


Expression Profiles of Zebrafish (Danio rerio) Lysozymes and Preparation of c-Type Lysozyme with High Bacteriolytic Activity against Vibrio vulnificus


Abstract
:1. Introduction
2. Results
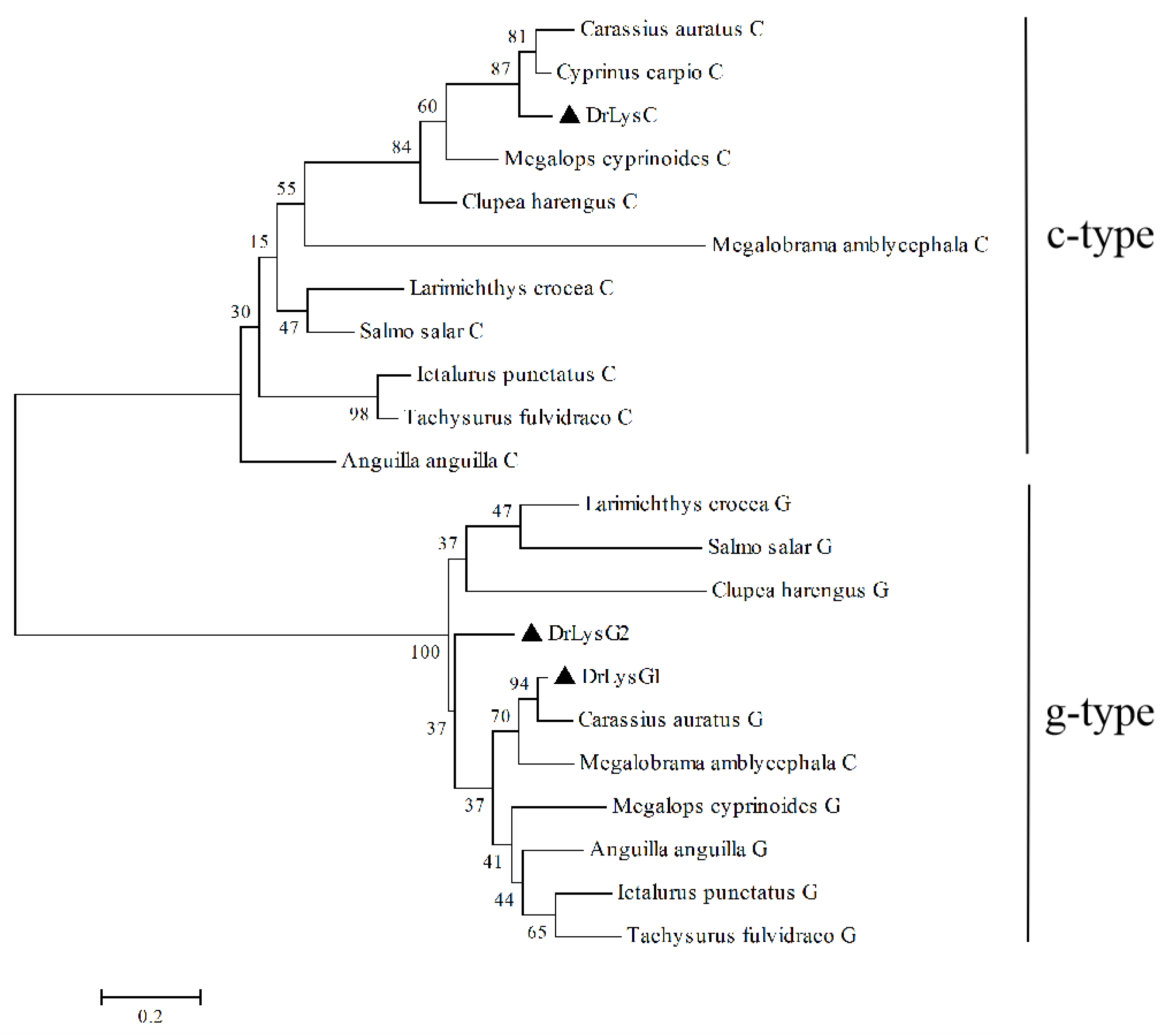
2.1. Composition and Constitutive Expression of Zebrafish Lysozymes
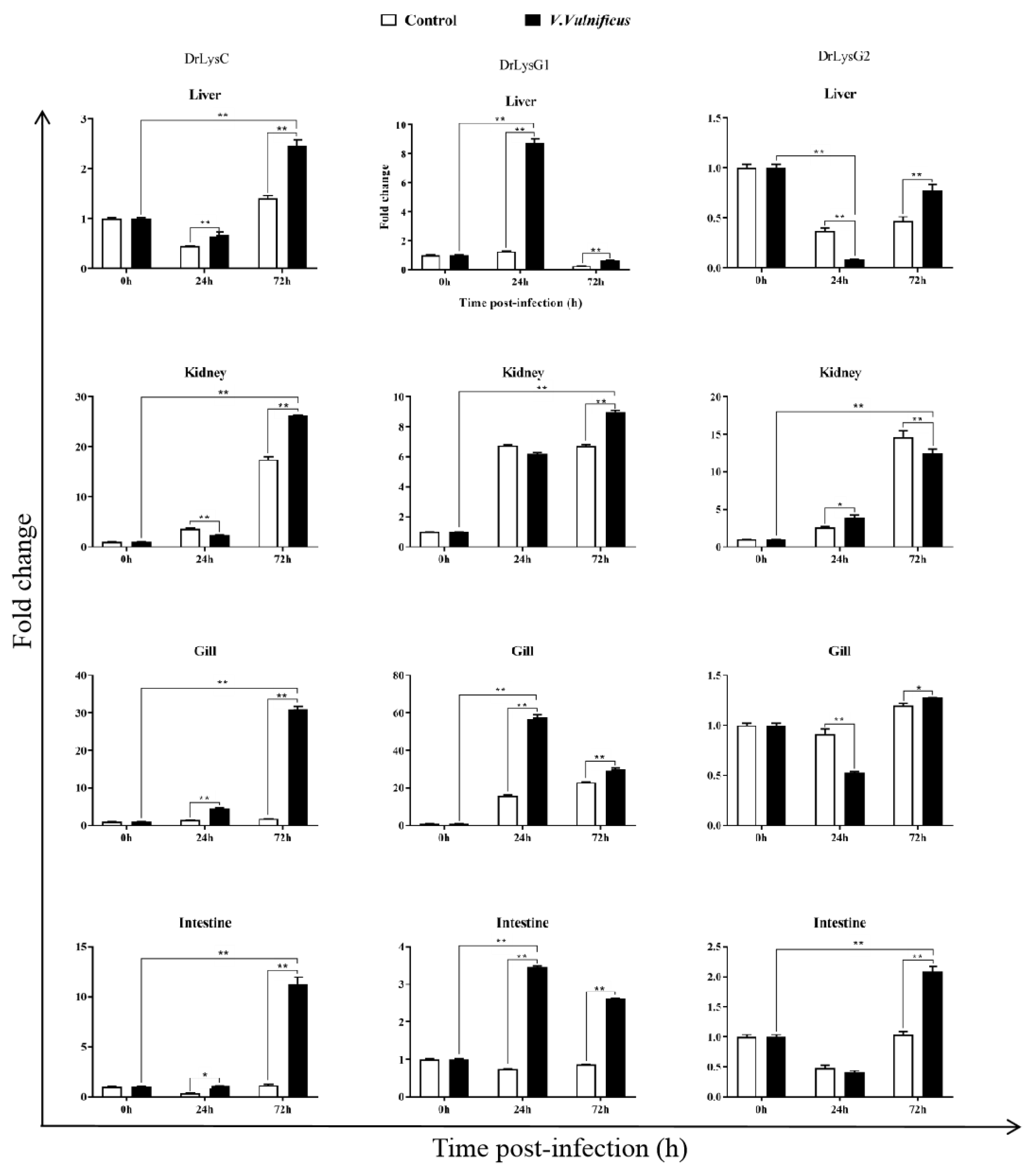
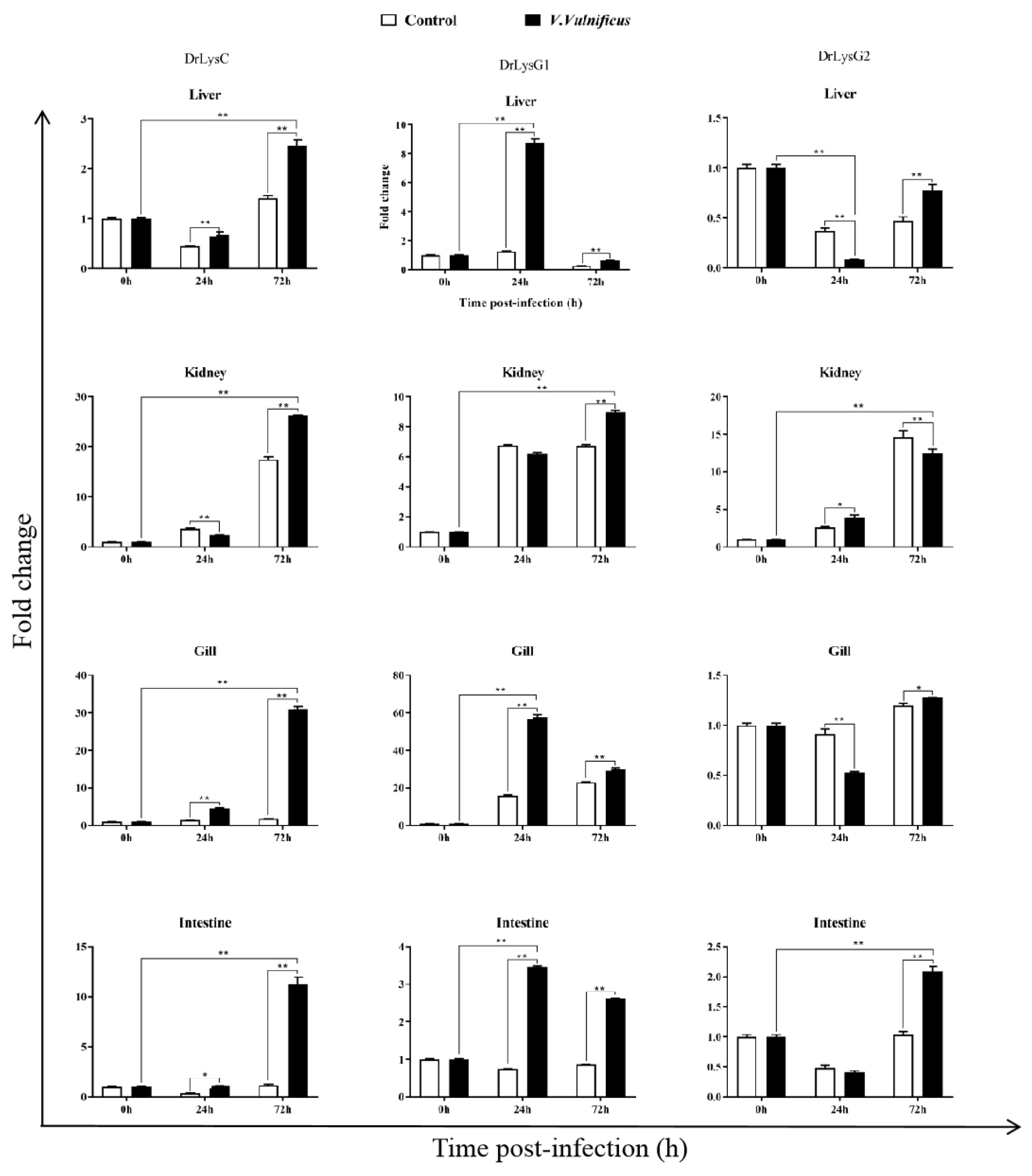
2.2. Expression of the Lysozymes in V. vulnificus-Infected Zebrafish
2.3. Expression of DrLysC and DrLysG1 in E. coli and Antimicrobial Activity Analysis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Culture Conditions
4.2. Experimental Fishes
4.3. Sequence and Phylogenetic Analysis
4.4. Bacterial Challenge
4.5. Sample Collection
4.6. RNA Extraction and qPCR Assay
4.7. Expression and Purification of Recombinant Protein
4.8. Determination of the Antimicrobial Activity of Over-Expressed rDrLysC and rDrLysG1
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferraboschi, P.; Ciceri, S.; Grisenti, P. Applications of Lysozyme, an Innate Immune Defense Factor, as an Alternative Antibiotic. Antibiotics 2021, 10, 1534. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.P.; Sun, B.G.; Li, J.; Sun, L. Characterization of a c-type lysozyme of Scophthalmus maximus: Expression, activity, and antibacterial effect. Fish Shellfish Immunol. 2013, 34, 46–54. [Google Scholar] [CrossRef]
- Cai, S.; Zhang, Y.; Wu, F.; Wu, R.; Yang, S.; Li, Y.; Xu, Y. Identification and functional characterization of a c-type lysozyme from Fenneropenaeus penicillatus. Fish Shellfish Immunol. 2019, 88, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, R.; Kim, J.; Kim, M.; Balasundaram, C.; Heo, M. Molecular characterization, phylogeny, and expression pattern of c-type lysozyme in kelp grouper, Epinephelus bruneus. Fish Shellfish Immunol. 2011, 31, 588–594. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Huang, Y.; Cai, J.; Huang, X.; Fu, J.; Qin, Q. Molecular cloning and characterization of c-type lysozyme gene in orange-spotted grouper, Epinephelus coioides. Fish Shellfish Immunol. 2012, 33, 186–196. [Google Scholar] [CrossRef]
- Fernandez-Trujillo, M.A.; Porta, J.; Manchado, M.; Borrego, J.J.; Alvarez, M.C.; Bejar, J. c-Lysozyme from Senegalese sole (Solea senegalensis): cDNA cloning and expression pattern. Fish Shellfish Immunol. 2008, 25, 697–700. [Google Scholar] [CrossRef]
- Sha, Z.X.; Wang, Q.L.; Liu, Y.; Chen, S.L. Identification and expression analysis of goose-type lysozyme in half-smooth tongue sole (Cynoglossus semilaevis). Fish Shellfish Immunol. 2012, 32, 914–921. [Google Scholar] [CrossRef]
- Savan, R.; Aman, A.; Sakai, M. Molecular cloning of G type lysozyme cDNA in common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2003, 15, 263–268. [Google Scholar] [CrossRef]
- Sun, B.J.; Wang, G.L.; Xie, H.X.; Gao, Q.; Nie, P. Gene structure of goose-type lysozyme in the mandarin fish Siniperca chuatsi with analysis on the lytic activity of its recombinant in Escherichia coli. Aquaculture 2005, 252, 106–113. [Google Scholar] [CrossRef]
- Wenbiao, Z.; Chen, T.; Xinhua, C. Molecular characterization of goose-type lysozyme homologue of large yellow croaker and its involvement in immune response induced by trivalent bacterial vaccine as an acute-phase protein. Immunol. Lett. 2007, 113, 107–116. [Google Scholar] [CrossRef]
- Larsen, A.N.; Solstad, T.; Svineng, G.; Seppola, M.; Jorgensen, T.O. Molecular characterisation of a goose-type lysozyme gene in Atlantic cod (Gadus morhua L.). Fish Shellfish Immunol. 2009, 26, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Sun, J.-S.; Li, S. The g-type lysozyme of Scophthalmus maximus has a broad substrate spectrum and is involved in the immune response against bacterial infection. Fish Shellfish Immunol. 2010, 30, 630–637. [Google Scholar] [CrossRef]
- Hikima, J.; Minagawa, S.; Hirono, I.; Aoki, T. Molecular cloning, expression and evolution of the Japanese flounder goose-type lysozyme gene, and the lytic activity of its recombinant protein. Biochim. Biophys. Acta. 2001, 1520, 35–44. [Google Scholar] [CrossRef]
- Wei, Q.; Mu, C.; Wang, C.; Zhao, J. Molecular characterization, expression, and antibacterial activity of a c-type lysozyme isolated from the manila clam, Ruditapes philippinarum. Fish Shellfish Immunol. 2018, 81, 502–508. [Google Scholar] [CrossRef]
- Kong, X.; Wang, S.; Jiang, H.; Nie, G.; Li, X. Responses of acid/alkaline phosphatase, lysozyme, and catalase activities and lipid peroxidation to mercury exposure during the embryonic development of goldfish Carassius auratus. Aquat. Toxicol. 2012, 120, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Tian, S.; Luo, K.; Zhang, Y.; Pan, H.; Zhang, W.; Mai, K. Dietary recombinant human lysozyme improves the growth, intestinal health, immunity and disease resistance of Pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2022, 121, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Whang, I.; Lee, Y.; Lee, S.; Oh, M.J.; Jung, S.J.; Choi, C.Y.; Lee, W.S.; Kim, H.S.; Kim, S.-J.; Lee, J. Characterization and expression analysis of a goose-type lysozyme from the rock bream Oplegnathus fasciatus, and antimicrobial activity of its recombinant protein. Fish Shellfish Immunol. 2011, 30, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, L.; Ke, L.; Chen, X.; Lin, C. CpG-ODN 2007 protects zebrafish (Danio rerio) against Vibrio vulnificus infection. Aquac. Res. 2021, 52, 897–905. [Google Scholar] [CrossRef]
- Liu, F.; Wen, Z. Cloning and expression pattern of the lysozyme C gene in zebrafish. Mech. Dev. 2002, 113, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, S. The role of lysozyme and complement in the antibacterial activity of zebrafish (Danio rerio) egg cytosol. Fish Shellfish Immunol. 2010, 29, 773–777. [Google Scholar] [CrossRef] [PubMed]
- Irwin, D.M.; Gong, Z. Molecular evolution of vertebrate goose-type lysozyme genes. J. Mol. Evol. 2003, 56, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.; Wan, Q.; Bathige, S.; Lee, J. Molecular characterization, transcriptional profiling, and antibacterial potential of G-type lysozyme from seahorse (Hippocampus abdominalis). Fish Shellfish Immunol. 2016, 58, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Meijuan, W.; Xianliang, Z.; Xianghui, K.; Li, W.; Dan, J.; Hongxu, Z. Molecular characterization and expressing analysis of the c-type and g-type lysozymes in Qihe crucian carp Carassius auratus. Fish Shellfish Immunol. 2016, 52, 210–220. [Google Scholar] [CrossRef]
- Zhang, S.; Xu, Q.; Boscari, E.; Du, H.; Qi, Z.; Li, Y.; Huang, J.; Di, J.; Yue, H.; Li, C.; et al. Characterization and expression analysis of g- and c-type lysozymes in Dabry’s sturgeon (Acipenser dabryanus). Fish Shellfish Immunol. 2018, 76, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Grinde, B. Lysozyme from rainbow trout, Salmo gairdneri Richardson, as an antibacterial agent against fish pathogens. J. Fish Dis. 1989, 12, 95–104. [Google Scholar] [CrossRef]
- Gong, Y.; Kim, Y.R. Therapeutic potential of otilonium bromide against Vibrio vulnificus. Res. Microbiol. 2022, 174, 103992. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Amino Acids | Molecular Weight (kD) | Signal Peptide | Conserved Cysteine Residue (Cys) | Disulfide Bond |
|---|---|---|---|---|---|
| DrLysC | 151 | 17.1 | Yes | 8 | 4 |
| DrLysG1 | 196 | 21.6 | Yes | 2 | 0 |
| DrLysG2 | 191 | 21.1 | No | 2 | 0 |
| Gene Symbol | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| β-actin | CACTTCACGCCGACTCAAAC | TCGGGGATGCTTATTTGCCA |
| DrLysC | GGCGTGGATGTCCTCGTGT | TCGGTGGGTCTTAAACCTGCT |
| DrLysG1 | CTGGTAGGTGCGTGGGACA | GGGCAACAACATCATTAGCG |
| DrLysG2 | CAAGTGTAAAATCTTCAAAGTTGCCA | TCGCCCCATCCTTTCAACA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Chen, X.; Song, T.-Y.; Ge, J.-Q. Expression Profiles of Zebrafish (Danio rerio) Lysozymes and Preparation of c-Type Lysozyme with High Bacteriolytic Activity against Vibrio vulnificus. Antibiotics 2022, 11, 1803. https://doi.org/10.3390/antibiotics11121803
Chen H, Chen X, Song T-Y, Ge J-Q. Expression Profiles of Zebrafish (Danio rerio) Lysozymes and Preparation of c-Type Lysozyme with High Bacteriolytic Activity against Vibrio vulnificus. Antibiotics. 2022; 11(12):1803. https://doi.org/10.3390/antibiotics11121803
Chicago/Turabian StyleChen, Hua, Xi Chen, Tie-Ying Song, and Jun-Qing Ge. 2022. "Expression Profiles of Zebrafish (Danio rerio) Lysozymes and Preparation of c-Type Lysozyme with High Bacteriolytic Activity against Vibrio vulnificus" Antibiotics 11, no. 12: 1803. https://doi.org/10.3390/antibiotics11121803
APA StyleChen, H., Chen, X., Song, T.-Y., & Ge, J.-Q. (2022). Expression Profiles of Zebrafish (Danio rerio) Lysozymes and Preparation of c-Type Lysozyme with High Bacteriolytic Activity against Vibrio vulnificus. Antibiotics, 11(12), 1803. https://doi.org/10.3390/antibiotics11121803