


New Evidence for the Role of Pituitary Adenylate Cyclase-Activating Polypeptide as an Antimicrobial Peptide in Teleost Fish

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Results
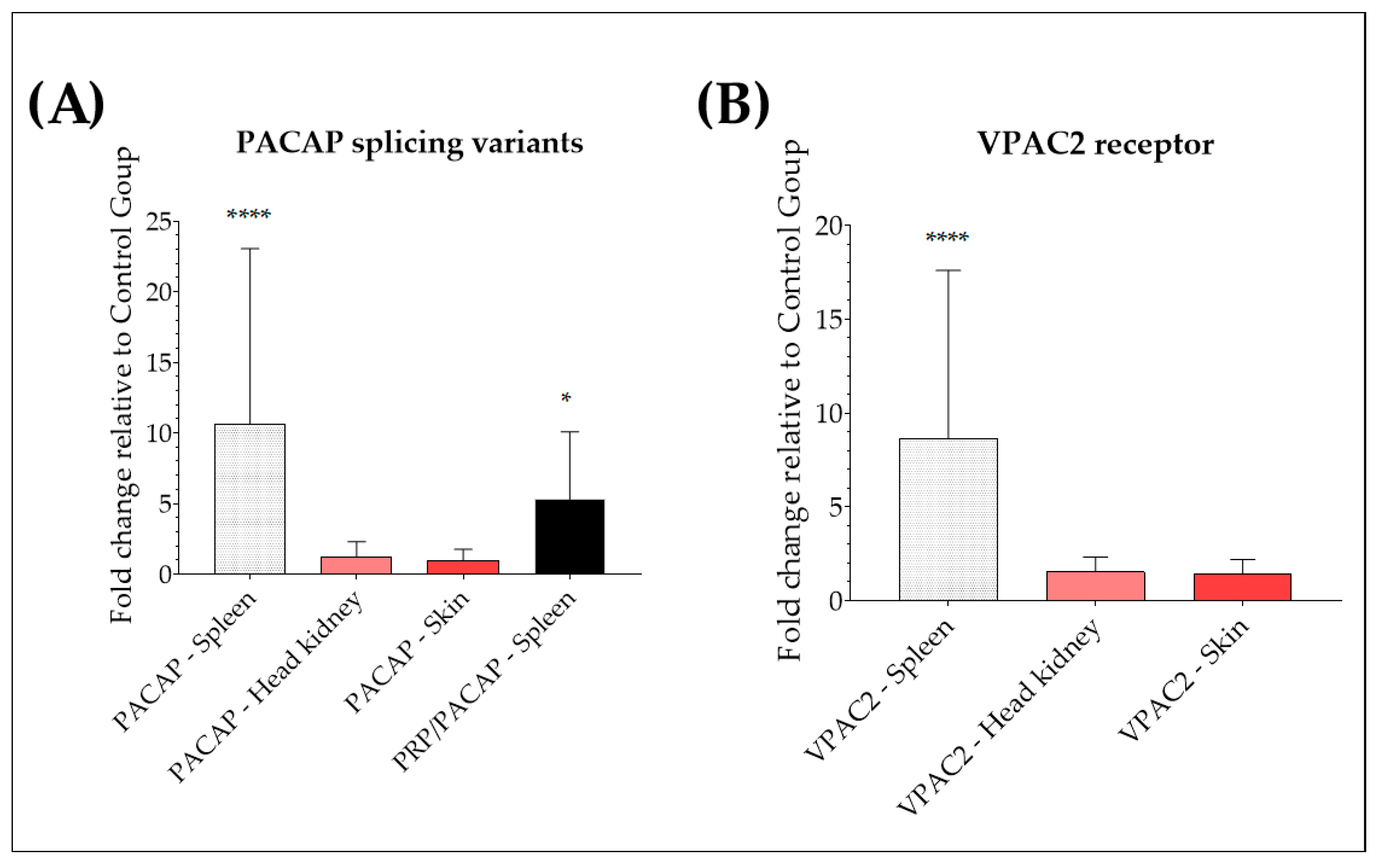
2.1. Up-Regulation of PACAP Splicing Variants and VPAC2 in the Spleen of Yersinia ruckeri-Infected Trout
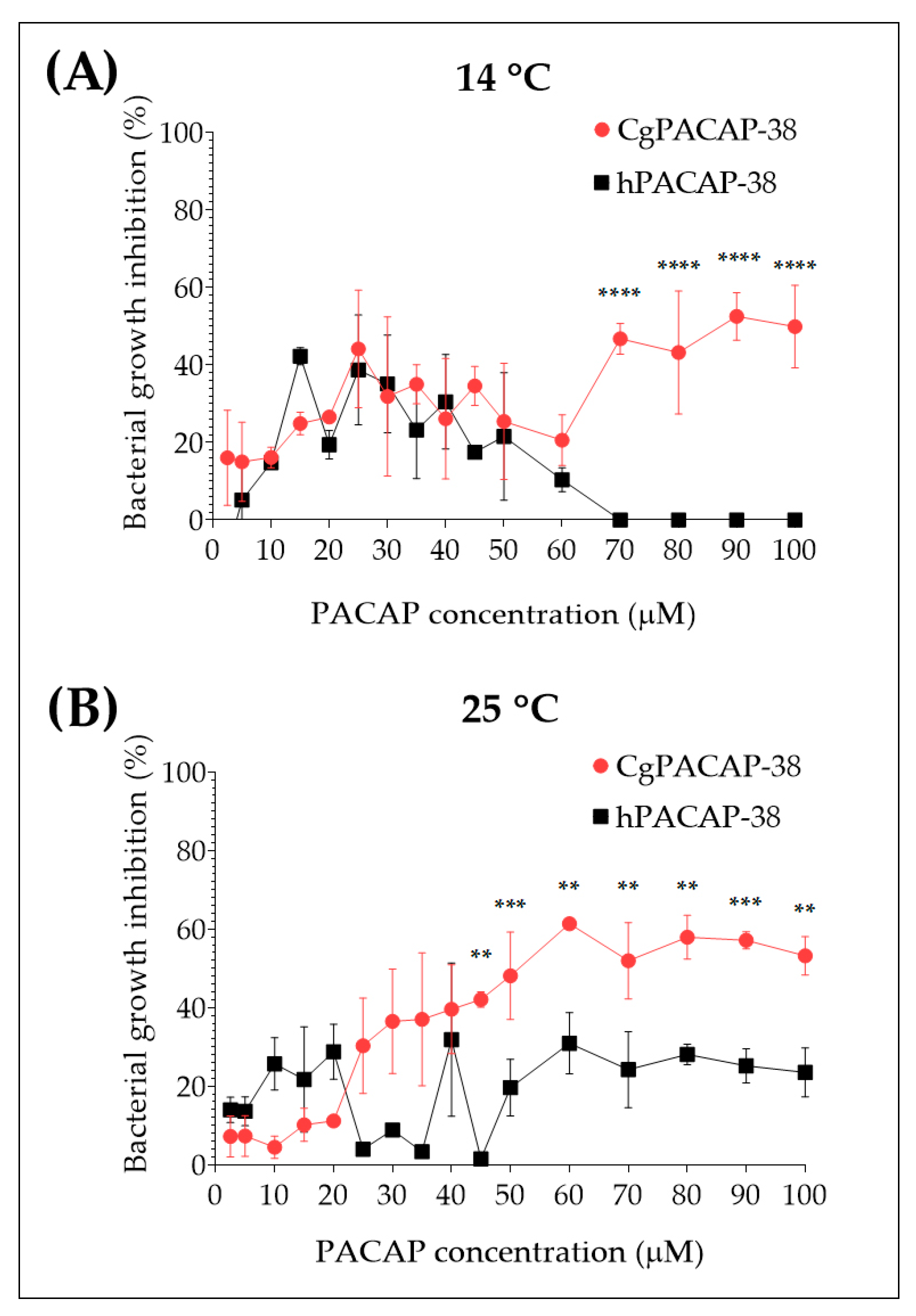
2.2. Direct Antimicrobial Activity of Synthetic PACAP-38 against Yersinia ruckeri in Tryptic Soy Broth
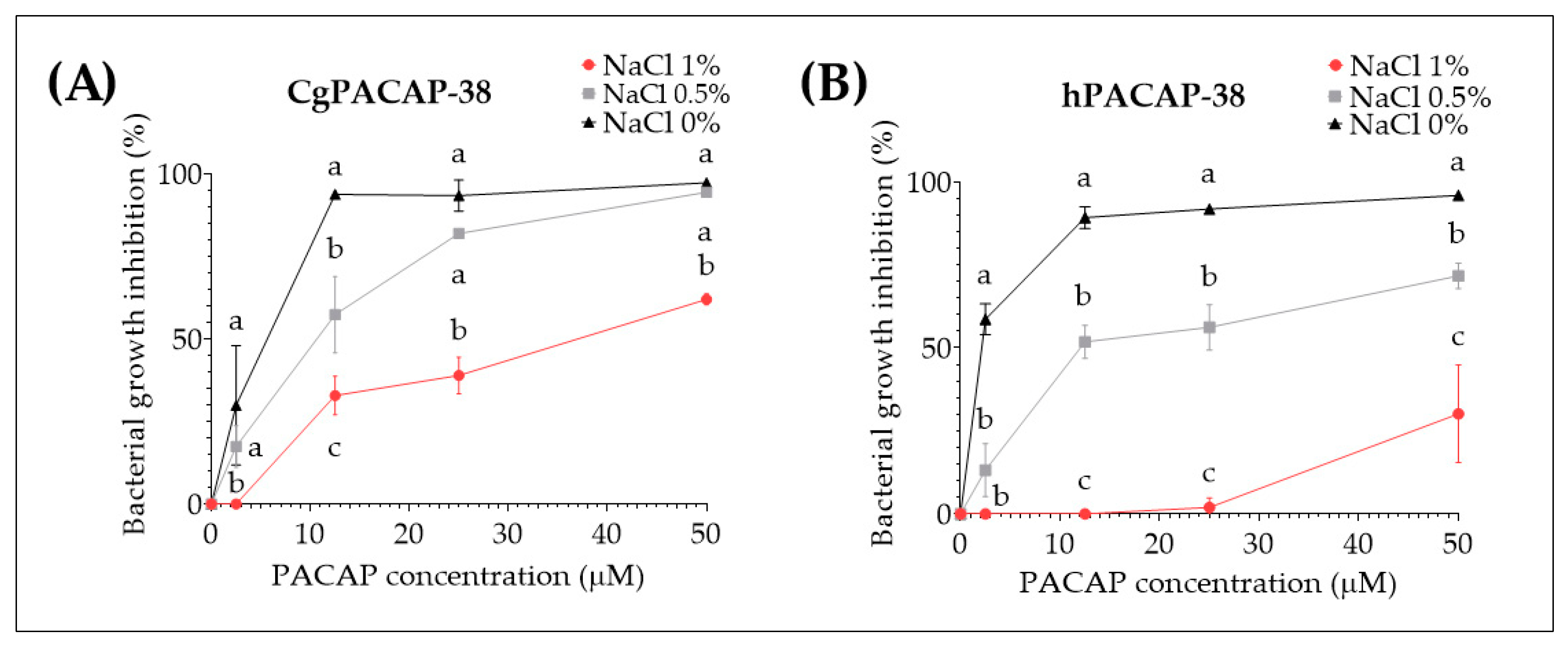
2.3. Effect of Different Salt Concentrations in Luria-Bertani Media on the Direct Antimicrobial Activity of Synthetic PACAP-38
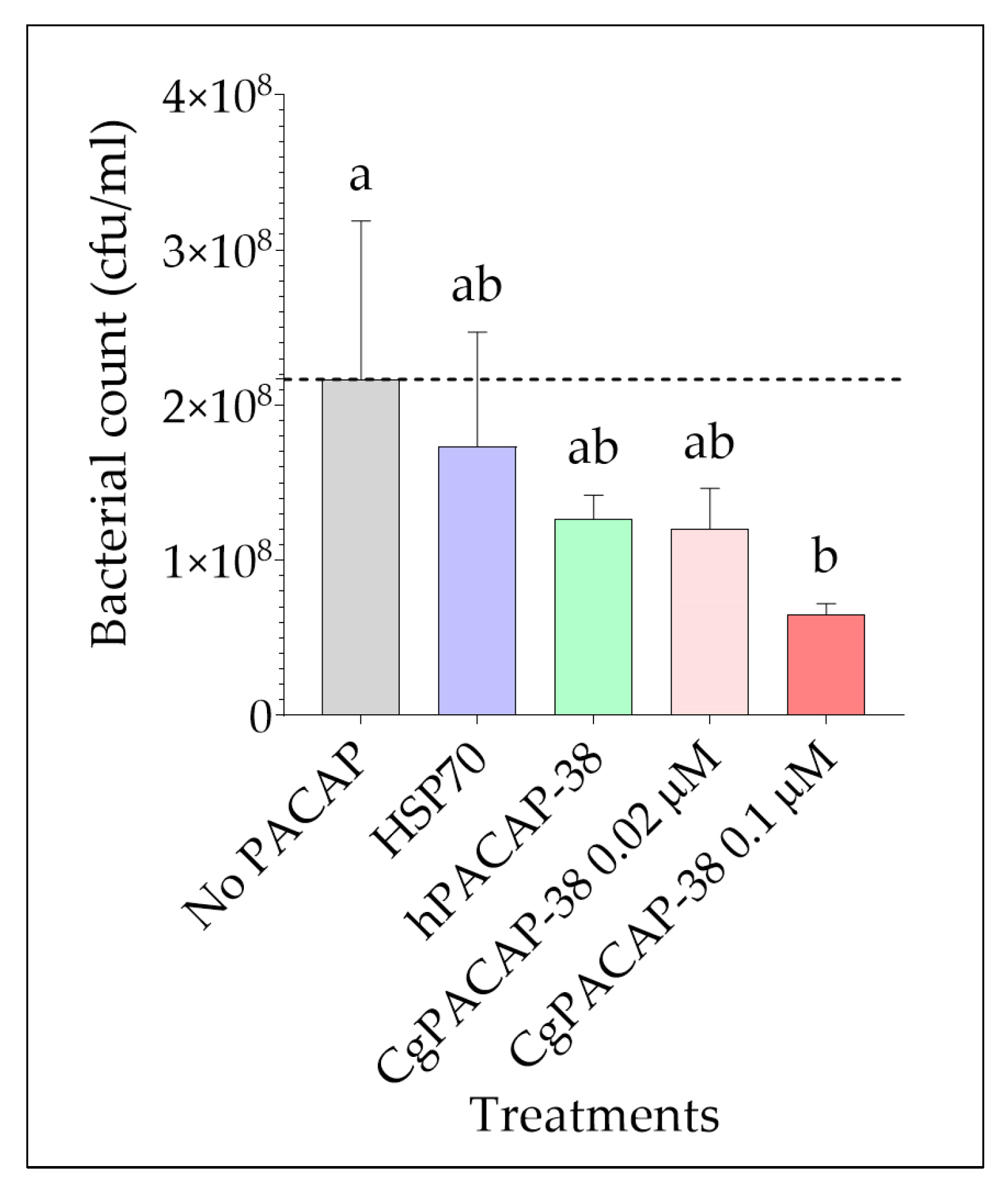
2.4. Effect of PACAP-38 on Yersinia ruckeri Bacterial Growth in Infected RTS11
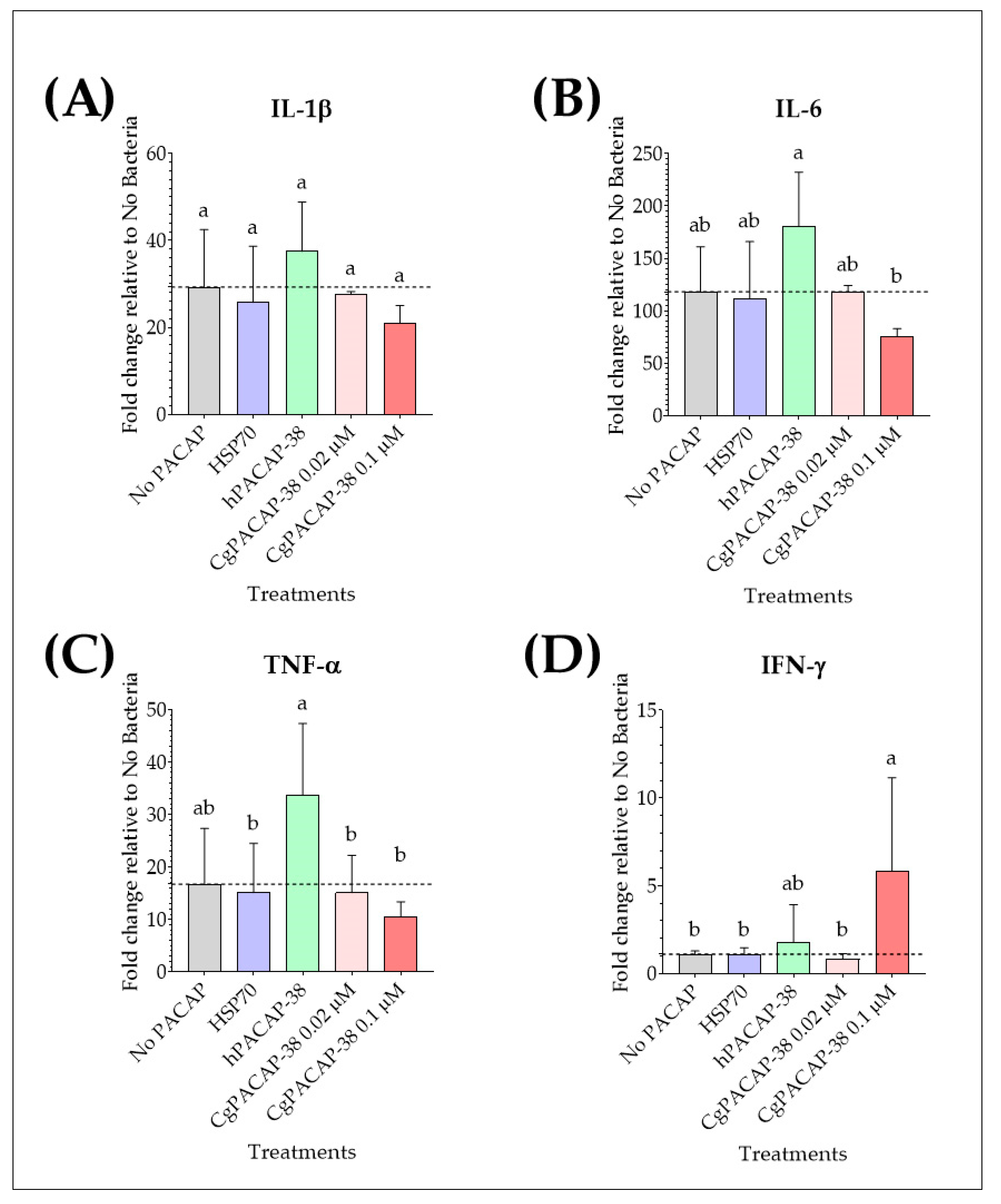
2.5. Effect of PACAP-38 on Pro-Inflammatory Cytokines in RTS11 Infected with Yersinia ruckeri
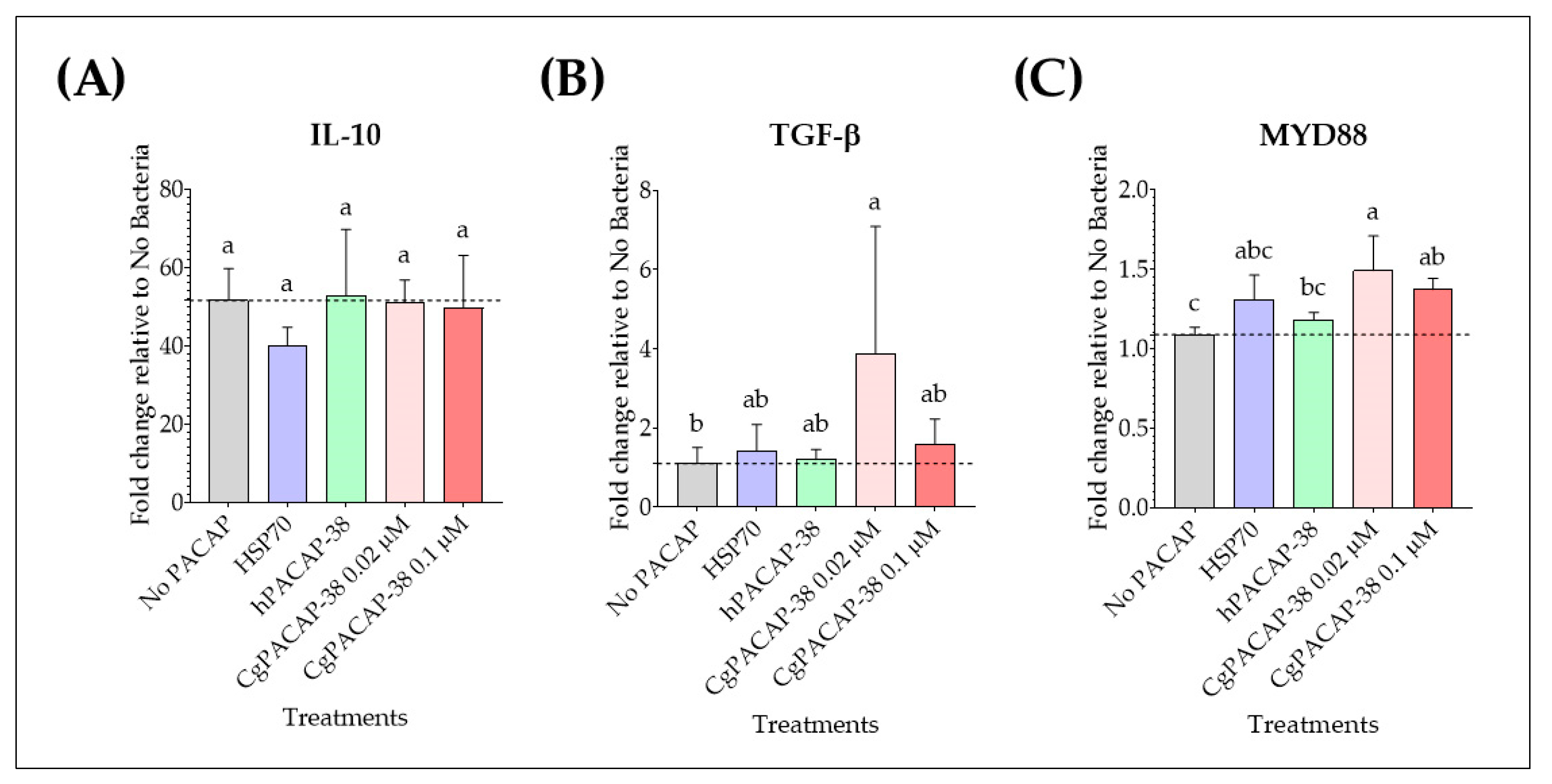
2.6. Effect of PACAP-38 on Anti-Inflammatory Cytokines, MYD88 Signal Transduction Adaptor Gene, and Antimicrobial Peptides in RTS11 Infected with Yersinia ruckeri
3. Discussion
4. Materials and Methods
4.1. Animals
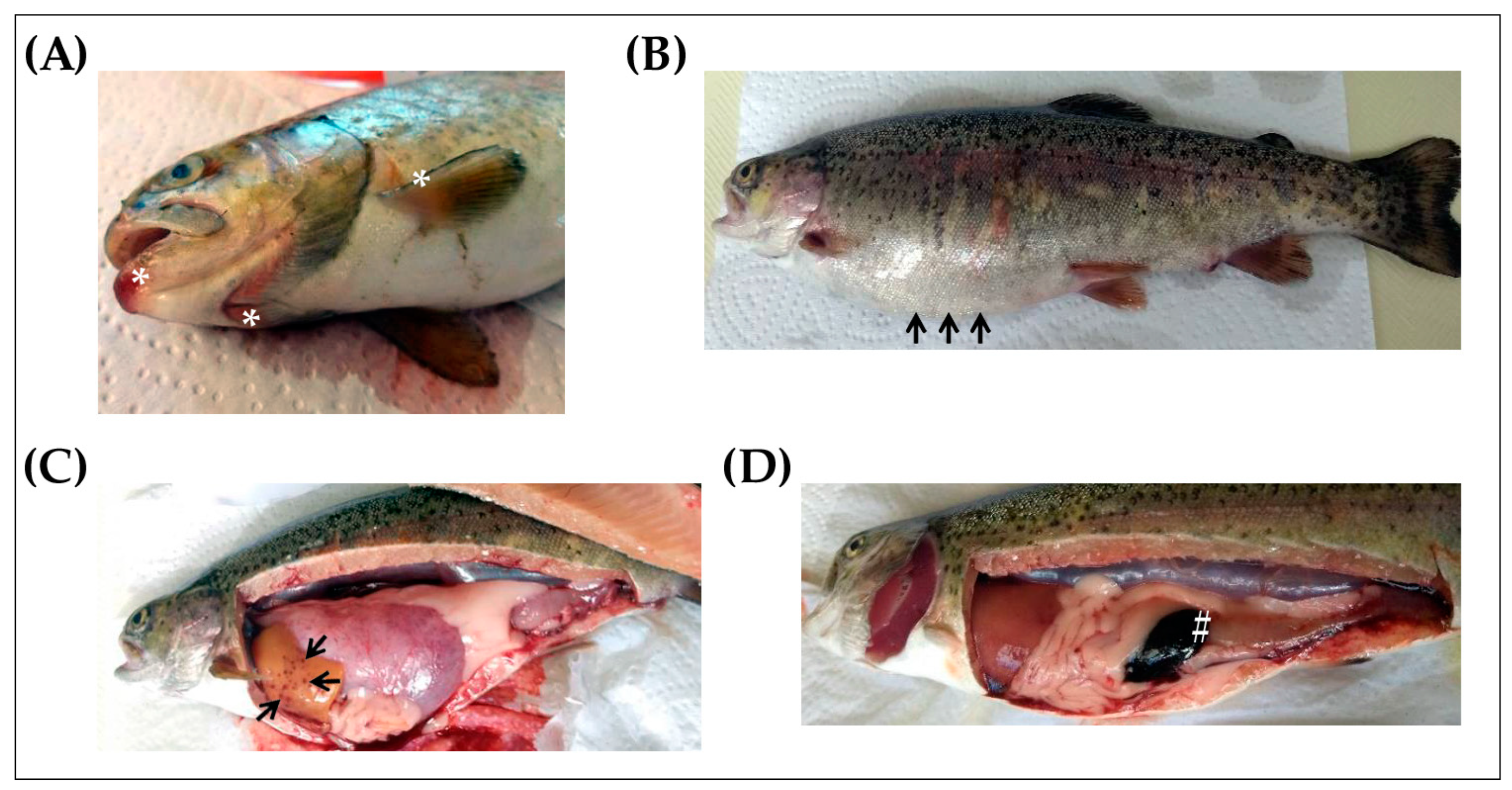
4.2. Sample Collection from Naturally Infected Fish and Clinically Healthy Trout
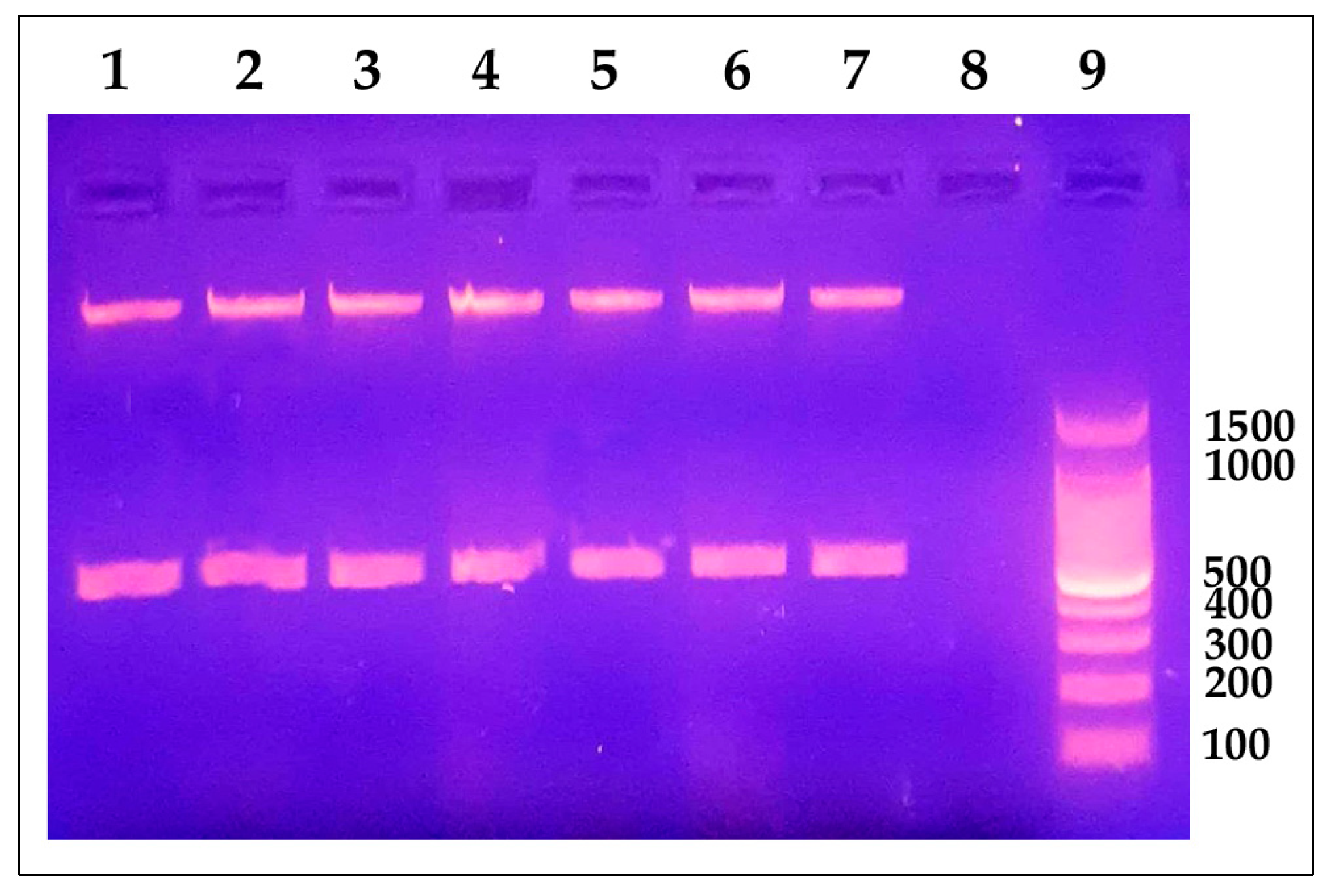
4.3. Identification of Y. ruckeri in Naturally Infected Fish and Clinically Healthy Trout
4.4. RNA Extraction and cDNA Synthesis
4.5. Gene Expression by qRT-PCR
4.6. In Vitro Trials to Determine the Activity of PACAP as an Antimicrobial and Immunostimulant Agent
4.6.1. Peptide Synthesis and Bacterial Strain
4.6.2. Direct Antibacterial Activity
4.6.3. Pre-Treatment of RTS11 with PACAP Followed by Infection with Live Y. ruckeri
4.6.4. RNA Extraction and cDNA Synthesis
4.6.5. Gene Expression by qRT-PCR
4.7. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Augustyniak, D.; Nowak, J.; T Lundy, F. Direct and Indirect Antimicrobial Activities of Neuropeptides and Their Therapeutic Potential. Curr. Protein Pept. Sci. 2012, 13, 723–738. [Google Scholar] [CrossRef] [PubMed]
- Starr, C.G.; Maderdrut, J.L.; He, J.; Coy, D.H.; Wimley, W.C. Pituitary Adenylate Cyclase-Activating Polypeptide Is a Potent Broad-Spectrum Antimicrobial Peptide: Structure-Activity Relationships. Peptides 2018, 104, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Toth, D.; Szabo, E.; Tamas, A.; Juhasz, T.; Horvath, G.; Fabian, E.; Opper, B.; Szabo, D.; Maugeri, G.; D’Amico, A.G. Protective Effects of PACAP in Peripheral Organs. Front. Endocrinol. 2020, 11, 377. [Google Scholar] [CrossRef] [PubMed]
- Gorgoglione, B.; Carpio, Y.; Secombes, C.J.; Taylor, N.G.; Lugo, J.M.; Estrada, M.P. Viral and Bacterial Septicaemic Infections Modulate the Expression of PACAP Splicing Variants and VIP/PACAP Receptors in Brown Trout Immune Organs. Fish Shellfish Immunol. 2015, 47, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Lugo, J.M.; Tafalla, C.; Oliva, A.; Pons, T.; Oliva, B.; Aquilino, C.; Morales, R.; Estrada, M.P. Evidence for Antimicrobial and Anticancer Activity of Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) from North African Catfish (Clarias gariepinus): Its Potential Use as Novel Therapeutic Agent in Fish and Humans. Fish Shellfish Immunol. 2019, 86, 559–570. [Google Scholar] [CrossRef]
- Semple, S.L.; Rodríguez-Ramos, T.; Carpio, Y.; Lumsden, J.S.; Estrada, M.P.; Dixon, B. PACAP Is Lethal to Flavobacterium psychrophilum Through Either Direct Membrane Permeabilization or Indirectly, by Priming the Immune Response in Rainbow Trout Macrophages. Front. Immunol. 2019, 10, 926. [Google Scholar] [CrossRef]
- Velázquez, J.; Pérez, G.; Semple, S.L.; Rodríguez-Ramos, T.; Díaz-Rosales, P.; Ordás, M.D.C.; Lugo, J.M.; Dixon, B.; Tafalla, C.; Estrada, M.P.; et al. First in Vivo Evidence of Pituitary Adenylate Cyclase-Activating Polypeptide Antiviral Activity in Teleost. Fish Shellfish Immunol. 2020, 103, 58–65. [Google Scholar] [CrossRef]
- Rodríguez, A.; Velázquez, J.; González, L.; Rodríguez-Ramos, T.; Dixon, B.; Miyares, F.H.; Morales, A.; González, O.; Estrada, M.P.; Carpio, Y. PACAP Modulates the Transcription of TLR-1/TLR-5/MyD88 Pathway Genes and Boosts Antimicrobial Defenses in Clarias gariepinus. Fish Shellfish Immunol. 2021, 115, 150–159. [Google Scholar] [CrossRef]
- Romalde, J.L.; MagariÑos, B.; Barja, J.L.; Toranzo, A.E. Antigenic and Molecular Characterization of Yersinia ruckeri Proposal for a New Intraspecies Classification. Syst. Appl. Microbiol. 1993, 16, 411–419. [Google Scholar] [CrossRef]
- Ross, A.; Rucker, R.; Ewing, W. Description of a Bacterium Associated with Redmouth Disease of Rainbow Trout (Salmo gairdneri). Can. J. Microbiol. 1966, 12, 763–770. [Google Scholar] [CrossRef]
- Furones, M.D.; Rodgers, C.J.; Munn, C.B. Yersinia ruckeri, the Causal Agent of Enteric Redmouth Disease (ERM) in Fish. Annu. Rev. Fish Dis. 1993, 3, 105–125. [Google Scholar]
- Kumar, G.; Menanteau-Ledouble, S.; Saleh, M.; El-Matbouli, M. Yersinia ruckeri, the Causative Agent of Enteric Redmouth Disease in Fish. Vet. Res. 2015, 46, 103. [Google Scholar] [CrossRef]
- Tobback, E.; Decostere, A.; Hermans, K.; Haesebrouck, F.; Chiers, K. Yersinia ruckeri Infections in Salmonid Fish. J. Fish Dis. 2007, 30, 257–268. [Google Scholar] [PubMed]
- Altinok, I. The Infectious Route of Yersinia ruckeri Is Affected by Salinity. Bull.-Eur. Assoc. Fish Pathol. 2004, 24, 253–259. [Google Scholar]
- Horne, M.; Barnes, A. Enteric Redmouth Disease (Yersinia ruckeri); CAB International: Wallingford, UK, 1999. [Google Scholar]
- Raida, M.; Buchmann, K. Temperature-Dependent Expression of Immune-Relevant Genes in Rainbow Trout Following Yersinia ruckeri Vaccination. Dis. Aquat. Organ. 2007, 77, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Raida, M.K.; Buchmann, K. Bath Vaccination of Rainbow Trout (Oncorhynchus mykiss Walbaum) against Yersinia ruckeri: Effects of Temperature on Protection and Gene Expression. Vaccine 2008, 26, 1050–1062. [Google Scholar] [CrossRef]
- Raida, M.K.; Buchmann, K. Development of Adaptive Immunity in Rainbow Trout, Oncorhynchus mykiss (Walbaum) Surviving an Infection with Yersinia ruckeri. Fish Shellfish Immunol. 2008, 25, 533–541. [Google Scholar] [CrossRef]
- Raida, M.K.; Nylén, J.; Holten-Andersen, L.; Buchmann, K. Association between Plasma Antibody Response and Protection in Rainbow Trout Oncorhynchus mykiss Immersion Vaccinated against Yersinia ruckeri. PLoS ONE 2011, 6, e18832. [Google Scholar] [CrossRef]
- Raida, M.K.; Holten-Andersen, L.; Buchmann, K. Association between Yersinia Ruckeri Infection, Cytokine Expression and Survival in Rainbow Trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2011, 30, 1257–1264. [Google Scholar] [CrossRef]
- Skov, J.; Kania, P.W.; Holten-Andersen, L.; Fouz, B.; Buchmann, K. Immunomodulatory Effects of Dietary β-1,3-Glucan from Euglena Gracilis in Rainbow Trout (Oncorhynchus mykiss) Immersion Vaccinated against Yersinia ruckeri. Fish Shellfish Immunol. 2012, 33, 111–120. [Google Scholar] [CrossRef]
- Naderi Farsani, M.; Hoseinifar, S.H.; Rashidian, G.; Ghafari Farsani, H.; Ashouri, G.; Van Doan, H. Dietary Effects of Coriandrum Sativum Extract on Growth Performance, Physiological and Innate Immune Responses and Resistance of Rainbow Trout (Oncorhynchus mykiss) against Yersinia ruckeri. Fish Shellfish Immunol. 2019, 91, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Terzi, E.; Kucukkosker, B.; Bilen, S.; Kenanoglu, O.N.; Corum, O.; Özbek, M.; Parug, S.S. A Novel Herbal Immunostimulant for Rainbow Trout (Oncorhynchus mykiss) against Yersinia ruckeri. Fish Shellfish Immunol. 2021, 110, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Stone, M. The Use of Prebiotics to Improve Fish Health. Doctoral Dissertation, University of Plymouth, Plymouth, UK, 2005. [Google Scholar]
- Kim, D.-H.; Austin, B. Innate Immune Responses in Rainbow Trout (Oncorhynchus mykiss, Walbaum) Induced by Probiotics. Fish Shellfish Immunol. 2006, 21, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Soltani, M.; Pakzad, K.; Taheri-Mirghaed, A.; Mirzargar, S.; Shekarabi, S.P.H.; Yosefi, P.; Soleymani, N. Dietary Application of the Probiotic Lactobacillus plantarum 426951 Enhances Immune Status and Growth of Rainbow Trout (Oncorhynchus mykiss) Vaccinated Against Yersinia ruckeri. Probiotics Antimicrob. Proteins 2019, 11, 207–219. [Google Scholar] [CrossRef]
- Quintanilla-Pineda, M.; Achou, C.; Díaz, J.; Gutiérrez-Falcon, A.; Bravo, M.; Herrera-Muñoz, J.; Peña-Navarro, N.; Alvarado, C.; Ibañez, F.; Marzo, F. In Vitro Evaluation of Postbiotics Produced from Bacterial Isolates Obtained from Rainbow Trout and Nile Tilapia against the Pathogens Yersinia ruckeri and Aeromonas salmonicida Subsp. salmonicida. Foods 2023, 12, 861. [Google Scholar] [CrossRef]
- Yang, H.; Zhujin, D.; Marana, M.H.; Dalsgaard, I.; Rzgar, J.; Heidi, M.; Asma, K.M.; Per, K.W.; Kurt, B. Immersion Vaccines against Yersinia ruckeri Infection in Rainbow Trout: Comparative Effects of Strain Differences. J. Fish Dis. 2021, 44, 1937–1950. [Google Scholar] [CrossRef]
- Pajdak-Czaus, J.; Platt-Samoraj, A.; Szweda, W.; Siwicki, A.K.; Terech-Majewska, E. Yersinia ruckeri—A Threat Not Only to Rainbow Trout. Aquac. Res. 2019, 50, 3083–3096. [Google Scholar] [CrossRef]
- Chettri, J.K.; Mehrdana, F.; Hansen, E.B.; Ebbensgaard, A.; Overgaard, M.; Lauritsen, A.; Dalsgaard, I.; Buchmann, K. Antimicrobial Peptide CAP 18 and Its Effect on Yersinia Ruckeri Infections in Rainbow Trout Oncorhynchus mykiss (Walbaum): Comparing Administration by Injection and Oral Routes. J. Fish Dis. 2017, 40, 97–104. [Google Scholar] [CrossRef]
- Cardoso, J.C.; Garcia, M.G.; Power, D.M. Tracing the Origins of the Pituitary Adenylate-Cyclase Activating Polypeptide (PACAP). Front. Neurosci. 2020, 14, 366. [Google Scholar] [CrossRef]
- Carpio, Y.; Lugo, J.M.; León, K.; Morales, R.; Estrada, M.P. Novel Function of Recombinant Pituitary Adenylate Cyclase-Activating Polypeptide as Stimulator of Innate Immunity in African Catfish (Clarias gariepinus) Fry. Fish Shellfish Immunol. 2008, 25, 439–445. [Google Scholar] [CrossRef]
- Lugo, J.M.; Carpio, Y.; Oliva, A.; Morales, A.; Estrada, M.P. Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP): A Regulator of the Innate and Acquired Immune Functions in Juvenile Fish. Fish Shellfish Immunol. 2010, 29, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Lugo, J.M.; Oliva, A.; Morales, A.; Reyes, O.; Garay, H.E.; Herrera, F.; Cabrales, A.; Pérez, E.; Estrada, M.P. The Biological Role of Pituitary Adenylate Cyclase-activating Polypeptide (PACAP) in Growth and Feeding Behavior in Juvenile Fish. J. Pept. Sci. 2010, 16, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Herrera, F.; Velázquez, J.; Lugo, J.M.; Orellana, P.; Ruiz, J.; Vega, M.; Romero, A.; Santos, N.; Ramsés, G.; Rodríguez-Ramos, T.; et al. Oral Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) Formulation Modified Muscle Fatty Acid Profile and Cytokines Transcription in Head Kidney in Rainbow Trout (Oncorhynchus mykiss) Fingerlings. Aquac. Rep. 2021, 20, 100772. [Google Scholar] [CrossRef]
- Holm, G.E.; Sherman, J.M. Salt effects in bacterial growth I. preliminary paper. J. Bacteriol. 1921, 6, 511–519. [Google Scholar] [CrossRef]
- Peters, A.C.; Thomas, L.; Wimpenny, J.W.T. Effects of Salt Concentration on Bacterial Growth on Plates with Gradients of PH and Temperature. FEMS Microbiol. Lett. 1991, 77, 309–314. [Google Scholar] [CrossRef][Green Version]
- Breidt, F.; Romick, T.L.; Fleming, H.P. A rapid method for the determination of bacterial growth kinetics. J. Rapid Methods Autom. Microbiol. 1994, 3, 59–68. [Google Scholar] [CrossRef]
- Porter, E.M.; Van Dam, E.; Valore, E.V.; Ganz, T. Broad-Spectrum Antimicrobial Activity of Human Intestinal Defensin 5. Infect. Immun. 1997, 65, 2396–2401. [Google Scholar] [CrossRef]
- Wangkahart, E.; Scott, C.; Secombes, C.J.; Wang, T. Re-Examination of the Rainbow Trout (Oncorhynchus mykiss) Immune Response to Flagellin: Yersinia ruckeri Flagellin Is a Potent Activator of Acute Phase Proteins, Anti-Microbial Peptides and pro-Inflammatory Cytokines in vitro. Dev. Comp. Immunol. 2016, 57, 75–87. [Google Scholar] [CrossRef]
- Gomariz, R.P.; Juarranz, Y.; Abad, C.; Arranz, A.; Leceta, J.; Martinez, C. VIP–PACAP System in Immunity. Ann. N. Y. Acad. Sci. 2006, 1070, 51–74. [Google Scholar] [CrossRef]
- Flaishon, L.; Topilski, I.; Shoseyov, D.; Hershkoviz, R.; Fireman, E.; Levo, Y.; Marmor, S.; Shachar, I. Cutting Edge: Anti-Inflammatory Properties of Low Levels of IFN-γ. J. Immunol. 2002, 168, 3707–3711. [Google Scholar] [CrossRef]
- Wiegertjes, G.F.; Wentzel, A.S.; Spaink, H.P.; Elks, P.M.; Fink, I.R. Polarization of Immune Responses in Fish: The ‘Macrophages First’ Point of View. Mol. Immunol. 2016, 69, 146–156. [Google Scholar] [CrossRef]
- Gonzalez-Rey, E. Keeping the Balance between Immune Tolerance and Pathogen Immunity with Endogenous Neuropeptides. Neuroimmunomodulation 2010, 17, 161–164. [Google Scholar] [CrossRef]
- Langer, I.; Jeandriens, J.; Couvineau, A.; Sanmukh, S.; Latek, D. Signal Transduction by VIP and PACAP Receptors. Biomedicines 2022, 10, 406. [Google Scholar] [CrossRef] [PubMed]
- Santana, P.A.; Guzmán, F.; Forero, J.C.; Luna, O.F.; Mercado, L. Hepcidin, Cathelicidin-1 and IL-8 as Immunological Markers of Responsiveness in Early Developmental Stages of Rainbow Trout. Dev. Comp. Immunol. 2016, 62, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Chettri, J.K.; Raida, M.K.; Kania, P.W.; Buchmann, K. Differential Immune Response of Rainbow Trout (Oncorhynchus mykiss) at Early Developmental Stages (Larvae and Fry) against the Bacterial Pathogen Yersinia Ruckeri. Dev. Comp. Immunol. 2012, 36, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Zhang, L.; Chen, P. Membrane Internalization Mechanisms and Design Strategies of Arginine-Rich Cell-Penetrating Peptides. Int. J. Mol. Sci. 2022, 23, 9038. [Google Scholar] [CrossRef] [PubMed]
- American Veterinary Medical Association. Guidelines for the Euthanasia of Animals; AVMA: Schaumbg, IL, USA, 2013. [Google Scholar]
- Roberts, J.C.; Syme, D.A. Effects of Using Tricaine Methanesulfonate and Metomidate before Euthanasia on the Contractile Properties of Rainbow Trout (Oncorhynchus Mykiss) Myocardium. J. Am. Assoc. Lab. Anim. Sci. 2016, 55, 565–569. [Google Scholar]
- Meyers, T.R. Standard Necropsy Procedures for Finfish. In NWFHS Laboratory Procedures Manual, 5th ed.; US Fish and Wildlife Service: Washington, DC, USA, 2009; pp. 64–74. [Google Scholar]
- Gibello, A.; Blanco, M.; Moreno, M.; Cutuli, M.; Domenech, A.; Domínguez, L.; Fernández-Garayzábal, J. Development of a PCR Assay for Detection of Yersinia Ruckeri in Tissues of Inoculated and Naturally Infected Trout. Appl. Environ. Microbiol. 1999, 65, 346–350. [Google Scholar] [CrossRef]
- Lugo, J.M.; Tafalla, C.; Leceta, J.; Gomariz, R.P.; Estrada, M.P. Differential Expression Pattern of Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP) Alternative Splicing Variants and Its Receptors in the Immune System of Rainbow Trout (Oncorhynchus Mykiss). Fish Shellfish Immunol. 2011, 30, 734–738. [Google Scholar] [CrossRef]
- Chaves-Pozo, E.; Zou, J.; Secombes, C.J.; Cuesta, A.; Tafalla, C. The Rainbow Trout (Oncorhynchus Mykiss) Interferon Response in the Ovary. Mol. Immunol. 2010, 47, 1757–1764. [Google Scholar] [CrossRef]
- Inoue, Y.; Kamota, S.; Ito, K.; Yoshiura, Y.; Ototake, M.; Moritomo, T.; Nakanishi, T. Molecular Cloning and Expression Analysis of Rainbow Trout (Oncorhynchus Mykiss) Interleukin-10 CDNAs. Fish Shellfish Immunol. 2005, 4, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Broekman, D.C.; Guðmundsson, G.H.; Maier, V.H. Differential Regulation of Cathelicidin in Salmon and Cod. Fish Shellfish Immunol. 2013, 35, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Otvos, L.; Cudic, M. Broth Microdilution Antibacterial Assay of Peptides. In Peptide Characterization and Application Protocols; Humana Press Inc.: Totowa, NJ, USA, 2007; pp. 309–320. [Google Scholar]
- Sever, L.; Vo, N.T.; Bols, N.C.; Dixon, B. Expression of Tapasin in Rainbow Trout Tissues and Cell Lines and up Regulation in a Monocyte/Macrophage Cell Line (RTS11) by a Viral Mimic and Viral Infection. Dev. Comp. Immunol. 2014, 44, 86–93. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ to 3′) * | Reference |
|---|---|---|
| Y. ruckeri 16S rRNA | F: GCGAGGAGGAAGGGTTAAGTG R: GAAGGCACCAAGGCATCTCTG | Gibello et al. [52] |
| PACAP | F: AAATTGCTATAAGAAGTCCCCCATC R: GTATTTCTTGACTGCCATTTGCTTT | Lugo et al. [53] |
| PRP/PACAP | F: CTGGGTCAGTTATCAGCAAGAAAT R: TGTCTATACCTTTTCCCAAGGACTG | Lugo et al. [53] |
| VPAC2 | F: CTCACTGTGACACGACAGTGATTCC R: GCTTTTCAGTTTCACCTCAACTTGT | Lugo et al. [53] |
| IL-1β | F: CCACAAAGTGCATTTGAAC R: GCAACCTCCTCTAGGTGC | Semple et al. [6] |
| IL-6 | F: CTTCTACACGCTATCTCTCACTC R: CGTCTGTCCCGAGCT | Semple et al. [6] |
| TNF-α | F: GTGCAAAAGATACCCACC R: CACTGCACGGTGTCAG | Semple et al. [6] |
| IFN-γ | F: GAAGGCTCTGTCCGAGTTCA R: TGTGTGATTTGAGCCTCTGG | Chaves-Pozo et al. [54] |
| IL-10 | F: GCCTTCTCCACCATCAGAGAC R: GATGCTGTCCATAGCGTGAC | Inoue et al. [55] |
| TGF-β | F: TGTGGGGAGACAACACAAGG R: AAACCAGCGCCATCAAAAAGG | This study |
| MYD88 | F: GACAAAGTTTGCCCTCAGTCTCT R: CCGTCAGGAACCTCAGGATACT | This study |
| Cathelicidin-1 | F: ATGGGAAACTAATGATGTGC R: CGGTCAGTGTTGAGGGTATT | Broekman et al. [56] |
| Hepcidin | F: GCTTCTGCTGCAAATTCTGAGG R: GTACAAGACTGAGGTTGTGCAG | This study |
| EF-1α | F: CGCACAGTAACACCGAAACTAATTAAGC R: GCCTCCGCACTTGTAGATCAGATG | Semple et al. [6] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velázquez, J.; Rodríguez-Cornejo, T.; Rodríguez-Ramos, T.; Pérez-Rodríguez, G.; Rivera, L.; Campbell, J.H.; Al-Hussinee, L.; Carpio, Y.; Estrada, M.P.; Dixon, B. New Evidence for the Role of Pituitary Adenylate Cyclase-Activating Polypeptide as an Antimicrobial Peptide in Teleost Fish. Antibiotics 2023, 12, 1484. https://doi.org/10.3390/antibiotics12101484
Velázquez J, Rodríguez-Cornejo T, Rodríguez-Ramos T, Pérez-Rodríguez G, Rivera L, Campbell JH, Al-Hussinee L, Carpio Y, Estrada MP, Dixon B. New Evidence for the Role of Pituitary Adenylate Cyclase-Activating Polypeptide as an Antimicrobial Peptide in Teleost Fish. Antibiotics. 2023; 12(10):1484. https://doi.org/10.3390/antibiotics12101484
Chicago/Turabian StyleVelázquez, Janet, Tania Rodríguez-Cornejo, Tania Rodríguez-Ramos, Geysi Pérez-Rodríguez, Laura Rivera, James Hugh Campbell, Lowia Al-Hussinee, Yamila Carpio, Mario Pablo Estrada, and Brian Dixon. 2023. "New Evidence for the Role of Pituitary Adenylate Cyclase-Activating Polypeptide as an Antimicrobial Peptide in Teleost Fish" Antibiotics 12, no. 10: 1484. https://doi.org/10.3390/antibiotics12101484
APA StyleVelázquez, J., Rodríguez-Cornejo, T., Rodríguez-Ramos, T., Pérez-Rodríguez, G., Rivera, L., Campbell, J. H., Al-Hussinee, L., Carpio, Y., Estrada, M. P., & Dixon, B. (2023). New Evidence for the Role of Pituitary Adenylate Cyclase-Activating Polypeptide as an Antimicrobial Peptide in Teleost Fish. Antibiotics, 12(10), 1484. https://doi.org/10.3390/antibiotics12101484