.E.; Fossen, J.; Müller-Bunz, H.; Andes, D.R.; Kavanagh, K.; Patil, S.A.; Baumann, M.; Tacke, M. Continuous flow synthesis and antimicrobial evaluation of NHC* silver carboxylate derivatives of SBC3 in vitro and in vivo. Metallomics 2021, 13, mfaa011. [Google Scholar] [CrossRef]
- O’Loughlin, J.; Napolitano, S.; Alkhathami, F.; O’Beirne, C.; Marhöfer, D.; O’Shaughnessy, M.; Howe, O.; Tacke, M.; Rubini, M. The Antibacterial Drug Candidate SBC3 is a Potent Inhibitor of Bacterial Thioredoxin Reductase. ChemBioChem 2021, 22, 1093–1098. [Google Scholar] [CrossRef]
- Alexander, J.W. History of the medical use of silver. Surg. Infect. 2009, 10, 289–292. [Google Scholar] [CrossRef]
- Politano, A.D.; Campbell, K.T.; Rosenberger, L.H.; Sawyer, R.G. Use of silver in the prevention and treatment of infections: Silver review. Surg. Infect. 2013, 14, 8–20. [Google Scholar] [CrossRef]
- Amiri-Dashatan, N.; Koushki, M.; Abbaszadeh, H.A.; Rostami-Nejad, M.; Rezaei-Tavirani, M. Proteomics applications in health: Biomarker and drug discovery and food industry. Iran. J. Pharm. Res. 2018, 17, 1523–1536. [Google Scholar]
- van Duin, D.; Paterson, D.L. Multidrug-Resistant Bacteria in the Community: Trends and Lessons Learned. Infect. Dis. Clin. N. Am. 2016, 30, 377–390. [Google Scholar] [CrossRef]
- Goldsworthy, M.J.H. Gene expression of Pseudomonas aeruginosa and MRSA within a catheter-associated urinary tract infection biofilm model. Biosci. Horiz. 2008, 1, 28–37. [Google Scholar] [CrossRef]
- Trizna, E.Y.; Yarullina, M.N.; Baidamshina, D.R.; Mironova, A.V.; Akhatova, F.S.; Rozhina, E.V.; Fakhrullin, R.F.; Khabibrakhmanova, A.M.; Kurbangalieva, A.R.; Bogachev, M.I.; et al. Bidirectional alterations in antibiotics susceptibility in Staphylococcus aureus—Pseudomonas aeruginosa dual-species biofilm. Sci. Rep. 2020, 10, 14849. [Google Scholar] [CrossRef]
- Reigada, I.; San-Martin-Galindo, P.; Gilbert-Girard, S.; Chiaro, J.; Cerullo, V.; Savijoki, K.; Nyman, T.A.; Fallarero, A.; Miettinen, I. Surfaceome and Exoproteome Dynamics in Dual-Species Pseudomonas aeruginosa and Staphylococcus aureus Biofilms. Front. Microbiol. 2021, 12, 672975. [Google Scholar] [CrossRef]
- Turner, R.J. Is silver the ultimate antimicrobial bullet? Antibiotics 2018, 7, 112. [Google Scholar] [CrossRef]
- Dakal, T.C.; Kumar, A.; Majumdar, R.S.; Yadav, V. Mechanistic basis of antimicrobial actions of silver nanoparticles. Front. Microbiol. 2016, 7, 1831. [Google Scholar] [CrossRef]
- Sharif, S.; Kim, S.J.; Labischinski, H.; Schaefer, J. Characterization of peptidoglycan in fem-deletion mutants of methicillin-resistant staphylococcus aureus by solid-state NMR. Biochemistry 2009, 48, 3100–3108. [Google Scholar] [CrossRef]
- Klein, K.; Sonnabend, M.S.; Frank, L.; Leibiger, K.; Franz-Wachtel, M.; Macek, B.; Trunk, T.; Leo, J.C.; Autenrieth, I.B.; Schütz, M.; et al. Deprivation of the Periplasmic Chaperone SurA Reduces Virulence and Restores Antibiotic Susceptibility of Multidrug-Resistant Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 100. [Google Scholar] [CrossRef]
- Ebbensgaard, A.; Mordhorst, H.; Aarestrup, F.M.; Hansen, E.B. The role of outer membrane proteins and lipopolysaccharides for the sensitivity of escherichia coli to antimicrobial peptides. Front. Microbiol. 2018, 9, 2153. [Google Scholar] [CrossRef]
- Farhana, A.; Khan, Y.S. Biochemistry, Lipopolysaccharide; StatPearls Publishing: Tampa, FL, USA, 2020. [Google Scholar]
- Johnson, L.; Mulcahy, H.; Kanevets, U.; Shi, Y.; Lewenza, S. Surface-localized spermidine protects the Pseudomonas aeruginosa: Outer membrane from antibiotic treatment and oxidative stress. J. Bacteriol. 2012, 194, 813–826. [Google Scholar] [CrossRef]
- Hentzer, M.; Teitzel, G.M.; Balzer, G.J.; Heydorn, A.; Molin, S.; Givskov, M.; Parsek, M.R. Alginate overproduction affects pseudomonas aeruginosa biofilm structure and function. J. Bacteriol. 2001, 183, 5395–5401. [Google Scholar] [CrossRef]
- Jones, J.C.; Grotewold, N.; Wozniak, D.; Gloag, E. Pseudomonas aeruginosa Initiates a Rapid and Specific Transcriptional Response during Surface Attachment. J. Bacteriol. 2022, 204, e00086-22. [Google Scholar] [CrossRef]
- Persat, A.; Inclan, Y.F.; Engel, J.N.; Stone, H.A.; Gitai, Z. Type IV pili mechanochemically regulate virulence factors in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2015, 112, 7563–7568. [Google Scholar] [CrossRef]
- Shi, W.; Sun, H. Type IV pilus-dependent motility and its possible role in bacterial pathogenesis. Infect. Immun. 2002, 70, 1–4. [Google Scholar] [CrossRef]
- Beaussart, A.; Baker, A.E.; Kuchma, S.L.; El-Kirat-Chatel, S.; Otoole, G.A.; Dufrêne, Y.F. Nanoscale adhesion forces of Pseudomonas aeruginosa type IV pili. ACS Nano 2014, 8, 10723–10733. [Google Scholar] [CrossRef]
- Tammam, S.; Sampaleanu, L.M.; Koo, J.; Manoharan, K.; Daubaras, M.; Burrows, L.L.; Howell, P.L. PilMNOPQ from the Pseudomonas aeruginosa type IV pilus system form a transenvelope protein interaction network that interacts with PilA. J. Bacteriol. 2013, 195, 2126–2135. [Google Scholar] [CrossRef]
- Nguyen, Y.; Sugiman-Marangos, S.; Harvey, H.; Bell, S.D.; Charlton, C.L.; Junop, M.S.; Burrows, L.L. Pseudomonas aeruginosa minor pilins prime type IVa pilus assembly and promote surface display of the PilY1 adhesin. J. Biol. Chem. 2015, 290, 601–611. [Google Scholar] [CrossRef]
- Marko, V.A.; Kilmury, S.L.N.; MacNeil, L.T.; Burrows, L.L. Pseudomonas aeruginosa type IV minor pilins and PilY1 regulate virulence by modulating FimS-AlgR activity. PLoS Pathog. 2018, 14, e1007074. [Google Scholar] [CrossRef]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Microbiol. 2009, 7, 654–665. [Google Scholar] [CrossRef]
- Coburn, B.; Sekirov, I.; Finlay, B.B. Type III secretion systems and disease. Clin. Microbiol. Rev. 2007, 20, 535–549. [Google Scholar] [CrossRef]
- Dillon, M.M.; Almeida, R.N.D.; Laflamme, B.; Martel, A.; Weir, B.S.; Desveaux, D.; Guttman, D.S. Molecular evolution of Pseudomonas syringae type iii secreted effector proteins. Front. Plant Sci. 2019, 10, 148. [Google Scholar] [CrossRef]
- Rangel, S.M.; Diaz, M.H.; Knoten, C.A.; Zhang, A.; Hauser, A.R. The Role of ExoS in Dissemination of Pseudomonas aeruginosa during Pneumonia. PLoS Pathog. 2015, 11, e1004945. [Google Scholar] [CrossRef]
- Emma, K.; Josephine, T.; José, D.A.; Alicia, R.; Arne, R. PopB-PcrV Interactions Are Essential for Pore Formation in the Pseudomonas aeruginosa Type III Secretion System Translocon. MBio 2022, 13, e0238122. [Google Scholar]
- McCaw, M.L.; Lykken, G.L.; Singh, P.K.; Yahr, T.L. ExsD is a negative regulator of the Pseudomonas aeruginosa type III secretion regulon. Mol. Microbiol. 2002, 46, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Ye, M.; Li, X.; Li, J.; Deng, Z.; Yao, Y.-F.; Ou, H.Y. Identification and characterization of an antibacterial type VI secretion System in the carbapenem-resistant strain Klebsiella pneumoniae HS11286. Front. Cell. Infect. Microbiol. 2017, 7, 442. [Google Scholar] [CrossRef]
- Chen, L.; Zou, Y.; Kronfl, A.A.; Wu, Y. Type VI secretion system of Pseudomonas aeruginosa is associated with biofilm formation but not environmental adaptation. Microbiologyopen 2020, 9, e991. [Google Scholar] [CrossRef]
- Yoon, S.S.; Hennigan, R.F.; Hilliard, G.M.; Ochsner, U.A.; Parvatiyar, K.; Kamani, M.C.; Allen, H.L.; DeKievit, T.R.; Gardner, P.R.; Schwab, U.; et al. Pseudomonas aeruginosa anaerobic respiration in biofilms: Relationships to cystic fibrosis pathogenesis. Dev. Cell 2002, 3, 593–603. [Google Scholar] [CrossRef]
- Masamba, P.; Kappo, A.P. Parasite survival and disease persistence in cystic fibrosis, schistosomiasis and pathogenic bacterial diseases: A role for universal stress proteins? Int. J. Mol. Sci. 2021, 22, 10878. [Google Scholar] [CrossRef]
- Boes, N.; Schreiber, K.; Schobert, M. SpoT-triggered stringent response controls usp gene expression in Pseudomonas aeruginosa. J. Bacteriol. 2008, 190, 7189–7199. [Google Scholar] [CrossRef]
- Trunk, K.; Benkert, B.; Quäck, N.; Münch, R.; Scheer, M.; Garbe, J.; Jänsch, L.; Trost, M.; Wehland, J.; Buer, J.; et al. Anaerobic adaptation in Pseudomonas aeruginosa: Definition of the Anr and Dnr regulons. Environ. Microbiol. 2010, 12, 1719–1733. [Google Scholar] [CrossRef]
- Arat, S.; Bullerjahn, G.S.; Laubenbacher, R. A network biology approach to denitrification in Pseudomonas aeruginosa. PLoS ONE 2015, 10, e0118235. [Google Scholar] [CrossRef]
- Klünemann, T.; Preuß, A.; Adamczack, J.; Rosa, L.F.M.; Harnisch, F.; Layer, G.; Blankenfeldt, W. Crystal Structure of Dihydro-Heme d1 Dehydrogenase NirN from Pseudomonas aeruginosa Reveals Amino Acid Residues Essential for Catalysis. J. Mol. Biol. 2019, 431, 3246–3260. [Google Scholar] [CrossRef]
- Storbeck, S.; Saha, S.; Krausze, J.; Klink, B.U.; Heinz, D.W.; Layer, G. Crystal structure of the heme d 1 biosynthesis enzyme NirE in complex with its substrate reveals new insights into the catalytic mechanism of S-adenosyl-L-methionine-dependent uroporphyrinogen III methyltransferases. J. Biol. Chem. 2011, 286, 26754–26767. [Google Scholar] [CrossRef]
- Adamczack, J.; Hoffmann, M.; Papke, U.; Haufschildt, K.; Nicke, T.; Bröring, M.; Sezer, M.; Weimar, R.; Kuhlmann, U.; Hildebrandt, P.; et al. NirN protein from Pseudomonas aeruginosa is a novel electron-bifurcating dehydrogenase catalyzing the last step of Heme d1 biosynthesis. J. Biol. Chem. 2014, 289, 30753–30762. [Google Scholar] [CrossRef]
- Kawasaki, S.; Arai, H.; Kodama, T.; Igarashi, Y. Gene cluster for dissimilatory nitrite reductase (nir) from Pseudomonas aeruginosa: Sequencing and identification of a locus for heme d1 biosynthesis. J. Bacteriol. 1997, 179, 235–242. [Google Scholar] [CrossRef]
- Schreiber, K.; Krieger, R.; Benkert, B.; Eschbach, M.; Arai, H.; Schobert, M.; Jahn, D. The anaerobic regulatory network required for Pseudomonas aeruginosa nitrate respiration. J. Bacteriol. 2007, 189, 4310–4314. [Google Scholar] [CrossRef]
- Lee, Y.; Song, S.; Sheng, L.; Zhu, L.; Kim, J.S.; Wood, T.K. Substrate binding protein DppA1 of ABC transporter DppBCDF increases biofilm formation in Pseudomonas aeruginosa by inhibiting Pf5 prophage lysis. Front. Microbiol. 2018, 9, 30. [Google Scholar] [CrossRef]
- Kaiser, J.C.; Heinrichs, D.E. Branching out: Alterations in bacterial physiology and virulence due to branched-chain amino acid deprivation. MBio 2018, 9, e01188-18. [Google Scholar] [CrossRef]
- Puchalska, P.; Crawford, P.A. Multi-dimensional Roles of Ketone Bodies in Fuel Metabolism, Signaling, and Therapeutics. Cell Metab. 2017, 25, 262–284. [Google Scholar] [CrossRef]
- Wang, Z.; Bie, P.; Cheng, J.; Lu, L.; Cui, B.; Wu, Q. The ABC transporter YejABEF is required for resistance to antimicrobial peptides and the virulence of Brucella melitensis. Sci. Rep. 2016, 6, 31876. [Google Scholar] [CrossRef]
- López, C.A.; Travers, T.; Pos, K.M.; Zgurskaya, H.I.; Gnanakaran, S. Dynamics of Intact MexAB-OprM Efflux Pump: Focusing on the MexA-OprM Interface. Sci. Rep. 2017, 7, 16521. [Google Scholar] [CrossRef]
- Gotoh, N.; Tsujimoto, H.; Nomura, A.; Okamoto, K.; Tsuda, M.; Nishino, T. Functional replacement of OprJ by OprM in the MexCD-OprJ multidrug efflux system of Pseudomonas aeruginosa. FEMS Microbiol. Lett. 1998, 165, 21–27. [Google Scholar] [CrossRef]
- Wright, B.W.; Kamath, K.S.; Krisp, C.; Molloy, M.P. Proteome profiling of Pseudomonas aeruginosa PAO1 identifies novel responders to copper stress. BMC Microbiol. 2019, 19, 69. [Google Scholar] [CrossRef]
- Kwong, W.K.; Zheng, H.; Moran, N.A. Convergent evolution of a modified, acetate-driven TCA cycle in bacteria. Nat. Microbiol. 2017, 2, 17067. [Google Scholar] [CrossRef]
- Serbanescu, D.; Ojkic, N.; Banerjee, S. Cellular resource allocation strategies for cell size and shape control in bacteria. FEBS J. 2021, 289, 7891–7906. [Google Scholar] [CrossRef]
- Arenz, S.; Wilson, D.N. Bacterial protein synthesis as a target for antibiotic inhibition. Cold Spring Harb. Perspect. Med. 2016, 6, a025361. [Google Scholar] [CrossRef]
- Piatek, M.; O’Beirne, C.; Beato, Z.; Tacke, M.; Kavanagh, K. Exposure of Candida parapsilosis to the silver(I) compound SBC3 induces alterations in the proteome and reduced virulence. Metallomics 2022, 14, mfac046. [Google Scholar] [CrossRef]
- Rowland, E.A.; Snowden, C.K.; Cristea, I.M. Protein lipoylation: An evolutionarily conserved metabolic regulator of health and disease. Curr. Opin. Chem. Biol. 2018, 42, 75–85. [Google Scholar] [CrossRef]
- Teoh, W.P.; Chen, X.; Laczkovich, I.; Alonzo, F. Staphylococcus aureus adapts to the host nutritional landscape to overcome tissue-specific branched-chain fatty acid requirement. Proc. Natl. Acad. Sci. USA 2021, 118, e2022720118. [Google Scholar] [CrossRef]
- Grayczyk, J.P.; Harvey, C.J.; Laczkovich, I.; Alonzo, F. A Lipoylated Metabolic Protein Released by Staphylococcus aureus Suppresses Macrophage Activation. Cell Host Microbe 2017, 2, 678–687.e9. [Google Scholar] [CrossRef]
- Laczkovich, I.; Teoh, W.P.; Flury, S.; Grayczyk, J.P.; Zorzoli, A.; Alonzo, F. Increased flexibility in the use of exogenous lipoic acid by Staphylococcus aureus. Mol. Microbiol. 2018, 109, 150–168. [Google Scholar] [CrossRef]
- Cui, W.Q.; Qu, Q.W.; Wang, J.P.; Bai, J.W.; Bello-Onaghise, G.; Li, Y.A.; Zhou, Y.H.; Chen, X.R.; Liu, X.; Zheng, S.D.; et al. Discovery of potential anti-infective therapy targeting glutamine synthetase in Staphylococcus xylosus. Front. Chem. 2019, 7, 381. [Google Scholar] [CrossRef]
- Wang, H.; Wang, M.; Yang, X.; Xu, X.; Hao, Q.; Yan, A.; Hu, M.; Lobinski, R.; Li, H.; Sun, H. Antimicrobial silver targets glyceraldehyde-3-phosphate dehydrogenase in glycolysis of: E. coli. Chem. Sci. 2019, 10, 7193–7199. [Google Scholar] [CrossRef]
- Wang, H.; Wang, M.; Xu, X.; Gao, P.; Xu, Z.; Zhang, Q.; Li, H.; Yan, A.; Kao, R.Y.T.; Sun, H. Multi-target mode of action of silver against Staphylococcus aureus endows it with capability to combat antibiotic resistance. Nat. Commun. 2021, 12, 3331. [Google Scholar] [CrossRef]
- Tomlinson, K.L.; Lung, T.W.F.; Dach, F.; Annavajhala, M.K.; Gabryszewski, S.J.; Groves, R.A.; Drikic, M.; Francoeur, N.J.; Sridhar, S.H.; Smith, M.L.; et al. Staphylococcus aureus induces an itaconate-dominated immunometabolic response that drives biofilm formation. Nat. Commun. 2021, 12, 1399. [Google Scholar] [CrossRef]
- Margalit, A.; Carolan, J.C.; Sheehan, D.; Kavanagh, K. The Aspergillus fumigatus secretome alters the proteome of Pseudomonas aeruginosa to stimulate bacterial growth: Implications for co-infection. Mol. Cell. Proteom. 2020, 19, 1346–1359. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).