
The Palaeobiology of the False Mako Shark, Parotodus benedenii (Le Hon, 1871): A View from the Pliocene Mediterranean Sea




{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
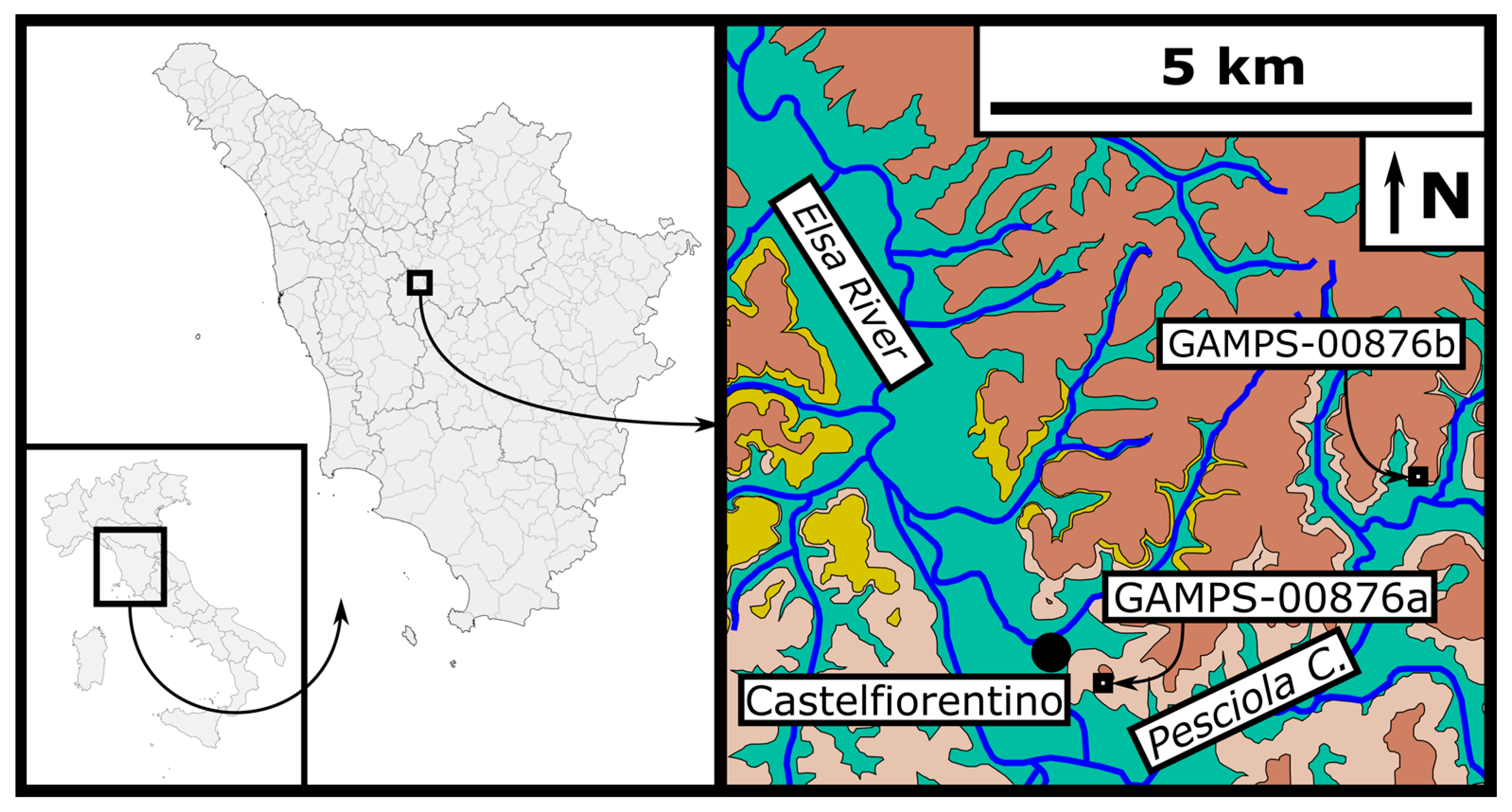
2. Stratigraphic Framework
3. Materials and Methods
3.1. Repository
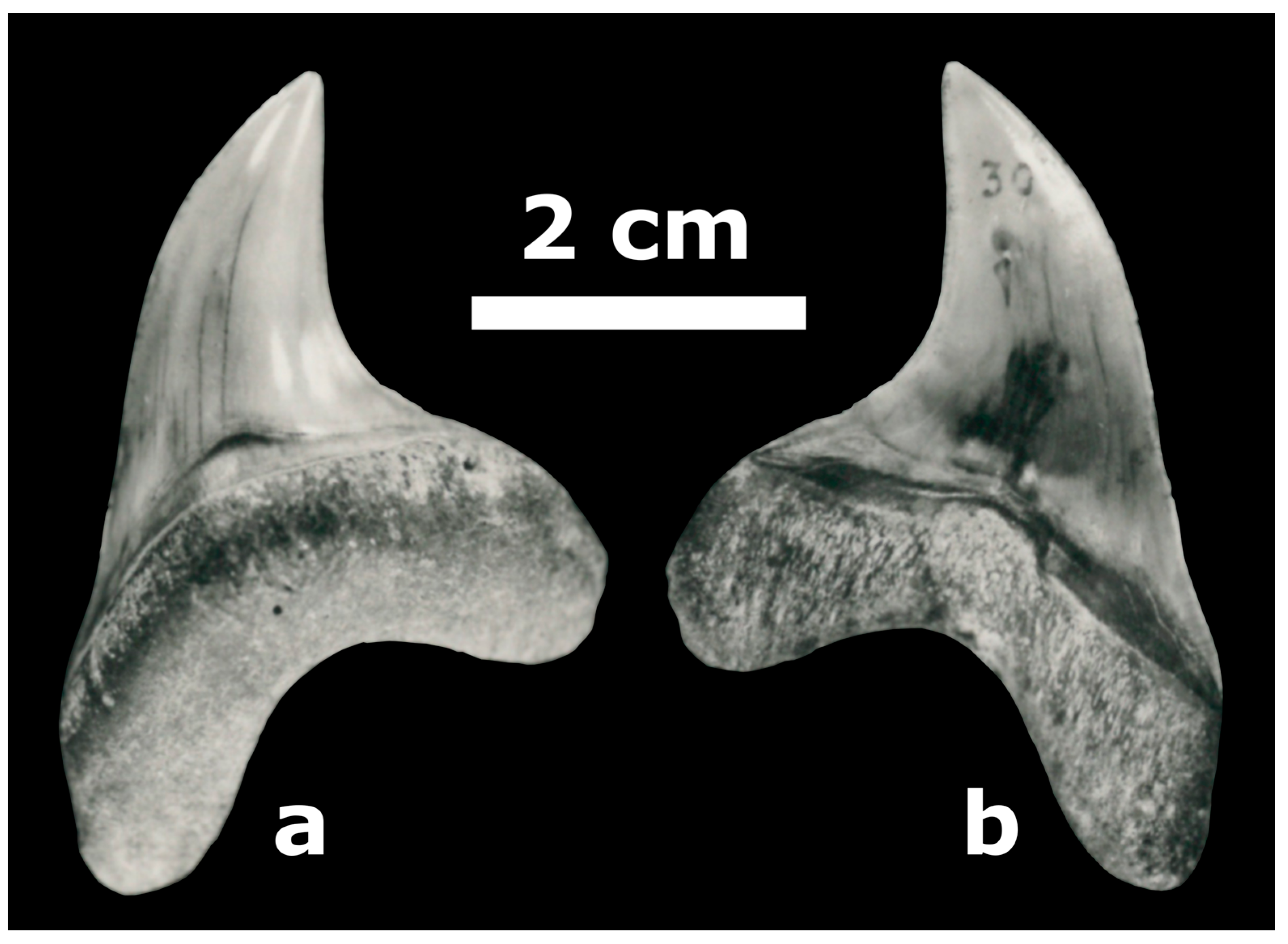
3.2. Specimen Documentation
4. Systematics
5. The Palaeobiology of Parotodus benedenii: An Updated Review
5.1. Body Size
5.2. Body Shape
5.3. Habitat Preferences
5.4. Trophic Ecology
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cappetta, H. Handbook of Paleoichthyology, Vol. 3E. Chondrichthyes: Mesozoic and Cenozoic Elasmobranchii: Teeth; Dr. Friedrich Pfeil: Munich, Germany, 2012; pp. 1–512. [Google Scholar]
- Ward, D.J.; Nakatani, D.; Bernard, E.L. A new species of Parotodus (Lamniformes: Otodontidae) from the Oligocene of Japan. In Proceedings of the 61st PalAss Annual Meeting, London, UK, 17–18 December 2017. [Google Scholar]
- Canevet, J.-M. Des dents de requins fossiles! 9. Le genre Parotodus: Le faux requin-mako. Fossiles-Rev. Fr. Paléontol. 2019, 37, 39–50. [Google Scholar]
- Kent, B.W. The cartilaginous fishes (chimaeras, sharks, and Rays) of Calvert Cliffs, Maryland, USA. In The Geology and Vertebrate Paleontology of Calvert Cliffs, Maryland; Godfrey, S.J., Ed.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2018; pp. 45–156. [Google Scholar]
- Le Hon, H. Préliminaire d’un Mémoire sur les Poissons Fossiles Tertiaires de Belgique; Muquardt: Bruxelles, Belgium, 1871; pp. 1–15. [Google Scholar]
- Menesini, E. Ittiodontoliti miocenici di Terra d’Otranto. Palaeontogr. Ital. 1969, 65, 1–61. [Google Scholar]
- De Ceuster, J. Stratigrafische interpretatie van jongcenozoische afzettingen bij Rumst (België, provincie Antwerpen) en beschrijving van de in een post-mioceen basisgrind aangetroffen vissenfauna, II. Systematische beschrijvingen en conclusies. Meded. Werkgr. Tert. Kwart. Geol. 1976, 13, 119–172. [Google Scholar]
- Applegate, S.P.; Espinosa-Arrubarrena, L. The Fossil History of Carcharodon and Its Possible Ancestor, Cretolamna. A Study in Tooth Identification. In Great White Sharks. The Biology of Carcharodon carcharias; Klimley, A.P., Ainley, D., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 19–36. [Google Scholar] [CrossRef]
- Cappetta, H. Modification du statut générique de quelques espèces de sélaciens crétacés et tertiaires. Palaeovertebrata 1980, 10, 29–42. [Google Scholar]
- Heinzl, L. Collecting fossil shark teeth in the Lee Creek Mine, Aurora, Nc. M.A.P.S. Dig. 2000, 23, 110–117. [Google Scholar]
- Fitzgerald, E.; Schmidt, R. Fossils of Beaumaris; Museum of Victoria: Melbourne, Australia, 2015; pp. 1–15. [Google Scholar]
- Shimada, K.; Chandler, R.E.; Lam, O.L.T.; Tanaka, T.; Ward, D.J. A new elusive otodontid shark (Lamniformes: Otodontidae) from the lower Miocene, and comments on the taxonomy of otodontid genera, including the ‘megatoothed’ clade. Hist. Biol. 2017, 29, 704–714. [Google Scholar] [CrossRef]
- Herman, J. Réflexions sur la systematique des Galeoidei et sur les affinites du genre Cetorhinus, a l’occasion de la découverte d’éléments de la denture d’un exemplaire fossile dans le sables du Kattendijk à Kallo (Pliocène inférieur, Belgique). Ann. Soc. Géol. Belg. 1979, 102, 357–377. [Google Scholar]
- Siverson, M. A New Large Lamniform Shark from the Uppermost Gearle Sandstone (Cenomanian, Late Cretaceous) of Western Australia. Transactions of the Royal Society of Edinburgh. Earth Sci. 1999, 90, 49–65. [Google Scholar] [CrossRef]
- Purdy, R.; Schneider, V.; Applegate, S.; McLellan, J.; Meyer, R.; Slaughter, R. The Neogene Sharks, Rays, and Bony Fishes from Lee Creek Mine, Aurora, North Carolina. In Geology and Paleontology of the Lee Creek Mine, North Carolina, 3rd ed.; Smithsonian Contributions to Paleobiology; Ray, C.E., Bohaska, D.J., Eds.; Smithsonian Institution Press: Washington, DC, USA, 2001; Volume 90, pp. 71–202. [Google Scholar]
- Iserbyt, A.; De Schutter, P.J. Quantitative analysis of elasmobranch assemblages from two successive Ypresian (early Eocene) facies at Marke, western Belgium. Geol. Belg. 2012, 15, 146–153. [Google Scholar]
- Mas, G. Presència de Parotodus benedeni (Le Hon, 1871) (Pisces: Chondrichthyes: Otodontidae) al Neogen de Mallorca i Menorca (Illes Balears, Mediterrania occidental). Consideracions taxonomiques i paleoambientals. Boll. Soc. Hist. Nat. Balears 2003, 46, 85–90. [Google Scholar]
- González-Barba, G. Descripción de la Asociación Faunistica de Elasmobranquios Fósiles del Conglomerado Basal en las Formaciones San Gregorio y el Cien (Oligoceno Temprano) de Baja California Sur, México. Ph.D. Thesis, Centro Interdisciplinario de Ciencias Marinas, La Paz, Mexico, 2008. [Google Scholar]
- Betancort, J.F.; Lomoschitz, A.; Meco, J. Early Pliocene fishes (Chondrichthyes, Osteichthyes) from Gran Canaria and Fuerteventura (Canary Islands, Spain). Estud. Geol. 2016, 72, e054. [Google Scholar] [CrossRef]
- Carrillo-Briceño, J.D.; Villafaña, J.A.; De Gracia, C.; Flores-Alcívar, F.F.; Kindlimann, R.; Abella, J. Diversity and paleoenvironmental implications of an elasmobranch assemblage from the Oligocene–Miocene boundary of Ecuador. PeerJ 2020, 8, e9051. [Google Scholar] [CrossRef] [PubMed]
- Malyshkina, T.P.; Ward, D.J.; Nazarkin, M.V.; Nam, G.S.; Kwon, S.H.; Lee, J.H.; Kim, T.W.; Kim, D.K.; Baek, D.S. Miocene Elasmobranchii from the Duho Formation, South Korea. Hist. Biol. 2023, 35, 1726–1741. [Google Scholar] [CrossRef]
- Belyaev, G.M.; Glikman, L.S. The teeth of sharks on the floor of the Pacific Ocean. Trudy Inst. Okaenol. Akad. Nauk. SSSR 1970, 88, 236–251. (In Russian) [Google Scholar]
- Roux, C.; Geistdoerfer, P. Dents de requins et bulles tympaniques de cétacés: Noyaux de nodules polymétalliques récoltés dans l’océan Indien. Cybium 1988, 12, 129–137. [Google Scholar]
- Ward, D.J.; Bonavia, C.G. Additions to, and a review of, the Miocene shark and ray fauna of Malta. Cent. Medit. Nat. 2001, 3, 131–146. [Google Scholar]
- Vera-Peláez, J.L.; Lozano-Francisco, M.C.; Pascual-Alayón, P.; Báez, J.C. Primera cita de Paratodus benedeni (Le Hon, 1871) (Chondrichthyes, Lamnidae) en los depósitos del Mioceno superior (Tortoniense) de Antequera (Málaga) y del Plioceno inferior (Zancliense) del Puerto de Santa María (Cádiz), sur de España. Rev. Bras. Paleontol. 2016, 19, 341–346. [Google Scholar] [CrossRef]
- Marsili, S.; Carnevale, G.; Danese, E.; Bianucci, G.; Landini, W. Early Miocene vertebrates from Montagna della Maiella, Italy. Ann. Paléontol. 2007, 93, 27–66. [Google Scholar] [CrossRef]
- Marsili, S. Analisi Sistematica, Paleoecologica e Paleobiogeografica della Selaciofauna Plio-Pleistocenica del Mediterraneo. Ph.D. Thesis, Università di Pisa, Pisa, Italy, 2007. [Google Scholar]
- Manganelli, G.; Benocci, A.; Spadini, V. The scientific bibliography of Roberto Lawley (1818–1881) and his contribution to the study of fossil sharks. Arch. Nat. Hist. 2003, 33, 267–281. [Google Scholar] [CrossRef]
- Lawley, R. Nuovi Studi Sopra i Pesci ed Altri Vertebrati Fossili delle Colline Toscane; Tipografia dell’arte della Stampa: Firenze, Italy, 1876; pp. 1–122. [Google Scholar]
- Landini, W. Revisione degli «ittiodontoliti pliocenici» della Collezione Lawley. Palaeontogr. Ital. 1977, 70, 92–134. [Google Scholar]
- Landini, W. Nuovi Studi Sugli Ittiodontoliti della Collezione Lawley. Master’s Thesis, Università di Pisa, Pisa, Italy, 1972. [Google Scholar]
- Benvenuti, M.; Del Conte, S.; Scarselli, N.; Dominici, S. Hinterland basin development and infilling through tectonic and eustatic processes: Latest Messinian-Gelasian Valdelsa Basin, Northern Apennines, Italy. Basin Res. 2014, 26, 387–402. [Google Scholar] [CrossRef]
- Capellini, G. Balene fossili toscane. III Idiocetus Guicciardini. Mem. R. Accad. Sci. Ist. Bologna 1905, 6, 71–80. [Google Scholar]
- Azzaroli, A. The cervid genus Pseudodama n.g. in the Villafranchian of Tuscany. Palaeontogr. Ital. 1992, 79, 1–41. [Google Scholar]
- Borselli, V.; Cozzini, F. Il recupero di un cetaceo fossile in località Ponte a Elsa (Pisa). Museol. Sci. 1991, 8, 9–22. [Google Scholar]
- Bianucci, G. A new record of baleen whale from the Pliocene of Tuscany (Italy). Atti Soc. Tosc. Sci. Nat. Mem. Ser. A 1995, 102, 101–104. [Google Scholar]
- Rustioni, M.; Mazza, P. Taphonomic analysis of Tapirus arvernensis remains from the lower Valdarno (Tuscany, Central Italy). Geobios 2001, 34, 469–474. [Google Scholar] [CrossRef]
- Abbazzi, L.; Benvenuti, M.; Ceci, M.E.; Esu, D.; Faranda, C.; Rook, L.; Tangocci, F. The end of the Lago-Mare time in the SE Valdelsa Basin (central Italy): Interference between local tectonism and regional sea-level rise. Geodiversitas 2008, 30, 611–639. [Google Scholar]
- Dominici, S.; Rook, L.; Benvenuti, M.; Abbazzi, L. Tapir remains in paralic deposits of Pliocene age in Lower Valdarno (Tuscany, Italy): Facies analysis and taphonomy. Geobios 1995, 28, 131–135. [Google Scholar] [CrossRef]
- Dominici, S.; Danise, S.; Benvenuti, M. Pliocene stratigraphic paleobiology in Tuscany and the fossil record of marine megafauna. Earth-Sci. Rev. 2018, 176, 277–310. [Google Scholar] [CrossRef]
- Collareta, A.; Bosselaers, M.; Bianucci, G. Jumping from turtles to whales: A Pliocene fossil record depicts an ancient dispersal of Chelonibia on mysticetes. Riv. Ital. Paleontol. Strat. 2016, 122, 35–44. [Google Scholar] [CrossRef]
- Collareta, A.; Casati, S.; Catanzariti, R.; Di Cencio, A. First record of the knifetooth sawfish Anoxypristis (Elasmobranchii: Rhinopristiformes) from the Pliocene of Tuscany (central Italy). Neues Jahrb. Geol. Paläontol. Abh. 2017, 284, 289–297. [Google Scholar] [CrossRef]
- Collareta, A.; Casati, S.; Zuffi, M.A.L.; Di Cencio, A. First authentic record of the freshwater turtle Mauremys from the upper Pliocene of Italy, with a new occurrence of the rarely reported ichnotaxon Thatchtelithichnus holmani. Carnets Geol. 2020, 20, 301–313. [Google Scholar] [CrossRef]
- Collareta, A.; Merella, M.; Casati, S.; Di Cencio, A. Did titanic stingrays wander the Pliocene Mediterranean Sea? Some notes on a giant-sized myliobatoid stinger from the Piacenzian of Italy. Neues Jahrb. Geol. Paläontol. Abh. 2020, 298, 155–164. [Google Scholar] [CrossRef]
- Collareta, A.; Merella, M.; Casati, S.; Coletti, G.; Di Cencio, A. Another thermophilic” Miocene survivor” from the Italian Pliocene: A geologically young occurrence of the pelagic eagle ray Aetobatus in the Euro-Mediterranean region. Carnets Geol. 2021, 21, 203–214. [Google Scholar] [CrossRef]
- Collareta, A.; Mollen, F.H.; Merella, M.; Casati, S.; Di Cencio, A. Remarkable multicuspid teeth in a new elusive skate (Chondrichthyes, Rajiformes) from the Mediterranean Pliocene. PalZ 2021, 95, 117–128. [Google Scholar] [CrossRef]
- Merella, M.; Collareta, A.; Casati, S.; Di Cencio, A.; Bianucci, G. Pliocene Geotourism: Innovative Projects for Valorizing the Paleontological Heritage of Three Different-Staged Quarries of Tuscany (Central Italy). Geoheritage 2023, 15, 82. [Google Scholar] [CrossRef]
- Glikman, L.S. Paleogene Sharks and Their Stratigraphic Importance; Akad. Nauk S.S.S.R.: Moscow, Russia, 1964; pp. 1–228. (In Russian) [Google Scholar]
- Leriche, M. Les poissons tertiaires de Belgique. IV. Les poissons néogènes. Mém. Mus. Roy. Hist. Nat. Belg. 1926, 32, 367–472. [Google Scholar]
- Reinecke, T.; Louwyw, S.; Havekost, U.; Moths, H. The Elasmobranch Fauna of the Late Burdigalian, Miocene, at Werder-Uesen, Lower Saxony, Germany, and Its Relationships with Early Miocene Faunas in the North Atlantic, Central Paratethys and Mediterranean. Palaeontos 2011, 20, 1–207. [Google Scholar]
- Collareta, A.; Kindlimann, R.; Baglioni, A.; Landini, W.; Sarti, G.; Altamirano, A.; Urbina, M.; Bianucci, G. Dental Morphology, Palaeoecology and Palaeobiogeographic Significance of a New Species of Requiem Shark (Genus Carcharhinus) from the Lower Miocene of Peru (East Pisco Basin, Chilcatay Formation). J. Mar. Sci. Eng. 2022, 10, 1466. [Google Scholar] [CrossRef]
- ICZN [International Commission on Zoological Nomenclature]. International Code of Zoological Nomenclature, 4th ed.; International Trust for Zoological Nomenclature: London, UK, 1999; pp. 1–106. [Google Scholar]
- Dubois, A. Genitives of species and subspecies nomina derived from personal names should not be emended. Zootaxa 2007, 1550, 49–68. [Google Scholar] [CrossRef]
- Diesel Abegg, A.; Malta-Borges, L.; Tavares de Carvalho, J.D.; Mario-Da-Rosa, C.; Di Mare, R.A. Cases of unjustified emendations of scientific names dedicated to people in tarantulas (Mygalomorphae: Theraphosidae). Rev. Colomb. Entomol. 2016, 42, 201–202. [Google Scholar] [CrossRef]
- Ranzani, C. De Naturali Historia Dissertations; Novis Commentariis Accademie Instituti Scientiarium Bononiensis: Bologna, Italia, 1844; pp. 1–41. [Google Scholar]
- Kent, B.W.; Powell, G.W., Jr. Reconstructed Dentition of the Rare Lamnoid Shark, Parotodus benedeni (Le Hon) from the Yorktown Formation (Early Pliocene) at Lee Creek Mine, North Carolina. Mosasaur 1999, 6, 1–10. [Google Scholar]
- Kent, B.W. Speculations on the Size and Morphology of the Extinct Lamnoid Shark, Parotodus benedeni (Le Hon). Mosasaur 1999, 6, 11–15. [Google Scholar]
- Marsili, S. Systematic, paleoecologic and paleobiogeographic analysis of the Plio-Pleistocene Mediterranean elasmobranch fauna. Atti Soc. Tosc. Sci. Nat. Mem. Ser. A 2008, 113, 81–88. [Google Scholar]
- Everaert, S. Parotodus benedenii (Le Hon, 1871) uit het Zand van Antwerpen (Midden Mioceen) te Berchem (Antwerpen, België). Afzett. Werkgr. Tert. Kwart. Geol. 2020, 41, 36–40. [Google Scholar]
- Boessenecker, R.W.; Ehret, D.J.; Long, D.J.; Churchill, M.; Martin, E.; Boessenecker, S.J. The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: A view from the eastern North Pacific. PeerJ 2019, 7, e6088. [Google Scholar] [CrossRef]
- Pimiento, C.; Clements, C.F. When did Carcharocles megalodon become extinct? A new analysis of the fossil record. PLoS ONE 2014, 9, e111086. [Google Scholar] [CrossRef]
- Goto, M.; Kobayashi, F.; Osawa, S. On the teeth of the genus Isurus from Tomioka City, Gumma Prefecture, Japan (preliminary report). J. Geol. Soc. Jpn. 1978, 84, 271–272. [Google Scholar] [CrossRef]
- Adnet, S.; Balbino, A.C.; Antunes, M.T.; Marín-Ferrer, J.M. New fossil teeth of the White Shark (Carcharodon carcharias) from the Early Pliocene of Spain. Implication for its paleoecology in the Mediterranean. Neues Jahrb. Geol. Palaontol. Abh. 2010, 256, 7–16. [Google Scholar] [CrossRef]
- Collareta, A.; Casati, S.; Di Cencio, A.; Bianucci, G. The deep past of the white shark, Carcharodon carcharias, in the Mediterranean Sea: A synthesis of its palaeobiology and palaeoecology. Life 2023, 13, 2085. [Google Scholar] [CrossRef]
- Greenfield, T. List of skeletal material from megatooth sharks (Lamniformes, Otodontidae). Paleoichthys 2022, 4, 1–9. [Google Scholar]
- Greenfield, T. Additions to “List of skeletal material from megatooth sharks”, with a response to Shimada (2022). Paleoichthys 2022, 6, 6–11. [Google Scholar]
- Shimada, K. Phylogenetic affinity of the extinct shark family Otodontidae within Lamniformes remains uncertain–Comments on “List of skeletal material from megatooth sharks (Lamniformes, Otodontidae)” by Greenfield. Paleoichthys 2022, 6, 1–5. [Google Scholar]
- Dolton, H.R.; Jackson, A.L.; Deaville, R.; Hall, J.; Hall, G.; McManus, G.; Perkins, M.W.; Rolfe, R.A.; Snelling, E.P.; Houghton, J.D.R.; et al. Regionally endothermic traits in planktivorous basking sharks Cetorhinus maximus. Endanger. Species Res. 2023, 51, 227–232. [Google Scholar] [CrossRef]
- Griffiths, L.M.; Eagle, R.A.; Kim, S.L.; Flores, R.; Becker, M.A.; Maisch, H.M., IV; Trayler, R.B.; Chan, R.L.; McCormack, J.; Akhtara, A.A.; et al. Endothermic physiology of extinct megatooth sharks. Proc. Natl. Acad. Sci. USA 2023, 120, e221815312. [Google Scholar] [CrossRef]
- Kuga, N. Revision of the Neogene Mackerel Shark Genus Isurus from Japan. Mem. Fac. Sci. Kyoto Univ. Ser. Geol. Miner. 1985, 51, 1–20. [Google Scholar]
- Compagno, L.J.V. Alternative life-history styles of cartilaginous fishes in time and space. Environ. Biol. Fishes 1990, 28, 33–75. [Google Scholar] [CrossRef]
- Boustany, A.M.; Davis, S.F.; Pyle, P.; Anderson, S.D.; Le Boeuf, B.J.; Block, B.A. Expanded niche for white sharks. Nature 2002, 415, 35–36. [Google Scholar] [CrossRef]
- Compagno, L.J.V. FAO species catalogue. Sharks of the world. In An Annotated and Illustrated Catalogue of Shark Species Known to Date. Part 1. Hexanchiformes to Lamniformes; FAO: Rome, Italy, 1984; Volume 4, pp. 1–258. [Google Scholar]
- Collareta, A.; Merella, M.; Casati, S.; Di Cencio, A.; Bianucci, G. Smoking guns for cold cases: The find of a Carcharhinus tooth piercing a fossil cetacean rib, with notes on the feeding ecology of some Mediterranean Pliocene requiem sharks. Neues Jahrb. Geol. Paläontol. Abh. 2022, 305, 145–152. [Google Scholar] [CrossRef]
- Dominici, S.; Danise, S.; Cau, S.; Freschi, A. The awkward record of fossil whales. Earth-Sci. Rev. 2020, 205, 103057. [Google Scholar] [CrossRef]
- Noriega, J.I.; Cione, A.L.; Aceñolaza, F.G. Shark tooth marks on Miocene balaenopterid cetacean bones from Argentina. Neues Jahrb. Geol. Paläontol. Abh. 2007, 245, 185–192. [Google Scholar] [CrossRef]
- Collareta, A.; Cigala Fulgosi, F.; Bianucci, G. A new kogiid sperm whale from northern Italy supports psychrospheric conditions in the early Pliocene Mediterranean Sea. Acta Palaeontol. Polon. 2019, 64, 609–626. [Google Scholar] [CrossRef]
- Godfrey, S.J.; Lowry, A.J. The ichnospecies Linichnus bromleyi on a Miocene baleen whale radius preserving multiple shark bite-shake traces suggests scavenging. Carnets Geol. 2021, 21, 391–398. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collareta, A.; Casati, S.; Di Cencio, A. The Palaeobiology of the False Mako Shark, Parotodus benedenii (Le Hon, 1871): A View from the Pliocene Mediterranean Sea. J. Mar. Sci. Eng. 2023, 11, 1990. https://doi.org/10.3390/jmse11101990
Collareta A, Casati S, Di Cencio A. The Palaeobiology of the False Mako Shark, Parotodus benedenii (Le Hon, 1871): A View from the Pliocene Mediterranean Sea. Journal of Marine Science and Engineering. 2023; 11(10):1990. https://doi.org/10.3390/jmse11101990
Chicago/Turabian StyleCollareta, Alberto, Simone Casati, and Andrea Di Cencio. 2023. "The Palaeobiology of the False Mako Shark, Parotodus benedenii (Le Hon, 1871): A View from the Pliocene Mediterranean Sea" Journal of Marine Science and Engineering 11, no. 10: 1990. https://doi.org/10.3390/jmse11101990
APA StyleCollareta, A., Casati, S., & Di Cencio, A. (2023). The Palaeobiology of the False Mako Shark, Parotodus benedenii (Le Hon, 1871): A View from the Pliocene Mediterranean Sea. Journal of Marine Science and Engineering, 11(10), 1990. https://doi.org/10.3390/jmse11101990