
The Detection of Potential Native Probiotics Lactobacillus spp. against Salmonella Enteritidis, Salmonella Infantis and Salmonella Kentucky ST198 of Lebanese Chicken Origin

 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Screening of Lactobacilli and Their Anti-Salmonella Activity
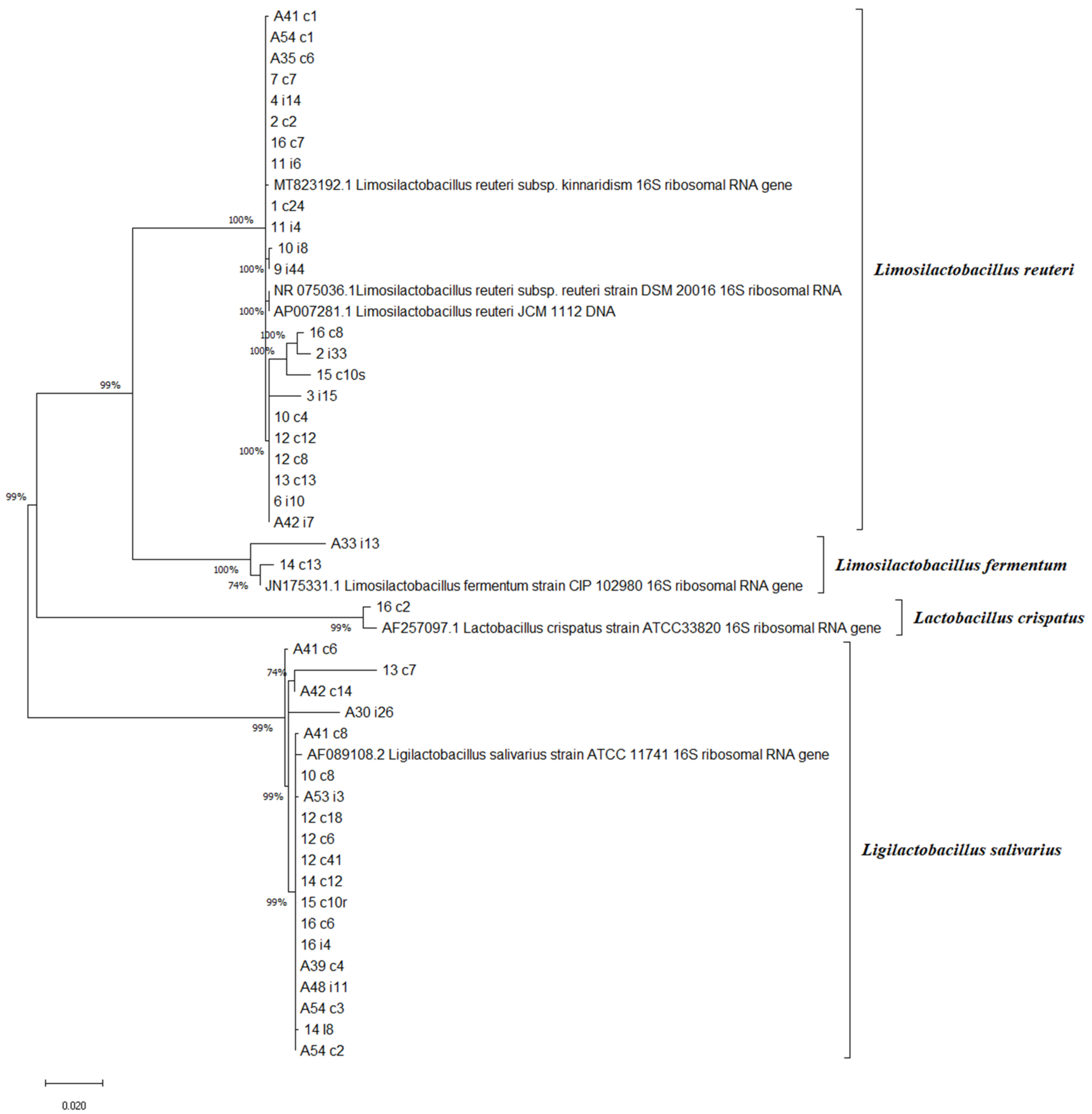
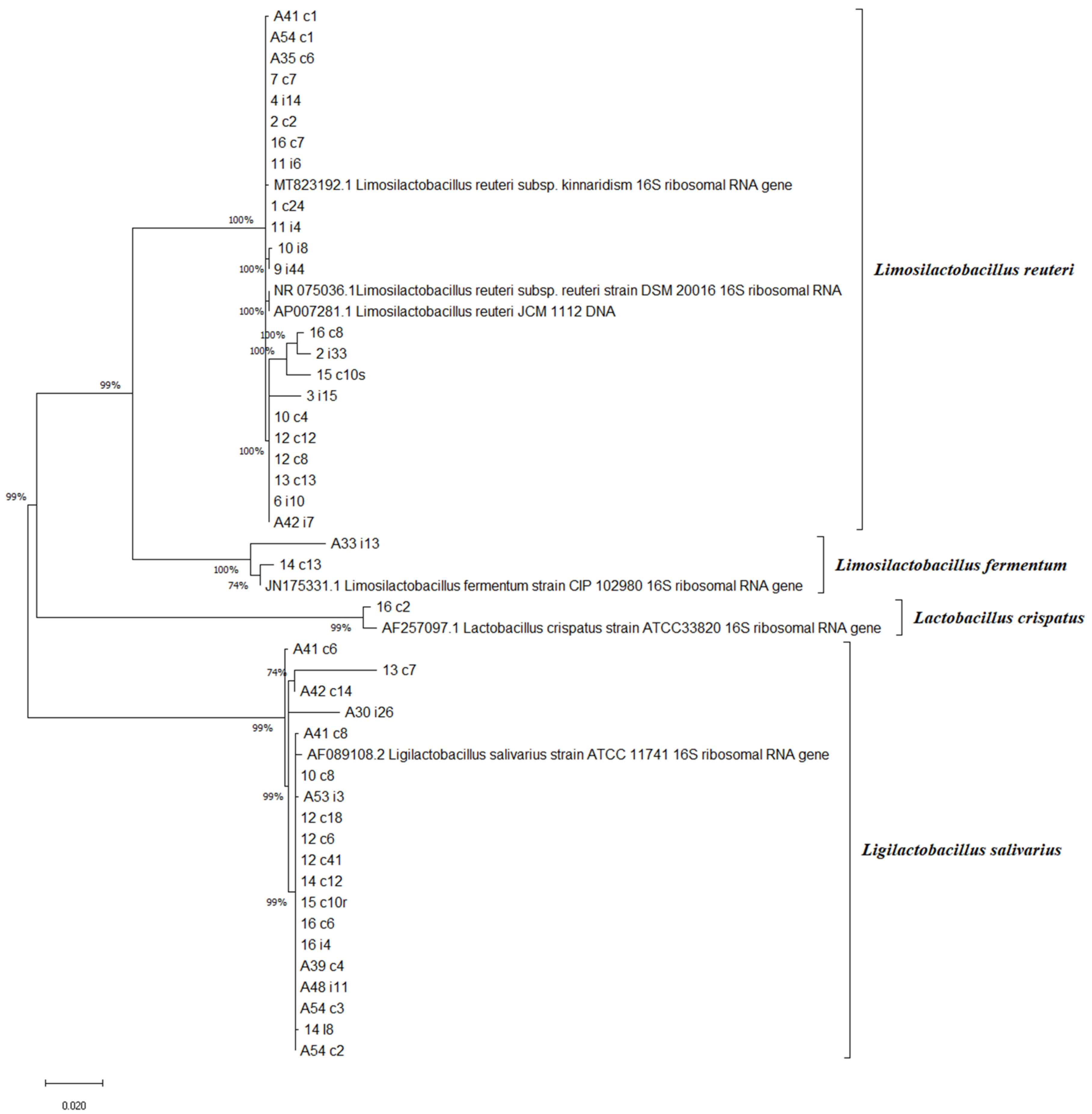
2.2. Genotypic Identification of Lactobacilli Isolates with Phylogenetic Relations
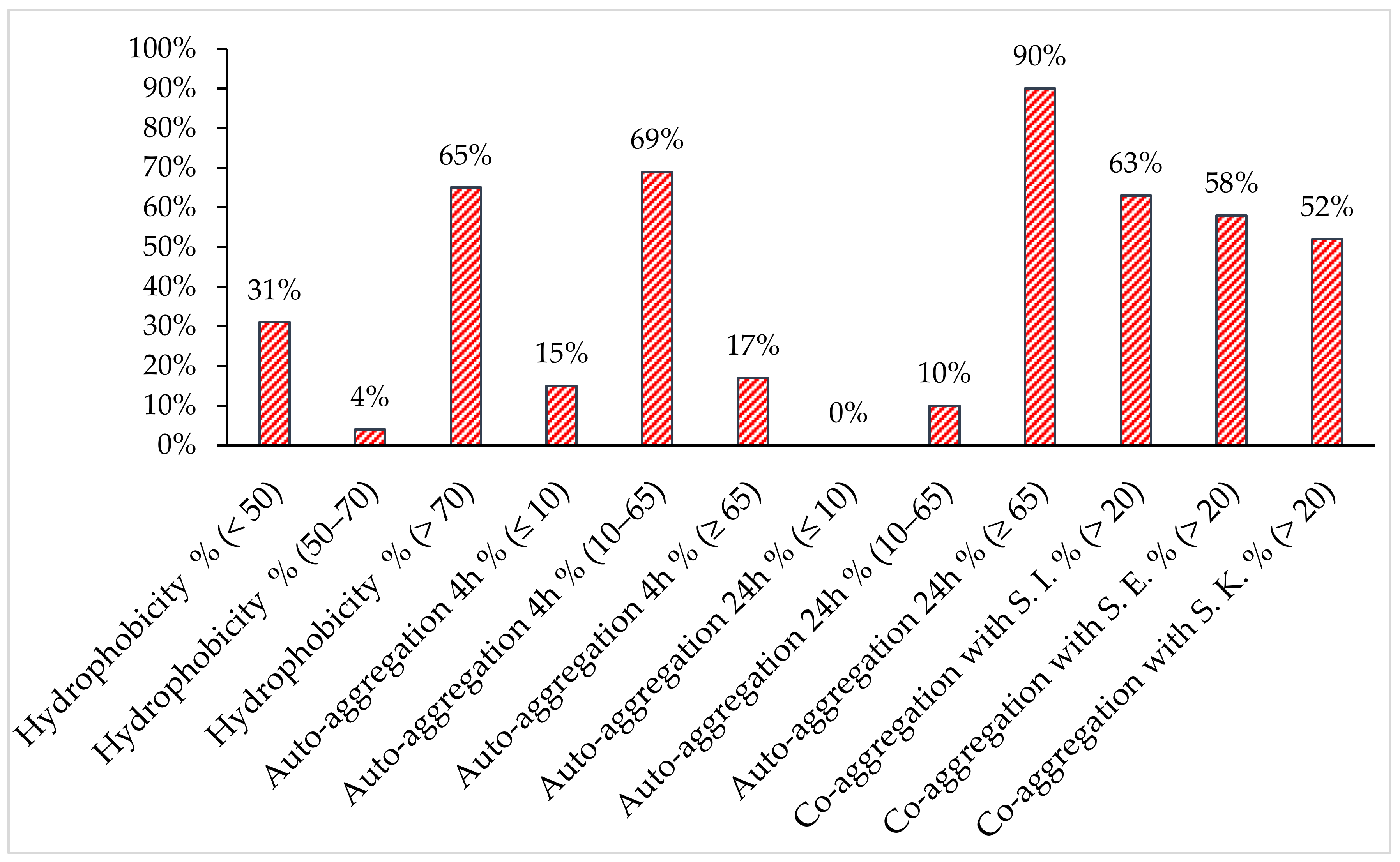
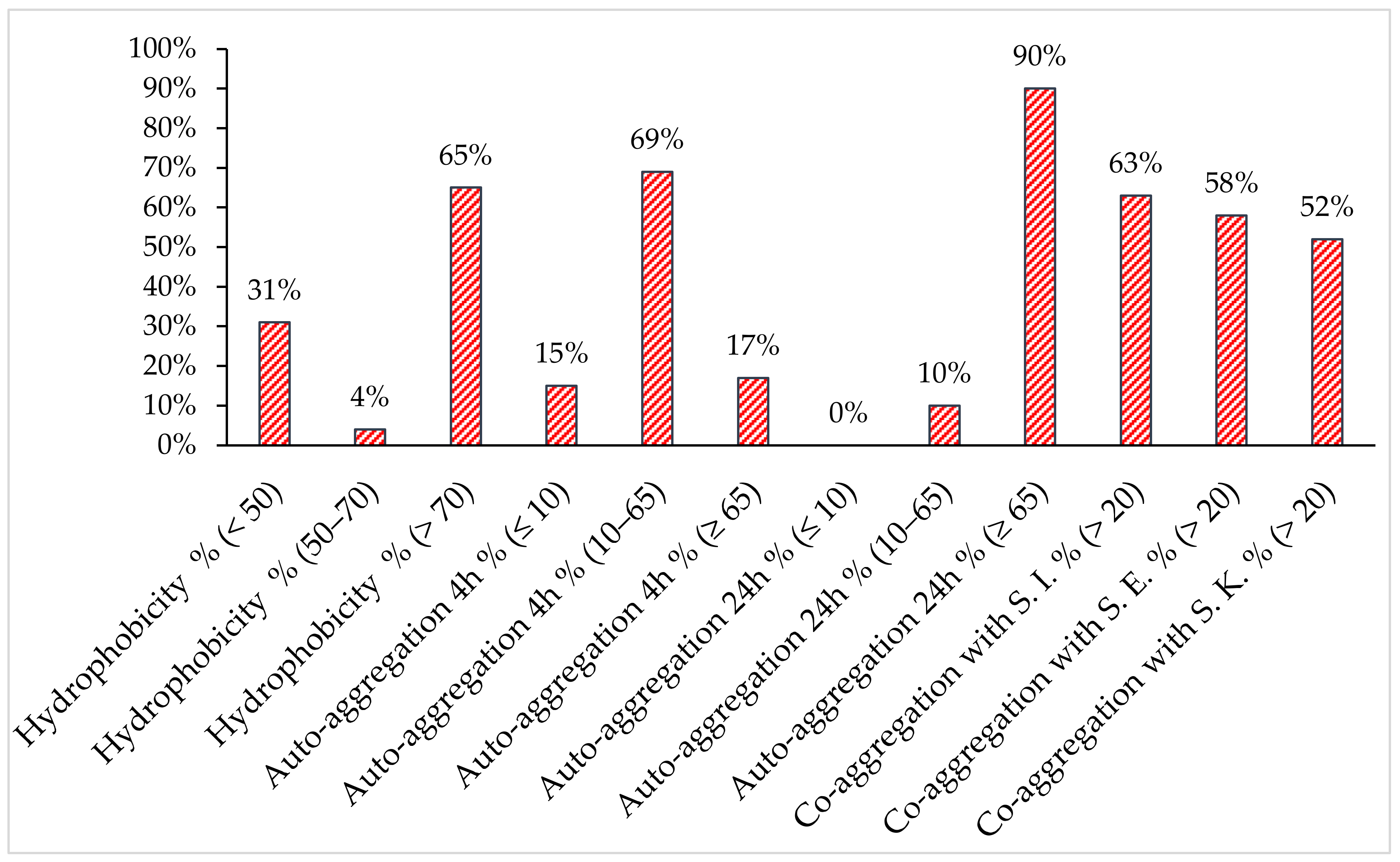
2.3. Analysis of Surface Properties
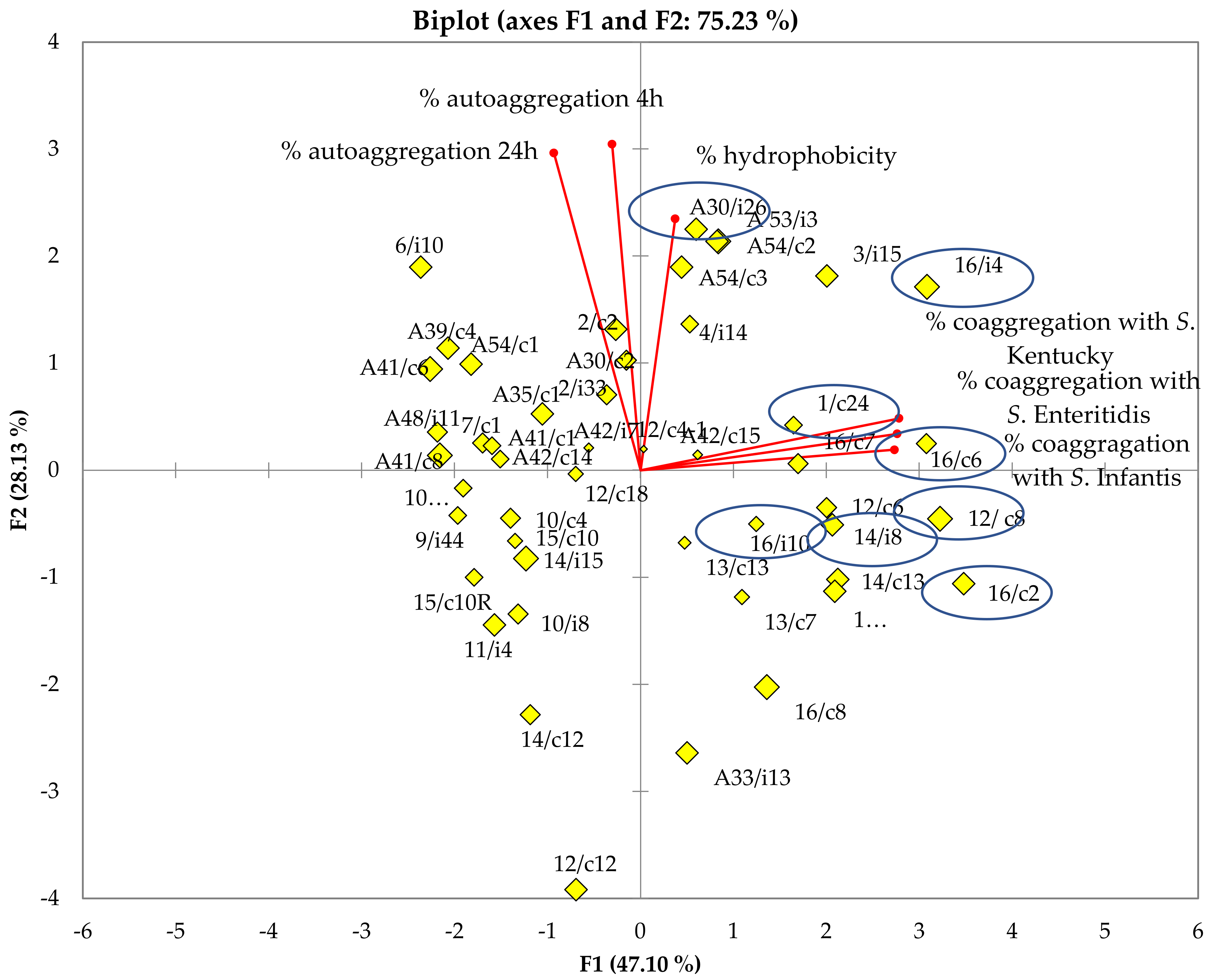
2.4. Hydrophobicity and Auto/co-Aggregation Correlation
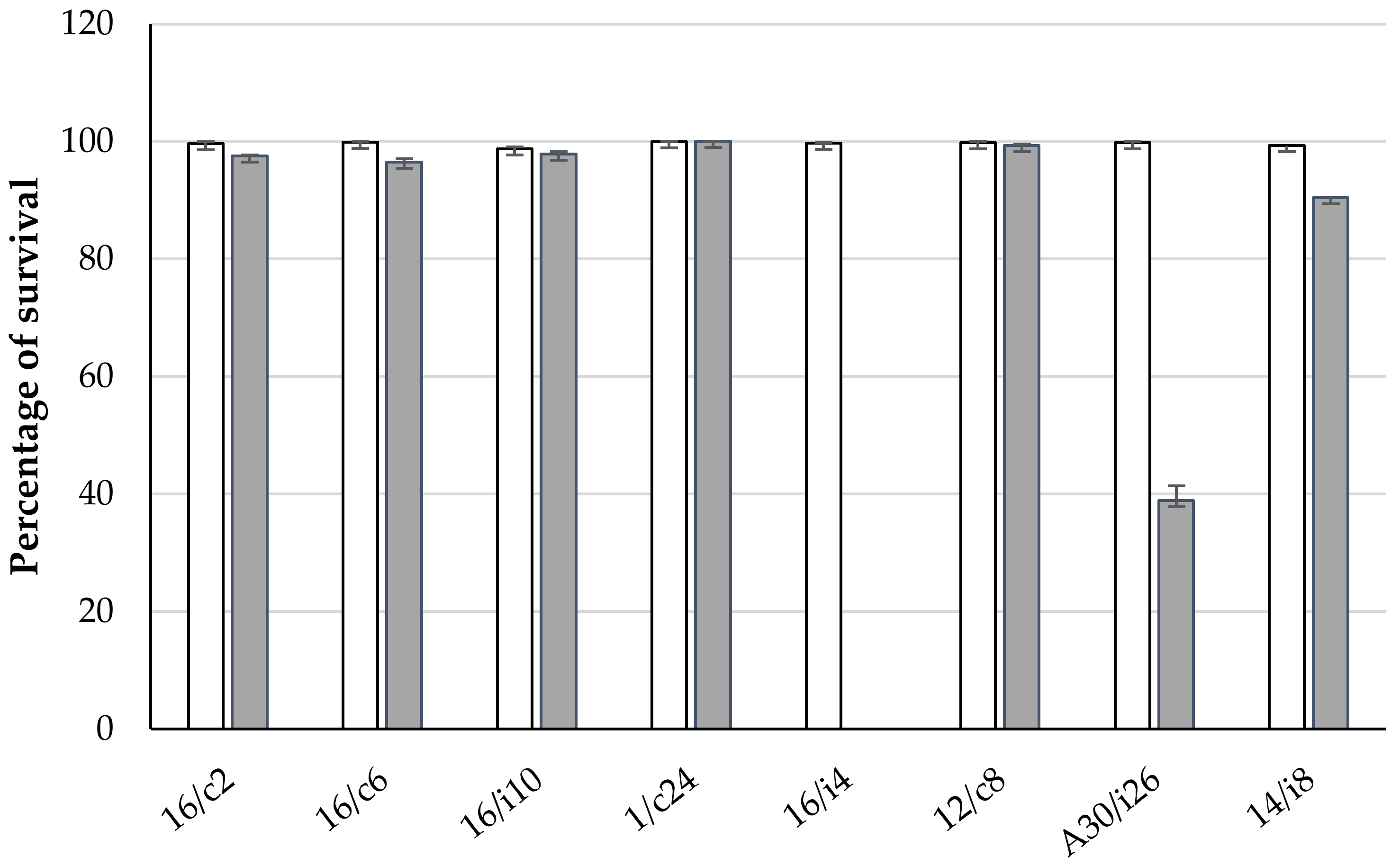
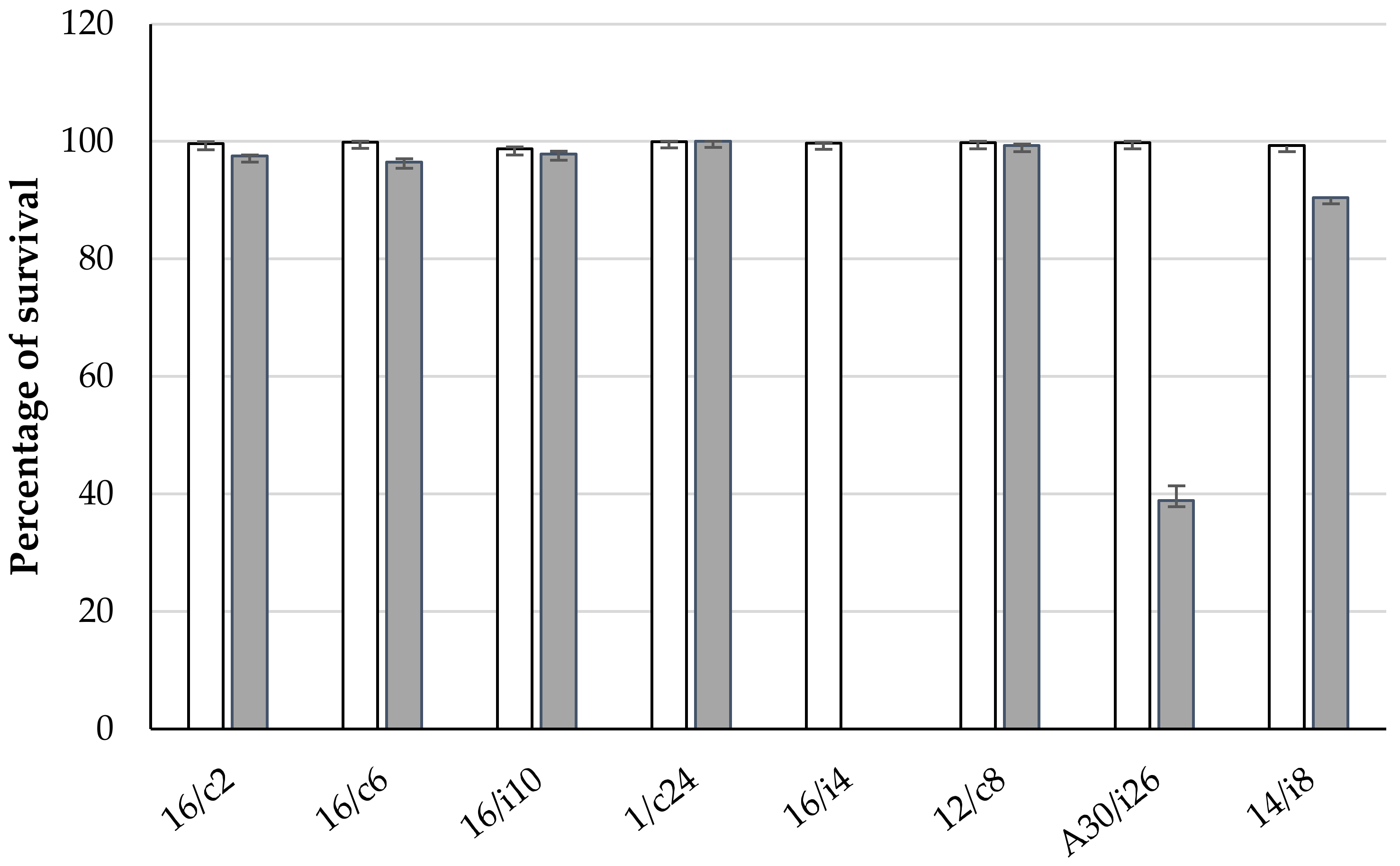
2.5. Assays for Tolerance to Simulated Gastrointestinal Conditions of Chickens
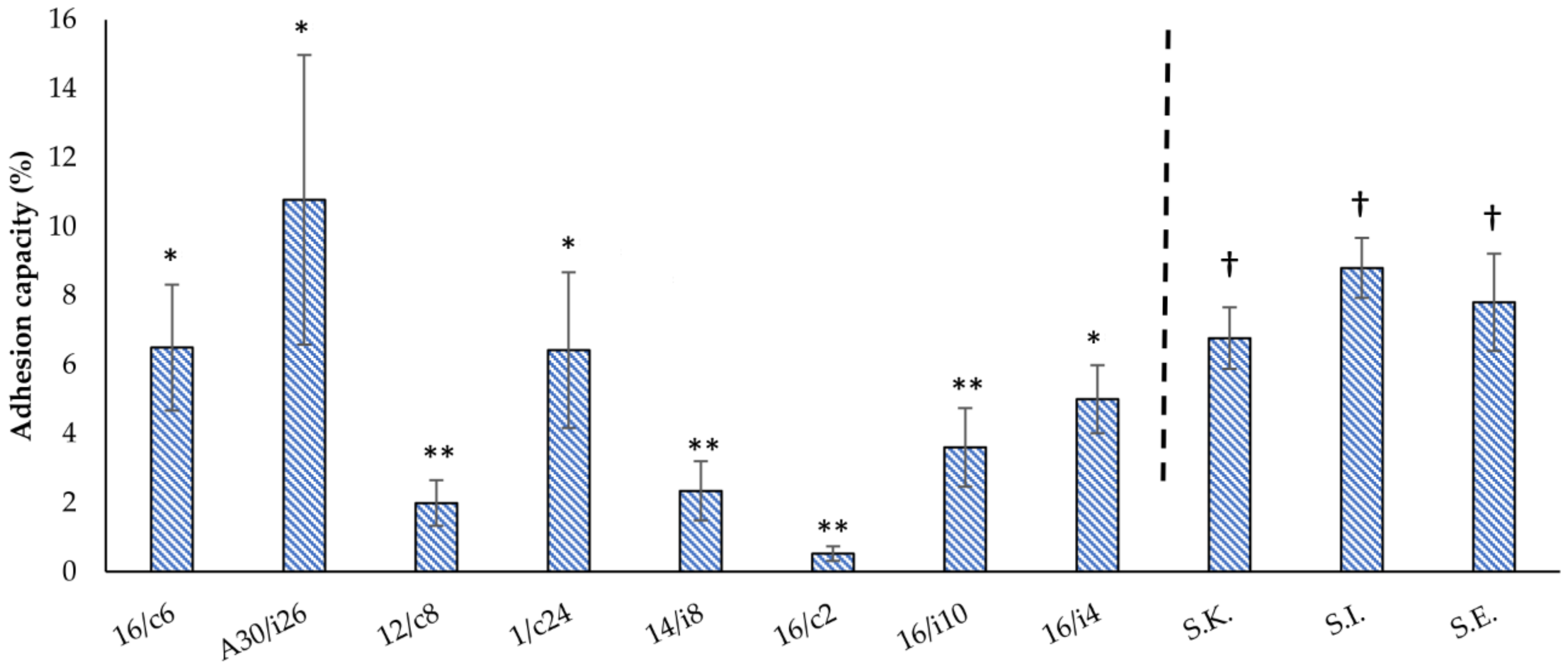
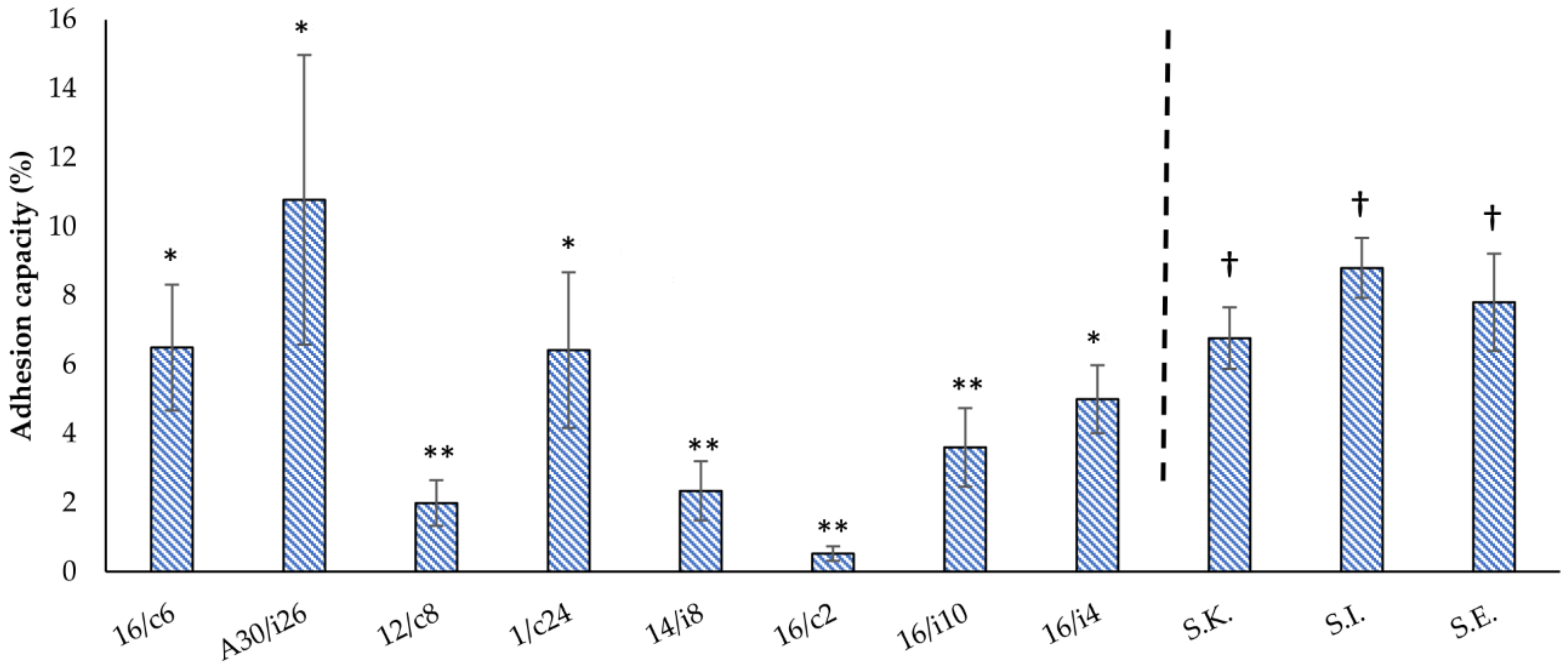
2.6. Adhesion Assays
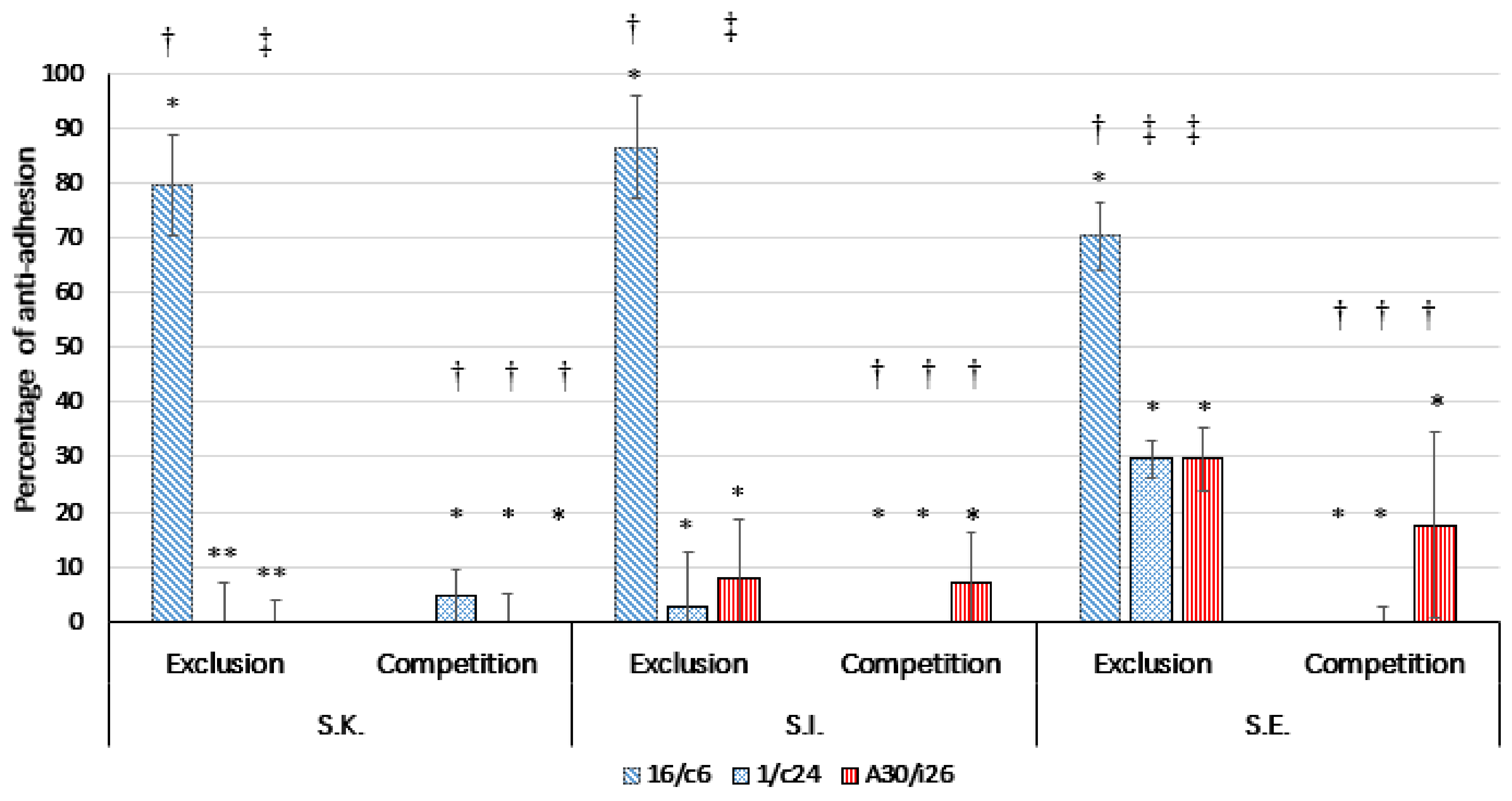
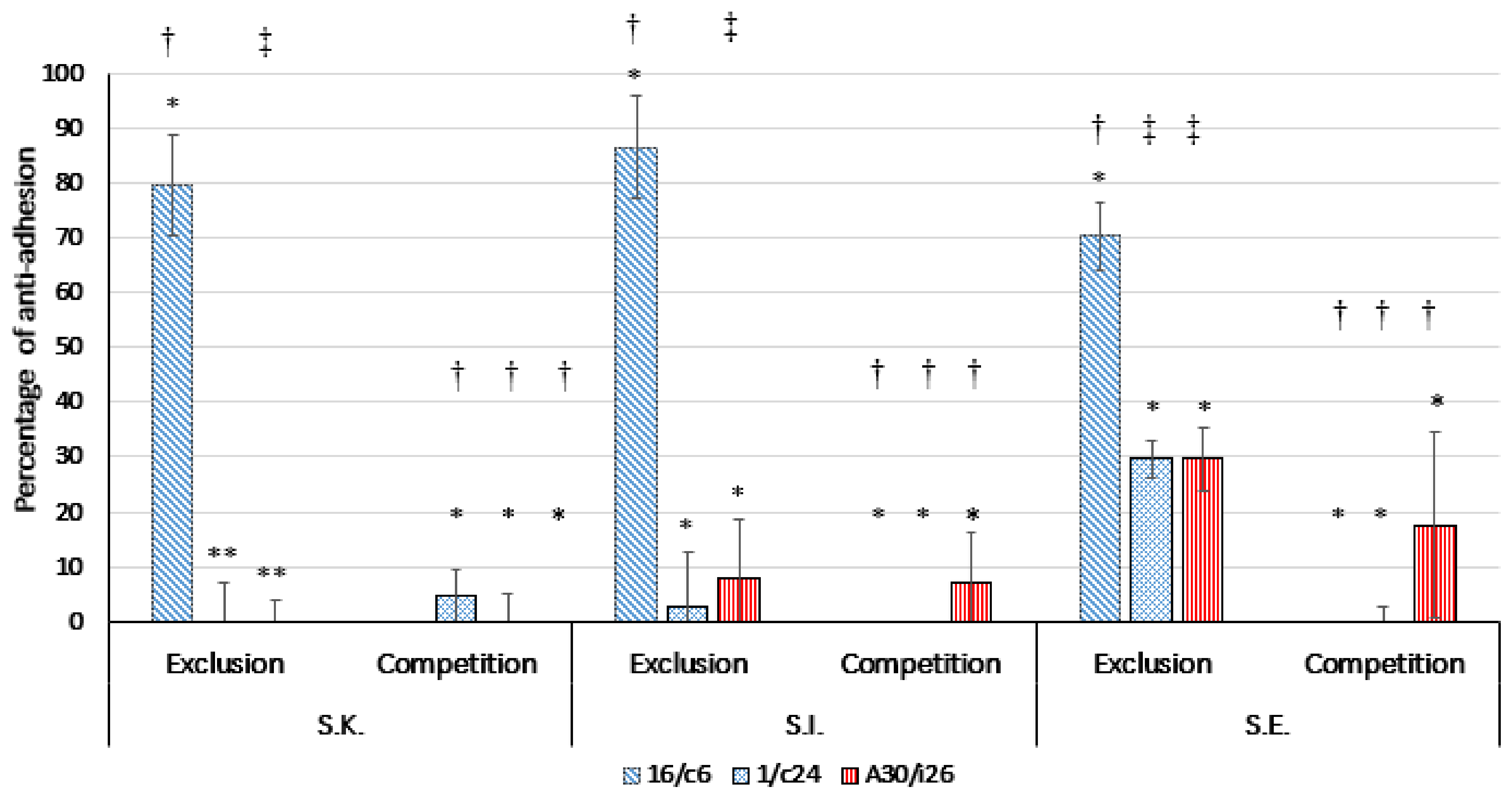
2.7. Competition/Exclusion Assays
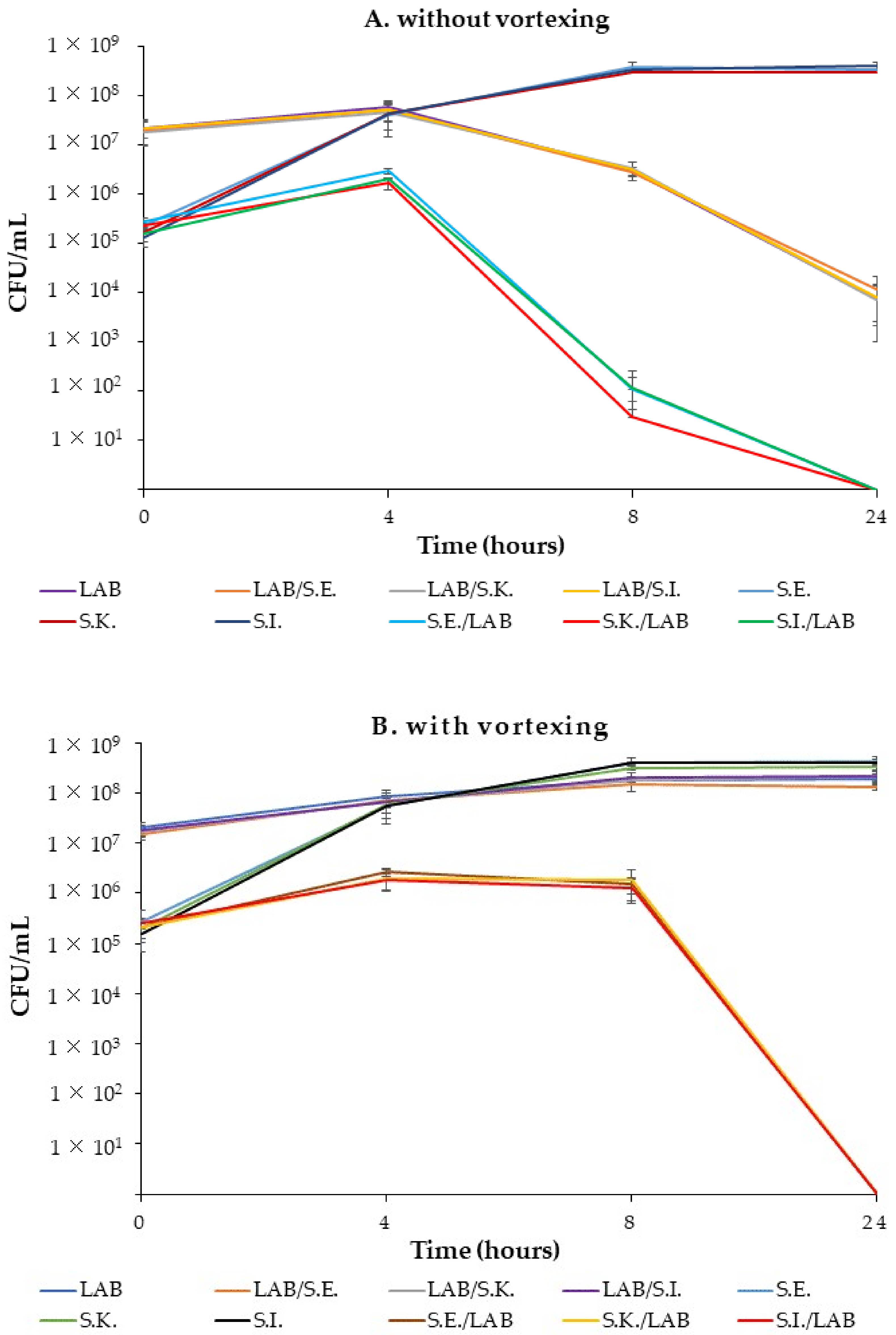
2.8. Co-Culture Growth Kinetics
3. Discussion
4. Materials and Methods
4.1. Isolation and Phenotypic Characterization of Lactobacillus spp.
4.2. Salmonella Isolates
4.3. Assessment of the Lactobacilli Antagonism
4.4. Selection of Strains Based on Their Phenotypic Aggregation
4.5. Species Identification and Phylogenetic Relationships
4.6. Cell Surface Properties
4.6.1. Auto-Aggregation and Co-Aggregation Assays
4.6.2. Hydrophobicity Assays
4.7. In Vitro Cell Tolerance to Gastrointestinal Conditions
4.8. Cell Culture
4.8.1. Cell Line and Growth Conditions
4.8.2. Adhesion to Caco-2 Cells
4.8.3. Inhibition of the Adhesion of Salmonella to Caco-2 Cells
4.9. Co-Culture Growth Kinetic Study
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority/European Centre for Disease Prevention and Control. The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2016. EFSA J. 2017, 15, e04676. [Google Scholar] [CrossRef]
- Tan, S.M.; Lee, S.M.; Dykes, G.A. Buffering Effect of Chicken Skin and Meat Protects Salmonella Enterica Strains against Hydrochloric Acid but Not Organic Acid Treatment. Food Control 2014, 42, 329–334. [Google Scholar] [CrossRef]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The Role of Poultry Meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [PubMed]
- El Hage, R.; El Rayess, Y.; Bonifait, L.; El Hafi, B.; Baugé, L.; Viscogliosi, E.; Hamze, M.; Mathieu, F.; Matar, G.M.; Chemaly, M. A National Study through a ‘Farm-to-Fork’ Approach to Determine Salmonella Dissemination along with the Lebanese Poultry Production Chain. Zoonoses Public Health 2022, 69, 499–513. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Yu, Z. Intestinal Microbiome of Poultry and Its Interaction with Host and Diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Ferri, M.; Ranucci, E.; Romagnoli, P.; Giaccone, V. Antimicrobial Resistance: A Global Emerging Threat to Public Health Systems. Crit. Rev. Food Sci. Nutr. 2017, 53, 2857–2876. [Google Scholar] [CrossRef]
- Franco, A.; Leekitcharoenphon, P.; Feltrin, F.; Alba, P.; Cordaro, G.; Iurescia, M.; Tolli, R.; D’Incau, M.; Staffolani, M.; Di Giannatale, E.; et al. Emergence of a Clonal Lineage of Multidrug-Resistant ESBL-Producing Salmonella Infantis Transmitted from Broilers and Broiler Meat to Humans in Italy between 2011 and 2014. PLoS ONE 2015, 10, e144802. [Google Scholar] [CrossRef]
- Regulation (EC) No 1831/2003. Regulation (EC) No 1831/2003 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition. Off. J. Eur. Communities 2003, L268, 15.
- Krysiak, K.; Konkol, D.; Korczyński, M. Review Overview of the Use of Probiotics in Poultry Production. Animals 2021, 11, 1620. [Google Scholar] [CrossRef]
- Foley, S.L.; Nayak, R.; Hanning, I.B.; Johnson, T.J.; Han, J.; Ricke, S.C. Population Dynamics of Salmonella Enterica Serotypes in Commercial Egg and Poultry Production. Appl. Environ. Microbiol. 2011, 77, 4273–4279. [Google Scholar] [CrossRef]
- FAO; WHO. Guidelines for the Evaluation of Probiotics in Food; FAO: Rome, Italy; WHO: Geneva, Switzerland, 2002. [Google Scholar]
- Spivey, M.A.; Dunn-Horrocks, S.L.; Duong, T. Epithelial Cell Adhesion and Gastrointestinal Colonization of Lactobacillus in Poultry. Poult. Sci. 2014, 93, 2910–2919. [Google Scholar] [CrossRef] [PubMed]
- Saint-Cyr, M.J.; Guyard-Nicodème, M.; Messaoudi, S.; Chemaly, M.; Cappelier, J.M.; Dousset, X.; Haddad, N. Recent Advances in Screening of Anti-Campylobacter Activity in Probiotics for Use in Poultry. Front. Microbiol. 2016, 7, 553. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Forssten, S.; Hibberd, A.A.; Lyra, A.; Stahl, B. Probiotic Approach to Prevent Antibiotic Resistance. Ann. Med. 2016, 48, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Quezada, S.; Gomez-Llorente, C.; Plaza-Diaz, J.; Chenoll, E.; Ramón, D.; Matencio, E.; Bermudez-Brito, M.; Genovés, S.; Romero, F.; Gil, A.; et al. Competitive Inhibition of Three Novel Bacteria Isolated from Faeces of Breast Milk-Fed Infants against Selected Enteropathogens. Br. J. Nutr. 2013, 109 (Suppl. 2), S63–S69. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wang, L.; Zhou, L.; Yang, X.; Zhao, X. Using in Vitro Immunomodulatory Properties of Lactic Acid Bacteria for Selection of Probiotics against Salmonella Infection in Broiler Chicks. PLoS ONE 2016, 11, e0147630. [Google Scholar] [CrossRef] [PubMed]
- Rantala, M.; Nurmi, E. Prevention of the Growth of Salmonella Infantis in Chicks by the Flora of the Alimentary Tract of Chickens. Br. Poult. Sci. 1973, 14, 627–630. [Google Scholar] [CrossRef]
- Ayeni, A.O.; Ruppitsch, W.; Ayeni, F.A. Characterization of Bacteria in Nigerian Yogurt as Promising Alternative to Antibiotics in Gastrointestinal Infections. J. Diet. Suppl. 2018, 211, 1–11. [Google Scholar] [CrossRef]
- Yadav, A.K.; Tyagi, A.; Kumar, A.; Panwar, S.; Grover, S.; Saklani, A.C.; Hemalatha, R.; Batish, V.K. Adhesion of Lactobacilli and Their Anti-Infectivity Potential. Crit. Rev. Food Sci. Nutr. 2017, 57, 2042–2056. [Google Scholar] [CrossRef]
- Vineetha, P.G.; Tomar, S.; Saxena, V.K.; Susan, C.; Sandeep, S.; Adil, K.; Mukesh, K. Screening of Lactobacillus Isolates from Gastrointestinal Tract of Guinea Fowl for Probiotic Qualities Using in vitro Tests to Select Species-Specific Probiotic Candidates. Br. Poult. Sci. 2016, 57, 474–482. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap; Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Chen, X.; Xu, J.; Shuai, J.; Chen, J.; Zhang, Z.; Fang, W. The S-Layer Proteins of Lactobacillus Crispatus Strain ZJ001 Is Responsible for Competitive Exclusion against Escherichia coli O157:H7 and Salmonella Typhimurium. Int. J. Food Microbiol. 2007, 115, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Albazaz, R.I.; Byukunal Bal, E.B. Microflora of Digestive Tract in Poultry. Kahramanmaraş Sütçü İmam Üniversitesi Doğa Bilim. Derg. 2014, 17, 39–42. [Google Scholar] [CrossRef]
- Babot, J.D.; Argañaraz-Martínez, E.; Saavedra, L.; Apella, M.C.; Perez Chaia, A. Selection of Indigenous Lactic Acid Bacteria to Reinforce the Intestinal Microbiota of Newly Hatched Chicken—Relevance of in Vitro and Ex Vivo Methods for Strains Characterization. Res. Vet. Sci. 2014, 97, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Schillinger, U.; Lucke, F.K. Antimicrobial Activity of Lactobacillus Sake Isolated from Meat. Appl. Environ. Microbiol. 1989, 55, 1901–1906. [Google Scholar] [CrossRef]
- Adetoye, A.; Pinloche, E.; Adeniyi, B.A.; Ayeni, F.A. Characterization and Anti-Salmonella Activities of Lactic Acid Bacteria Isolated from Cattle Faeces. BMC Microbiol. 2018, 18, 96. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.L.; Thorsen, L.; Schwan, R.F.; Jespersen, L. Strain-Specific Probiotics Properties of Lactobacillus fermentum, Lactobacillus plantarum and Lactobacillus crevis Isolates from Brazilian Food Products. Food Microbiol. 2013, 36, 22–29. [Google Scholar] [CrossRef]
- Gupta, A.; Tiwari, S.K. Plantaricin LD1: A Bacteriocin Produced by Food Isolate of Lactobacillus plantarum LD1. Appl. Biochem. Biotechnol. 2014, 172, 3354–3362. [Google Scholar] [CrossRef]
- García-Cayuela, T.; Korany, A.M.; Bustos, I.; Gómez de Cadiñanos, L.P.; Requena, T.; Peláez, C.; Martínez-Cuesta, M.C. Adhesion Abilities of Dairy Lactobacillus plantarum Strains Showing an Aggregation Phenotype. Food Res. Int. 2014, 57, 44–50. [Google Scholar] [CrossRef]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and Aggregation Ability of Probiotic Strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef]
- Potočnjak, M.; Pušić, P.; Frece, J.; Abram, M.; Janković, T.; Gobin, I. Three New Lactobacillus plantarum Strains in the Probiotic Toolbox against Gut Pathogen Salmonella enterica Serotype Typhimurium. Food Technol. Biotechnol. 2017, 55, 48–54. [Google Scholar] [CrossRef]
- Nishiyama, K.; Sugiyama, M.; Mukai, T. Adhesion Properties of Lactic Acid Bacteria on Intestinal Mucin. Microorganisms 2016, 4, 34. [Google Scholar] [CrossRef]
- Polak-Berecka, M.; Waśko, A.; Paduch, R.; Skrzypek, T.; Sroka-Bartnicka, A. The Effect of Cell Surface Components on Adhesion Ability of Lactobacillus rhamnosus. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2014, 106, 751–762. [Google Scholar] [CrossRef]
- Collado, M.C.; Surono, I.; Meriluoto, J.; Salminen, S. Indigenous Dadih Lactic Acid Bacteria: Cell-Surface Properties and Interactions with Pathogens. J. Food Sci. 2007, 72, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Ait Seddik, H.; Bendali, F.; Cudennec, B.; Drider, D. Anti-Pathogenic and Probiotic Attributes of Lactobacillus salivarius and Lactobacillus plantarum Strains Isolated from Feces of Algerian Infants and Adults. Res. Microbiol. 2017, 168, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Solieri, L.; Bianchi, A.; Mottolese, G.; Lemmetti, F.; Giudici, P. Tailoring the Probiotic Potential of Non-Starter Lactobacillus Strains from Ripened Parmigiano Reggiano Cheese by in Vitro Screening and Principal Component Analysis. Food Microbiol. 2014, 38, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Singh, T.P.; Kaur, G.; Kapila, S.; Malik, R.K. Antagonistic Activity of Lactobacillus reuteri Strains on the Adhesion Characteristics of Selected Pathogens. Front. Microbiol. 2017, 8, 486. [Google Scholar] [CrossRef]
- Campana, R.; Van Hemert, S.; Baffone, W. Strain-Specific Probiotic Properties of Lactic Acid Bacteria and Their Interference with Human Intestinal Pathogens Invasion. Gut Pathog. 2017, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Handley, P.S.; Harty, D.W.; Wyatt, J.E.; Brown, C.R.; Doran, J.P.; Gibbs, A.C. A Comparison of the Adhesion, Coaggregation and Cell-Surface Hydrophobicity Properties of Fibrillar and Fimbriate Strains of Streptococcus salivarius. J. Gen. Microbiol. 1987, 133, 3207–3217. [Google Scholar] [CrossRef]
- De Souza, B.M.S.; Borgonovi, T.F.; Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Lactobacillus casei and Lactobacillus fermentum Strains Isolated from Mozzarella Cheese: Probiotic Potential, Safety, Acidifying Kinetic Parameters and Viability under Gastrointestinal Tract Conditions. Probiotics Antimicrob. Proteins 2018, 11, 382–396. [Google Scholar] [CrossRef]
- Del Re, B.; Sgorbati, B.; Miglioli, M.; Palenzona, D. Adhesion, Autoaggregation and Hydrophobicity of 13 Strains of Bifidobacterium longum. Lett. Appl. Microbiol. 2000, 31, 438–442. [Google Scholar] [CrossRef]
- Zommiti, M.; Connil, N.; Hamida, J.B.; Ferchichi, M. Probiotic Characteristics of Lactobacillus curvatus DN317, a Strain Isolated from Chicken Ceca. Probiotics Antimicrob. Proteins 2017, 9, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Abhisingha, M.; Dumnil, J.; Pitaksutheepong, C. Selection of Potential Probiotic Lactobacillus with Inhibitory Activity Against Salmonella and Fecal Coliform Bacteria. Probiotics Antimicrob. Proteins 2018, 10, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, A.; Laubitz, D.; Antushevich, H.; Zabielski, R.; Grzesiuk, E. Competition of Lactobacillus paracasei with Salmonella Enterica for Adhesion to Caco-2 Cells. J. Biomed. Biotechnol. 2008, 2008, 357964. [Google Scholar] [CrossRef] [PubMed]
- Jessie Lau, L.Y.; Chye, F.Y. Antagonistic Effects of Lactobacillus plantarum 0612 on the Adhesion of Selected Foodborne Enteropathogens in Various Colonic Environments. Food Control 2018, 91, 237–247. [Google Scholar] [CrossRef]
- Hai, D.; Lu, Z.; Huang, X.; Lv, F.; Bie, X. In Vitro Screening of Chicken-Derived Lactobacillus Strains That Effectively Inhibit Salmonella Colonization and Adhesion. Foods 2021, 10, 569. [Google Scholar] [CrossRef] [PubMed]
- Malik, A.; Sakamoto, M.; Hanazaki, S.; Osawa, M.; Suzuki, T.; Tochigi, M.; Kakii, K. Coaggregation among Nonflocculating Bacteria Isolated from Activated Sludge. Appl. Environ. Microbiol. 2003, 69, 6056–6063. [Google Scholar] [CrossRef]
- Bian, X.; Evivie, S.E.; Muhammad, Z.; Luo, G.W.; Liang, H.Z.; Wang, N.N.; Huo, G.C. In Vitro Assessment of the Antimicrobial Potentials of Lactobacillus helveticus Strains Isolated from Traditional Cheese in Sinkiang China against Food-Borne Pathogens. Food Funct. 2016, 7, 789–797. [Google Scholar] [CrossRef]
- El Hage, R.; Losasso, C.; Longo, A.; Petrin, S.; Ricci, A.; Mathieu, F.; Abi Khattar, Z.; El Rayess, Y. Whole-Genome Characterisation of TEM-1 and CMY-2 β-Lactamase-Producing Salmonella Kentucky ST198 in Lebanese Broiler Chain. J. Glob. Antimicrob. Resist. 2020, 23, 408–416. [Google Scholar] [CrossRef]
- Saitou, N.N.M.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees’. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Rosenberg, M.; Gutnick, D.; Rosenberg, E. Adherence of Bacteria to Hydrocarbons: A Simple Method for Measuring Cell-Surface Hydrophobicity. FEMS Microbiol. Lett. 1980, 9, 29–33. [Google Scholar] [CrossRef]
- Buahom, J.; Siripornadulsil, S.; Siripornadulsil, W. Feeding with Single Strains Versus Mixed Cultures of Lactic Acid Bacteria and Bacillus subtilis KKU213 Affects the Bacterial Community and Growth Performance of Broiler Chickens. Arab. J. Sci. Eng. 2018, 43, 3417–3427. [Google Scholar] [CrossRef]
- Son, S.H.; Jeon, H.L.; Yang, S.J.; Lee, N.K.; Paik, H.D. In Vitro Characterization of Lactobacillus brevis KU15006, an Isolate from Kimchi, Reveals Anti-Adhesion Activity against Foodborne Pathogens and Antidiabetic Properties. Microb. Pathog. 2017, 112, 135–141. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Visual Aggregation | Auto-Aggregation 4 h (%) | Auto-Aggregation 24 h (%) | % Co-Aggregation with | Hydrophobicity (%) | ||
|---|---|---|---|---|---|---|---|
| S. Enteritidis | S. Infantis | S. Kentucky ST198 | |||||
| L. crispatus 16/c2 | Agg+/Agg− | 14.46 ± 2.78 | 58.67 ± 7.62 | 89.36 | 75.06 | 69.66 | 84.58 ± 1.92 |
| Ligilactobacillussalivarius 16/c6 | Agg− | 9.89 ± 3.63 | 95.91 ± 2.58 | 71.07 | 69.55 | 94.55 | 90.26 ± 3.91 |
| Ligilactobacillussalivarius 16/i4 | Agg+ | 76.23 ± 3.38 | 92.95 ± 10.5 | 82.49 | 80.45 | 79.94 | 82.25 ± 5.84 |
| Lactobacillus sp. 16/i10 | Agg+/Agg− | 6.16 ± 5.53 | 79.46 ± 1.18 | 45.60 | 34.32 | 63.51 | 98.36 ± 0.75 |
| Ligilactobacillussalivarius 14/i8 | Agg+/Agg− | 23.14 ± 5.29 | 73.47 ± 3.67 | 62.30 | 70.35 | 47.54 | 81.63 ± 1.2 |
| Limosilactobacillus reuteri 12/c8 | Agg+/Agg− | 33.93 ± 6.44 | 71.86 ± 1.89 | 83.47 | 73.87 | 80.00 | 52.66 ± 2.98 |
| Limosilactobacillus reuteri 1/c24 | Agg+/Agg− | 13.76 ± 1.87 | 91.81 ± 7.78 | 50.43 | 62.47 | 58.93 | 97.53 ± 0.96 |
| Ligilactobacillussalivarius A30/i26 | Agg+ | 76.15 ± 3.93 | 99.63 ± 0.26 | 49.54 | 25.71 | 60.00 | 98.84 ± 1.34 |
| Variables | Hydrophobicity (%) | Auto-Aggregation 4 h (%) | Auto-Aggregation 24 h (%) | Co-Aggregation with S. Infantis (%) | Co-Aggregation with S. Enteritidis (%) | Co-Aggregation with S. Kentucky (%) |
|---|---|---|---|---|---|---|
| Hydrophobicity (%) | 1 | |||||
| Auto-aggregation 4 h (%) | 0.2264 | 1 | ||||
| Auto-aggregation 24 h (%) | 0.2665 | 0.5302 | 1 | |||
| Co-aggregation with S. Infantis (%) | 0.0595 | −0.0537 | −0.1878 | 1 | ||
| Co-aggregation with S. Enteritidis (%) | 0.1524 | 0.0202 | −0.1880 | 0.8782 | 1 | |
| Co-aggregation with S. Kentucky (%) | 0.1496 | −0.0208 | −0.2181 | 0.8439 | 0.8887 | 1 |
| Experiment Number | Category, Age | Breed | Diet Formula |
|---|---|---|---|
| 1 | Broiler, 35 days | Cobb | High starch diet: corn 60%, soya 20%, wheat 20% |
| 2 | Broiler, 35 days | Cobb | High protein diet: soya 40%, corn 40%, wheat 20% |
| 3 | Broiler, 35 days | Cobb | High gluten diet: wheat 60%, soya 20%, corn 20% |
| 4 | Broiler, 35 days | Ross | High starch diet: corn 60%, soya 20%, wheat 20% |
| 5 | Broiler, 35 days | Ross | High protein diet: soya 40%, corn 40%, wheat 20% |
| 6 | Broiler, 35 days | Ross | High gluten diet: wheat 60%, soya 20%, corn 20% |
| 7 | Broiler, one day | Cobb | High starch diet: corn 60%, soya 20%, wheat 20% |
| 8 | Broiler, one day | Cobb | High protein diet: soya 40%, corn 40%, wheat 20% |
| 9 | Broiler, one day | Cobb | High gluten diet: wheat 60%, soya 20%, corn 20% |
| 10 | Broiler, one day | Ross | High starch diet: corn 60%, soya 20%, wheat 20% |
| 11 | Broiler, one day | Ross | High protein diet: soya 40%, corn 40%, wheat 20% |
| 12 | Broiler, one day | Ross | High gluten diet: wheat 60%, soya 20%, corn 20% |
| 13 | Layer, 69 weeks | Isa Brown | Normal feed: corn 40%, soya 32%, wheat 20% |
| 14 | Layer, 69 weeks | Isa White | Normal feed: corn 40%, soya 32%, wheat 20% |
| 15 | Layer, 27 weeks | Isa Brown | Normal feed: corn 40%, soya 32%, wheat 20% |
| 16 | Layer, 27 weeks | Isa White | Normal feed: corn 40%, soya 32%, wheat 20% |
| A | Broiler, 35 weeks | Ross | Normal feed: corn 40%, soya 32%, wheat 20% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Hage, R.; El Hage, J.; Snini, S.P.; Ammoun, I.; Touma, J.; Rachid, R.; Mathieu, F.; Sabatier, J.-M.; Abi Khattar, Z.; El Rayess, Y. The Detection of Potential Native Probiotics Lactobacillus spp. against Salmonella Enteritidis, Salmonella Infantis and Salmonella Kentucky ST198 of Lebanese Chicken Origin. Antibiotics 2022, 11, 1147. https://doi.org/10.3390/antibiotics11091147
El Hage R, El Hage J, Snini SP, Ammoun I, Touma J, Rachid R, Mathieu F, Sabatier J-M, Abi Khattar Z, El Rayess Y. The Detection of Potential Native Probiotics Lactobacillus spp. against Salmonella Enteritidis, Salmonella Infantis and Salmonella Kentucky ST198 of Lebanese Chicken Origin. Antibiotics. 2022; 11(9):1147. https://doi.org/10.3390/antibiotics11091147
Chicago/Turabian StyleEl Hage, Rima, Jeanne El Hage, Selma P. Snini, Imad Ammoun, Joseph Touma, Rami Rachid, Florence Mathieu, Jean-Marc Sabatier, Ziad Abi Khattar, and Youssef El Rayess. 2022. "The Detection of Potential Native Probiotics Lactobacillus spp. against Salmonella Enteritidis, Salmonella Infantis and Salmonella Kentucky ST198 of Lebanese Chicken Origin" Antibiotics 11, no. 9: 1147. https://doi.org/10.3390/antibiotics11091147
APA StyleEl Hage, R., El Hage, J., Snini, S. P., Ammoun, I., Touma, J., Rachid, R., Mathieu, F., Sabatier, J.-M., Abi Khattar, Z., & El Rayess, Y. (2022). The Detection of Potential Native Probiotics Lactobacillus spp. against Salmonella Enteritidis, Salmonella Infantis and Salmonella Kentucky ST198 of Lebanese Chicken Origin. Antibiotics, 11(9), 1147. https://doi.org/10.3390/antibiotics11091147