
Impact of Raised without Antibiotics Measures on Antimicrobial Resistance and Prevalence of Pathogens in Sow Barns
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Antifolates Are Largely Administered to Non-RWA Sows While β-Lactams Are Still Widely Used for Treatment of Sows in RWA Barns
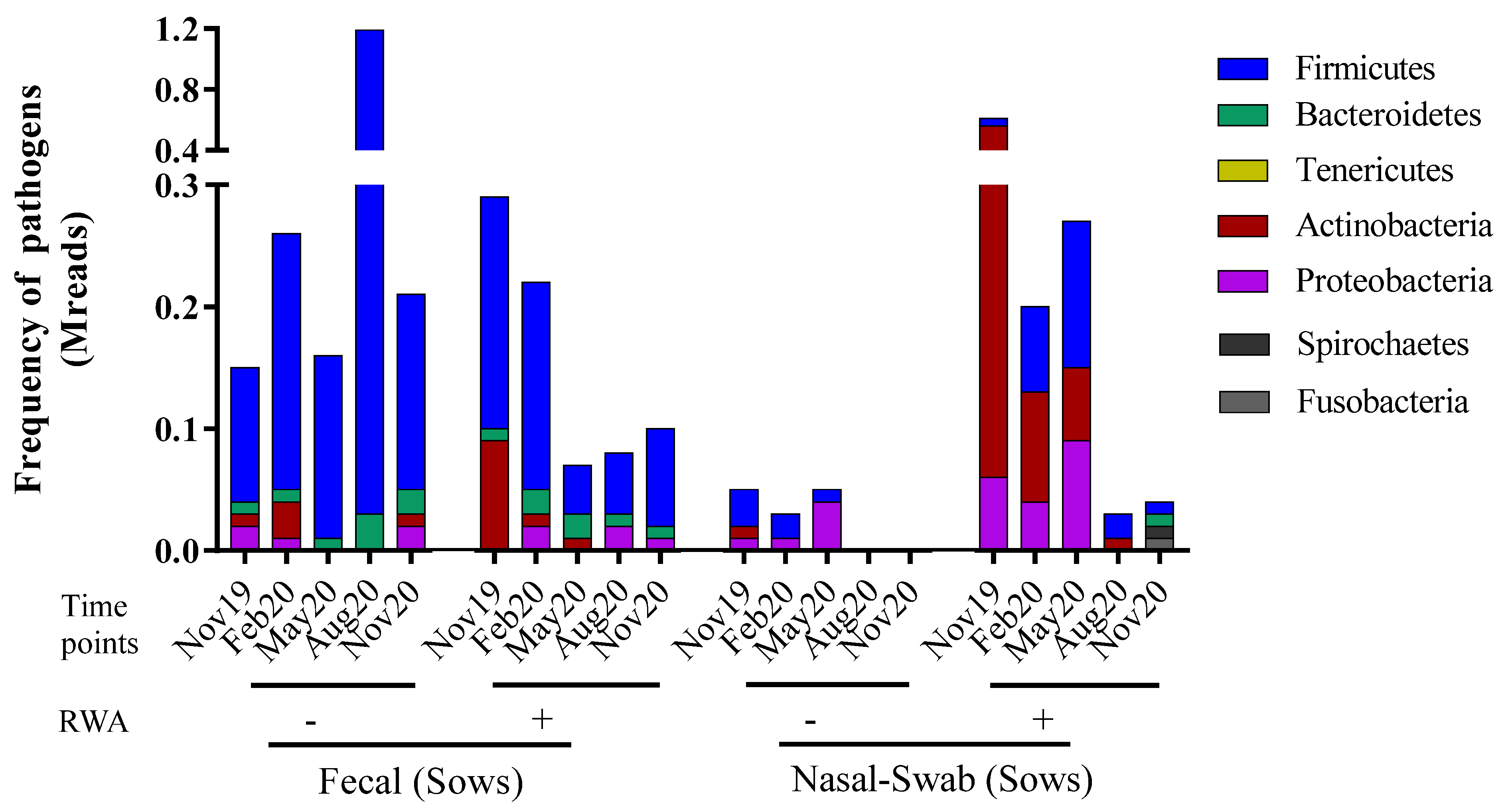
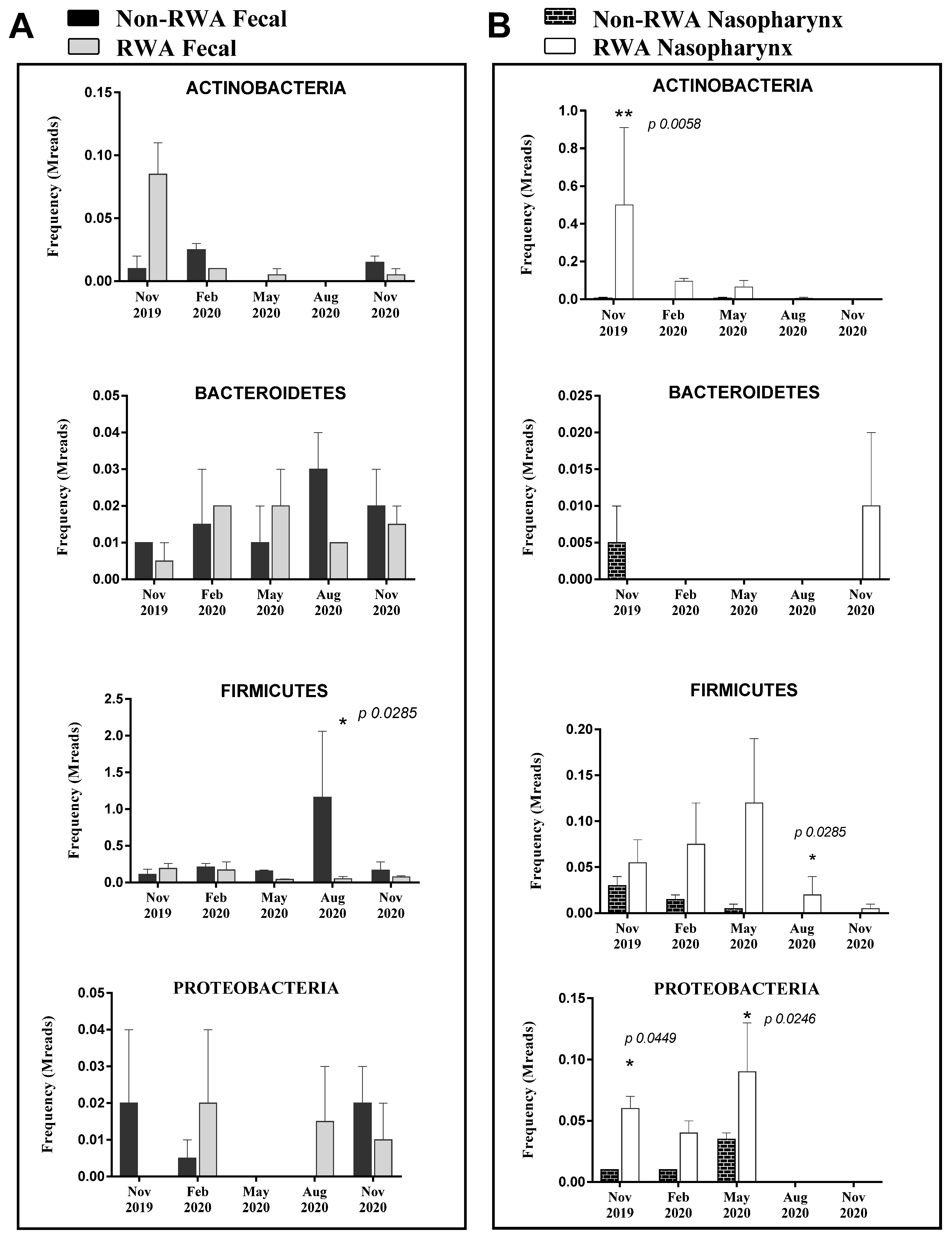
2.2. RWA Sows Exhibit More Pathogenic Actinobacteria, Firmicutes, and Proteobacteria in Nasopharynx Samples
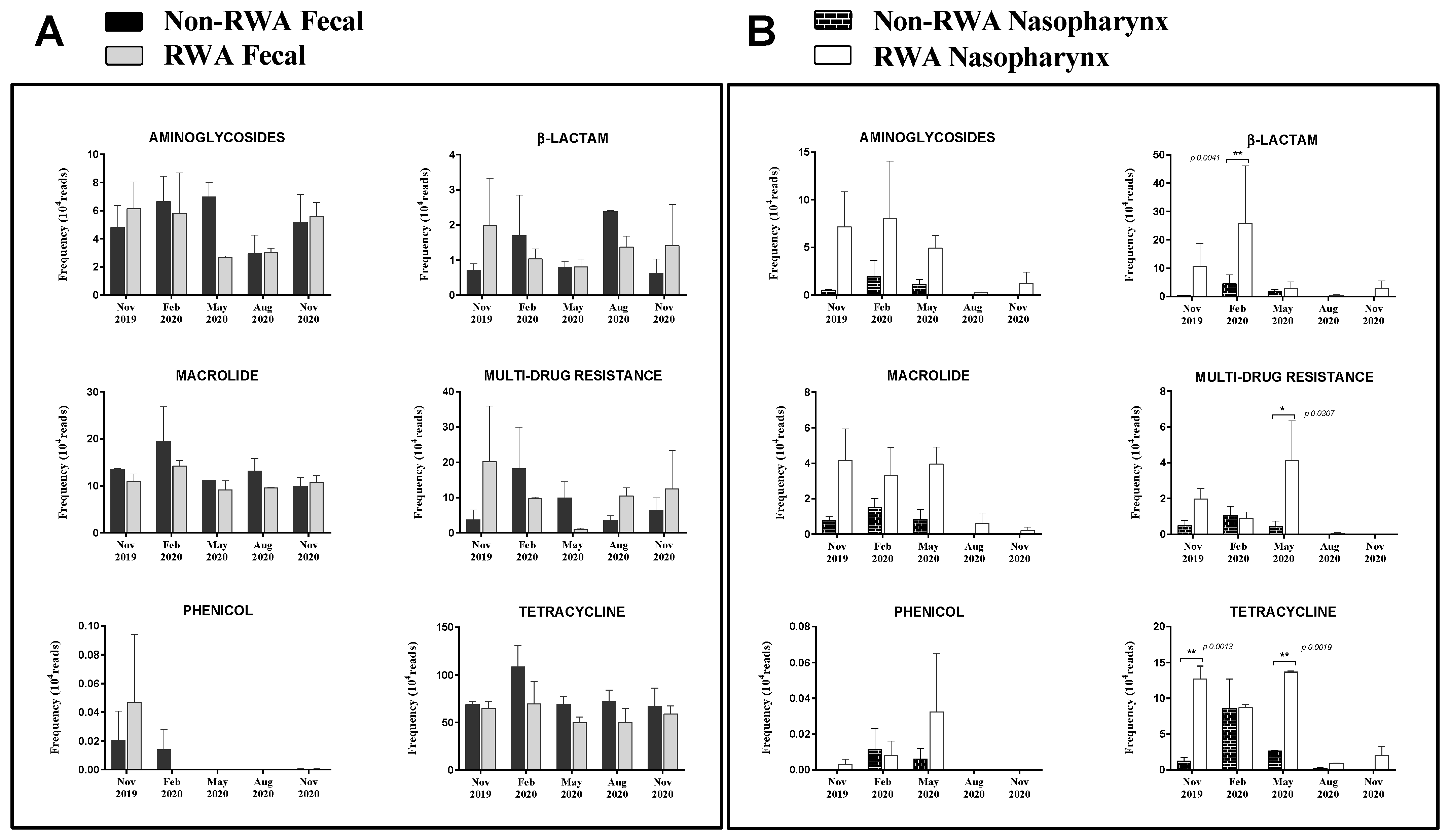
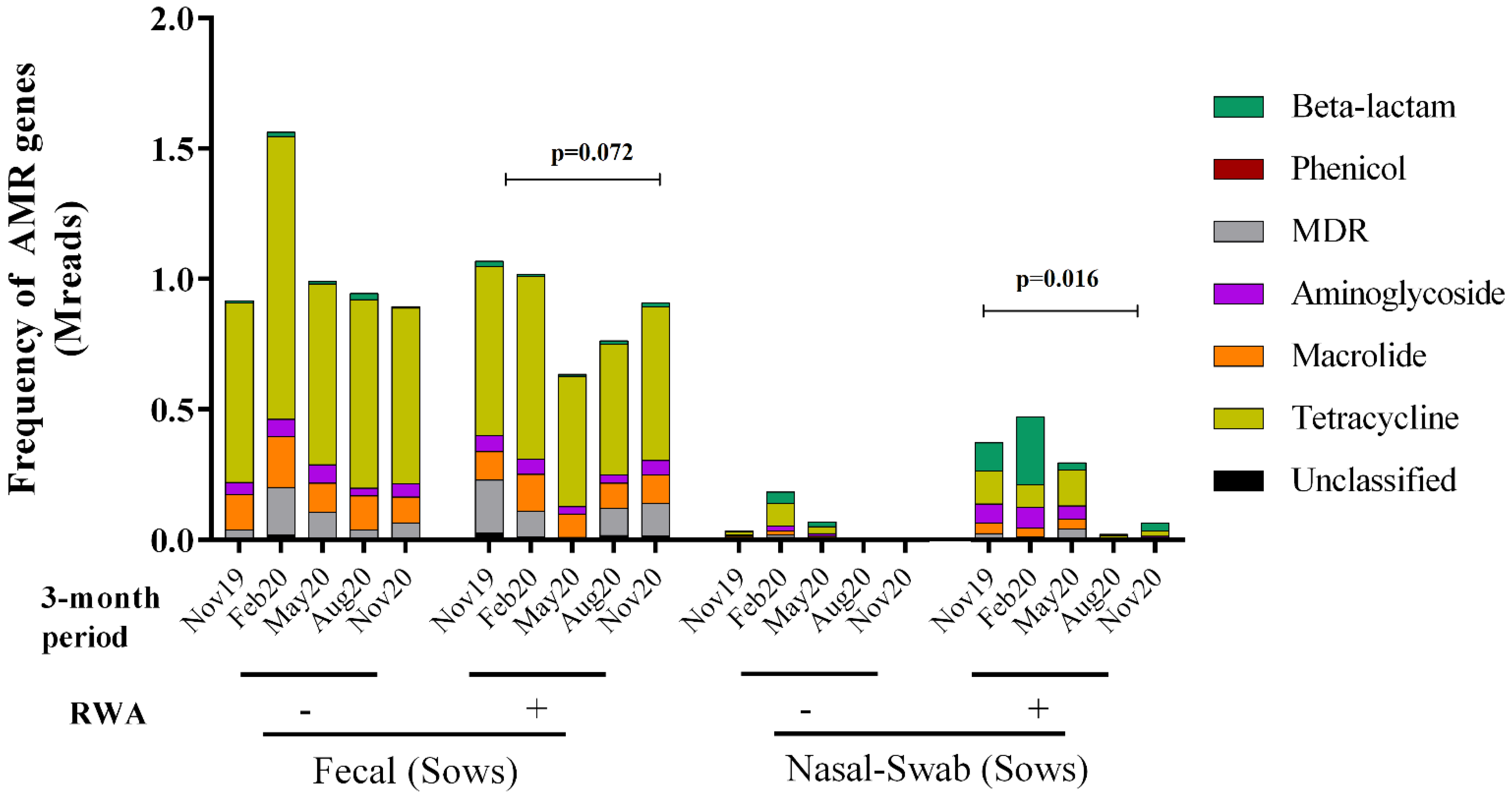
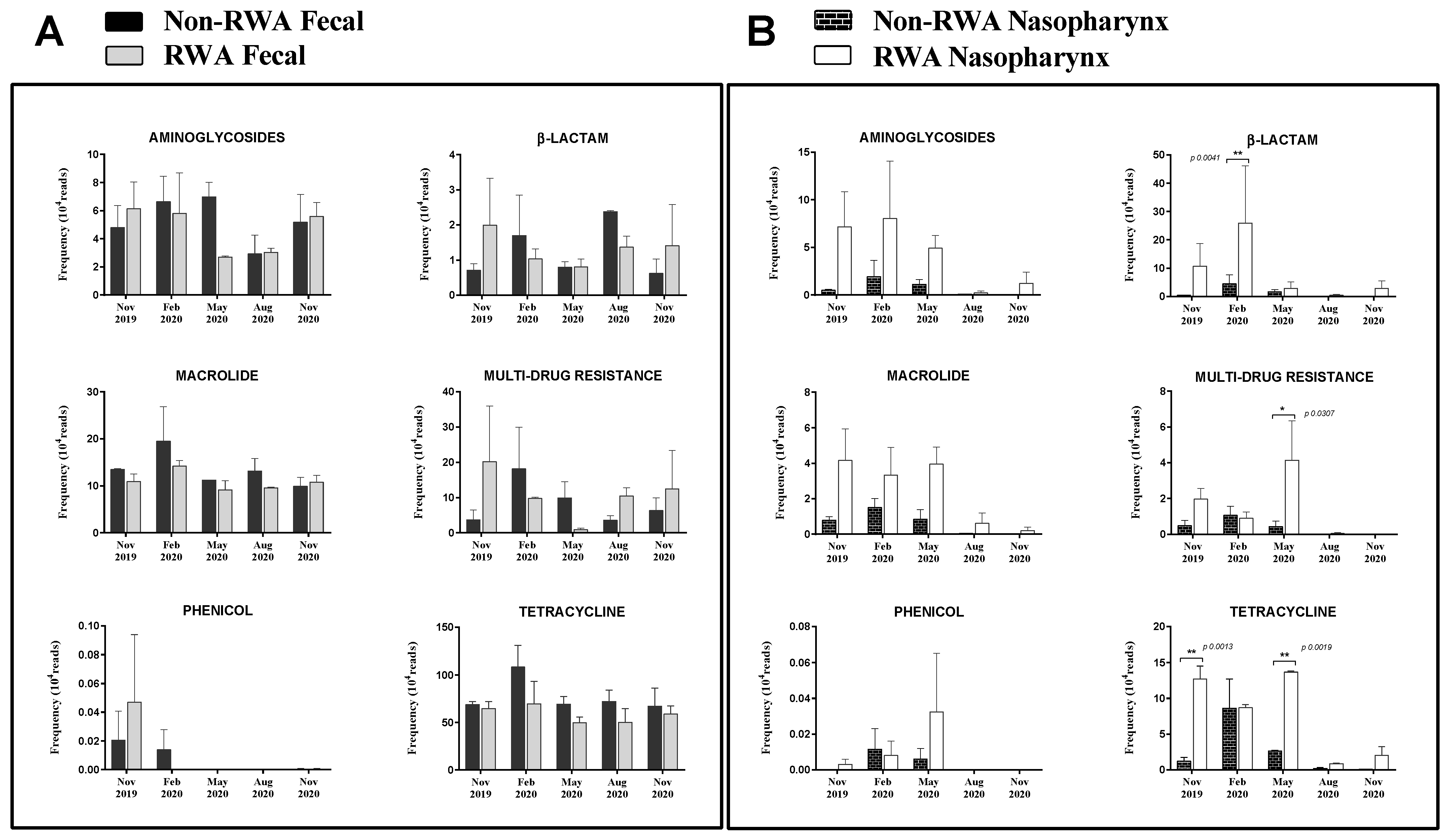
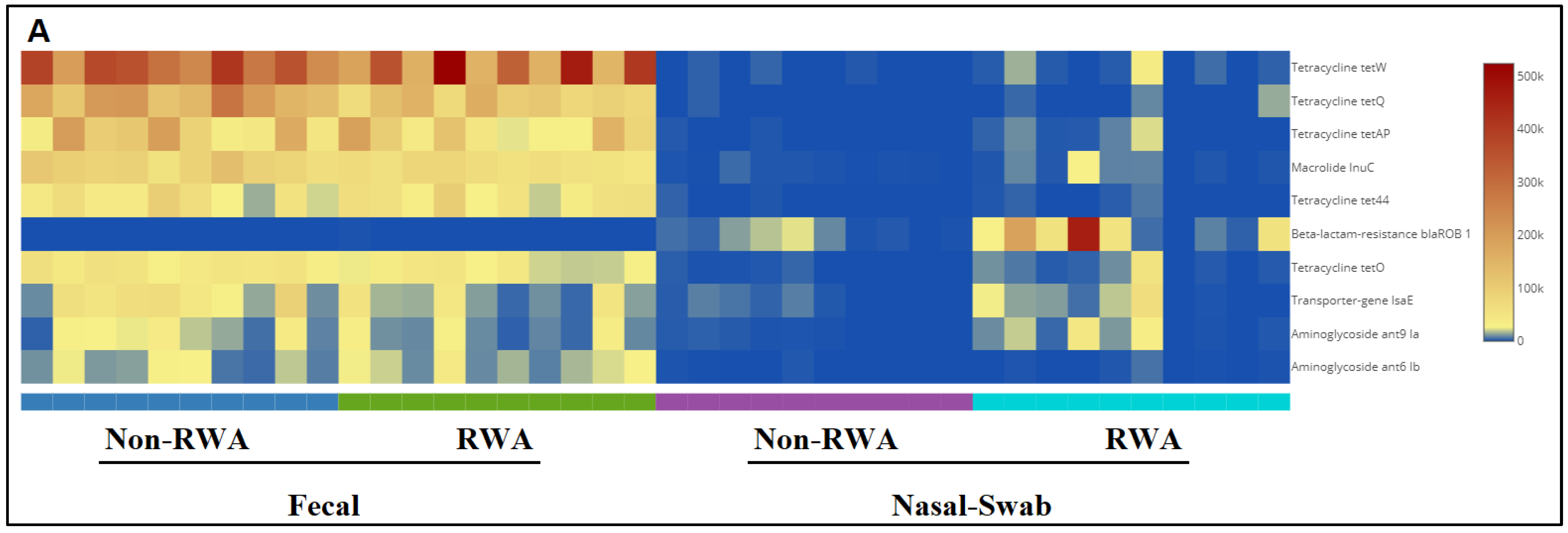
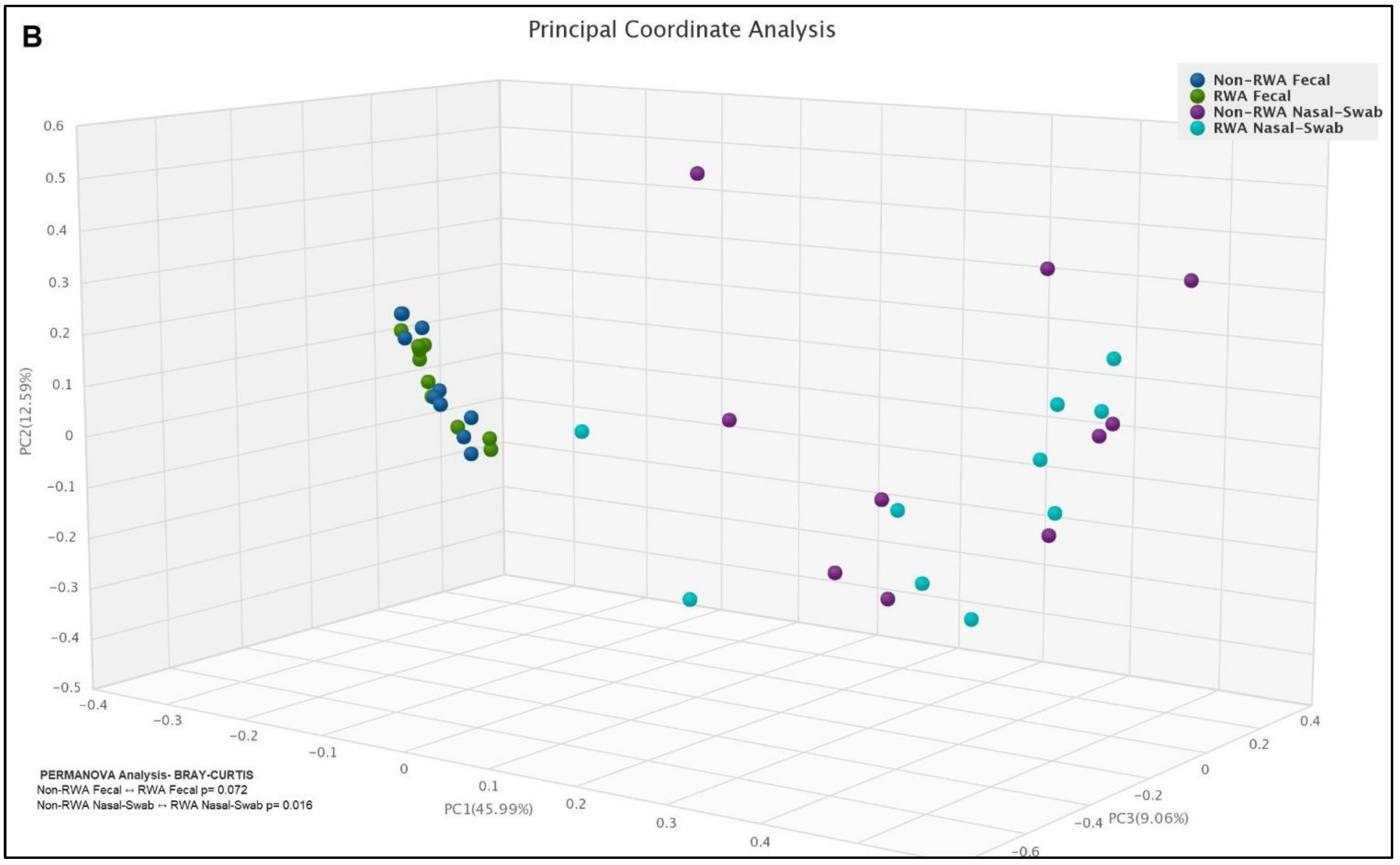
2.3. RWA Practices Increased AMR in the Sow Nasopharynx but Not in the Sow Gut
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Sample Collection
4.2. Whole Genome Sequencing (WGS) and Sequence Analyses
4.3. Prevalence of Pathogens and Resistome
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Antimicrobial Resistance Global Report on Surveillance. 2014. Available online: https://www.who.int/publications/i/item/9789241564748 (accessed on 25 May 2022).
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global trends in antimicrobial resistance in animals in low- and middle-income countries. Science 2019, 365, eaaw1944. [Google Scholar] [CrossRef]
- Public Health Agency of Canada. Canadian Antimicrobial Resistance Surveillance System Report. 2021. Available online: https://www.canada.ca/en/public-health/services/publications (accessed on 10 May 2022).
- Arnold, C.; Schupbach-Regula, G.; Hirsiger, P.; Malik, J.; Scheer, P.; Sidler, X.; Spring, P.; Perer-Egil, J.; Harisberger, M. Risk factors for oral antimicrobial consumption in Swiss fattening pigs farms—A case-control study. Porc. Health Manag. 2016, 2, 5. [Google Scholar] [CrossRef] [PubMed]
- Public Health Agency of Canada. Canadian Antimicrobial Resistance Surveillance System Report. 2016. Available online: https://www.canada.ca/en/public-health/services/publications (accessed on 13 May 2022).
- Carmo, L.P.; Schüpbach, G.; Müntener, C.; Alban, L.; Nielsen, L.R.; Magouras, I. Quantification of antimicrobial use in Swiss pigs: Comparison with other Swiss livestock species and with Danish pigs. In Proceedings of the Safe Pork Conference: Epidemiology and Control of Hazards in Pork Production Chain, Porto, Portugal, 7–10 September 2015. [Google Scholar]
- Callens, B.; Persoons, D.; Maes, D.; Laanen, M.; Postma, M.; Boyen, F. Prophylactic and metaphylactic antimicrobial use in Belgian fattening pig herds. Prev. Vet. Med. 2012, 106, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Rajić, A.; Reid-Smith, R.; Deckert, A.E.; Dewey, C.E.; McEwen, S.A. Reported antibiotic use in 90 swine farms in Alberta. Can. Vet. J. 2006, 47, 446–452. [Google Scholar]
- Dunlop, R.H.; McEwen, S.A.; Meek, A.H.; Clarke, R.C.; Black, W.D.; Friendship, R.M. Association among antimicrobial drug treatments and antimicrobial resistance of fecal Escherichia coli of swine on 34 farrrow-to-finish farms in Ontario, Canada. Prev. Vet. Med. 1998, 34, 283–305. [Google Scholar] [CrossRef]
- Statistics Canada. Hogs Statistics, Number of Hogs on Farms at End of Semi-Annual Period. 2022. Available online: https://www150.statcan.gc.ca/t1/tbl1/en/tv.action?pid=3210016001 (accessed on 3 May 2022).
- Cromwell, G.L. Why and how antibiotics are used in swine production. Anim. Biotechnol. 2002, 13, 7–27. [Google Scholar] [CrossRef] [PubMed]
- Arruda, A.G.; Deblais, L.; Hale, V.L.; Madden, C.; Pairis-Garcia, M.; Srivastava, V.; Kathayat, D.; Kumar, A.; Rajashekara, G. A cross-sectional study of the nasal and fecal microbiota of sows from different health status within six commercial swine farms. PeerJ 2021, 9, e12120. [Google Scholar] [CrossRef]
- Canadian Integrated Program for Antimicrobial Resistance Surveillance (CIPARS). Pigs. 2019. Available online: https://www.canada.ca/en/public-health/services/surveillance/canadian-integrated-program-antimicrobial-resistance-surveillance-cipars.html#wb-auto-4 (accessed on 10 May 2022).
- Chekabab, S.M.; Lawrence, J.R.; Alvarado, A.C.; Predicala, B.Z.; Korber, D.R. Piglet gut and in-barn manure from farms on a raised without antibiotics program display reduced antimicrobial resistance but an increased prevalence of pathogens. Antibiotics 2021, 10, 1152. [Google Scholar] [CrossRef]
- Jo, H.E.; Kwon, M.S.; Whon, T.W.; Kim, D.W.; Yun, M.; Lee, J.; Shin, M.Y.; Kim, S.H.; Choi, H.J. Alteration of gut microbiota after antibiotic exposure in finishing swine. Front. Microbiol. 2021, 12, 596002. [Google Scholar] [CrossRef]
- Zeineldin, M.; Aldridge, B.; Lowe, J. Antimicrobial effects on swine gastrointestinal microbiota and their accompanying antibiotic resistome. Front. Microbiol. 2019, 10, 1035. [Google Scholar] [CrossRef]
- Holman, D.B.; Chénier, M.R. Antimicrobial use in swine production and its effect on the swine gut microbiota and antimicrobial resistance. Can. J. Microbiol. 2015, 61, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Correa-Fiz, F.; Gonçalves dos Santos, J.M.; Illas, F.; Aragon, V. Antimicrobial removal on piglets promotes health and higher bacterial diversity in the nasal microbiota. Sci. Rep. 2019, 10, 1035. [Google Scholar] [CrossRef] [PubMed]
- Man, W.H.; de Steenhuijsen Piters, W.A.; Bogaert, D. The microbiota of the respiratory tract: Gatekeeper to respiratory health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef]
- Canadian Integrated Program. for Antimicrobial Resistance Surveillance (CIPARS) Annual Report. 2016. Available online: https://www.canada.ca/en/public-health/services/surveillance/canadian-integrated-program-antimicrobial-resistance-surveillance-cipars/cipars-reports/2016-annual-report-summary.html (accessed on 10 February 2022).
- Davies, P.R.; Singer, R.S. Antimicrobial use in wean to market pigs in the United States assessed via voluntary sharing of proprietary data. Zoonoses Public Health 2020, 67, 6–21. [Google Scholar] [CrossRef]
- Echtermann, T.; Muentener, C.; Sidler, X.; Kuemmerlen, D. Antimicrobial usage among different age categories and herd sizes in Swiss farrow-to-finish farms. Front. Vet. Sci. 2020, 7, 566529. [Google Scholar] [CrossRef]
- Visentin, M.; Zhao, R.; Goldman, I.D. The Antifolates. Hematol. Oncol. Clin. N. Am. 2012, 26, 629–648. [Google Scholar] [CrossRef]
- Takemura, Y.; Kobayashi, H.; Miyachi, H. Cellular and molecular mechanisms of resistance to antifolate drugs: New analogues and approaches to overcome the resistance. Int. J. Hematol. 1997, 66, 459–477. [Google Scholar] [CrossRef]
- Bergspica, I.; Kaprou, G.; Alexa, E.A.; Prieto, M.; Alvarez-Ordonez, A. Extended spectrum beta-lactamase (ESBL) producing Escherichia coli in pigs and pork meat in the European Union. Antibiotics 2020, 9, 678. [Google Scholar] [CrossRef]
- Hayer, S.S.; Rovira, A.; Olsen, K.; Johnson, T.J.; Vannucci, F.; Rendahl, A.; Perez, A.; Alvarez, J. Prevalence and trend analysis of antimicrobial resistance in clinical Escherichia coli isolates collected from diseased pigs in the USA between 2006 and 2016. Transbound. Emerg. Dis. 2020, 67, 1930–1941. [Google Scholar] [CrossRef]
- Hayer, S.S.; Rovira, A.; Olsen, K.; Johnson, T.J.; Vannucci, F.; Rendahl, A.; Perez, A.; Alvarez, J. Prevalence and time trend analysis of antimicrobial resistance in respiratory bacterial pathogens collected from diseased pigs in USA between 2006–2016. Res. Vet. Sci. 2020, 128, 135–144. [Google Scholar] [CrossRef]
- Mou, K.T.; Allen, H.K.; Alt, D.P.; Trachsel, J.; Hau, S.J.; Coetzee, J.F.; Holman, D.B.; Kellner, S.; Loving, C.L.; Brockmeier, S.L. Shifts in the nasal microbiota of swine in response to different dosing regimens of oxytetracycline administration. Vet. Microbiol. 2019, 237, 108386. [Google Scholar] [CrossRef]
- Zeineldin, M.; Aldridge, B.; Blair, B.; Kancer, K.; Lowe, J. Microbial shifts in the swine nasal microbiota in response to parenteral antimicrobial administration. Microb Pathog. 2018, 121, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Slifierz, M.J.; Friendship, R.M.; Weese, J.S. Longitudinal study of the early-life fecal and nasal microbiotas of the domestic pig. BMC Microbiol. 2015, 15, 184. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, L.; Liu, B.; Xia, J.; Liao, X.; Liu, Y. Development of aminoglycoside and β-lactamase resistance among intestinal microbiota of swine treated with lincomycin, chlortetracycline, and amoxicillin. Front. Microbiol. 2014, 5, 580. [Google Scholar] [CrossRef]
- De, R.; Mukhopadhyay, A.K.; Dutta, S. Metagenomic analysis of gut microbiome and resistome of diarrheal fecal samples from Kolkata, India, reveals the core and variable microbiota including signatures of microbial dark matter. Gut Pathog. 2020, 7, 12–32. [Google Scholar] [CrossRef]
- Holman, D.B.; Yang, W.; Alexander, T.W. Antibiotic treatment in feedlot cattle: A longitudinal study of the effect of oxytetracycline and tulathromycin on the fecal and nasopharyngeal microbiota. Microbiome 2019, 7, 86. [Google Scholar] [CrossRef] [PubMed]
- Agga, G.E.; Morgan Scott, H.; Vinasco, J.; Nagaraja, T.G.; Amachawadi, R.G.; Bai, J.; Norby, B.; Renter, D.G.; Dritz, S.S.; Nelssen, J.L.; et al. Effects of chlortetracycline and copper supplementation on the prevalence, distribution, and quantity of antimicrobial resistance genes in the fecal metagenome of weaned pigs. Prev. Vet. Med. 2015, 119, 179–189. [Google Scholar] [CrossRef]
- Looft, T.; Johnson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D.; Sul, W.J.; Stedtfeld, T.M.; Chai, B.; Cole, J.R.; et al. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef]
- Pakpour, S.; Jabaji, S.; Chenier, M.R. Frequency of antibiotic resistance in a swine facility 2.5 years after a ban on antibiotics. Microb. Ecol. 2012, 63, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Stanton, T.B.; Humphrey, S.B.; Stoffregen, W.C. Chlortetracycline-resistant intestinal bacteria in organically raised and feral swine. Appl. Environ. Microbiol. 2011, 77, 7167–7170. [Google Scholar] [CrossRef]
- Mathew, A.G.; Beckmann, M.A.; Saxton, A.M. A comparison of antibiotic resistance in bacteria isolated from swine herds in which antibiotics were used or excluded. J. Swine Health Prod. 2001, 9, 125–129. [Google Scholar]
- Frye, J.G.; Lindsey, R.L.; Meinersmann, R.J.; Berrang, M.E.; Jackson, C.R.; Englen, M.D.; Turpin, J.B.; Fedorka-Cray, P.J. Related antimicrobial resistance genes detected in different bacterial species co-isolated from swine fecal samples. Foodborne Pathog. Dis. 2011, 8, 663–679. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Biosafety in Microbiological and Biomedical Laboratories (BMBL), 6th ed.; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018. Available online: https://www.cdc.gov/labs/BMBL.html (accessed on 24 October 2020).
- Chekabab, S.M.; Lawrence, J.R.; Alvarado, A.C.; Predicala, B.Z.; Korber, D.R. A health metadata-based management approach for comparative analysis of high-throughput genetic sequences for quantifying antimicrobial resistance reduction in Canadian hog barns. Comput. Struct. Biotechnol. J. 2020, 18, 2629–2638. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Numbers of Animals | Type of Antibiotics | Antibiotics Used | |||
|---|---|---|---|---|---|
| Absolute Quantity | Relative Amount | ||||
| mg | Percent | DDDvetCA 1 | Percent | ||
| Non-RWA | |||||
| Sow (n = 78) | Antifolate | 425,496 | 77% | 925 | 72% |
| Β-lactam | 8000 | 1% | 13 | 1% | |
| Tetracycline | 119,500 | 22% | 347 | 27% | |
| Total antibiotics (mg) | 552,996 | 1285 | |||
| RWA | |||||
| Sow (n = 22) | β-lactam | 107,085 | 82% | 173 | 85% |
| Tetracycline | 23,250 | 18% | 31 | 15% | |
| Total antibiotics (mg) | 130,335 | 204 | |||
| Barns | Type of Antibiotics | Drug Name | Dosage | Treatment Route 1 |
|---|---|---|---|---|
| Non-RWA | ||||
| Antifolate | Trimidox (trimethoprim & sulfadoxine) | 1 mL/15 kg/day | IV or IM injection | |
| Tetracycline | Biomycin (Oxytetracycline) | 1 mL/10 kg/day | IM or subcutaneous | |
| β-lactam | Penicillin G | 6000 IU per kg (1 mL/50 kg) | IM injection | |
| Polyflex (ampicillin) | 6 mg/kg/day | IM injection | ||
| Excenel (ceftiofur) | 3.0 mg/kg/day for 3 days | IM injection | ||
| RWA | ||||
| β-lactam | Polyflex (ampicillin) | 6 mg/kg/day | IM injection | |
| Penicillin G | 6000 IU per kg (1 mL/50 kg) | IM injection | ||
| Tetracycline | Biomycin (Oxytetracycline) | 1 mL/10 kg/day | IM or subcutaneous |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarado, A.C.; Chekabab, S.M.; Predicala, B.Z.; Korber, D.R. Impact of Raised without Antibiotics Measures on Antimicrobial Resistance and Prevalence of Pathogens in Sow Barns. Antibiotics 2022, 11, 1221. https://doi.org/10.3390/antibiotics11091221
Alvarado AC, Chekabab SM, Predicala BZ, Korber DR. Impact of Raised without Antibiotics Measures on Antimicrobial Resistance and Prevalence of Pathogens in Sow Barns. Antibiotics. 2022; 11(9):1221. https://doi.org/10.3390/antibiotics11091221
Chicago/Turabian StyleAlvarado, Alvin C., Samuel M. Chekabab, Bernardo Z. Predicala, and Darren R. Korber. 2022. "Impact of Raised without Antibiotics Measures on Antimicrobial Resistance and Prevalence of Pathogens in Sow Barns" Antibiotics 11, no. 9: 1221. https://doi.org/10.3390/antibiotics11091221
APA StyleAlvarado, A. C., Chekabab, S. M., Predicala, B. Z., & Korber, D. R. (2022). Impact of Raised without Antibiotics Measures on Antimicrobial Resistance and Prevalence of Pathogens in Sow Barns. Antibiotics, 11(9), 1221. https://doi.org/10.3390/antibiotics11091221