
Molecular Detection of Carbapenemases in Enterobacterales: A Comparison of Real-Time Multiplex PCR and Whole-Genome Sequencing

 , and
, and
Abstract
1. Introduction
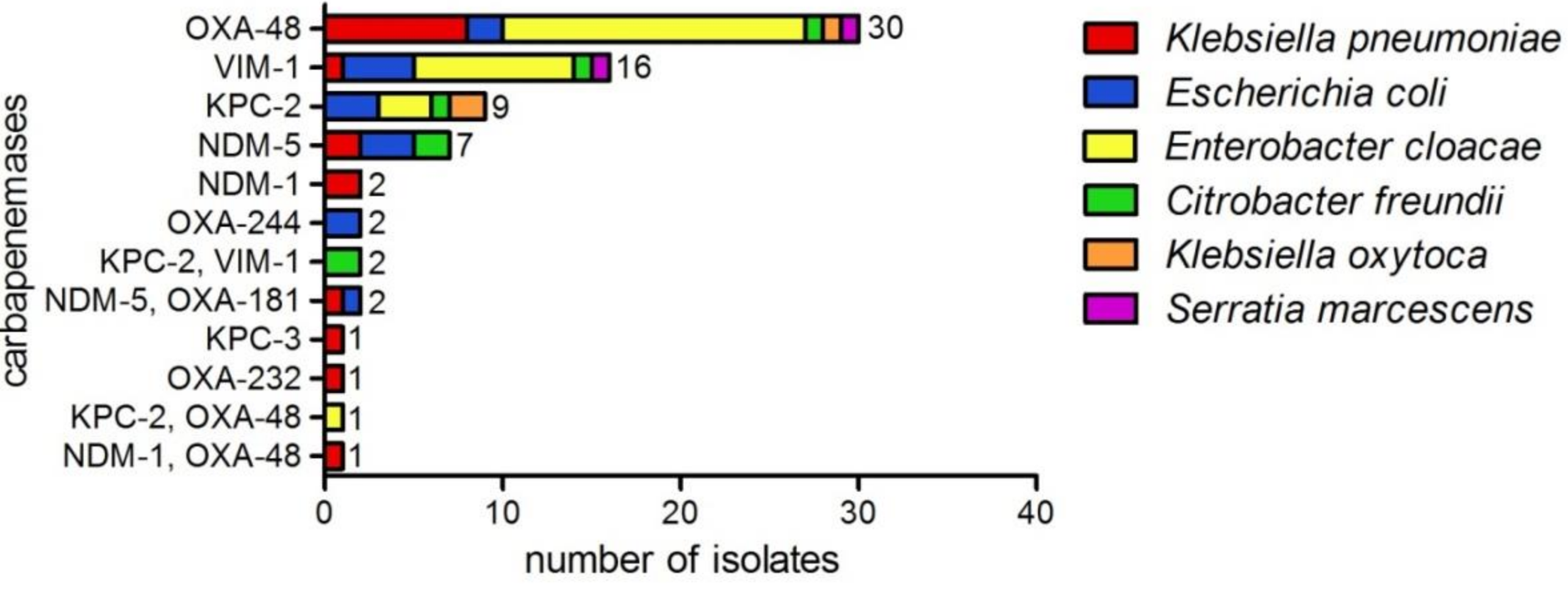
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Carbapenem Inactivation Method
4.3. DNA Extraction
4.4. Multiplex Real-Time PCR
4.5. Whole-Genome Sequencing
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
| Sample ID | Species | CIM | RT-qPCR | WGS | WGS Reanalyzed | Coverage | N50 |
|---|---|---|---|---|---|---|---|
| KE9539 | E. coli | positive | blaKPC | blaKPC-2 | 48 | 535,993 | |
| KE9246 | E. coli | positive | blaKPC | blaKPC-2 | 53 | 135,761 | |
| KE9526 | E. cloacae | positive | blaKPC | blaKPC-2 | 52 | 363,822 | |
| KE9478 | E. cloacae | positive | blaKPC | blaKPC-2 | 96 | 363,822 | |
| BK31926 | E. coli | positive | blaKPC | blaKPC-2 | 29 | 120,862 | |
| KE9621 | K. pneumoniae | positive | blaKPC | blaKPC-3 | 35 | 386,401 | |
| KE9498 | C. freundii | positive | blaKPC | blaKPC-2 | 31 | 200,582 | |
| KE9038 | K. oxytoca | positive | blaKPC | blaKPC-2 | 50 | 285,607 | |
| KE9326 | K. oxytoca | positive | blaKPC | negative | blaKPC-2 | 42 | 109,274 |
| KE9511 | C. freundii | positive | blaKPC, blaVIM | blaKPC-2, blaVIM-1 | 30 | 198,406 | |
| KE9378 | C. freundii | positive | blaKPC, blaVIM | blaKPC-2 | blaKPC-2, blaVIM-1 | 39 | 201,178 |
| KE9132 | E. cloacae | positive | blaKPC | blaKPC-2 | 49 | 363,822 | |
| KE9520 | K. pneumoniae | positive | blaNDM | blaNDM-5 | 53 | 186,575 | |
| KE9434 | K. pneumoniae | positive | blaNDM | blaNDM-5 | 34 | 292,061 | |
| KE9521 | E. coli | positive | blaNDM, blaOXA-48-like | blaNDM-5, blaOXA-181 | 61 | 106,471 | |
| KE9395 | E. coli | positive | blaNDM | blaNDM-5 | 54 | 94,083 | |
| KE9433 | E. coli | positive | blaNDM | blaNDM-5 | 36 | 214,212 | |
| KE9636 | K. pneumoniae | positive | blaNDM, blaOXA-48-like | blaNDM-1, blaOXA-48 | 27 | 383,090 | |
| KE9616 | C. freundii | positive | blaNDM | blaNDM-5 | 50 | 186,958 | |
| KE9522 | E. coli | positive | blaNDM | blaNDM-5 | 103 | 269,697 | |
| KE9593 | K. pneumoniae | positive | blaNDM, blaOXA-48-like | blaNDM-5, blaOXA-181 | 38 | 296,725 | |
| D3014 | C. freundii | positive | blaNDM | blaNDM-5 | 36 | 186,959 | |
| KE9449 | K. pneumoniae | positive | blaNDM | negative | blaNDM-1 | 25 | 220,843 |
| KE9500 | K. pneumoniae | positive | blaNDM | blaNDM-1 | 33 | 536,321 | |
| KE9382 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 27 | 374,725 | |
| KE9492 | K. pneumoniae | positive | blaOXA-48-like | blaOXA-232 | 30 | 242,997 | |
| KE9629 | E. coli | positive | blaOXA-48-like | blaOXA-244 | 33 | 238,467 | |
| KE9025 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 49 | 272,750 | |
| KE9469 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 76 | 374,315 | |
| KE9472 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 98 | 382,653 | |
| KE9424 | K. pneumoniae | positive | blaOXA-48-like | blaOXA-48 | 36 | 184,292 | |
| KE9499 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 66 | 486,681 | |
| KE9400 | K. pneumoniae | positive | blaOXA-48-like | blaOXA-48 | 45 | 208,351 | |
| KE9468 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 80 | 383,026 | |
| KE9638 | E. coli | positive | blaOXA-48-like | blaOXA-244 | 37 | 156,925 | |
| KE9493 | E. cloacae | positive | blaOXA-48-like, blaKPC | blaOXA-48 | blaKPC-2, blaOXA-48 | 44 | 530,933 |
| KE9456 | K. oxytoca | positive | blaOXA-48-like | blaOXA-48 | 28 | 223,596 | |
| KE9443 | K. pneumoniae | positive | blaOXA-48-like | blaOXA-48 | 27 | 225,118 | |
| KE9354 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 66 | 486,663 | |
| BK32270 | E. coli | positive | blaOXA-48-like | blaOXA-48 | 35 | 117,967 | |
| KE9626 | E. coli | positive | blaOXA-48-like | blaOXA-48 | 53 | 196,578 | |
| KE9208 | S. marcescens | positive | blaOXA-48-like | blaOXA-48 | 58 | 2,797,497 | |
| D2902 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 64 | 302,960 | |
| KE9541 | K. pneumoniae | positive | blaOXA-48-like | blaOXA-48 | 47 | 427,613 | |
| KE9554 | C. freundii | positive | blaOXA-48-like | blaOXA-48 | 39 | 165,554 | |
| KE9338 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 47 | 374,725 | |
| KE9355 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 109 | 486,681 | |
| KE9328 | K. pneumoniae | positive | blaOXA-48-like | blaOXA-48 | 43 | 274,145 | |
| KE9510 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 31 | 491,022 | |
| D3070 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 44 | 372,768 | |
| KE9428 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 36 | 486,663 | |
| KE9527 | E. cloacae | positive | blaOXA-48-like | negative | blaOXA-48 | 27 | 339,153 |
| D3018 | K. pneumoniae | positive | blaOXA-48-like | blaOXA-48 | 25 | 473,650 | |
| D3082 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 62 | 486,663 | |
| KE9637 | K. pneumoniae | positive | blaOXA-48-like | blaOXA-48 | 36 | 876,600 | |
| D3081 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 85 | 383,026 | |
| EX1012 | K. pneumoniae | positive | blaOXA-48-like | blaOXA-48 | 39 | 223,327 | |
| D3078 | E. cloacae | positive | blaOXA-48-like | blaOXA-48 | 54 | 486,828 | |
| KE9366 | E. coli | positive | blaVIM | blaVIM-1 | 38 | 215,473 | |
| KE9563 | E. cloacae | positive | blaVIM | blaVIM-1 | 35 | 377,920 | |
| KE9409 | E. cloacae | positive | blaVIM | blaVIM-1 | 46 | 486,118 | |
| KE9414 | E. cloacae | positive | blaVIM | blaVIM-1 | 38 | 161,463 | |
| KE9365 | K. pneumoniae | positive | blaVIM | blaVIM-1 | 32 | 232,474 | |
| KE9538 | S. marcescens | positive | blaVIM | blaVIM-1 | 40 | 1,130,420 | |
| KE9585 | E. cloacae | positive | blaVIM | blaVIM-1 | 25 | 287,090 | |
| KE9559 | C. freundii | positive | blaVIM | blaVIM-1 | 41 | 163,976 | |
| KE9549 | E. coli | positive | blaVIM | blaVIM-1 | 47 | 279,067 | |
| KE9548 | E. cloacae | positive | blaVIM | blaVIM-1 | 46 | 230,814 | |
| KE9579 | E. coli | positive | blaVIM | blaVIM-1 | 39 | 112,495 | |
| KE9474 | E. cloacae | positive | blaVIM | blaVIM-1 | 38 | 290,132 | |
| KE9462 | E. cloacae | positive | blaVIM | blaVIM-1 | 33 | 502,528 | |
| KE9560 | E. cloacae | positive | blaVIM | blaVIM-1 | 40 | 290,117 | |
| KE9575 | E. cloacae | positive | blaVIM | blaVIM-1 | 38 | 389,538 | |
| KE9536 | E. coli | positive | blaVIM | negative | blaVIM-1 | 44 | 377,920 |
| D2923 | E. cloacae | negative | negative | negative | 58 | 203,439 | |
| KE9347 | E. cloacae | negative | negative | negative | 79 | 439,426 | |
| KE9591 | E. cloacae | negative | negative | negative | 47 | 279,225 | |
| KE9576 | E. coli | negative | negative | negative | 27 | 228,481 | |
| KE9599 | E. coli | negative | negative | negative | 37 | 281,932 | |
| KE9623 | E. coli | negative | negative | negative | 50 | 93,960 | |
| KE9633 | K. aerogenes | negative | negative | negative | 40 | 495,847 | |
| KE9068 | C. freundii | negative | negative | negative | 46 | 176,242 | |
| KE8986 | E. cloacae | negative | negative | negative | 47 | 230,847 | |
| KE9344 | E. cloacae | negative | negative | negative | 48 | 235,301 | |
| KE9475 | E. cloacae | negative | negative | negative | 40 | 208,042 | |
| KE9083 | E. coli | negative | negative | negative | 57 | 208,544 | |
| KE9425 | K. aerogenes | negative | negative | negative | 48 | 902,223 | |
| KE9614 | K. aerogenes | negative | negative | negative | 62 | 429,809 | |
| D3017 | K. pneumoniae | negative | negative | negative | 27 | 232,937 | |
| KE9095 | K. pneumoniae | negative | negative | negative | 66 | 237,389 | |
| KE9171 | K. pneumoniae | negative | negative | negative | 54 | 481,561 | |
| KE9039 | S. marcescens | negative | negative | negative | 40 | 1,228,444 |
References
- Nordmann, P.; Dortet, L.; Poirel, L. Carbapenem resistance in Enterobacteriaceae: Here is the storm! Trends Mol. Med. 2012, 18, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215 (Suppl. 1), S28–S36. [Google Scholar] [CrossRef]
- Tzouvelekis, L.S.; Markogiannakis, A.; Psichogiou, M.; Tassios, P.T.; Daikos, G.L. Carbapenemases in Klebsiella pneumoniae and Other Enterobacteriaceae: An Evolving Crisis of Global Dimensions. Clin. Microbiol. Rev. 2012, 25, 682–707. [Google Scholar] [CrossRef]
- Delgado-Valverde, M.; Sojo-Dorado, J.; Pascual, Á.; Rodríguez-Baño, J. Clinical management of infections caused by multidrug-resistant Enterobacteriaceae. Ther. Adv. Infect. Dis. 2013, 1, 49–69. [Google Scholar] [CrossRef]
- Van Duin, D.; Doi, Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence 2017, 8, 460–469. [Google Scholar] [CrossRef]
- Exner, M.; Bhattacharya, S.; Christiansen, B.; Gebel, J.; Goroncy-Bermes, P.; Hartemann, P.; Heeg, P.; Ilschner, C.; Kramer, A.; Larson, E.; et al. Antibiotic resistance: What is so special about multidrug-resistant Gram-negative bacteria? GMS Hyg. Infect. Control 2017, 12, 3. [Google Scholar]
- Paterson, D.; Doi, Y. Carbapenemase-Producing Enterobacteriaceae. Semin. Respir. Crit. Care Med. 2015, 36, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Nordmann, P.; Cornaglia, G. Carbapenemase-producing Enterobacteriaceae: A call for action! Clin. Microbiol. Infect. 2012, 18, 411–412. [Google Scholar] [CrossRef]
- Tijet, N.; Boyd, D.; Patel, S.N.; Mulvey, M.R.; Melano, R.G. Evaluation of the Carba NP Test for Rapid Detection of Carbapenemase-Producing Enterobacteriaceae and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2013, 57, 4578–4580. [Google Scholar] [CrossRef]
- Amjad, A.; Mirza, I.; Abbasi, S.; Farwa, U.; Malik, N.; Zia, F. Modified Hodge test: A simple and effective test for detection of carbapenemase production. Iran. J. Microbiol. 2011, 3, 189–193. [Google Scholar]
- Van Dijk, K.; Voets, G.M.; Scharringa, J.; Voskuil, S.; Fluit, A.C.; Rottier, W.C.; Hall, M.A.L.; Stuart, J.W.T.C. A disc diffusion assay for detection of class A, B and OXA-48 carbapenemases in Enterobacteriaceae using phenyl boronic acid, dipicolinic acid and temocillin. Clin. Microbiol. Infect. 2014, 20, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Antonelli, A.; Arena, F.; Giani, T.; Colavecchio, O.L.; Valeva, S.V.; Paule, S.; Boleij, P.; Rossolini, G.M. Performance of the BD MAX™ instrument with Check-Direct CPE real-time PCR for the detection of carbapenemase genes from rectal swabs, in a setting with endemic dissemination of carbapenemase-producing Enterobacteriaceae. Diagn. Microbiol. Infect. Dis. 2016, 86, 30–34. [Google Scholar] [CrossRef]
- Hofko, M.; Mischnik, A.; Kaase, M.; Zimmermann, S.; Dalpke, A.H. Detection of Carbapenemases by Real-Time PCR and Melt Curve Analysis on the BD Max System. J. Clin. Microbiol. 2014, 52, 1701–1704. [Google Scholar] [CrossRef] [PubMed]
- Dallenne, C.; da Costa, A.; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef]
- Schurch, A.C.; van Schaik, W. Challenges and opportunities for whole-genome sequencing-based surveillance of antibiotic resistance. Ann. N. Y. Acad. Sci. 2017, 1388, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Baeza, L.L.; Pfennigwerth, N.; Greissl, C.; Göttig, S.; Saleh, A.; Stelzer, Y.; Gatermann, S.; Hamprecht, A. Comparison of five methods for detection of carbapenemases in Enterobacterales with proposal of a new algorithm. Clin. Microbiol. Infect. 2019, 25, 1286.e9–1286.e15. [Google Scholar] [CrossRef]
- Baeza, L.L.; Pfennigwerth, N.; Hamprecht, A. Rapid and Easy Detection of Carbapenemases in Enterobacterales in the Routine Laboratory Using the New Gene POC Carba/Revogene Carba C Assay. J. Clin. Microbiol. 2019, 57. [Google Scholar] [CrossRef]
- Han, R.; Guo, Y.; Peng, M.; Shi, Q.; Wu, S.; Yang, Y.; Zheng, Y.; Yin, D.; Hu, F. Evaluation of the Immunochromatographic NG-Test Carba 5, RESIST-5 O.O.K.N.V., and IMP K-SeT for Rapid Detection of KPC-, NDM-, IMP-, VIM-type, and OXA-48-like Carbapenemase Among Enterobacterales. Front. Microbiol. 2020, 11, 609856. [Google Scholar] [CrossRef] [PubMed]
- Ellington, M.; Ekelund, O.; Aarestrup, F.; Canton, R.; Doumith, M.; Giske, C.; Grundman, H.; Hasman, H.; Holden, M.; Hopkins, K.; et al. The role of whole genome sequencing in antimicrobial susceptibility testing of bacteria: Report from the EUCAST Subcommittee. Clin. Microbiol. Infect. 2017, 23, 2–22. [Google Scholar] [CrossRef]
- Pfennigwerth, N. Bericht des Nationalen Referenzzentrums (NRZ) für gramnegative Krankenhauserreger – Zeitraum 1. Januar 2017 – 31. Dezember 2017. J. Epidemiol. Bull. 2018, 28, 263–267. [Google Scholar]
- Pfennigwerth, N. Bericht des Nationalen Referenzzentrums (NRZ) für gramnegative Krankenhauserreger, 2018. J. Epidemiol. Bull. 2019, 31, 289–294. [Google Scholar]
- Pfennigwerth, N. Bericht des Nationalen Referenzzentrums (NRZ) für gramnegative Krankenhauserreger, 2019. J. Epidemiol. Bull. 2020, 26, 3–10. [Google Scholar]
- Nurjadi, D.; Zizmann, E.; Chanthalangsy, Q.; Heeg, K.; Boutin, S. Integrative Analysis of Whole Genome Sequencing and Phenotypic Resistance Toward Prediction of Trimethoprim-Sulfamethoxazole Resistance in Staphylococcus aureus. Front. Microbiol. 2020, 11, 607842. [Google Scholar] [CrossRef] [PubMed]
- Van der Zwaluw, K.; de Haan, A.; Pluister, G.N.; Bootsma, H.J.; de Neeling, A.J.; Schouls, L.M. The carbapenem inactivation method (CIM), a simple and low-cost alternative for the Carba NP test to assess phenotypic carbapenemase activity in gram-negative rods. PLoS ONE 2015, 10, e0123690. [Google Scholar] [CrossRef] [PubMed]
- Probst, K.; Boutin, S.; Bandilla, M.; Heeg, K.; Dalpke, A.H. Fast and automated detection of common carbapenemase genes using multiplex real-time PCR on the BD MAX™ system. J. Microbiol. Methods 2021, 185, 106224. [Google Scholar] [CrossRef] [PubMed]
- Nurjadi, D.; Boutin, S.; Dalpke, A.; Heeg, K.; Zanger, P. Draft Genome Sequence of Staphylococcus aureus Strain HD1410, Isolated from a Persistent Nasal Carrier. Genome Announc. 2018, 6, e00411–e00418. [Google Scholar] [CrossRef] [PubMed]
- Eichel, P.C.V.; Boutin, S.; Poschl, J.; Heeg, K.; Nurjadi, D. Altering antibiotic regimen as additional control measure in suspected multi-drug-resistant Enterobacter cloacae outbrake in neonatal intensive care unit. J. Hosp. Infect. 2019, 104, 144–149. [Google Scholar] [CrossRef]

| CIM | |||
|---|---|---|---|
| Positive | Negative | ||
| RT-qPCR | positive negative | 74 0 | 0 18 |
| WGS | positive negative | 70(74) 1 4(0) 1 | 0 18 |
| WGS | |||
|---|---|---|---|
| Positive | Negative | ||
| RT-qPCR | positive negative | 68(74) 1 0 | 6(0) 1 18 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Probst, K.; Nurjadi, D.; Heeg, K.; Frede, A.-M.; Dalpke, A.H.; Boutin, S. Molecular Detection of Carbapenemases in Enterobacterales: A Comparison of Real-Time Multiplex PCR and Whole-Genome Sequencing. Antibiotics 2021, 10, 726. https://doi.org/10.3390/antibiotics10060726
Probst K, Nurjadi D, Heeg K, Frede A-M, Dalpke AH, Boutin S. Molecular Detection of Carbapenemases in Enterobacterales: A Comparison of Real-Time Multiplex PCR and Whole-Genome Sequencing. Antibiotics. 2021; 10(6):726. https://doi.org/10.3390/antibiotics10060726
Chicago/Turabian StyleProbst, Katja, Dennis Nurjadi, Klaus Heeg, Anne-Marie Frede, Alexander H. Dalpke, and Sébastien Boutin. 2021. "Molecular Detection of Carbapenemases in Enterobacterales: A Comparison of Real-Time Multiplex PCR and Whole-Genome Sequencing" Antibiotics 10, no. 6: 726. https://doi.org/10.3390/antibiotics10060726
APA StyleProbst, K., Nurjadi, D., Heeg, K., Frede, A.-M., Dalpke, A. H., & Boutin, S. (2021). Molecular Detection of Carbapenemases in Enterobacterales: A Comparison of Real-Time Multiplex PCR and Whole-Genome Sequencing. Antibiotics, 10(6), 726. https://doi.org/10.3390/antibiotics10060726