


A Deep Learning Framework for Parameter Estimation in 1D Marine Ecosystem Model

 and
and 
Abstract
1. Introduction
2. Methods
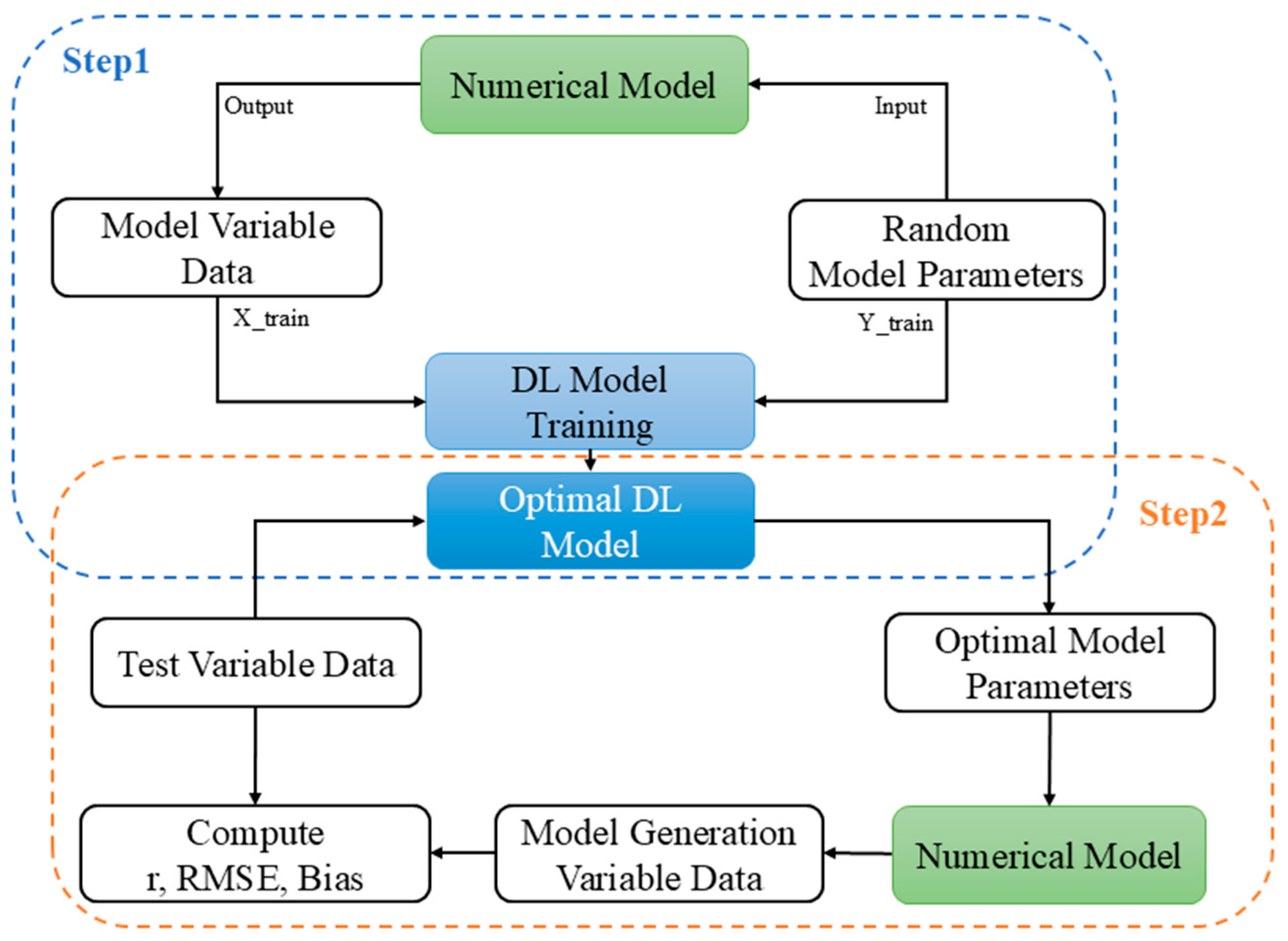
2.1. Experiment Setup
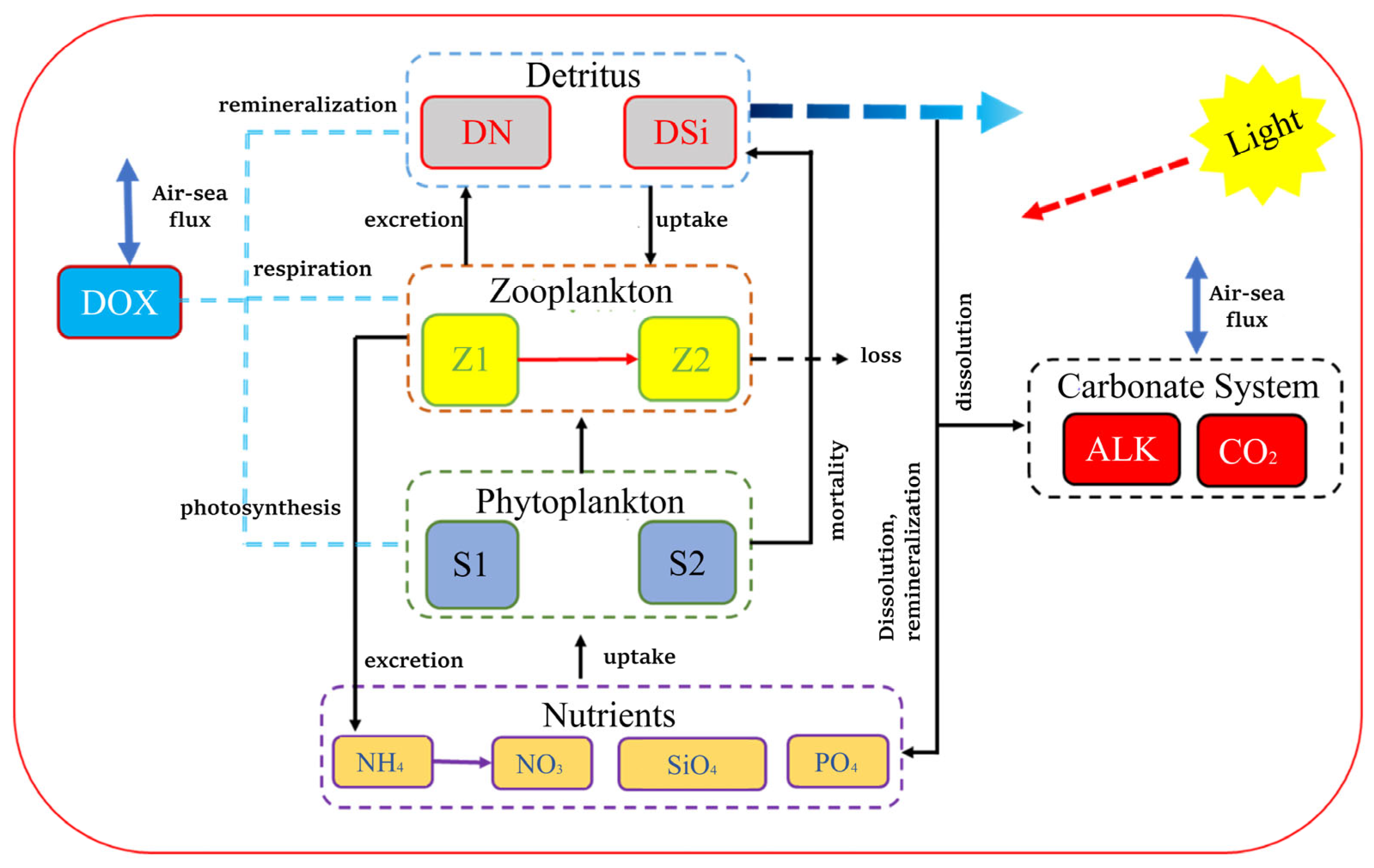
2.2. One-Dimensional Numerical Model
- (1)
- Phytoplankton-related Parameters
- (2)
- Zooplankton-related Parameters
- (3)
- Other Parameters
2.3. Deep Learning Framework
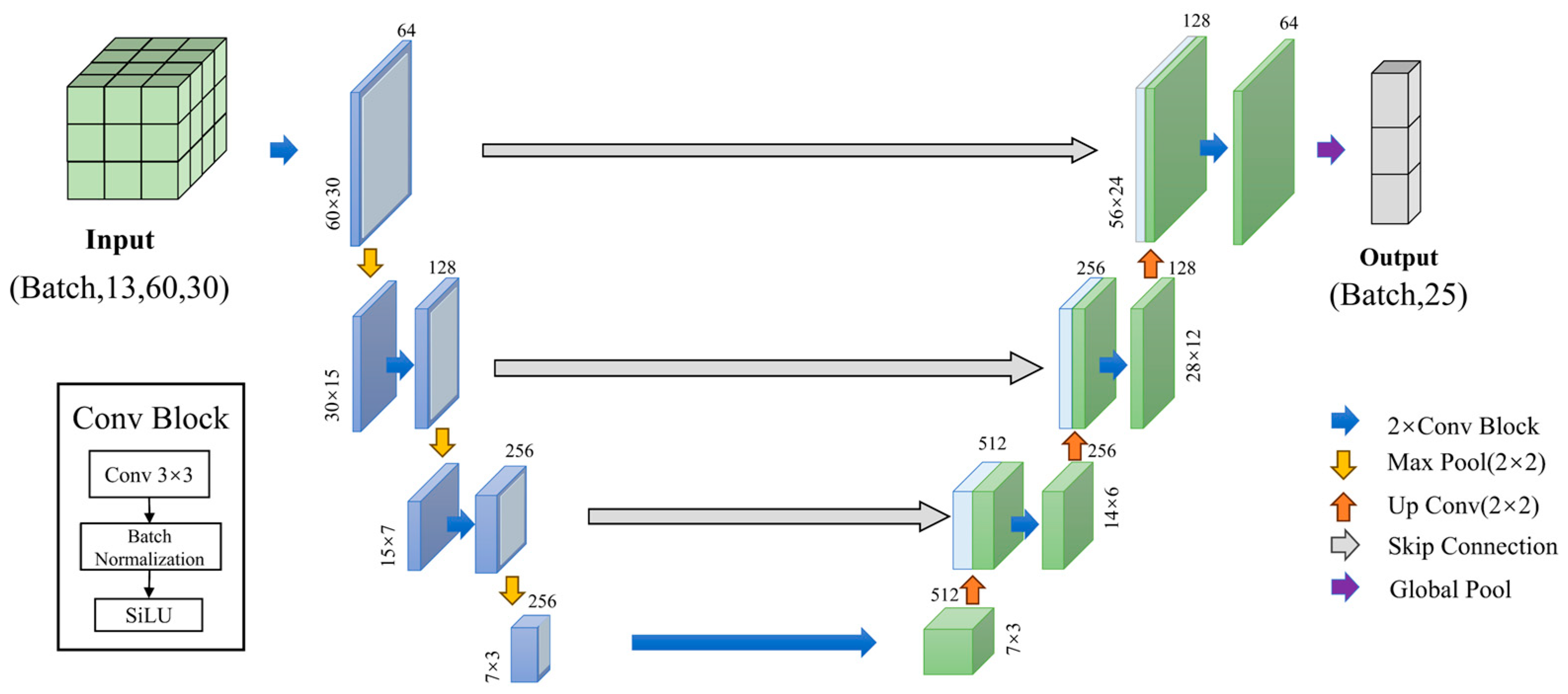
2.4. U-Net Model Training and Evaluation
- (1)
- Dataset Splitting
- (2)
- Data Preprocessing
- (3)
- Model Training
- (4)
- Performance Evaluation
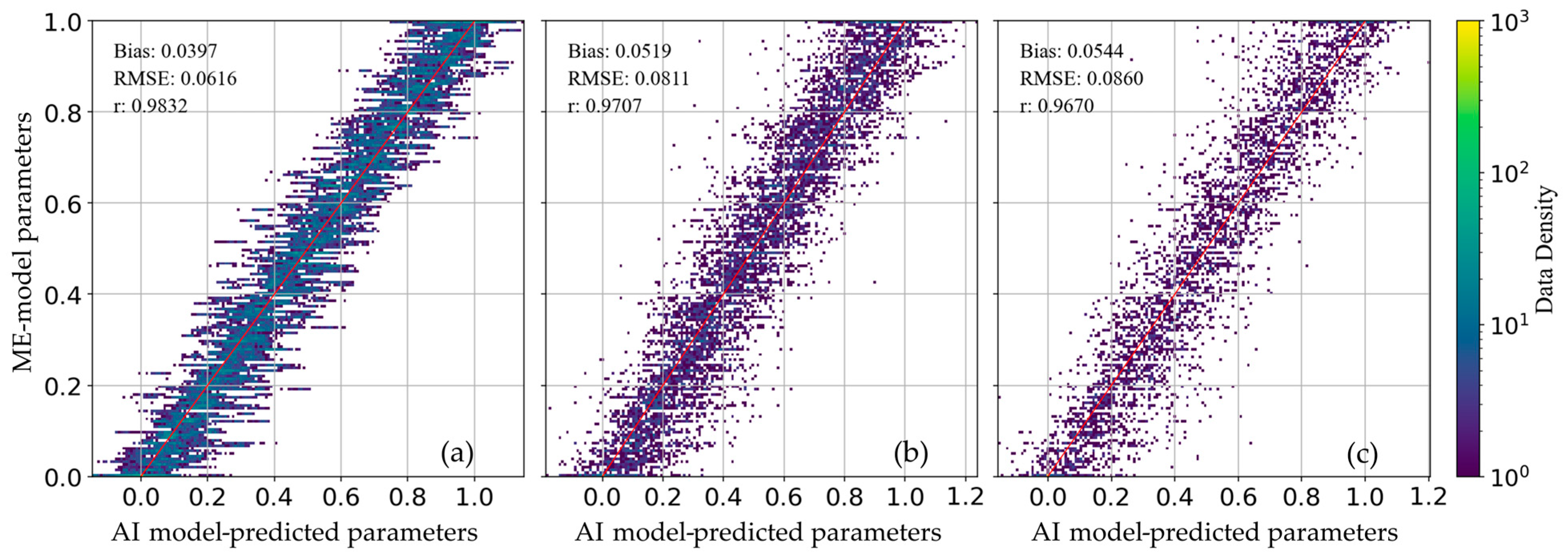
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Gattuso, J.; Magnan, A.; Bopp, L.; Cheung, W.; Duarte, C.; Hinkel, J.; McLeod, E.; Micheli, F.; Oschlies, A.; Williamson, P.; et al. Ocean Solutions to Address Climate Change and Its Effects on Marine Ecosystems. Front. Mar. Sci. 2018, 5, 337. [Google Scholar] [CrossRef]
- Doney, S.; Ruckelshaus, M.; Emmett Duffy, J.; Barry, J.; Chan, F.; English, C.; Galindo, H.; Grebmeier, J.; Hollowed, A.; Knowlton, N.; et al. Climate Change Impacts on Marine Ecosystems. Annu. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef]
- Shu, C.; Xiu, P.; Xing, X.; Qiu, G.; Ma, W.; Brewin, R.; Ciavatta, S. Biogeochemical Model Optimization by Using Satellite-Derived Phytoplankton Functional Type Data and BGC-Argo Observations in the Northern South China Sea. Remote Sens. 2022, 14, 1297. [Google Scholar] [CrossRef]
- Fang, W.; Geng, B.; Xiu, P. Typhoon Effects on the Vertical Chlorophyll Distribution on the Northern Shelf of the South China Sea. J. Geophys. Res. Ocean. 2022, 127, e2022JC019350. [Google Scholar] [CrossRef]
- Geng, B.; Xiu, P.; Liu, N.; He, X.; Chai, F. Biological Response to the Interaction of a Mesoscale Eddy and the River Plume in the Northern South China Sea. J. Geophys. Res. Ocean. 2021, 126, e2021JC017244. [Google Scholar] [CrossRef]
- Riley, G.A. Quantitative ecology of the plankton of the western North Atlantic. Bull. Bingham Oceanogr. Collect. 1949, 12, 1–169. [Google Scholar]
- Heinle, A.; Slawig, T. Internal dynamics of NPZD type ecosystem models. Ecol. Model. 2013, 254, 33–42. [Google Scholar] [CrossRef]
- Franks, P. NPZ models of plankton dynamics: Their construction, coupling to physics, and application. J. Oceanogr. 2002, 58, 379–387. [Google Scholar] [CrossRef]
- Fasham, M.; Ducklow, H.; McKelvie, S. A nitrogen-based model of plankton dynamics in the oceanic mixed layer. J. Mar. Res. 1990, 48, 591–639. [Google Scholar] [CrossRef]
- Allen, J.; Siddorn, J.; Blackford, J.; Gilbert, F. Turbulence as a control on the microbial loop in a temperate seasonally stratified marine systems model. J. Sea Res. 2004, 52, 1–20. [Google Scholar] [CrossRef]
- Fennel, W. Towards bridging biogeochemical and fish-production models. J. Mar. Syst. 2008, 71, 171–194. [Google Scholar] [CrossRef]
- Chai, F.; Dugdale, R.; Peng, T.; Wilkerson, F.; Barber, R. One-dimensional ecosystem model of the equatorial Pacific upwelling system. Part I: Model development and silicon and nitrogen cycle. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 2713–2745. [Google Scholar] [CrossRef]
- Xiu, P.; Chai, F. Modeled biogeochemical responses to mesoscale eddies in the South China Sea. J. Geophys. Res. Ocean. 2011, 116, C10006. [Google Scholar] [CrossRef]
- Guo, L.; Xiu, P.; Chai, F.; Xue, H.; Wang, D.; Sun, J. Enhanced chlorophyll concentrations induced by Kuroshio intrusion fronts in the northern South China Sea. Geophys. Res. Lett. 2017, 44, 11565–11572. [Google Scholar] [CrossRef]
- Ma, W.; Xiu, P.; Chai, F.; Li, H. Seasonal variability of the carbon export in the central South China Sea. Ocean Dyn. 2019, 69, 955–966. [Google Scholar] [CrossRef]
- Zhou, F.; Chai, F.; Huang, D.; Xue, H.; Chen, J.; Xiu, P.; Xuan, J.; Li, J.; Zeng, D.; Ni, X. Investigation of hypoxia off the Changjiang Estuary using a coupled model of ROMS-CoSiNE. Prog. Oceanogr. 2017, 159, 237–254. [Google Scholar] [CrossRef]
- Xiu, P.; Chai, F.; Curchitser, E.N.; Castruccio, F.S. Future changes in coastal upwelling ecosystems with global warming: The case of the California Current System. Sci. Rep. 2018, 8, 2866. [Google Scholar] [CrossRef]
- Fasham, M. Variations in the seasonal cycle of biological production in subarctic oceans: A model sensitivity analysis. Deep Sea Res. Part I Oceanogr. Res. Pap. 1995, 42, 1111–1149. [Google Scholar] [CrossRef]
- LöPtien, U.; Dietze, H. Effects of parameter indeterminacy in pelagic biogeochemical modules of Earth System Models on projections into a warming future: The scale of the problem. Glob. Biogeochem. Cycles 2017, 31, 1155–1172. [Google Scholar] [CrossRef]
- Gharamti, M.; Samuelsen, A.; Bertino, L.; Simon, E.; Korosov, A.; Daewel, U. Online tuning of ocean biogeochemical model parameters using ensemble estimation techniques: Application to a one-dimensional model in the North Atlantic. J. Mar. Syst. 2017, 168, 1–16. [Google Scholar] [CrossRef]
- Xiao, Y.; Friedrichs, M. The assimilation of satellite-derived data into a one-dimensional lower trophic level marine ecosystem model. J. Geophys. Res. Ocean. 2014, 119, 2691–2712. [Google Scholar] [CrossRef]
- Mattern, J.; Dowd, M.; Fennel, K. Particle filter-based data assimilation for a three-dimensional biological ocean model and satellite observations. J. Geophys. Res. Ocean. 2013, 118, 2746–2760. [Google Scholar] [CrossRef]
- Kuroda, H.; Kishi, M. A data assimilation technique applied to estimate parameters for the NEMURO marine ecosystem model. Ecol. Model. 2004, 172, 69–85. [Google Scholar] [CrossRef]
- Fennel, K.; Losch, M.; Schröter, J.; Wenzel, M. Testing a marine ecosystem model: Sensitivity analysis and parameter optimization. J. Mar. Syst. 2001, 28, 45–63. [Google Scholar] [CrossRef]
- Doron, M.; Brasseur, P.; Brankart, J. Stochastic estimation of biogeochemical parameters of a 3D ocean coupled physical–biogeochemical model: Twin experiments. J. Mar. Syst. 2011, 87, 194–207. [Google Scholar] [CrossRef]
- Hoshiba, Y.; Hirata, T.; Shigemitsu, M.; Nakano, H.; Hashioka, T.; Masuda, Y.; Yamanaka, Y. Biological data assimilation for parameter estimation of a phytoplankton functional type model for the western North Pacific. Ocean Sci. 2018, 14, 371–386. [Google Scholar] [CrossRef]
- Marjorie, A.; Raleigh, R.; Jerry, D. Ecosystem model complexity versus physical forcing: Quantification of their relative impact with assimilated Arabian Sea data. Deep Sea Res. Part II Top. Stud. Oceanogr. 2006, 53, 576–600. [Google Scholar]
- Hittawe, M.; Harrou, F.; Sun, Y.; Knio, O. Stacked Transformer Models for Enhanced Wind Speed Prediction in the Red Sea. In Proceedings of the 2024 IEEE 22nd International Conference on Industrial Informatics (INDIN), Beijing, China, 18–20 August 2024; pp. 1–7. [Google Scholar]
- LeCun, Y.; Bengio, Y.; Hinton, G. Deep learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef]
- Afzal, S.; Hittawe, M.M.; Ghani, S.; Jamil, T.; Knio, O.; Hadwiger, M.; Hoteit, I. The state of the art in visual analysis approaches for ocean and atmospheric datasets. Comput. Graph. Forum 2019, 38, 881–907. [Google Scholar] [CrossRef]
- Toni, T.; Welch, D.; Strelkowa, N.; Ipsen, A.; Stumpf, M.P. Approximate Bayesian computation scheme for parameter inference and model selection in dynamical systems. J. R. Soc. Interface 2009, 6, 187–202. [Google Scholar] [CrossRef]
- Gutenkunst, R.; Waterfall, J.; Casey, F.; Brown, K.; Myers, C.; Sethna, J. Universally sloppy parameter sensitivities in systems biology models. PLoS Comput. Biol. 2007, 3, e189. [Google Scholar] [CrossRef]
- Wang, Z.; Chai, F. CoSiNE Manual; Virginia Institute of Marine Science: Gloucester Point, VA, USA, 2017; p. 16. Available online: https://ccrm.vims.edu/schismweb/CoSiNE_manual_ZG_v5.pdf (accessed on 15 June 2025).
- Kishi, M.J.; Kashiwai, M.; Ware, D.M.; Megrey, B.A.; Eslinger, D.L.; Werner, F.E.; Noguchi-Aita, M.; Azumaya, T.; Fujii, M.; Hashimoto, S. NEMURO—A lower trophic level model for the North Pacific marine ecosystem. Ecol. Model. 2007, 202, 12–25. [Google Scholar] [CrossRef]
- Evans, G.T.; Parslow, J.S. A model of annual plankton cycles. Biol. Oceanogr. 1985, 3, 327–347. [Google Scholar]
- Wang, J.; Chen, J.; Chen, D.; Wu, J. LKM-UNet: Large Kernel Vision Mamba Unet for Medical Image Segmentation. In Proceedings of the Medical Image Computing and Computer Assisted Intervention—MICCAI 2024, Marrakesh, Morocco, 6–10 October 2024; pp. 360–370. [Google Scholar]
- Ronneberger, O.; Fischer, P.; Brox, T. U-net: Convolutional Networks for Biomedical Image Segmentation. In Proceedings of the Medical Image Computing and Computer-Assisted Intervention—MICCAI 2015: 18th international conference, Munich, Germany, 5–9 October 2015; pp. 234–241. [Google Scholar]
- Luo, Y.; Weng, E.; Wu, X.; Gao, C.; Zhou, X.; Zhang, L. Parameter identifiability, constraint, and equifinality in data assimilation with ecosystem models. Ecol. Appl. 2009, 19, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Reichstein, M.; Camps-Valls, G.; Stevens, B.; Jung, M.; Denzler, J.; Carvalhais, N.; Prabhat, F. Deep learning and process understanding for data-driven Earth system science. Nature 2019, 566, 195–204. [Google Scholar] [CrossRef]
- Simon, W. Statistical inference for noisy nonlinear ecological dynamic systems. Nature 2010, 466, 1102–1104. [Google Scholar]
- Anderson, T.; Pondaven, P. Non-redfield carbon and nitrogen cycling in the Sargasso Sea: Pelagic imbalances and export flux. Deep Sea Res. Part I Oceanogr. Res. Pap. 2003, 50, 573–591. [Google Scholar] [CrossRef]
- Ji, X.; Liu, G.; Gao, S.; Wang, H. Parameter sensitivity study of the biogeochemical model in the China coastal seas. Acta Oceanol. Sin. 2015, 34, 51–60. [Google Scholar] [CrossRef]
- Hemmings, J.; Challenor, P.; Yool, A. Mechanistic site-based emulation of a global ocean biogeochemical model (MEDUSA 1.0) for parametric analysis and calibration: An application of the Marine Model Optimization Testbed (MarMOT 1.1). Geosci. Model Dev. 2015, 8, 697–731. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of State Variables | Symbol | Tracer Number | Unit |
|---|---|---|---|
| Nitrate | NO3 | 1 | mmol/m3 |
| Silicate | SiO4 | 2 | mmol/m3 |
| Ammonium | NH4 | 3 | mmol/m3 |
| Small Phytoplankton | S1 | 4 | mmol/m3 |
| Diatom | S2 | 5 | mmol/m3 |
| Microzooplankton | Z1 | 6 | mmol/m3 |
| Mesozooplankton | Z2 | 7 | mmol/m3 |
| Detritus Nitrogen | DN | 8 | mmol/m3 |
| Detritus Silicon | DSi | 9 | mmol/m3 |
| Phosphate | PO4 | 10 | mmol/m3 |
| Dissolved Oxygen | DOX | 11 | mmol/m3 |
| Dioxide Carbon | CO2 | 12 | mmol/m3 |
| Alkalinity | ALK | 13 | meq/m−3 |
| Descriptions | Symbols | Values Threshold | Descriptions |
|---|---|---|---|
| Phytoplankton | gmaxs1 | 2.0–4.0 | Maximum growth rate for S1 |
| gmaxs2 | 2.0–4.0 | Maximum growth rate for S2 | |
| pis1 | 0.5–2.0 | Ammonium inhibition factor for S1 | |
| pis2 | 0.1–2.0 | Ammonium inhibition factor for S2 | |
| kno3s1 | 0.1–2.0 | half-saturation constants of NO3 | |
| knh4s1 | 0.1–2.0 | half-saturation constants of NH4 | |
| kpo4s1 | 0.1–2.0 | half-saturation constants of PO4 | |
| kno3s2 | 0.1–2.0 | half-saturation constants of NO3 | |
| knh4s2 | 0.1–2.0 | half-saturation constants of NH4 | |
| kpo4s2 | 0.1–2.0 | half-saturation constants of PO4 | |
| ksio4s2 | 0.1–2.0 | half-saturation constants of SiO4 | |
| ak1 | 0.0–1.0 | background light extinction | |
| ak2 | 0.0–1.0 | light extinction coefficient for phytoplankton | |
| gammas1 | 0.0–1.0 | Mortality rates for S1 | |
| gammas2 | 0.0–1.0 | Mortality rates for S2 | |
| Zooplankton | beta1 | 0.0–1.0 | Maximum grazing rates for Z1 |
| beta2 | 0.0–1.0 | Maximum grazing rates for Z2 | |
| gamma1 | 0.0–1.0 | the assimilation rates for Z1 | |
| gamma2 | 0.0–1.0 | the assimilation rates for Z2 | |
| gammaz | 0.0–0.1 | Mortality rate for Z1 and Z2 | |
| kex1 | 0.0–0.5 | They are the excretion rates for Z1 | |
| kex2 | 0.0–0.5 | They are the excretion rates for Z2 | |
| Others | wss2 | 0.0–10.0 | Settling velocities S1 |
| wsdn | 0.0–20.0 | Settling velocities DN | |
| wsdsi | 0.0–40.0 | Settling velocities DSi |
| Sequence Number | Parameter Names | Parameter Values | |||||
|---|---|---|---|---|---|---|---|
| Group 1 | Group 2 | Group 3 | |||||
| ME | AI | ME | AI | ME | AI | ||
| 1 | gmaxs1 | 2.453 | 2.521 | 2.207 | 2.155 | 2.515 | 2.521 |
| 2 | gmaxs2 | 2.765 | 2.920 | 2.827 | 2.973 | 2.657 | 2.920 |
| 3 | pis1 | 1.500 | 1.341 | 1.436 | 1.360 | 1.017 | 1.341 |
| 4 | pis2 | 0.374 | 0.108 | 0.156 | 0.269 | 0.105 | 0.108 |
| 5 | kno3s1 | 1.017 | 0.951 | 0.574 | 0.563 | 1.096 | 0.951 |
| 6 | knh4s1 | 0.371 | 0.374 | 0.198 | 0.188 | 0.435 | 0.374 |
| 7 | kpo4s1 | 0.231 | 0.224 | 0.371 | 0.351 | 0.263 | 0.224 |
| 8 | kno3s2 | 1.777 | 1.760 | 1.734 | 1.704 | 1.703 | 1.760 |
| 9 | knh4s2 | 0.243 | 0.231 | 0.418 | 0.399 | 0.299 | 0.231 |
| 10 | kpo4s2 | 0.203 | 0.219 | 0.368 | 0.4357 | 0.389 | 0.219 |
| 11 | ksio4s2 | 4.823 | 4.825 | 4.969 | 4.996 | 4.843 | 4.825 |
| 12 | ak1 | 0.627 | 0.619 | 0.669 | 0.652 | 0.655 | 0.619 |
| 13 | ak2 | 0.035 | 0.034 | 0.035 | 0.035 | 0.033 | 0.034 |
| 14 | gammas1 | 0.039 | 0.039 | 0.035 | 0.035 | 0.038 | 0.039 |
| 15 | gammas2 | 0.036 | 0.037 | 0.028 | 0.029 | 0.834 | 0.846 |
| 16 | beta1 | 0.851 | 0.846 | 0.841 | 0.835 | 0.269 | 0.256 |
| 17 | beta2 | 0.256 | 0.256 | 0.275 | 0.274 | 0.269 | 0.256 |
| 18 | gamma1 | 0.816 | 0.810 | 0.804 | 0.828 | 0.829 | 0.810 |
| 19 | gamma2 | 0.492 | 0.496 | 0.474 | 0.475 | 0.484 | 0.496 |
| 20 | gammaz | 0.065 | 0.067 | 0.055 | 0.050 | 0.070 | 0.067 |
| 21 | kex1 | 0.099 | 0.098 | 0.061 | 0.061 | 0.098 | 0.098 |
| 22 | kex2 | 0.092 | 0.090 | 0.099 | 0.096 | 0.091 | 0.090 |
| 23 | wss2 | 0.845 | 0.822 | 0.946 | 0.965 | 0.875 | 0.822 |
| 24 | wsdn | 0.971 | 0.966 | 0.921 | 0.911 | 0.965 | 0.966 |
| 25 | wsdsi | 0.923 | 0.943 | 0.967 | 0.987 | 0.890 | 0.943 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, A.; Zhao, K.; Fang, W.; Shu, C.; Zhou, R.; Li, Q.; Huang, X.; Lei, X.; Xu, M.; Jiang, H.; et al. A Deep Learning Framework for Parameter Estimation in 1D Marine Ecosystem Model. J. Mar. Sci. Eng. 2025, 13, 1228. https://doi.org/10.3390/jmse13071228
Li A, Zhao K, Fang W, Shu C, Zhou R, Li Q, Huang X, Lei X, Xu M, Jiang H, et al. A Deep Learning Framework for Parameter Estimation in 1D Marine Ecosystem Model. Journal of Marine Science and Engineering. 2025; 13(7):1228. https://doi.org/10.3390/jmse13071228
Chicago/Turabian StyleLi, Ao, Kewei Zhao, Weiwei Fang, Chan Shu, Runjie Zhou, Qiuyi Li, Xiaolong Huang, Xiaohong Lei, Menghan Xu, Haoyu Jiang, and et al. 2025. "A Deep Learning Framework for Parameter Estimation in 1D Marine Ecosystem Model" Journal of Marine Science and Engineering 13, no. 7: 1228. https://doi.org/10.3390/jmse13071228
APA StyleLi, A., Zhao, K., Fang, W., Shu, C., Zhou, R., Li, Q., Huang, X., Lei, X., Xu, M., Jiang, H., & Mu, L. (2025). A Deep Learning Framework for Parameter Estimation in 1D Marine Ecosystem Model. Journal of Marine Science and Engineering, 13(7), 1228. https://doi.org/10.3390/jmse13071228