

Composition and Distribution of Microeukaryotic Communities in the Surface Sediments of Five Geographic Regions of Bohai Sea Based on 18S rDNA Amplicon Sequencing


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
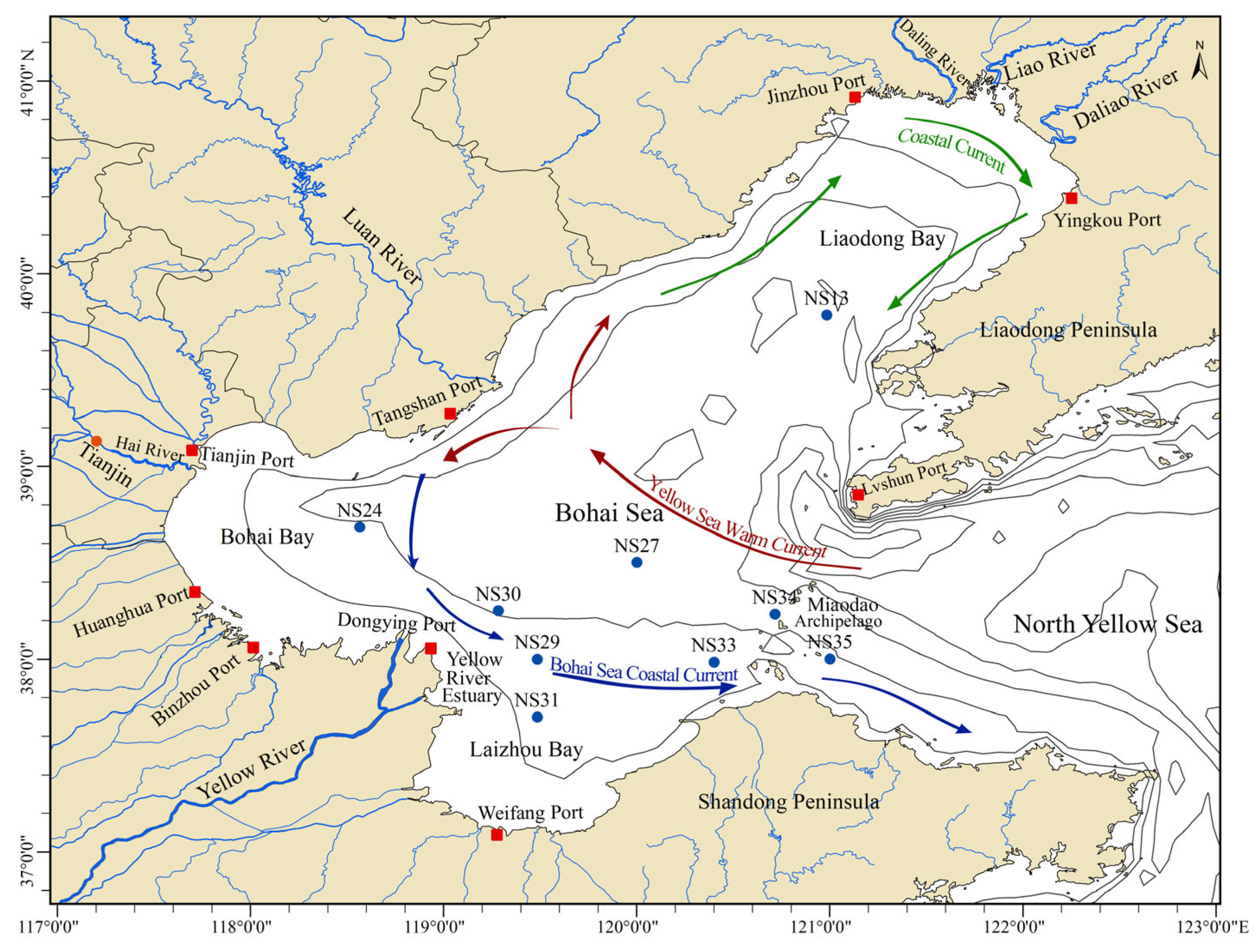
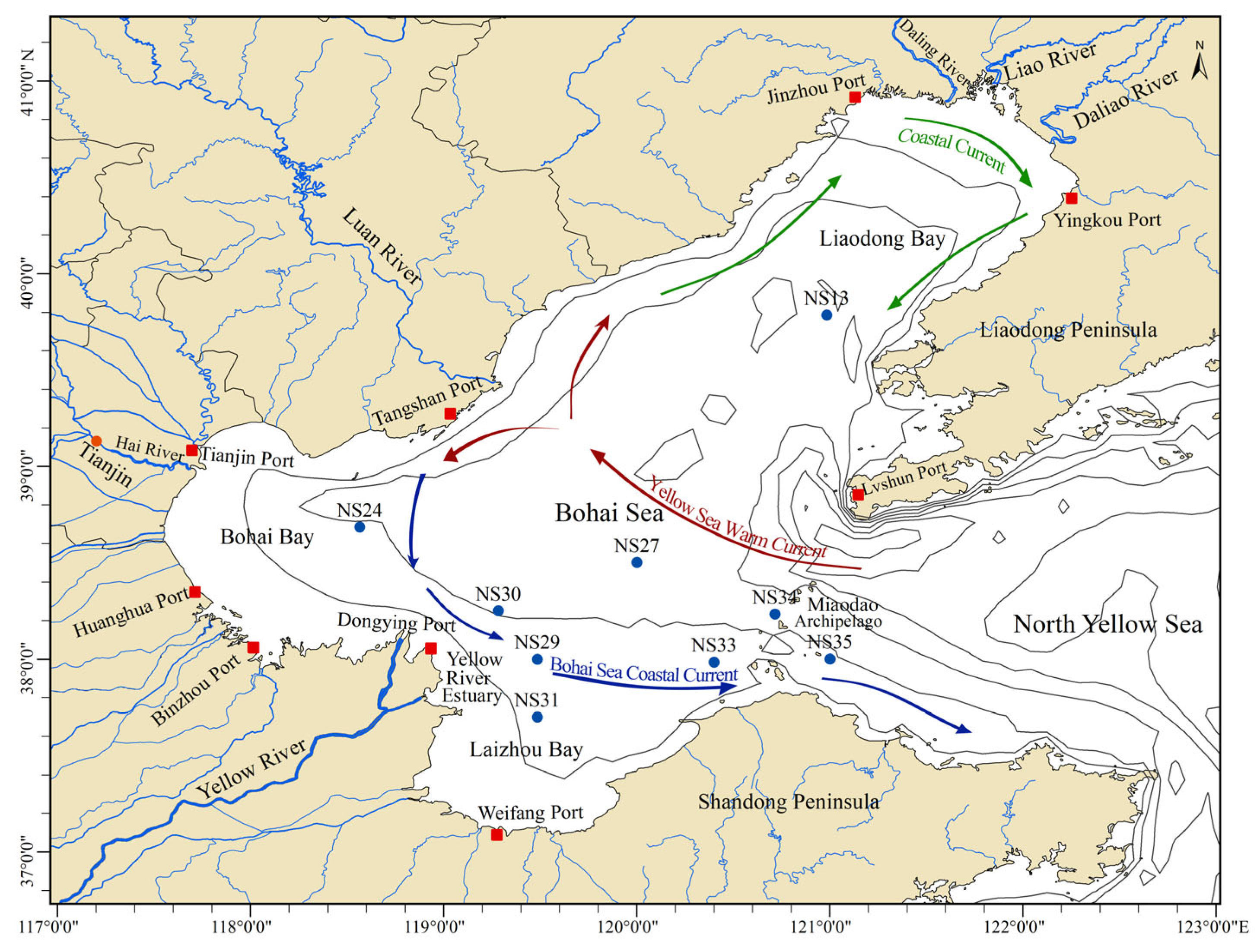
2.1. Description of Sampling Sites and Sample Collection
2.2. DNA Extraction and Amplicon Sequencing
2.3. Paired-End Reads Assembly and Species Annotation
2.4. Cluster and Community Composition Analysis
2.5. Environmental Correlation Analysis
3. Results
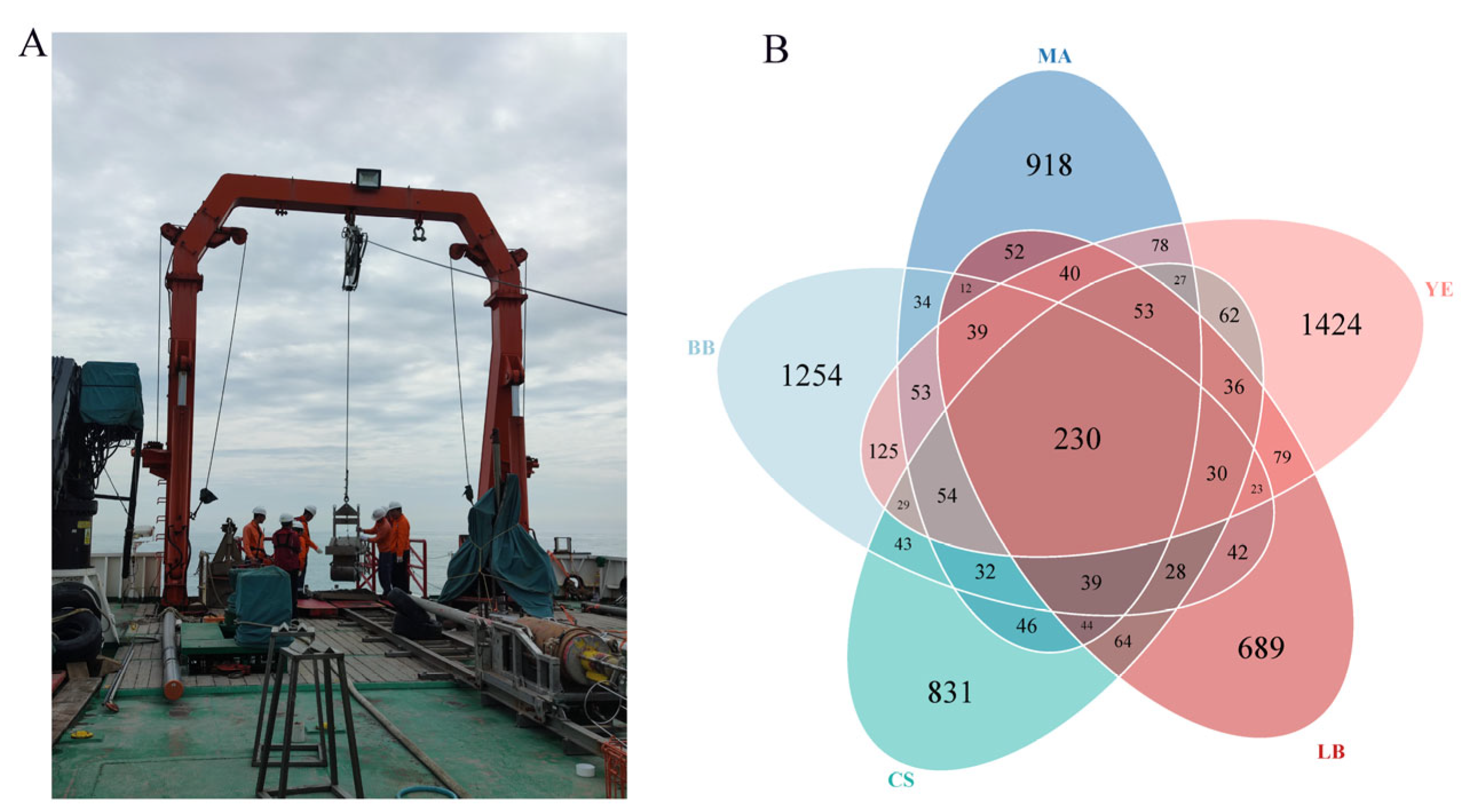
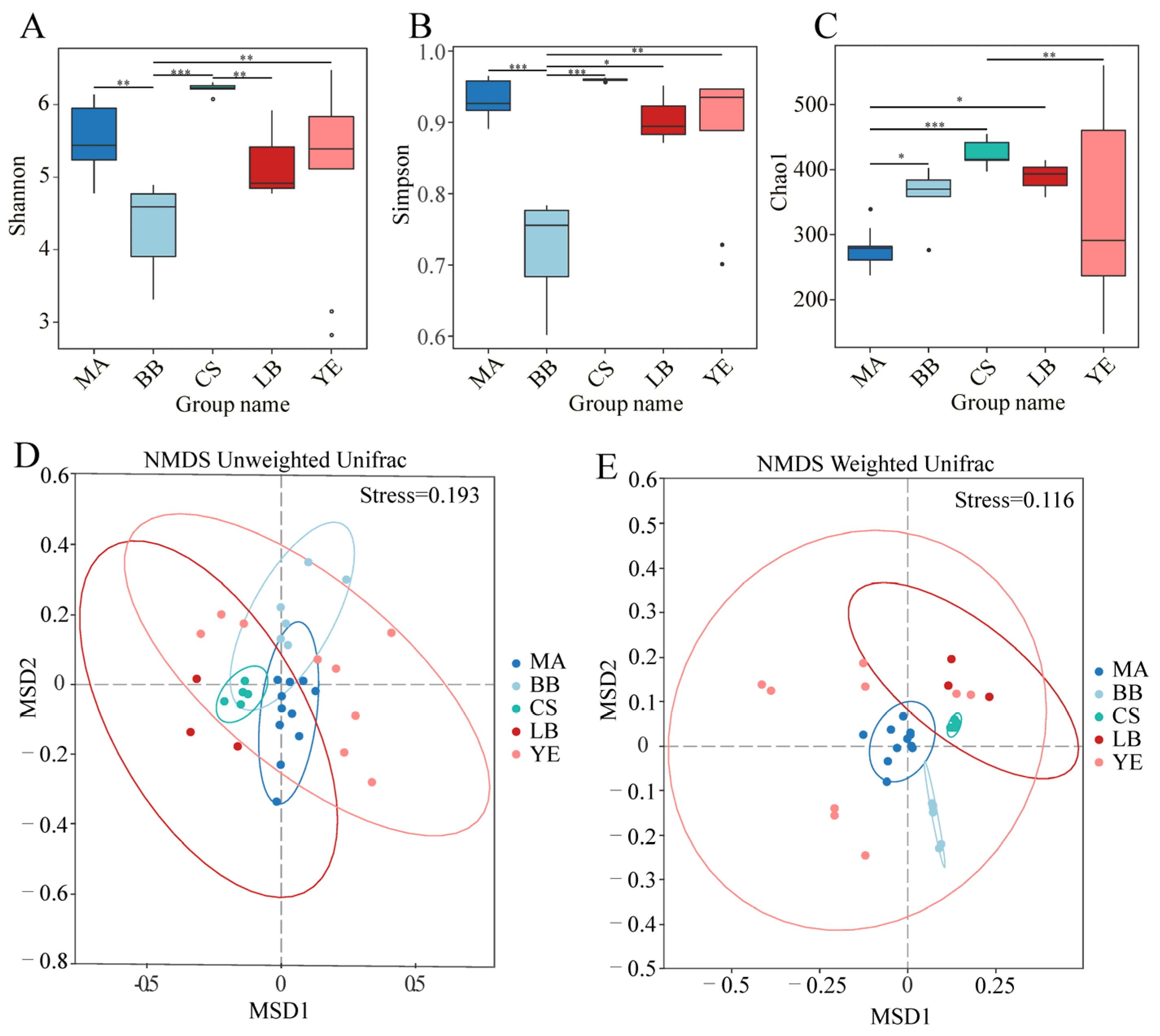
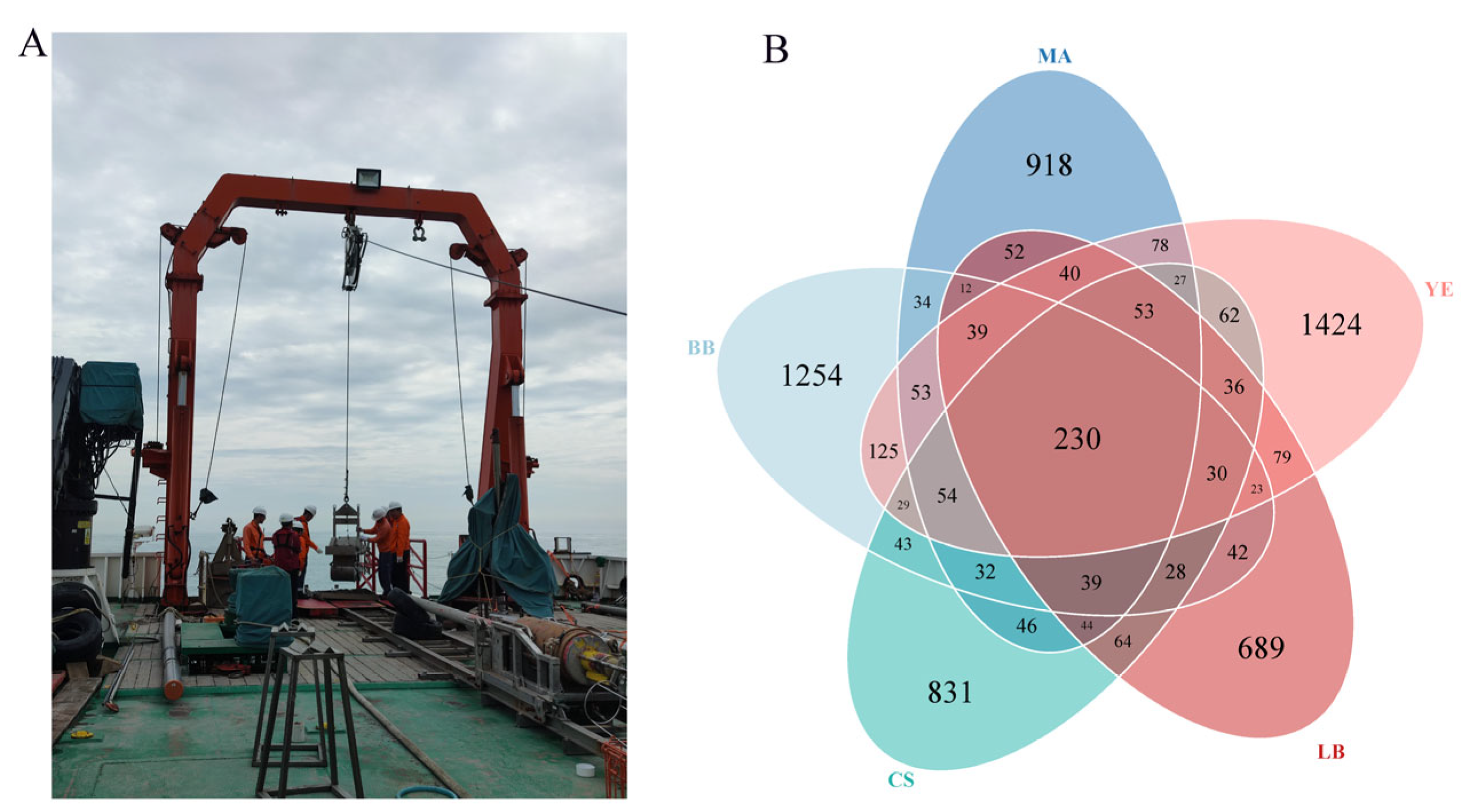
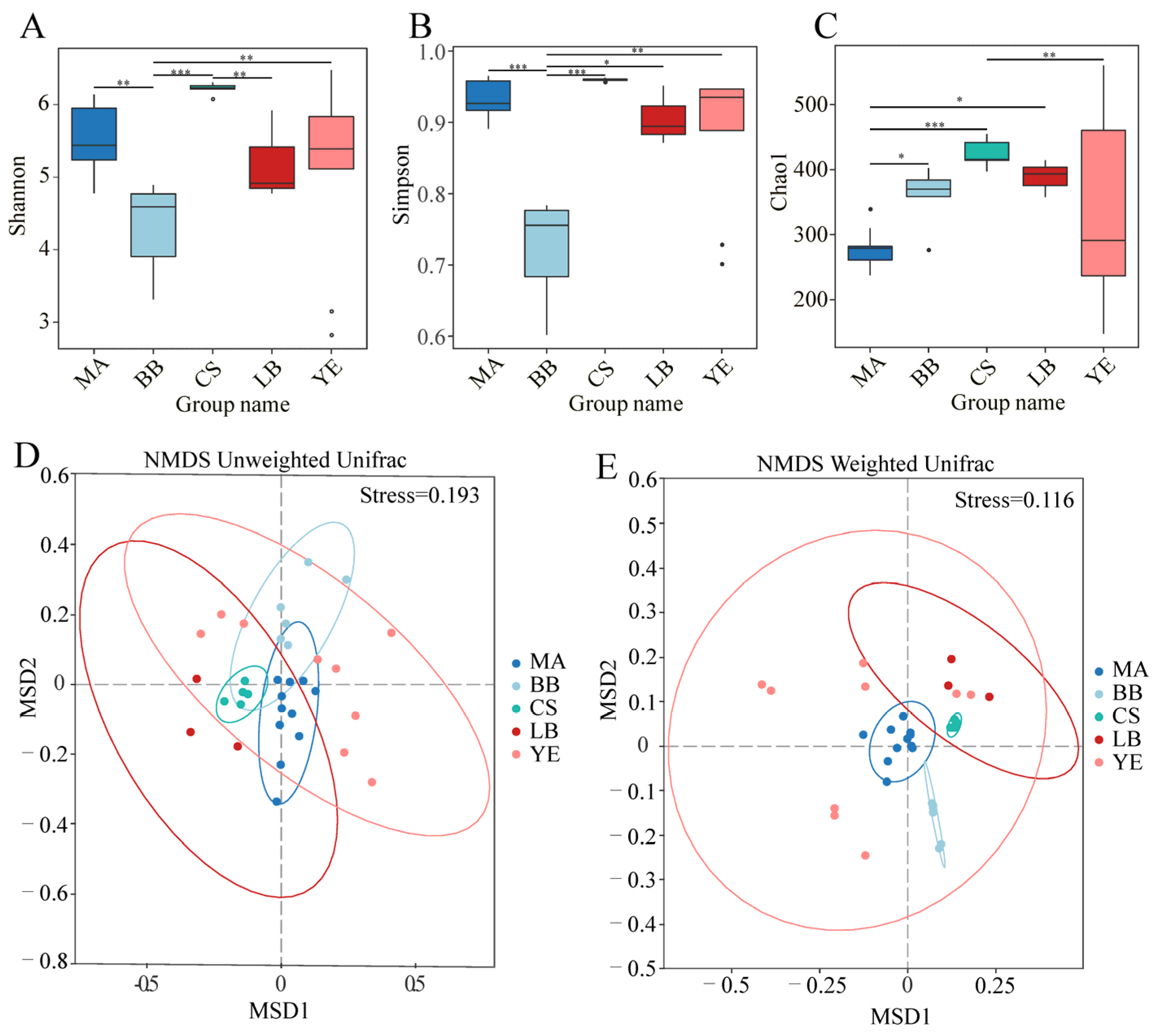
3.1. Microeukaryotic Communities of Five Typical Regions in Bohai Sea
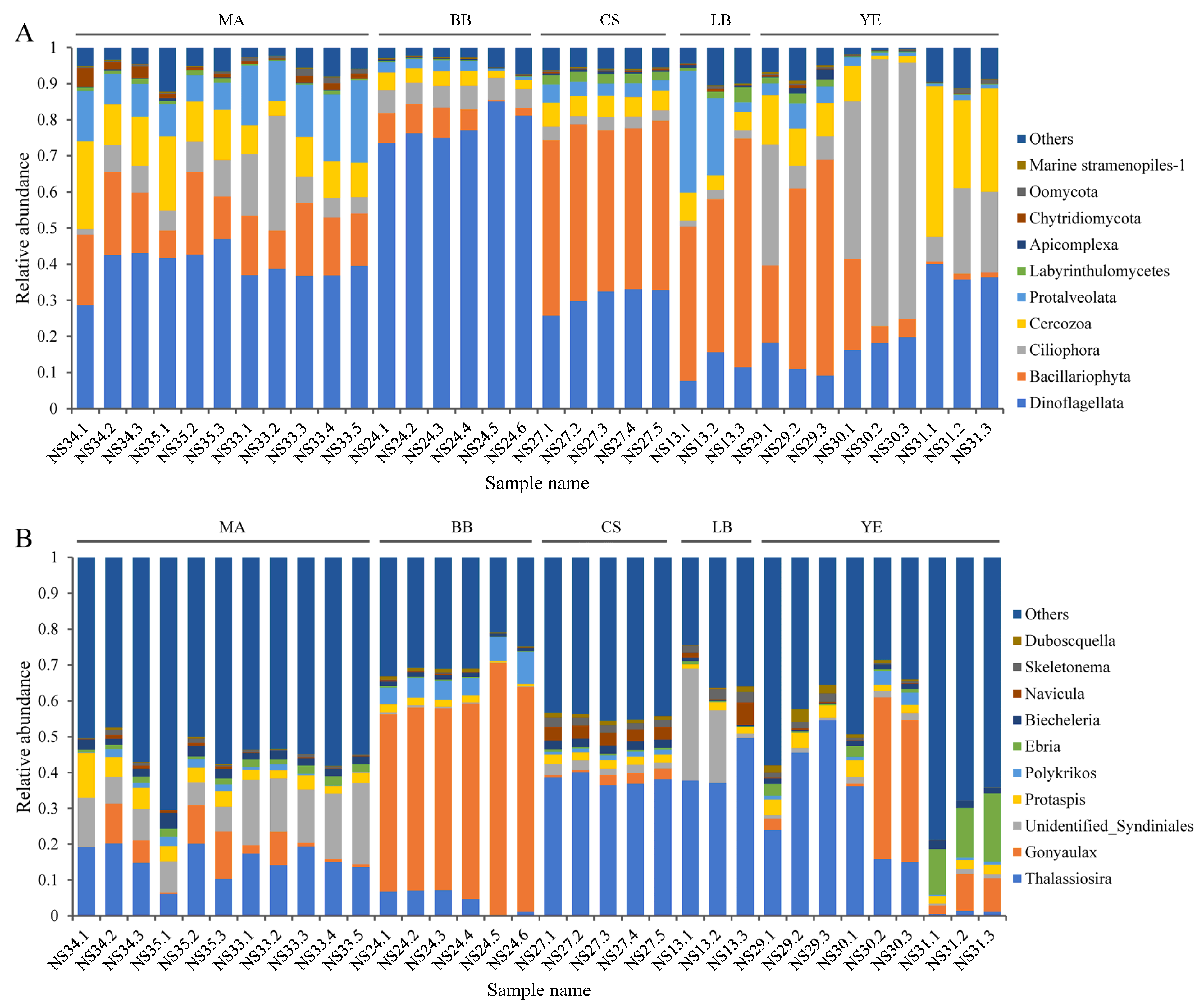
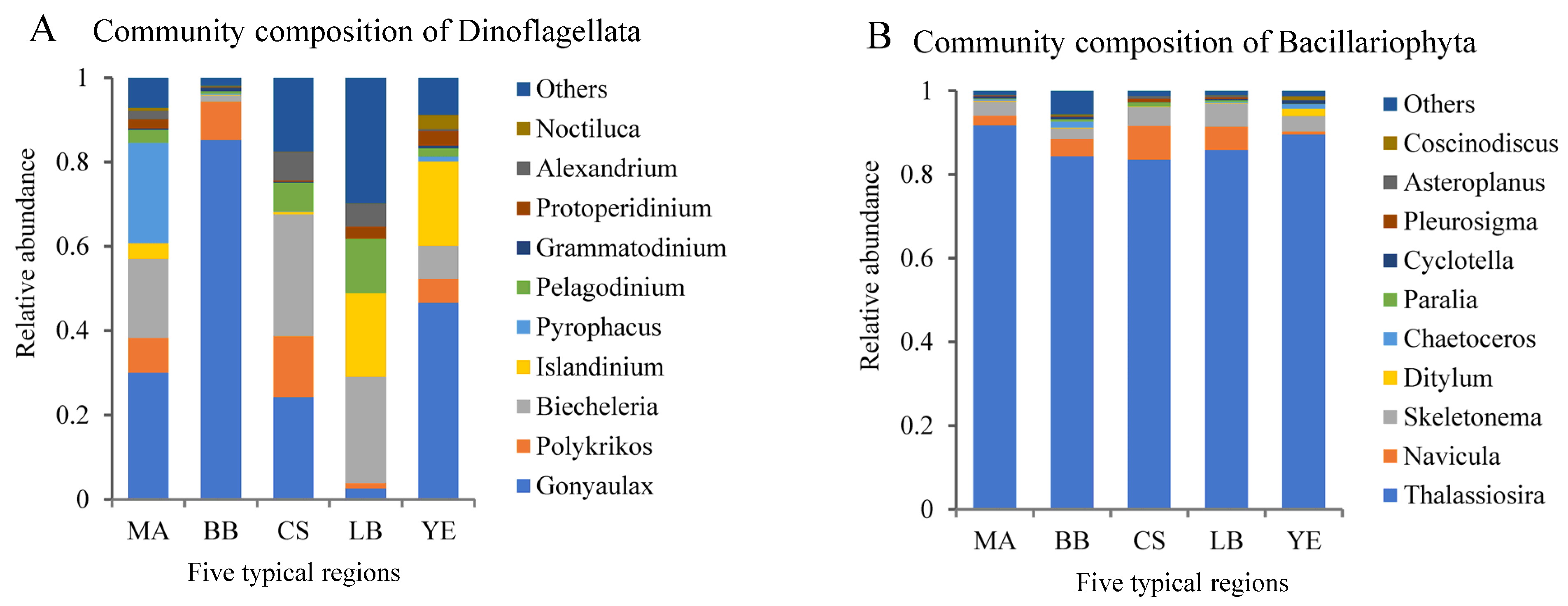
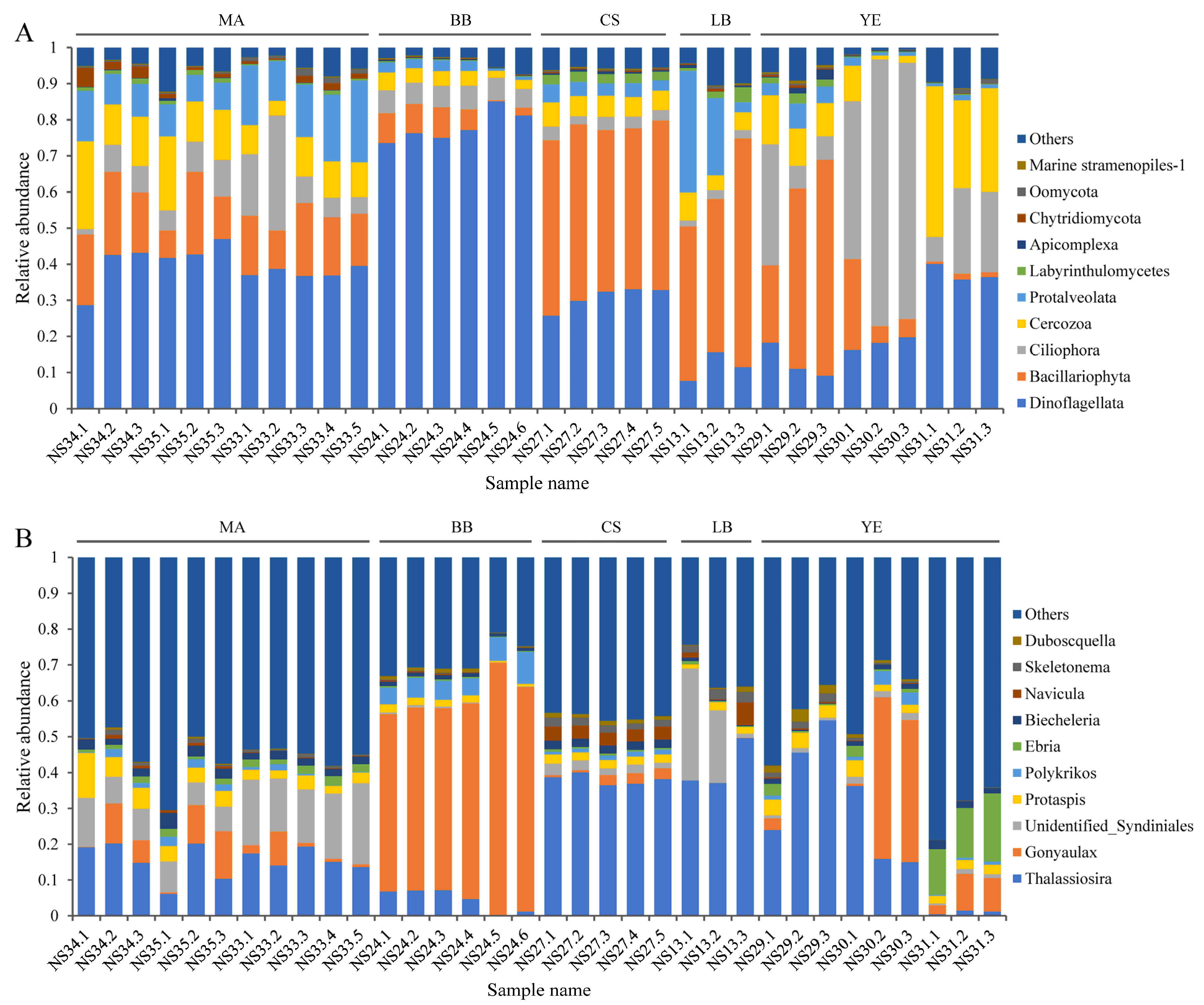
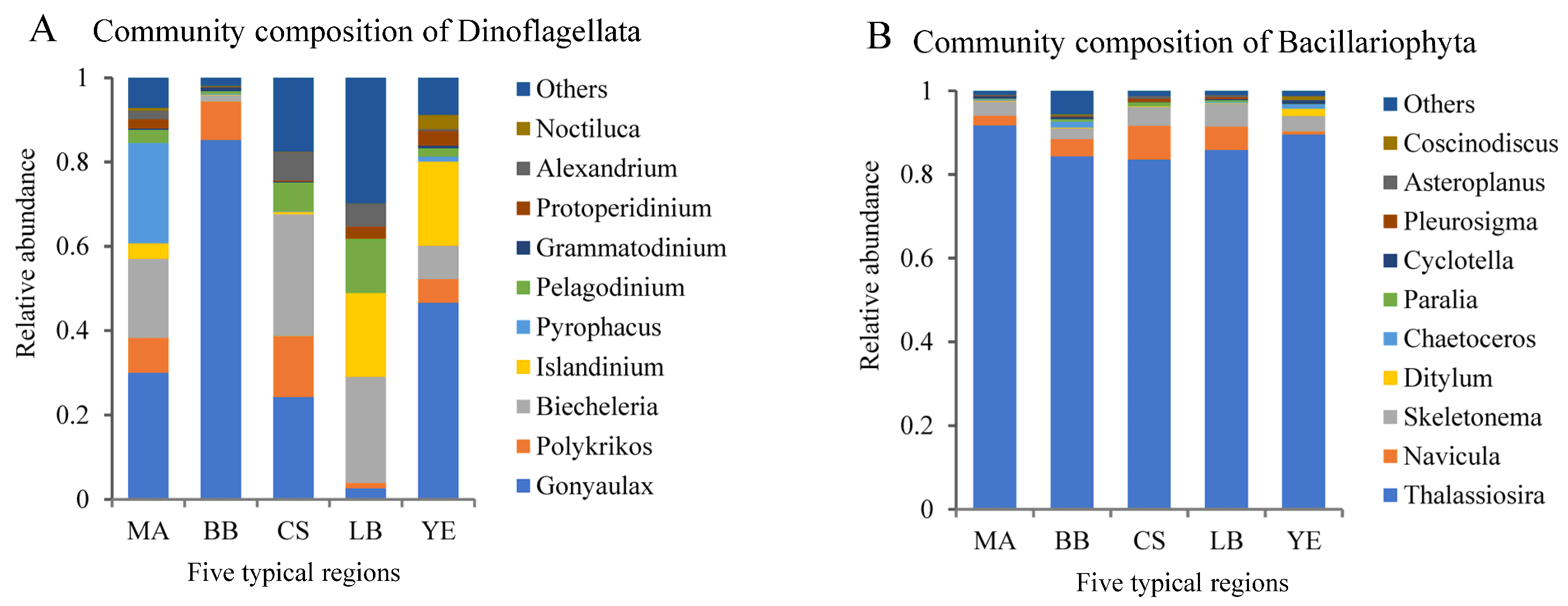
3.2. Composition and Distribution of Dominant Microeukaryotic Taxa
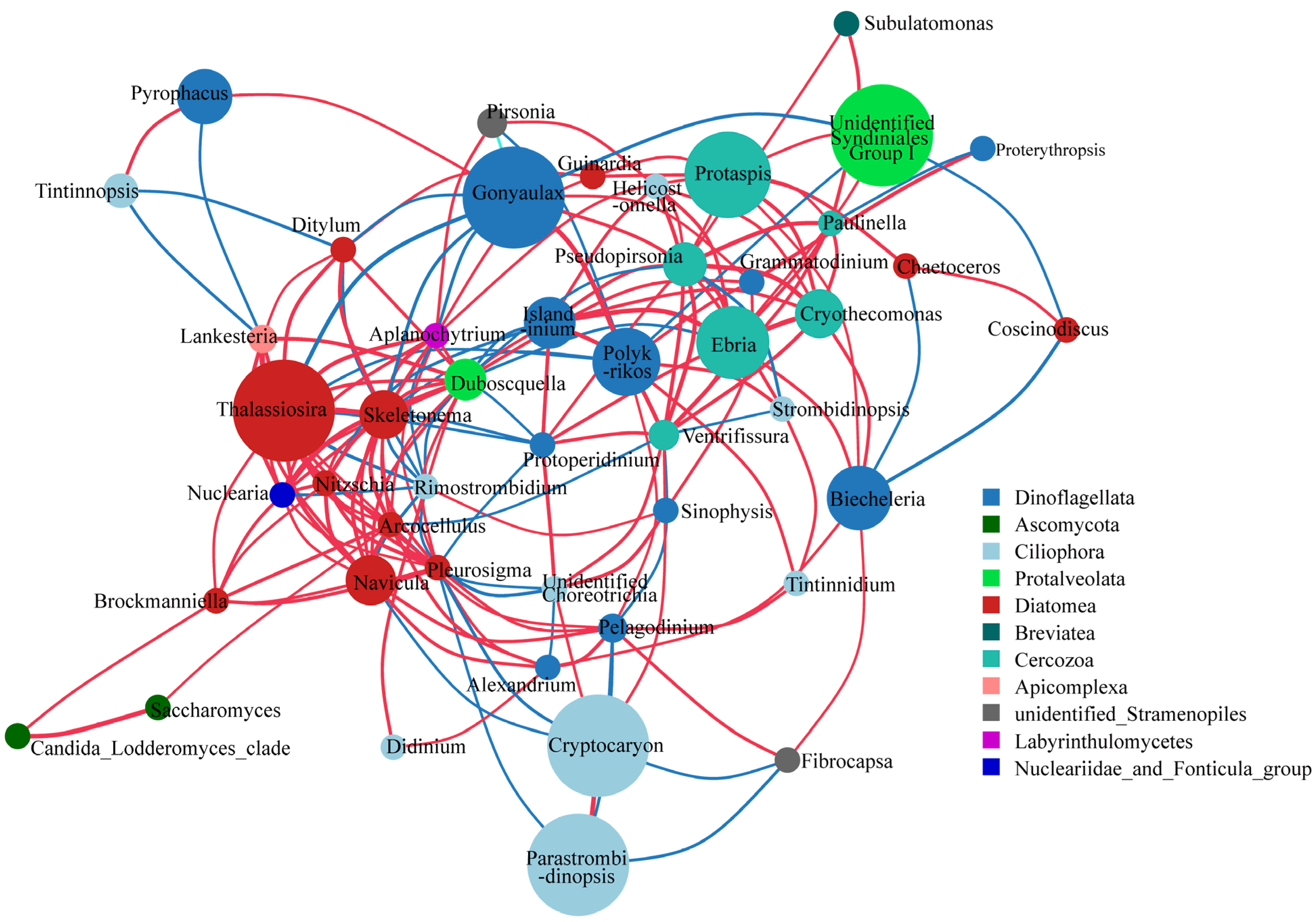
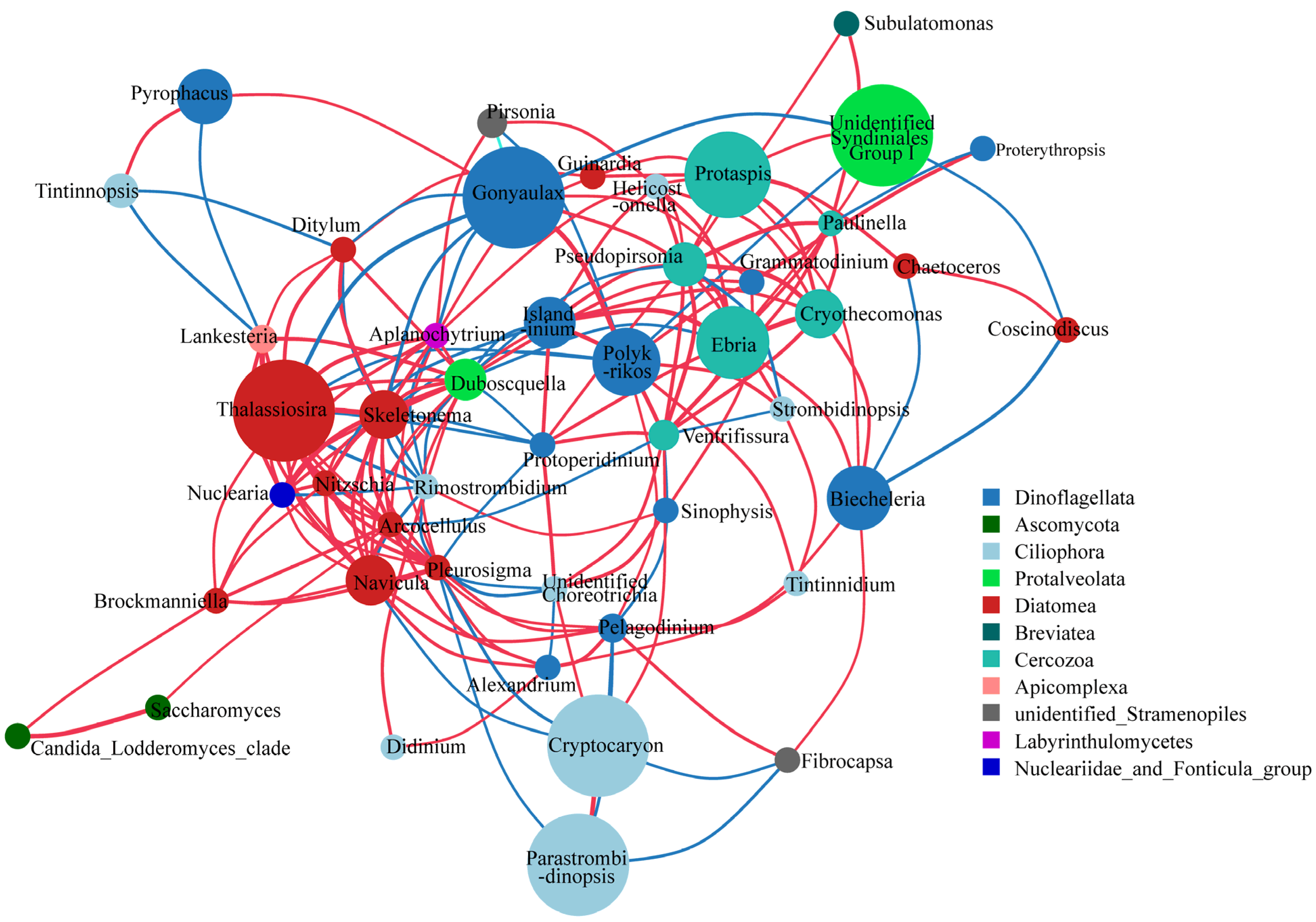
3.3. Species Relationship Networks of Microeukaryotic Communities
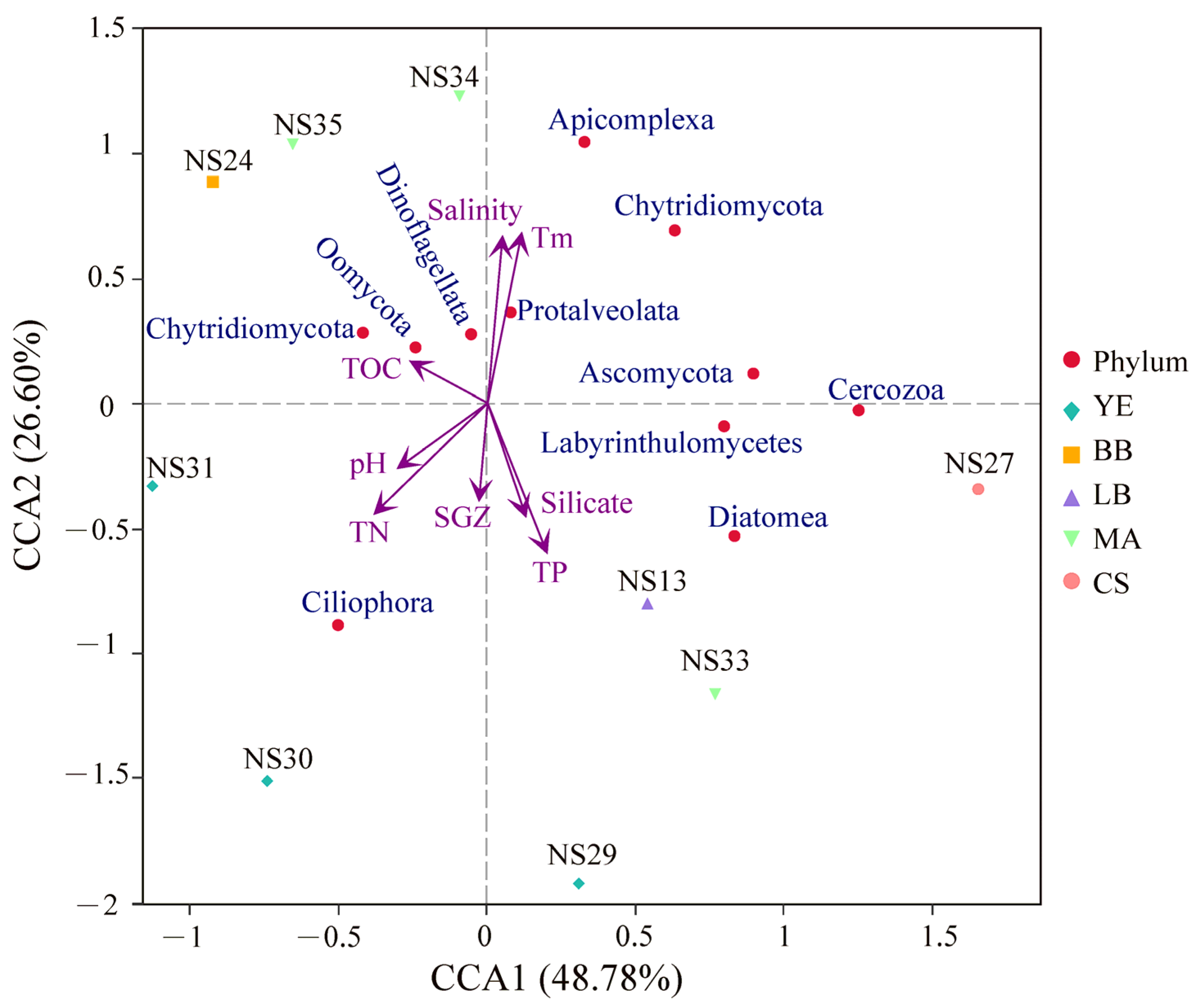
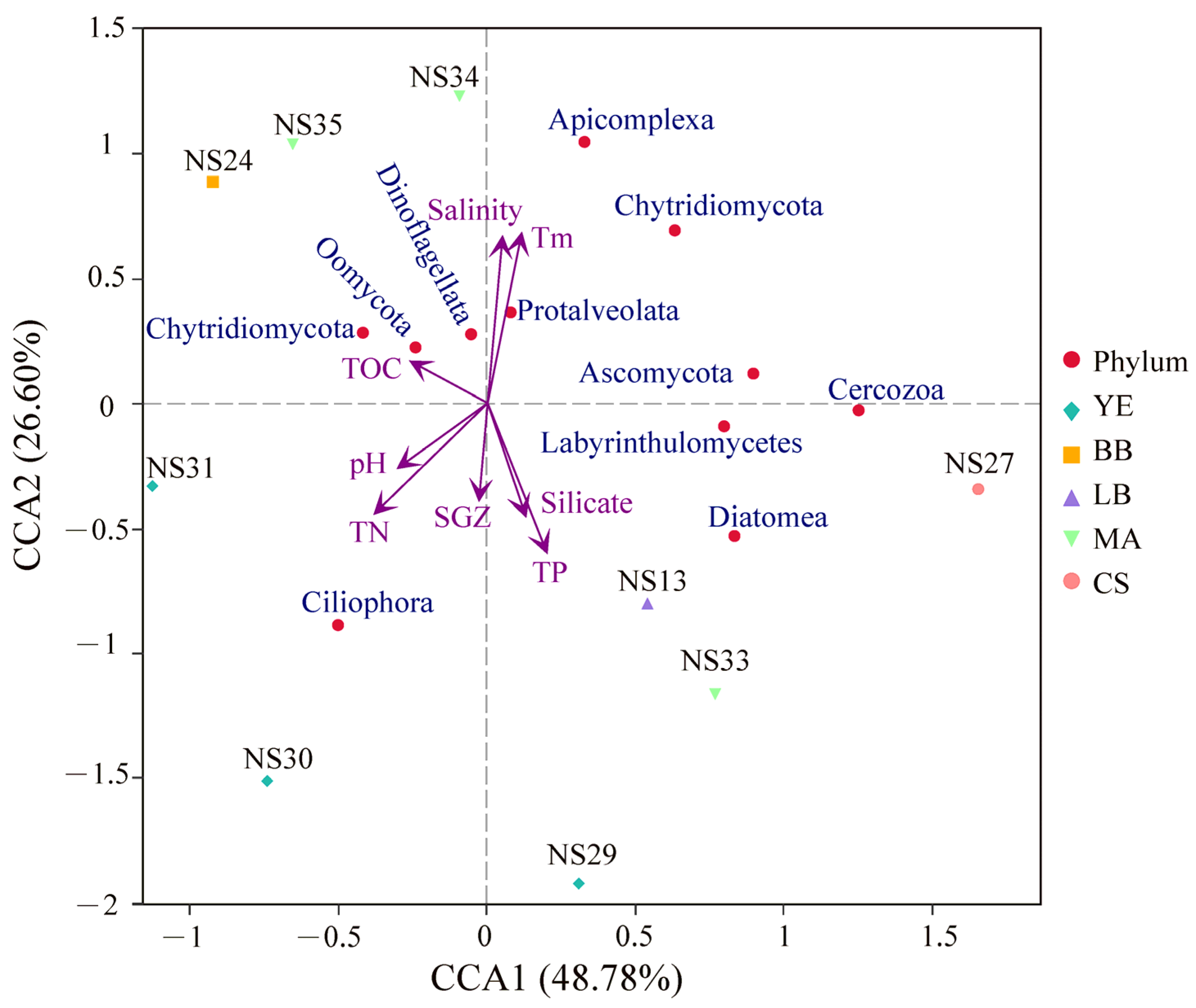
3.4. Correlation Analysis between Microeukaryotic Communities and Environmental Factors
4. Discussion
4.1. Microeukaryotic Alpha Diversity Affected by Environmental Variables
4.2. Dominant Microeukaryotic Assemblages Alter Spatially with Environmental Variation
4.3. Regime Shifts in Diatom–Dinoflagellate Composition Reflected Eutrophication
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ni, D.; Zhang, Z.; Liu, X. Benthic ecological quality assessment of the Bohai Sea, China using marine biotic indices. Mar. Pollut. Bull. 2019, 142, 457–464. [Google Scholar] [CrossRef]
- Liu, D.; Liu, L.; Di, B.; Wang, Y.; Wang, Y. Paleoenvironmental analyses of surface sediments from the Bohai Sea, China, using diatoms and silicoflagellates. Mar. Micropaleontol. 2015, 114, 46–54. [Google Scholar] [CrossRef]
- Wu, G.; Li, J.; Luo, W. Spatial distribution, source apportionment, and assessment of marine water quality parameters in the Bohai Sea, China. Mar. Pollut. Bull. 2023, 195, 115526. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, G.; Xu, Y.; Li, X.; Wang, J.; Sui, J. Community characteristics of macrozoobenthos in the Miaodao Archipelago in summer. Mar. Sci. 2023, 47, 96–110. [Google Scholar]
- Liu, B.; Wu, X.; Liu, X.; Gong, M. Assessment of ecological stress caused by maritime vessels based on a comprehensive model using AIS data: Case study of the Bohai Sea, China. Ecol. Indic. 2021, 126, 107592. [Google Scholar] [CrossRef]
- Liu, J.; Feng, Y.; Zhang, Y.; Liang, N.; Wu, H.; Liu, F. Allometric releases of nitrogen and phosphorus from sediments mediated by bacteria determines water eutrophication in coastal river basins of Bohai Bay. Ecotoxicol. Environ. Saf. 2022, 235, 113426. [Google Scholar] [CrossRef]
- Wei, Y.; Cui, H.; Hu, Q.; Bai, Y.; Qu, K.; Sun, J.; Cui, Z. Eutrophication status assessment in the Laizhou Bay, Bohai Sea: Further evidence for the ecosystem degradation. Mar. Pollut. Bull. 2022, 181, 113867. [Google Scholar] [CrossRef]
- Gu, X.; Liu, S.; Li, Y.; Ouyang, W.; He, M.; Liu, X.; Lin, C. A review of sources, status, and risks of microplastics in the largest semi-enclosed sea of China, the Bohai Sea. Chemosphere 2022, 306, 135564. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fu, R.; Li, H.; Zhang, Y.; Lu, M.; Xiao, K.; Zhang, X.; Zheng, C.; Xiong, Y. Heavy metal contamination in surface sediments: A comprehensive, large-scale evaluation for the Bohai Sea, China. Environ. Pollut. 2020, 260, 113986. [Google Scholar] [CrossRef]
- Li, X.Y.; Yu, R.C.; Richardson, A.J.; Sun, C.; Eriksen, R.; Kong, F.Z.; Zhou, Z.X.; Geng, H.X.; Zhang, Q.C.; Zhou, M.J. Marked shifts of harmful algal blooms in the Bohai Sea linked with combined impacts of environmental changes. Harmful Algae 2023, 121, 102370. [Google Scholar] [CrossRef]
- Kang, B.; Pecl, G.T.; Lin, L.; Sun, P.; Zhang, P.; Li, Y.; Zhao, L.; Peng, X.; Yan, Y.; Shen, C.; et al. Climate change impacts on China’s marine ecosystems. Rev. Fish Biol. Fisher. 2021, 31, 599–629. [Google Scholar] [CrossRef]
- Wei, Y.; Ding, D.; Gu, T.; Xu, Y.; Sun, X.; Qu, K.; Sun, J.; Cui, Z. Ocean acidification and warming significantly affect coastal eutrophication and organic pollution: A case study in the Bohai Sea. Mar. Pollut. Bull. 2023, 186, 114380. [Google Scholar] [CrossRef]
- Jiang, S.; Fan, W.; Chen, L.; Chen, J.; Li, B. Spatio-temporal distribution of macrobenthos and benthic ecological health status in the Bohai Sea and the northern Yellow Sea, China. Mar. Pollut. Bull. 2023, 196, 115671. [Google Scholar] [CrossRef]
- Xu, Z.; Shi, H.; Li, N.; Zhao, R.; Liu, X. Macrobenthic community structure and its relationship with environmental factors in the southern waters of the Miaodao Archipelago. Res. Environ. Sci. 2015, 28, 704–712. [Google Scholar]
- Shi, Y.; Zhang, G.; Zhang, G.; Wen, Y.; Guo, Y.; Peng, L.; Xu, W.; Sun, J. Species and functional diversity of marine macrobenthic community and benthic habitat quality assessment in semi-enclosed waters upon recovering from eutrophication, Bohai Bay, China. Mar. Pollut. Bull. 2022, 181, 113918. [Google Scholar] [CrossRef] [PubMed]
- Popovic, A.; Parkinson, J. Characterization of eukaryotic microbiome using 18S amplicon sequencing. Methods Mol. Biol. 2018, 1849, 29–48. [Google Scholar]
- Xu, H.; Zhang, S.; Ma, G.; Zhang, Y.; Li, Y.; Pei, H. 18S rRNA gene sequencing reveals significant influence of anthropogenic effects on microeukaryote diversity and composition along a river-to-estuary gradient ecosystem. Sci. Total Environ. 2020, 705, 135910. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, Z.; Cheng, F.; He, L.; Cao, X.; Song, X. Molecular diversity and ecological characteristics of the eukaryotic phytoplankton community in the coastal waters of the Bohai Sea, China. Harmful Algae 2017, 61, 13–22. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; Wong, J.; Heiner, C.; Oh, S.; Theriot, C.M.; Gulati, A.S.; McGill, S.K.; Dougherty, M.K. High-throughput amplicon sequencing of the full-length 16S rRNA gene with single-nucleotide resolution. Nucleic Acids Res. 2019, 47, e103. [Google Scholar] [CrossRef] [PubMed]
- Amir, A.; McDonald, D.; Navas-Molina, J.A.; Kopylova, E.; Morton, J.T.; Zech Xu, Z.; Kightley, E.P.; Thompson, L.R.; Hyde, E.R.; Gonzalez, A.; et al. Deblur rapidly resolves single-nucleotide community sequence patterns. mSystems 2017, 2, e00191-16. [Google Scholar] [CrossRef]
- Taft, D.H.; Akre, S.; Madrid, N.; Knoesen, A.; Mills, D.A.; Lewis, Z.T. Resident microbes of lactation rooms and daycares. PeerJ 2019, 7, e8168. [Google Scholar] [CrossRef]
- GB 17378.4-2007; Specification for Marine Monitoring of China—Part 5: Sediment Analysis. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China: Beijing, China, 2007.
- GB/T 9742-2008; Chemical Reagent—General Method for the Determination of Silicate. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China: Beijing, China, 2008.
- GB/T12736.8-2007; Oceanographic Survey—Part 8: Marine Geology and Geophysics Survey. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China: Beijing, China, 2007.
- Kanso, E.A.; Lopes, R.M.; Strickler, J.R.; Dabiri, J.O.; Costello, J.H. Teamwork in the viscous oceanic microscale. Proc. Natl. Acad. Sci. USA 2021, 118, e2018193118. [Google Scholar] [CrossRef]
- Riera, R.; Becerro, M.A.; Stuart-Smith, R.D.; Delgado, J.D.; Edgar, G.J. Out of sight, out of mind: Threats to the marine biodiversity of the Canary Islands (NE Atlantic Ocean). Mar. Pollut. Bull. 2014, 86, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Jost, L. Partitioning diversity into independent alpha and beta components. Ecology 2007, 88, 2427–2439. [Google Scholar] [CrossRef]
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yang, T.; Shan, X.; Jin, X.; Teng, G.; Wei, C. Stable isotope analysis of food web structure and the contribution of carbon sources in the sea adjacent to the Miaodao Archipelago (China). Fishes 2022, 7, 32. [Google Scholar] [CrossRef]
- Worm, B.; Barbier, E.B.; Beaumont, N.; Duffy, J.E.; Folke, C.; Halpern, B.S.; Jackson, J.B.; Lotze, H.K.; Micheli, F.; Palumbi, S.R.; et al. Impacts of biodiversity loss on ocean ecosystem services. Science 2006, 314, 787–790. [Google Scholar] [CrossRef]
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D’Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; et al. A global map of human impact on marine ecosystems. Science 2008, 319, 948–952. [Google Scholar] [CrossRef]
- Hu, L.; Zou, A.; Zhang, P.; Liu, H.; Mei, X.; Hu, Z.; Lei, Y.; Wang, X.; Wang, X.; Chen, Q.; et al. Surface sediment Cu characteristics and the possible influencing factors in the northern four-island area of Penglai district, Yantai city, China. Water Air Soil. Pollut. 2023, 234, 177. [Google Scholar] [CrossRef]
- Shan, X.; Sun, P.; Jin, X.; Li, X.; Dai, F. Long-term changes in fish assemblage structure in the Yellow River Estuary ecosystem, China. Mar. Coast. Fisher. 2013, 5, 65–78. [Google Scholar] [CrossRef]
- O’Brien, P.A.; Tan, S.; Yang, C.; Frade, P.R.; Andreakis, N.; Smith, H.A.; Miller, D.J.; Webster, N.S.; Zhang, G.; Bourne, D.G. Diverse coral reef invertebrates exhibit patterns of phylosymbiosis. Isme J. 2020, 14, 2211–2222. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, X.; Huang, L.; Bao, F.; Xiong, S.; Wang, K.; Zhang, D. Microeukaryotic biogeography in the typical subtropical coastal waters with multiple environmental gradients. Sci. Total Environ. 2018, 635, 618–628. [Google Scholar] [CrossRef]
- Dimitriadis, C.; Evagelopoulos, A.; Koutsoubas, D. Functional diversity and redundancy of soft bottom communities in brackish waters areas: Local vs regional effects. J. Exp. Mar. Biol. Ecol. 2012, 426, 53–59. [Google Scholar] [CrossRef]
- Xin, M.; Wang, B.; Xie, L.; Sun, X.; Wei, Q.; Liang, S.; Chen, K. Long-term changes in nutrient regimes and their ecological effects in the Bohai Sea, China. Mar. Pollut. Bull. 2019, 146, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Guo, Y.; Liu, H.; Zhang, G.; Zhang, X.; Thangaraj, S.; Sun, J. Water quality shifts the dominant phytoplankton group from diatoms to dinoflagellates in the coastal ecosystem of the Bohai Bay. Mar. Pollut. Bull. 2022, 183, 114078. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Achterberg, E.P.; Li, K.; Zhang, J.; Xin, M.; Wang, X. Governance pathway for coastal eutrophication based on regime shifts in diatom-dinoflagellate composition of the Bohai and Baltic Seas. Water Res. 2023, 250, 121042. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, L.; Gu, H.; Li, S.; Li, S. Species diversity and eco-geographic distribution of dinoflagellate cysts in the Bohai Sea, China. Chin. Bull. Bot. 2012, 47, 125–132. [Google Scholar]
- Sathish Kumar, P.; Kumaraswami, M.; Ezhilarasan, P.; Durga Rao, G.; Sivasankar, R.; Ranga Rao, V.; Ramu, K. Blooming of Gonyaulax polygramma along the southeastern Arabian Sea: Influence of upwelling dynamics and anthropogenic activities. Mar. Pollut. Bull. 2020, 151, 110817. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.B.; Kurian, S.; Vijayan, N.T.; Gauns, M.; Khichi, R.; Pratihary, A.K.; Borker, S.G.; Shenoy, D.M. Recurrence of Gonyaulax polygramma bloom in the southeastern Arabian Sea. Environ. Monit. Assess. 2023, 195, 635. [Google Scholar] [CrossRef]
- Bharathi, M.D.; Muthukumar, C.; Sathishkumar, R.S.; Ramu, K.; Murthy, M.V.R. First report on the occurrence of Gonyaulax polygramma bloom during the onset of Noctiluca scintillans bloom along the Tuticorin coast, southeast coast of India. Mar. Pollut. Bull. 2023, 195, 115523. [Google Scholar] [CrossRef]
- Hofmann, R.; Tietje, M.; Aberhan, M. Diversity partitioning in Phanerozoic benthic marine communities. Proc. Natl. Acad. Sci. USA 2019, 116, 79–83. [Google Scholar] [CrossRef]
- Stephens, C.R.; Heau, J.G.; González, C.; Ibarra-Cerdeña, C.N.; Sánchez-Cordero, V.; González-Salazar, C. Using biotic interaction networks for prediction in biodiversity and emerging diseases. PLoS ONE 2009, 4, e5725. [Google Scholar] [CrossRef] [PubMed]
- Luna, P.; Corro, E.J.; Antoniazzi, R.; Dáttilo, W. Measuring and linking the missing part of biodiversity and ecosystem function: The diversity of biotic interactions. Diversity 2020, 12, 86. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Tuzhilina, E.; Tozzi, L.; Hastie, T. Canonical correlation analysis in high dimensions with structured regularization. Stat. Modelling 2023, 23, 203–227. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Gao, Y.H.; Chen, C.P.; Liang, J.R.; Sun, L.; Zhen, Y.; Qiao, L. Seasonal variations of phytoplankton assemblages and its relation to environmental variables in a scallop culture sea area of Bohai Bay, China. Mar. Pollut. Bull. 2016, 113, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Guo, S. Dinoflagellate heterotrophy. Acta Ecol. Sin. 2011, 31, 6270–6286. [Google Scholar]
- Wang, X.; Yang, S.; Zhang, Q. Coupling effect of phytoplankton community structure and environmental factors in the Bohai Sea of China. Mar. Pollut. Bull. 2022, 179, 113707. [Google Scholar] [CrossRef]
- Li, L.; Wang, Y.; Liu, D. Phytoplankton shifts in the Central Bohai Sea over the last 250 years reflect eutrophication and input from the Yellow River. Ecol. Indic. 2021, 126, 107676. [Google Scholar] [CrossRef]
- Tang, Y.; Huangfu, J.; Huang, R.; Chen, W. Surface warming reacceleration in offshore China and its interdecadal effects on the East Asia-Pacific climate. Sci. Rep. 2020, 10, 14811. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Sun, J.; Wei, Y.; Liu, Z.; Xin, Y.; Guo, Y.; Gu, T. Seasonal shift of a phytoplankton (>5 µm) community in Bohai Sea and the adjacent Yellow Sea. Diversity 2021, 13, 65. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Ge, H.; Song, C.; Li, C.; Liu, S. Composition and Distribution of Microeukaryotic Communities in the Surface Sediments of Five Geographic Regions of Bohai Sea Based on 18S rDNA Amplicon Sequencing. J. Mar. Sci. Eng. 2024, 12, 410. https://doi.org/10.3390/jmse12030410
Zhang W, Ge H, Song C, Li C, Liu S. Composition and Distribution of Microeukaryotic Communities in the Surface Sediments of Five Geographic Regions of Bohai Sea Based on 18S rDNA Amplicon Sequencing. Journal of Marine Science and Engineering. 2024; 12(3):410. https://doi.org/10.3390/jmse12030410
Chicago/Turabian StyleZhang, Wenquan, Huameng Ge, Chengbing Song, Chengcheng Li, and Shenghao Liu. 2024. "Composition and Distribution of Microeukaryotic Communities in the Surface Sediments of Five Geographic Regions of Bohai Sea Based on 18S rDNA Amplicon Sequencing" Journal of Marine Science and Engineering 12, no. 3: 410. https://doi.org/10.3390/jmse12030410
APA StyleZhang, W., Ge, H., Song, C., Li, C., & Liu, S. (2024). Composition and Distribution of Microeukaryotic Communities in the Surface Sediments of Five Geographic Regions of Bohai Sea Based on 18S rDNA Amplicon Sequencing. Journal of Marine Science and Engineering, 12(3), 410. https://doi.org/10.3390/jmse12030410