

Adaptability of Toxoptera aurantii (Hemiptera: Aphididae) to Different Tea Cultivars

Abstract
:1. Introduction
2. Materials and Methods
2.1. Aphids
2.2. Plants
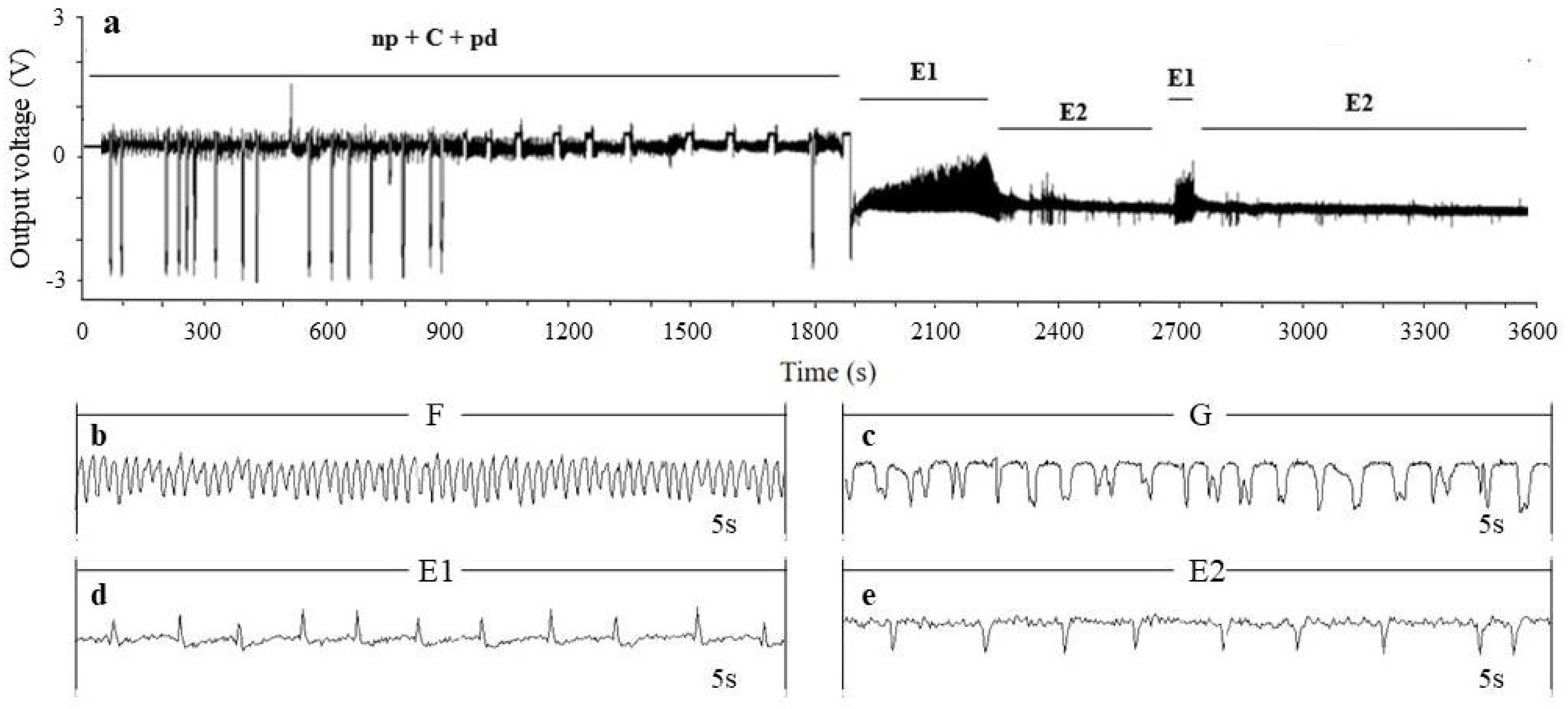
2.3. EPG Recording
2.4. Population Dynamics
2.5. Statistical Analysis
3. Results
3.1. Feeding Behaviour of Toxoptera aurantii on Seven Tea Cultivars
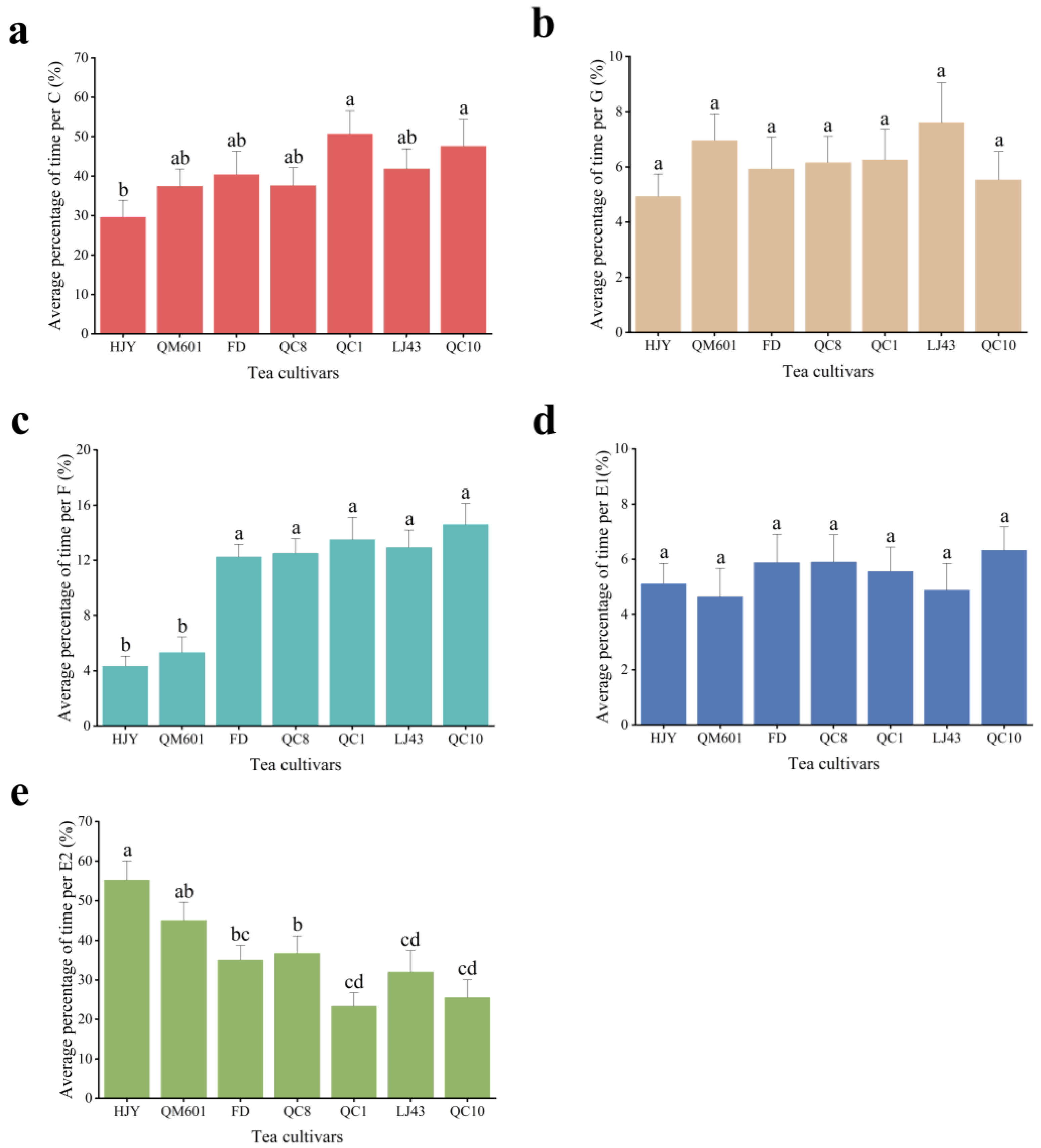
3.1.1. Probing Stage EPG Parameters of Toxoptera aurantii
3.1.2. Phloem-Stage EPG Parameters of Toxoptera aurantii
3.1.3. Proportion of Each Waveform of Toxoptera aurantii
3.2. Population Dynamics of Toxoptera aurantii on Seven Tea Cultivars
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Plant responses to herbivory, wounding, and infection. Int. J. Mol. Sci. 2022, 23, 7031. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.X.; Chi, H.; Zhang, J.; Zhou, Q.; Zhang, R.J. Life-table analysis of the performance of Nilaparvata lugens (Hemiptera: Delphacidae) on two wild rice species. J. Econ. Entomol. 2010, 103, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Li, F.Q. The Population Dynamics and Different Expressed Genes of Myzus persicae (Sulzer) on Different Host Plants. Master’s Thesis, Henan Agricultural University, Zhengzhou, China, 2013. [Google Scholar]
- Jiang, W.B.; Cheng, Q.; Lu, C.H.; Chen, W.L.; Zhao, D.G.; He, Y.Q. Different host plants distinctly influence the adaptability of Myzus persicae (Hemiptera: Aphididae). Agriculture 2022, 12, 2162. [Google Scholar] [CrossRef]
- Zheng, X.M.; Tao, Y.T.; Chi, H.; Wan, F.H.; Chu, D. Adaptability of small brown planthopper to four rice cultivars using life table and population projection method. Sci. Rep. 2017, 7, 42399. [Google Scholar] [CrossRef] [PubMed]
- Chesnais, Q.; Couty, A.; Uzest, M.; Brault, V.; Ameline, A. Plant infection by two different viruses induce contrasting changes of vectors fitness and behavior. Insect Sci. 2017, 26, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, J.Y.; Xu, P.J.; Liu, Y.J.; Dong, Y.H.; Zang, Y.; Cai, X.J.; Ren, G.W. Changes in the nutrient composition of tobacco plants after Potato virus Y infection and their effects on the growth and development of the vector Myzus persicae (Hemiptera: Aphididae). Acta Entomol. Sin. 2020, 63, 181–190. [Google Scholar] [CrossRef]
- Li, C.M. Adaptability of Bemisia tabaci (Gennadius) to the Host Plants and Plant Secondary Substances. Master’s Thesis, Yangzhou University, Yangzhou, China, 2009. [Google Scholar]
- Wang, Y.X.; Chen, H.F.; Yin, Z.Y.; Chen, W.L.; Lu, L.T. The genetic adaptations of Toxoptera aurantia facilitated its rapid multiple plant hosts dispersal and invasion. Genomics 2022, 114, 110472. [Google Scholar] [CrossRef]
- Fang, Y.; Qiao, G.X.; Zhang, G.X. Morphological adaptation of aphid species on different host plant leaves. Acta Entomol. Sin. 2011, 54, 157–178. [Google Scholar] [CrossRef]
- Golizadeh, A.; Abedi, Z.; Borzoui, E.; Golikhajeh, N.; Jafary, M. Susceptibility of five sugar beet cultivars to the black bean aphid, aphis fabae scopoli (Hemiptera: Aphididae). Neotrop. Entomol. 2016, 45, 427–432. [Google Scholar] [CrossRef]
- Han, B.Y.; Chen, Z.M. The differences between probing behaviour of tea aphids on different parts of tea tree. J. Plant Prot. 2001, 28, 7–11. [Google Scholar]
- Helden, M.V.; Tjallingii, W.F. Tissue localization of lettuce resistance to the aphid Nasonovia ribisnigri using electrical penetration graphs. Exp. Appl. 1993, 68, 269–278. [Google Scholar] [CrossRef]
- Zhao, R.N.; Wu, C.X.; He, Y.Q.; Yu, C.; Liu, J.F.; Li, T.S.; Zhou, C.Y.; Chen, W.L. Different host plants distinctly influence the feeding ability of the brown citrus aphid Toxoptera citricida. Insects 2021, 12, 864. [Google Scholar] [CrossRef] [PubMed]
- Tjallingii, W.F. Salivary secretions by aphids interacting with proteins of phloem wound responses. J. Exp. Bot. 2006, 57, 739–745. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.Y.; Liu, L.F.; Yu, X.P.; Han, B.Y. Evaluation of the resistance of different tea cultivars to tea aphids by EPG technique. J. Integr. Agric. 2012, 11, 2028–2034. [Google Scholar] [CrossRef]
- Garzo, E.; Soria, M.L.; Gómez-Guillamon, M.L.; Fereres, A. Feeding behavior of Aphis gossypii on resistant accessions of different melon genotypes (Cucumis melo). Phytoparasitica 2002, 30, 129–140. [Google Scholar] [CrossRef]
- Peng, H.C.; Walker, G.P. Sieve element occlusion provides resistance against aphis gossypii in TGR-1551 melons. Insect Sci. 2018, 1, 33–48. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.X.; Chang, W.J.; Zhan, Y.D.; Liu, Z.; Liu, Y. Investigation on the resistance of wheat germplasm resources to aphid based on fuzzy recognition and electrical penetration graph (EPG) techniques. Chin. J. Appl. Ecol. 2020, 31, 3248–3254. [Google Scholar] [CrossRef]
- Ouyang, Y.T.; Tian, S.; Hu, L.; Zhao, D.G.; Chen, Z.W.; He, Y.Q. Effects of physical and chemical properties of tea leaves on feeding selectivity of Toxoptera aurantii Boyer de Fonscolombe. Plant Prot. 2023. submitted. [Google Scholar] [CrossRef]
- Sarria, E.; Cid, M.; Garzo, E.; Fereres, A. Excel workbook for automatic parameter calculation of EPG data. Comput. Electron. Agric. 2009, 67, 35–42. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Electrical nature of recorded signals during stylet penetration by aphids. Entomol. Exp. Appl. 1985, 38, 177–186. [Google Scholar] [CrossRef]
- He, Y.Q.; Zhang, Y.Q.; Chen, J.N.; Chen, W.L.; Zeng, X.Y.; Chen, H.T.; Ding, W. Effects of Aphidius gifuensis on the feeding behavior and potato virus Y transmission ability of Myzus persicae. Insect Sci. 2017, 25, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Kanissery, R.; Ammar, E.-D.; Cabral, I.; Markle, L.T.; Patt, J.M.; Stelinski, L.L. Feeding behavior of Asian citrus psyllid [Diaphorina citri (Hemiptera: Liviidae)] nymphs and adults on common weeds occurring in cultivated citrus described using electrical penetration graph recordings. Insects 2020, 11, 48. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Guo, Z.; Yang, Z.; Han, H.; Wang, S.; Xu, H.; Yang, X.; Yang, F.; Wu, Q.; Xie, W.; et al. Whitefly hijacks a plant detoxification gene that neutralizes plant toxins. Cell 2021, 184, 1693–1705. [Google Scholar] [CrossRef] [PubMed]
- He, Y.Q.; Jiang, W.B.; Ding, W.; Chen, W.L.; Zhao, D.G. Effects of PVY-infected tobacco plants on the adaptation of Myzus persicae (Hemiptera: Aphididae). Insects 2022, 13, 1120. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.L.; Qiao, C.Y.; Kong, H.L.; Lu, M.; Mao, N.; Zhu, S.D. Adaptation of Bemisia tabaci biotype Q (Hemiptera: Aleyrodidae) to different tomato cultivars. J. Plant Prot. 2015, 42, 734–740. [Google Scholar] [CrossRef]
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.J.; He, K.L.; Wang, Z.Y. Comparison of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: Potential risks to potato and tobacco crops. Insect Sci. 2021, 28, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Acharya, R.; Malekera, M.J.; Dhungana, S.K.; Sharma, S.R.; Lee, K.Y. Impact of rice and potato host plants is higher on the reproduction than growth of corn strain Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 256. [Google Scholar] [CrossRef]
- Brito, N.F.; Moreira, M.F.; Melo, A.C.A. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef]
- Luo, C.; Yue, M.; Xu, H.F.; Zhang, Z.L. Application of electrical penetration graph (EPG) in entomological studies and new findings. Acta Entomol. Sin. 2005, 48, 437–443. [Google Scholar] [CrossRef]
- Zhou, H.; Li, Y.; Teng, Z.W.; Du, L.D.; Wan, F.H.; Zhou, H.X. EPG-based comparison of feeding behaviors of three piercing-sucking pests on apple seedlings. Acta Entomol. Sin. 2020, 63, 1207–1214. [Google Scholar] [CrossRef]
- Kempema, L.A.; Cui, X.; Holzer, F.M.; Walling, L.L. Arabidopsis transcriptome changes in response to phloem-feeding silverleaf whitefly nymphs. Similarities and distinctions in responses to aphids. Plant Physiol. 2007, 143, 849–865. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Li, G.H.; Ding, W.B.; Zhang, Y.J.; Li, Y.Z. EPG analysis of feeding behavior of Sogatella furcifera (Hemiptera: Delphacidae) on different rice varieties. Acta Entomol. Sin. 2014, 57, 335–342. [Google Scholar] [CrossRef]
- Zehnder, C.B.; Hunter, M.D. Effects of nitrogen deposition on the interaction between an aphid and its host plant. Ecol. Entomol. 2008, 33, 24–30. [Google Scholar] [CrossRef]
- Pobozniak, M.; Gaborska, M.; Wojtowicz, T. Resistance and tolerance of ten carrot cultivars to the hawthorn-carrot aphid, Dysaphis crataegi Kalt., in Poland. PLoS ONE 2021, 16, e0247978. [Google Scholar] [CrossRef] [PubMed]
- Lantos, E.; Schliephake, E.; Krämer, R.; Will, T.; Nothnagel, T. Feeding behavior of Myzus persicae on asparagus species susceptible and resistant to Asparagus virus 1. Entomol. Exp. Appl. 2019, 167, 360–369. [Google Scholar] [CrossRef]
- Shih, P.Y.; Sugio, A.; Christophe-Simon, J. Molecular mechanisms underlying host plant specificity in aphids. Annu. Rev. Entomol. 2023, 68, 431–450. [Google Scholar] [CrossRef]
- Cao, H.H.; Wu, J.; Zhang, Z.F.; Liu, T.X. Phloem nutrition of detached cabbage leaves varies with leaf age and influences performance of the green peach aphid, Myzus persicae. Entomol. Exp. Appl. 2018, 166, 452–459. [Google Scholar] [CrossRef]
- Ahmed, N.; Darshanee, H.L.C.; Fu, W.; Hu, X.; Fan, Y.; Liu, T. Resistance of seven cabbage cultivars to green peach aphid (Hemiptera: Aphididae). J. Econ. Entomol. 2018, 111, 909–916. [Google Scholar] [CrossRef]
- Feng, H.; Han, H.L.; Pu, P.; Wei, D.; Wang, J.; Liu, Y.H. Effects of five host plant species on the life history and population growth parameters of Myzus persicae (Hemiptera: Aphididae). J Insect Sci. 2019, 19, 15. [Google Scholar] [CrossRef]
- Zhao, R.N. CTV-Mediated Host Selection and Population Fitness of Brown Citrus Aphid, Toxoptera citricida (Kirkaldy) (Homoptera: Aphidiae). Doctoral Thesis, Guizhou University, Guizhou, China, 2022. [Google Scholar]
- Lu, C.; Shen, N.; Jiang, W.; Xie, B.; Zhao, R.; Zhou, G.; Zhao, D.; He, Y.; Chen, W. Different tea germplasms distinctly influence the adaptability of Toxoptera aurantii (Hemiptera: Aphididae). Insects 2023, 14, 695. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Plant secondary metabolites modulate insect behavior-steps toward addiction. Front. Physiol. 2018, 9, 364. [Google Scholar] [CrossRef] [PubMed]
- Korada, R.R. Plant volatiles and insect herbivore interactions: An overview. J. Appl. Zool. Res. 2017, 28, 122–137. [Google Scholar]
- Gao, X.F.; Li, H.L. The relationship between tea leaf structure and selectivity of tea aphids on tea germplasms. Fujian J. Agric. Sci. 2014, 29, 256–260. [Google Scholar] [CrossRef]
- Liu, W.J.; Li, L.Y.; Gao, X.K.; Zhang, K.X.; Li, D.Y.; Luo, J.Y.; Cui, J.J.; Yang, J. Screening of EPG-based parameters for measuring resistance to Aphis gossypii. Chin. J. Biol. Control 2022, 38, 1193–1201. [Google Scholar] [CrossRef]
- Douglas, A. Phloem-sap feeding by animals: Problems and solutions. J. Exp. Bot. 2006, 57, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.H.; Liu, H.R.; Zhang, Z.F.; Liu, T.X. The green peach aphid Myzus persicae perform better on pre-infested Chinese cabbage Brassica pekinensis by enhancing host plant nutritional quality. Sci. Rep. 2016, 6, 21954. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.M.; Zhang, J.X.; Sun, Q.Y.; Ye, T.; Xia, X.J.; Zhang, R.; Ding, Y. Feeding preference and adaptation of Ectropis grisescens (Lepidoptera: Geometridae) to different tea cultivars and their relationship with nutritional components in leaves of tea plants. Acta Entomol. Sin. 2018, 61, 1300–1309. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| EPG Parameter | HJY | QM601 | FD | QC8 | QC1 | LJ43 | QC10 |
|---|---|---|---|---|---|---|---|
| Number of probes | 2.87 ± 0.32 c | 4.93 ± 0.64 b | 5.73 ± 0.78 b | 5.67 ± 0.69 b | 9.00 ± 0.83 a | 6.33 ± 0.83 b | 9.53 ± 0.80 a |
| Sum time of np wave (min) | 65.32 ± 7.89 b | 59.02 ± 9.49 b | 60.50 ± 7.48 b | 89.35 ± 11.19 a | 50.34 ± 7.21 b | 52.19 ± 6.74 b | 47.53 ± 6.76 b |
| Time to first probe from start of EPG recording (min) | 3.32 ± 1.12 b | 9.42 ± 1.87 a | 8.76 ± 1.78 a | 9.92 ± 1.38 a | 8.77 ± 2.44 a | 10.50 ± 1.97 a | 11.46 ± 2.19 a |
| Duration of 1st probe (min) | 39.25 ± 5.18 | 43.14 ± 4.68 | 41.76 ± 5.97 | 43.71 ± 5.98 | 33.15 ± 4.84 | 29.65 ± 5.26 | 37.56 ± 5.34 |
| Number of probes to the 1st E1 | 2.53 ± 0.26 c | 3.67 ± 0.44 bc | 4.60 ± 0.54 ab | 4.53 ± 0.47 ab | 6.20 ± 0.76 a | 4.87 ± 0.62 ab | 6.00 ± 0.87 a |
| Number of pd | 123.07 ± 9.13 | 99.00 ± 10.08 | 106.93 ± 10.09 | 110.80 ± 6.83 | 125.40 ± 11.20 | 117.00 ± 15.33 | 116.20 ± 8.36 |
| Mean duration of pd (s) | 5.27 ± 0.22 | 5.45 ± 0.16 | 5.20 ± 0.21 | 5.39 ± 0.13 | 5.70 ± 0.25 | 5.67 ± 0.20 | 5.51 ± 0.21 |
| Number of short probes (C < 3 min) | 2.53 ± 0.35 b | 3.00 ± 0.45 b | 4.33 ± 0.60 b | 4.40 ± 0.81 b | 6.47 ± 0.18 a | 4.13 ± 0.59 b | 6.60 ± 0.89 a |
| Total duration of C (min) | 84.45 ± 7.65 d | 109.10 + 10.73 cd | 116.89 ± 12.52 bc | 98.87 ± 8.98 cd | 151.82 ± 9.49 a | 125.91 ± 9.75 abc | 143.80 ± 12.95 ab |
| Number of G | 0.93 ± 0.18 | 1.00 ± 0.20 | 0.87 ± 0.17 | 1.13 ± 0.19 | 1.07 ± 0.25 | 1.20 ± 0.26 | 1.07 ± 0.18 |
| Duration of G (min) | 14.35 ± 3.06 | 20.80 ± 3.48 | 16.68 ± 3.67 | 15.93 ± 2.99 | 18.51 ± 3.74 | 22.53 ± 3.70 | 16.60 ± 4.28 |
| Number of F | 0.60 ± 0.16 c | 1.00 ± 0.24 bc | 1.87 ± 0.36 a | 2.00 ± 0.32 a | 1.93 ± 0.30 a | 2.07 ± 0.27 a | 2.13 ± 0.34 a |
| Duration of F (min) | 12.77 ± 2.87 b | 15.42 ± 3.00 b | 36.60 ± 6.24 a | 33.71 ± 5.67 a | 41.51 ± 5.70 a | 39.10 ± 6.51 a | 45.60 ± 5.72 a |
| EPG Parameter | HJY | QM601 | FD | QC8 | QC1 | LJ43 | QC10 |
|---|---|---|---|---|---|---|---|
| Number of E1 | 1.47 ± 0.19 | 1.93 ± 0.23 | 2.07 ± 0.27 | 1.87 ± 0.22 | 1.73 ± 0.21 | 2.13 ± 0.36 | 2.00 ± 0.20 |
| Duration of 1st E | 96.60 ± 7.86 a | 70.06 ± 10.22 a | 53.92 ± 10.18 bc | 56.12 ± 9.37 bc | 37.39 ± 5.25 cd | 45.50 ± 5.27 cd | 29.87 ± 5.06 d |
| Total duration of E1 (min) | 14.90 ± 2.02 | 13.99 ± 2.35 | 17.50 ± 2.03 | 15.67 ± 2.04 | 16.94 ± 2.04 | 13.83 ± 2.18 | 19.55 ± 2.31 |
| Number of E2 | 2.00 ± 0.26 a | 1.73 ± 0.21 ab | 1.47 ± 0.19 ab | 1.53 ± 0.19 ab | 1.40 ± 0.16 b | 1.67 ± 0.21 ab | 1.33 ± 0.16 b |
| Time from 1st probe to 1st E2 | 84.52 ± 9.55 c | 96.72 ± 11.92 bc | 114.21 ± 11.59 abc | 103.64 ± 11.34 abc | 131.56 ± 12.16 a | 120.59 ± 13.50 ab | 126.47 ± 11.43 ab |
| Number of sustained E2 (>10 min) | 1.33 ± 0.19 | 1.20 ± 0.17 | 1.00 ± 0.17 | 1.07 ± 0.18 | 1.13 ± 0.19 | 1.27 ± 0.23 | 0.87 ± 0.17 |
| Total duration of E2 (min) | 156.92 ± 10.38 a | 132.54 ± 12.38 ab | 102.33 ± 12.22 bc | 96.60 ± 11.07 cd | 68.71 ± 11.07 d | 95.34 ± 11.56 cd | 75.80 ± 12.47 cd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Lu, C.; Jiang, W.; Chen, W.; Fan, J.; Niu, S.; Zhao, D. Adaptability of Toxoptera aurantii (Hemiptera: Aphididae) to Different Tea Cultivars. Agriculture 2023, 13, 2039. https://doi.org/10.3390/agriculture13102039
He Y, Lu C, Jiang W, Chen W, Fan J, Niu S, Zhao D. Adaptability of Toxoptera aurantii (Hemiptera: Aphididae) to Different Tea Cultivars. Agriculture. 2023; 13(10):2039. https://doi.org/10.3390/agriculture13102039
Chicago/Turabian StyleHe, Yingqin, Changhao Lu, Wenbin Jiang, Wenlong Chen, Jinjuan Fan, Suzhen Niu, and Degang Zhao. 2023. "Adaptability of Toxoptera aurantii (Hemiptera: Aphididae) to Different Tea Cultivars" Agriculture 13, no. 10: 2039. https://doi.org/10.3390/agriculture13102039
APA StyleHe, Y., Lu, C., Jiang, W., Chen, W., Fan, J., Niu, S., & Zhao, D. (2023). Adaptability of Toxoptera aurantii (Hemiptera: Aphididae) to Different Tea Cultivars. Agriculture, 13(10), 2039. https://doi.org/10.3390/agriculture13102039