Abstract
Wheat is a widely cultivated cereal throughout the world and stress caused by low temperatures significantly affects all stages of wheat development. Seed priming is an effective method to produce stress-resistant plants. This work was carried out to determine whether different priming methods (hormo-, halo-, osmo-, and hydropriming) can increase the resistance of wheat to low-temperature conditions (10 °C). The effect of priming on growth, as well as the biochemical and physiological performance of wheat seedlings were monitored. In general, priming had a significant stimulatory effect on the monitored characteristics. Hormo- and halopriming had a positive effect on the growth, vigor index, and total soluble protein content of wheat seedlings. Additionally, hormopriming reduced the malondialdehyde (MDA) content in wheat seedlings compared to unprimed seeds. A dominant effect on antioxidant enzymes (superoxide-dismutase, catalase, ascorbate peroxidase, guaiacol peroxidase, and pyrogallol peroxidase) was recorded after seed priming with KNO3. The effectiveness of priming was also confirmed through the increased content of phenolic compounds (including flavonoids), and total antioxidant activity. The HPLC analysis showed increased content of chlorogenic acid, catechin, 4-hydroxy benzoic acid, sinapic acid, rutin, naringin, and quercetin in primed wheat seedlings compared to unprimed grown seedlings under low-temperature conditions with the best effects achieved by hormo- and hydropriming. It is concluded that seed priming can be regarded as a promising approach for increasing the resistance of wheat seedlings to low-temperature stress.
1. Introduction
Since it is produced on more land than any other crop and accounts for approximately 26% of the worldwide production of cereals, Triticum aestivum L. (wheat) is one of the most important crops for humanity [1,2]. Wheat grains are a rich source of phytochemicals, including vitamin E, resistant starch, β-glucan, carotenoids, lignans, inulin, sterols, phytates, and phenolic acids [3]. Therefore, wheat is the leading grain in consumption primarily due to its nutritional value but also due to relatively easy harvesting and processing [4]. Despite the increasing demand for wheat production, cultivation of wheat is challenged with difficulties since wheat is sensitive to various abiotic stresses [5]. The most important limiting factors for worldwide production include late frost, high temperature, drought, and heavy rainfall, and these abiotic stresses can significantly reduce crop yield as well as their quality [6,7,8]. Recently, low-temperature stress has been pointed out as a serious challenge for cereal production [9], causing damage to wheat production in the United States [10,11], Australia [12,13,14], Europe [7], and China [15].
Temperature stresses are among the most pivotal abiotic stresses, which inhibit the development of many crops by limiting metabolic processes [16]. Heat stress inhibits germination by preventing imbibition, thereby preventing plant sprouting due to loss of seed vitality [17]. A serious negative effect on wheat growth, productivity, and grain yield quality has been recorded after heat stress [18]. Low-temperature stresses can stop wheat growth by forming ice crystals causing mechanical injuries and metabolic dysfunction, resulting in different metabolic changes (including disruption of metabolic regulations, accumulation of osmolytes, modification of carbohydrate metabolism and photosynthetic properties), and inactivation of many metabolic enzymes [19,20]. By increasing the content of reactive oxygen species (ROS) and malondialdehyde (MDA), as well as by inhibiting photosynthesis, low temperature inhibits the growth of wheat [21]. Xiao et al. [22] suggest that low-temperature stress (even short term) severely impairs wheat growth by limiting photosynthetic capacity through reduced leaf area. The negative impact of temperature stress on the growth and yield of wheat was recorded by Shah et al. [23]. In cereals, low-temperature stress in the vegetative phase limits main reactions in plant cells, resulting in wilting, leaf chlorosis, and necrosis [24,25]. Low temperatures were found to significantly reduce the number of ears per plant, as well as the number of grains per ears in wheat, leading to a decrease in crop quality [26,27]. By developing defense mechanisms to survive under low-temperature conditions, plants have increased their tolerance to stress [20,28]. This process is achieved by molecular, biochemical, and physiological changes, starting with membrane transformation into a rigid structure [29,30]. It has been observed that the concentration of proteins and the activity of peroxidases increased under the influence of low temperature [31]. In addition, improved expression of genes and antioxidant capacity increases crop tolerance to low temperature [32] together with the activation of the biosynthesis of phenolic compounds and the prevention of their oxidation [33]. In addition, the accumulation of soluble sugars significantly contributes to the protection and stabilization of cell membranes during cold acclimatization [32]. To maintain yield stability and crop quality, and to reduce the inhibitory impact of low-temperature stresses, it is necessary to develop methods that improve plant resistance in critical growth stages [27].
The priming technique can be an important method for the production of stress-resistant plants [34,35]. The priming method represents the process of seed hydration with solutions of different osmotic potentials that trigger certain metabolic processes (protein synthesis, repair, or synthesis of new mitochondria) that initiate germination but do not allow the emergence of a radicle [36]. In addition, seed priming improves seedling rooting and growth, vigor index, and ultimately crop yield [37]. Several different approaches to priming methods are commercially applied, including halo-, osmo-, hydro-, chemo- hormo-, or biotic priming [36]. The effects depend on the potential of the agents for priming, crop types, and stress intensity [34]. It was previously confirmed that the priming method increases the wheat tolerance to high temperature [18] and waterlogging [38]. Priming improves the resistance of plants by enabling seed germination, by modulating the expression of genes involved in the gibberellic and abscisic acid synthesis or stress tolerance [17,39]. Greater resistance of primed seeds is also associated with improved activation of DNA and RNA repair processes, antioxidant capacities, and ethylene biosynthesis [39,40].
There is little information on wheat resistance to low temperatures during germination, seedling emergence, or potential modeling of wheat tolerance to given abiotic stress by priming agents. The effects of different priming agents on germination and early seedling growth of wheat under low temperature (10 °C) were evaluated in this study. With the assumption that the biochemical and physiological changes in wheat seeds, caused by priming agents, can be important for seedling tolerance to low temperatures, we aimed to identify the most prominent priming agent for wheat seeds under low-temperature conditions. Therefore, the specific objectives were to investigate the ability of various priming agents to improve the germination potential and early seedling growth, as well as to quantify the low-temperature stress influence on the physiological and biochemical changes of seedlings grown from unprimed compared to primed seeds.
2. Materials and Methods
2.1. Design of Experiment
The winter wheat (Triticum aestivum L. cv. Belija) seeds obtained commercially were stored under optimal conditions (temperature 10–15 °C, air humidity 11–13%). The preparation and priming of wheat seeds were carried out according to Kanjevac et al. [41]. The applied priming agents were for hormopriming 1 mM gibberellic acid (GA3) or 1 mM indol-3-acetic acid (IAA), for halopriming 1% KNO3 or 1% MgSO4, for osmopriming 1% H2O2 or 0.01% ascorbic acid (AA), and for hydropriming distilled water (H2O). Seeds were primed for 12 h in the appropriate solution. After priming, seeds were dried (at room temperature for the next 48 h). Unprimed seeds (untreated) were set as a control sample. Dried primed seeds, together with the untreated ones, were placed in Petri boxes with filter paper and distilled water, and placed in a climate chamber (temperature 10 °C, photoperiod 16/8 h, air humidity 60%). With a moderate-continental climate in Serbia, chosen temperature corresponds to the temperature stress in late autumn which is the seeding time for this wheat cultivar. The low-temperature stress (10 °C) is applied until the two-leaf stage of tested wheat cultivar.
2.2. Seed Moisture Content
Different priming agents can lead to changes in seed moisture content and consequently to different biochemical–physiological performances of seedlings. To exclude this possibility, the seed moisture content was evaluated. The moisture content in wheat seeds was determined according to the oven method using a low constant temperature (101–105 °C) for 17 h [42]. Seed moisture content (SMC) was estimated for unprimed seeds before germination; for primed seeds after 12 h of different priming agents’ application, as well as after application of priming followed by desiccation before germination.
2.3. Germination, Growth, and Vigor Index
After the appearance of a 2 mm long radicle, wheat seeds were considered germinated. The number of germinated seeds was constant after the third day of incubation, and the total percentage of germination (GP) was calculated [43]. The seedling weight vigor index (SWVI) and seedling length vigor index (SLVI) was evaluated according to Bojović et al. [44].
2.4. Total Protein Concentration
To evaluate total soluble protein content in wheat leaves, the bovine serum albumin (BSA) was used as a standard solution [45]. The obtained values were expressed according to the fresh mass of plant material (mg g−1 FW).
2.5. Lipid Peroxidation
The extraction and lipid peroxidation determination in wheat leaves using MDA as a marker was carried out according to Jakovljević et al. [46]. The absorbance of the supernatant was determined at 532 and 600 nm, and content of MDA was expressed as nM g−1 FW.
2.6. Extraction and Analysis of Enzymes
The antioxidant enzymes were extracted from wheat leaves according to Jakovljević et al. [47]. Superoxide-dismutase (SOD) activity was determined by the spectrophotometric method according to Beauchamp and Fridovich [48]. One unit of SOD is the amount of enzyme that inhibits the NBT reduction by 50%. Catalase (CAT) activity was determined by the spectrophotometric method described in Goth [49]. One unit of CAT represents the amount of enzyme that decomposes 1 μM H2O2 per minute under assay conditions. Ascorbate peroxidase (A-POX) and guaiacol peroxidase (G-POX) activity was determined according to Jia et al. [50] while activity of pyrogallol peroxidase (P-POX) was determined according to Kukavica et al. [51]. Activity of all investigated enzymes is expressed as U mg−1 of protein.
2.7. Preparation of Plant Extracts
The aboveground part of the wheat seedlings was dried for 7 days (room temperature, air humidity 55–60%). The dried aboveground parts of the wheat seedlings were crushed to obtain plant powder. For the preparation of plant extracts, dry plant powder (1 g) was extracted with methanol (20 mL) (98%); after 48 h, extracted samples were filtered and then evaporated. The dry extract dissolved in methanol at a concentration of 1 mg/mL was used for further analysis.
2.8. Total Phenolic Compounds
Total phenolic content was estimated by the spectrophotometric method according to Singleton et al. [52]. A total of 0.5 mL of plant extract, 2.5 mL Folin–Ciocalteu reagent, and 2 mL NaHCO3 were incubated for 15 min at 45 °C, after which the absorbance was determined at 765 nm. The results were expressed as gallic acid equivalents (mg of GA g−1 of extract).
2.9. Total Flavonoids
The mixture of 1 mL of plant extract and 1 mL of AlCl3 dissolved in methanol was incubated (1 h at room temperature) and then the absorbance was determined at 415 nm [53]. The results were expressed as rutin equivalents (mg of RU g−1 of extract).
2.10. Total Antioxidant Activity
Serial dilutions of 1,1-diphenyl-2-picrylhydrazyl radical (DPPH) with methanol were made from 1 mg/mL plant extracts to obtain concentrations from 500 to 0.97 µg mL−1. The diluted solutions were mixed with the DPPH [54]. After incubation (30 min, room temperature, dark), the absorbance was determined (at 517 nm). The results were expressed as a percentage of DPPH radical inhibition, based on differences in the absorbance of plant extract samples and control samples (samples without plant extract solutions).
2.11. Quantification of Phenolic Acids and Flavonoids
2.11.1. Chemicals
Gallic acid, 4-hydroxy benzoic acid (4-HBA), 3.4-dihydroxy benzoic acid (3.4-DHBA), 3.5-dihydroxy benzoic acid (3.5-DHBA), catechin, chlorogenic acid, syringic acid, caffeic acid, epicatechin, p-coumaric acid, sinapic acid, ferulic acid, naringin, rutin, myricetin, quercetin, naringenin, apigenin and crysin were from Sigma-Aldrich (Steineheim, Germany). Formic acid and acetonitrile (HPLC grade) were purchased from J.T. Baker (Gliwice, Poland). Water was obtained from a Millipore Milli-Q system (Watford, UK).
2.11.2. Instruments
To determine the individual phenolic acids and flavonoids, the HPLC system (Shimadzu, Kyoto, Japan) consisted of degasser DGU-20A3, analytical pumps LC-20AT, manual injector 7125 and SPD M20A diode array detector, and CBM-20A system controller. A reversed-phase Hypersil GOLD aQ (150 × 4.6 mm, particle size 5 μm) column (Bellefonte, PA, USA) was used for separation. The chromatographic data were processed using LC Solution computer software (Shimadzu).
2.12. Statistical Analysis
The results were obtained with the SPSS for Windows (version 21) and expressed as the mean value of at least five repetitions ± standard error (SE) and presented in tables or figures. Data were analyzed using the ANOVA test (p ≤ 0.05). To determine the existence of differences between individual treatments, Tukey’s tests were used (p ≤ 0.05).
3. Results
3.1. Seed Moisture Content
Seed moisture content significantly affects physiological processes in seed. Based on this, seed moisture content was measured to confirm that the selected priming agents did not cause different seed moisture content after priming. The wheat seed moisture content measured immediately after priming with different priming agents ranged from 32.34 to 33.33%, without significant differences among the applied agents (Figure 1). Additionally, it was shown that the primed seed moisture content measured after desiccation was similar, while the control unprimed seed moisture content was significantly lower and amounted to 9.5%.
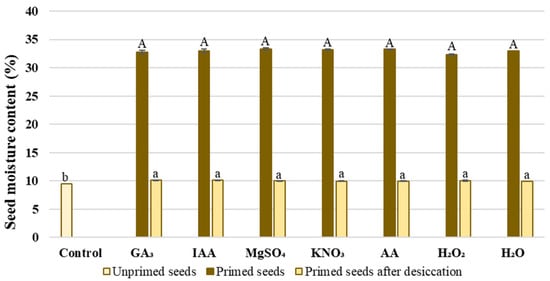
Figure 1.
The moisture content in unprimed wheat seeds, in primed seeds immediately after priming and in primed seeds after desiccation before germination; the values represent the means of five replicates ± standard error. Different small letters above bars indicates significant differences between unprimed seeds and primed seeds after desiccation. Same capital letters indicate significant differences after application of different priming agents using Tukey test (p ≤ 0.05).
3.2. Germination and Growth Characteristic
The low temperature did not affect the germination of wheat seeds since high germination percentage were recorded (>98%) in the shortest period (within 3 days) without significant differences between primed and unprimed seeds (Figure 2).
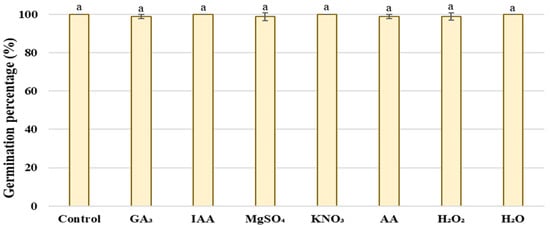
Figure 2.
Effect of different priming treatments on germination percentage (GP) (%) of wheat. Same letters above bars indicates that differences were not significant (p > 0.05) using Tukey test.
The application of priming had a significant impact on the monitored growth characteristics (Table 1). In general, applied agents had a stimulating effect on shoot and root growth compared to the unprimed seeds. The priming treatment with GA3 stands out as the treatment with the dominant effect (15.90 cm). The highest value for root length was recorded in the treatment with AA (10.71 cm) while priming treatment with KNO3 had the most prominent effect on fresh (146.00 mg) and dry mass (25.50 mg). Observed through the values obtained for SLVI (Table 1), the priming treatment with GA3 stands out as the treatment with the most prominent impact (SLVI = 2572.13). In the case of the seedling weight vigor index (Table 1), the most favorable impact was recorded after treatment with KNO3 (SLWI = 14.60).

Table 1.
Effect of different priming treatments on shoot and root length (cm), fresh and dry weight (mg), seedling weight vigor index (SWVI), and seedling length vigor index (SWVI) of wheat.
3.3. Total Soluble Proteins
In the conditions of low-temperature stress, in seedlings that were previously primed, a significant effect on the synthesis of total soluble proteins was determined compared to unprimed seedlings (Table 2). The measured protein concentrations in the leaves of wheat seedlings ranged from 98.51 to 172.96 mg g−1 FW, and the best effects were achieved after treatment with MgSO4 and AA.
Table 2.
Effect of different priming treatments on total soluble protein concentration (mg g−1 FW) and malondialdehyde (MDA) content (nM g−1 FW) in leaves of wheat seedlings.
3.4. Malondialdehyde Content (MDA)
The accumulation of MDA indicates damage of cells and formation of ROS under stressful conditions. In this regard, lower values of MDA content under stressful conditions may represent the tolerance capacity of plants. Compared to the control, the application of all priming agents significantly reduced MDA content and increased cell membrane stability (Table 2) with the most prominent effect achieved by hormopriming treatments (GA3 and IAA).
3.5. Antioxidant Enzymes Activity
The pre-sowing effect on the antioxidant enzyme activity depended on the applied priming agents (Figure 3 and Figure 4). Hormopriming caused a decrease in SOD and A-POX activity, and a significant increase in P-POX, G-POX, and CAT. SOD activities remained unaffected after seed halopriming with KNO3 compared to the control, while a stimulatory effect of this treatment on the activity of G-POX, P-POX, and CAT was noticed. Halopriming treatments differed in their effectiveness, since MgSO4 had a positive effect on CAT, while all other tested enzymes showed lower activity compared to the control. Osmopriming with H2O2 caused a significant decrease in SOD, A-POX, and CAT activity, while AA caused a significant decrease in SOD, A-POX, and G-POX. Hydropriming caused an increase in CAT activity.
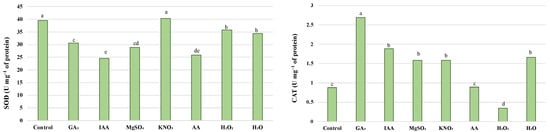
Figure 3.
Activity of SOD and CAT in leaves of wheat seedlings after different priming treatments. Different letters above bars indicates that differences were not significant (p > 0.05) using Tukey.
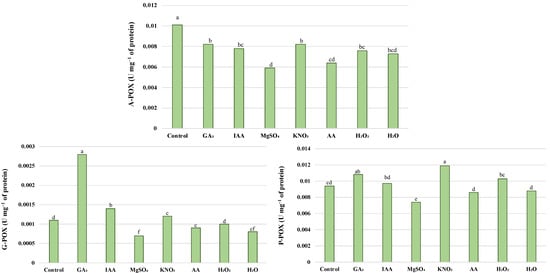
Figure 4.
Activity of A-POX, G-POX, and P-POX in leaves of wheat seedlings after different priming treatments. Different letters above bars indicates that differences were not significant (p > 0.05) using Tukey.
3.6. Total Phenolic Content and Concentration of Flavonoids
The measured values for phenolic compounds (Table 3) ranged from 19.28 to 22.50 mg of GA g−1 of extract. The highest values were recorded after treatment with KNO3 (22.50 mg of GA g−1 of extract) and AA (21.17 mg of GA g−1 of extract), without significant differences between treatments.
Table 3.
Total phenolic content (mg of GA g−1 of extract) and concentration of flavonoids (mg of RU g−1 of extract) after different priming treatments in the aboveground part of wheat seedlings.
The measured concentrations of the flavonoid concentration ranged from 25.39 to 26.88 mg of RU g−1 of extract, whereby all priming treatments significantly increased the concentration of flavonoids. The most favorable effect was after treatment with H2O2 and AA.
3.7. Antioxidant Activity
The neutralization of free DPPH radicals by plant extracts, depending on the applied treatments, is shown in Table 4. In general, wheat seedlings, obtained from the primed seed, had a higher antioxidant capacity compared to the unprimed seeds, whose activity directly depended on the concentration of the extract. The most significant influence on the inhibition of free DPPH radicals was recorded in plant extracts obtained from seeds primed with KNO3, followed by plants obtained from seeds primed with H2O and AA.
Table 4.
DPPH inhibition (%) in the aboveground part of wheat seedlings after different priming treatments.
3.8. Concentration of Phenolic Acids and Flavonoids
HPLC analysis confirmed that the application of the priming method caused significant changes in the concentration of both individual phenolic acids and flavonoids in wheat leaves (Table 5). In seedlings grown from unprimed seeds, the most abundant among phenolic acids were ferulic acid (7.06 μg of g−1 of dry extract), catechin (6.80 μg of g−1 of dry extract), and caffeic acid (1.11 μg of g−1 of dry extract). The concentration of most abundant phenolic acid in control (particularly ferulic acid and caffeic acid) decreased when plants were grown from primed seeds; however, synthesis of other phenolic acids increased, and the particular response depended on the applied priming agent. In general, except for ferulic and caffeic acid, seed priming caused an increase in the concentration of all detected phenolic acids under low-temperature conditions. Most significant effects were achieved for gallic acid, 4-HBA, and chlorogenic acid after hormopriming with GA3, and for gallic acid, sinapic acid, and catechin after hydropriming. In the case of flavonoids, in control plants, naringin (1.72 μg of g−1 of dry extract) and rutin (1.20 μg of g−1 of dry extract) were the most abundant. The stimulatory effect of the seed priming on flavonoid synthesis was recorded since an increase in the concentration of all detected flavonoids under low-temperature conditions can be seen compared to the control. After seed priming with GA3 concentration of naringin and quercetin was four-times higher, while hydropriming caused a 2.5-times increased synthesis of rutin.
Table 5.
The qualitative and quantitative composition of phenolic acids and flavonoids (μg of g−1 of dry extract) of wheat seedlings after different priming treatments.
4. Discussion
Stress caused by low temperatures seriously affects the distribution of species in nature, plant life cycle, as well as agricultural productivity [55,56]. The resistance of plants to temperature stress (low or high temperature), depends on plant species strength, and duration of stress [57]. Fluctuations of temperature can alter cellular homeostasis negatively affecting biochemical and physiological processes [58]. Suboptimal temperatures are the main limiting factor leading to crop reduction or complete failure [59] and the serious negative impact of low temperatures on wheat production was recorded [60]. In recent years, the priming method has been an effective technique for increasing crop tolerance to temperature stresses [61,62,63], which is achieved by modulating metabolic activities before germination, such as genetic and structural repair/protection, increased protein accumulation, activation of osmotic enzymes and activation of antioxidant capacity [39,64].
At the sowing stage, low-temperature stress leads directly to wheat crop failure by inhibiting/reducing germination and plant growth. Indirect low-temperature stress reduces plant density in the field [65]. Furthermore, temperature stress, caused by low temperature, reduces root length and biomass, thereby inhibiting normal nutrient and water uptake, resulting in a reduction in aboveground mass [61]. Similarly, an inhibitory effect on the reduction of size and area of leaves, as well as on shoot biomass was recorded [66]. In the reproductive phase, low temperature leads to significant functional and structural deformities, exemplified by a reduction in the number of ears, grains per ear, and ultimately yield [67]. The growth, organogenesis, and metabolic reactions in plants are also inhibited by low temperature, therefore increasing crop tolerance to cold is considered a key solution to increase optimization of their production and nutritional quality [68]. To the best of our knowledge, our study is the first report on the germination of a tested wheat cultivar (T. aestivum L. cv. Belija) under low-temperature conditions. Based on the obtained data, the germination of this cultivar can be considered independent of low temperature. Although there are no differences in germination, priming significantly contributed to growth as shown by SLVI and SWVI, so the application of GA3 and KNO3 as priming can contribute to the morphology of the tested wheat cultivar under conditions of temperature stress. GA3, as a growth regulator, induces various physiological responses in plants, thereby stimulating the germination and growth [69]. Application of GA3 induces plant development by increasing the content of amino acids in the embryo as well as by stimulating hydrolytic enzymes for breaking down starch necessary for embryo development [70]. In addition, gibberellic acid has a dynamic potential in increasing crop tolerance to abiotic stresses [71,72]. The beneficial effects of KNO3 as a priming agent may be associated with increased imbibition, stimulation of cell division and elongation, synthesis and/or repair of genetic material, the activation of enzymes for mobilization, including amylases, dehydrogenases, and phosphatases [73].
Under temperature stress, plants produce ROS which consequently leads to the creation of oxidative stress [74]. Oxidative stress leads to the oxidation of macromolecules such as proteins, lipids, and nucleic acids, thus causing not only cell damage but even complete cell destruction (death) [75]. The resulting changes are highly toxic but can be rapidly detoxified by various plant cellular mechanisms [76]. Seed priming can stimulate plants to eliminate oxidative stress, whose regulatory role is related to the neutralization of ROS, improvement of antioxidant defense mechanisms, accumulation of metabolites such as soluble proteins, or increased expression of genes responsible for the secondary metabolism [77,78]. In our study, a significant accumulation of proteins under low-temperature conditions was observed after the priming of wheat seeds, compared to unprimed. As a defense strategy, a high concentration of these proteins may enhance the ability of cells to retain water and can provide repair and structural protection of cell membranes against damage caused by abiotic stress [79]. A significant stimulating effect of priming treatment on the content of total soluble proteins under stressful conditions was recorded by Ashraf et al. [80]. In addition to higher protein content in primed wheat seedlings, seedlings obtained from primed seeds were also reported to produce lower MDA content. This is probably due to reduced free radical damage and greater membrane protection, thus preventing unsaturated fatty acid damage and electrolyte leakage, which is in agreement with Moori et al. [81]. According to Nawaz et al. [82], the lower MDA concentration can be attributed to the reorganization of cell membranes that improves membrane repair and antioxidant systems after priming, which synergistically eliminates oxidative damage. Superoxide dismutase rapidly catalyzes the dismutation of superoxide radicals into hydrogen peroxide, later converted into H2O by the activity of catalase and peroxidases [83]. In general, higher activities of antioxidant enzymes and reduced levels of MDA in membranes of primed wheat seedlings indicate the vital role of priming in improving antioxidant capacity under low-temperature conditions. Taking all priming agents into account, the dominant protective effect is achieved with KNO3 which unaffected SOD and stimulated the activity of G-POX, P-POX, and CAT. This can indicate the significant role of KNO3 as a pre-sowing treatment in improving the defense potential of wheat seedlings under low temperatures. Zhang et al. [84] suggest that the implementation of priming promotes expression of genes and achieves a stimulatory plant response to a given stress. Similarly, Liu et al. [85] showed that halopriming can improve wheat cold tolerance through the regulation of ROS metabolism, effectively maintaining membrane integrity under cold stress. This is in agreement with previous studies where it was found that the application of priming reduced the harmful effects of oxidative stress in wheat by increasing the activity of antioxidant enzymes [23,86]. Higher activities of antioxidant enzymes in primed seedlings were recorded earlier under different abiotic stress conditions [72,87,88].
Temperature stress leads to the increased production of secondary metabolites [89] and the higher content of phenolic compounds can be regarded as an adaptive mechanism in plant protection from oxidative damage, which consequently increases stress tolerance [90]. Phenolic compounds, including flavonoids, have a significant function in the elimination of ROS and plant defense responses to environmental limitations [91,92]. In this study, the application of pre-sowing treatments in the form of priming, low-temperature stress, and their interaction were significant for increasing the content of phenolics and flavonoids, and for antioxidant activity. Priming with KNO3, H2O, and AA increased the total phenolic content and concentration of flavonoids, which consequently improved the antioxidant capacity to remove free DPPH radicals in seedlings treated with these agents. Ashraf et al. [93] confirmed that priming can increase the concentration of phenolic compounds that are significant initiators of antioxidant potential. Similar notices were recorded by Hatami et al. [94].
The profile of wheat phenolic compounds was previously presented by Wang et al. [95] and Gupta et al. [3]. Frequently identified phenolic acids were syringic, p-hydroxybenzoic, ρ-coumaric, vanillic, gallic, ferulic, caffeic, and sinapic acids. Frequently identified flavonoids were quercetin, apigenin, luteolin, tricin, and chrysoeriol. A similar compound profile was identified in our study. Kowalska et al. [96] confirmed that phenolic compounds present in wheat show great variability in concentration depending on environmental factors. Exposure to low temperatures affects a wide range of phenolics in wheat seedlings. According to Ma et al. [97], stimulation of the synthesis of phenolic acids can be one of the key factors regarding the resistance of wheat. An increase in the phenolic acid content of wheat grain during temperature stress was observed by Shamloo et al. [98]. HPLC results of our study suggest that the priming method had a prominent effect on the production of all detected phenolic compounds (except for ferulic acid) under low-temperature stress compared to seedlings obtained from unprimed seeds, with the dominant effect observed after priming with GA3 and H2O. Considering the importance of phenolic acids in the abiotic stress mitigation [99], it could be presumed that seed priming can reduce the negative effects of cold stress on the biochemistry and physiology of wheat seedlings through the induction of phenolic acid and flavonoid synthesis (including gallic acid, chlorogenic acid, 4-HBA, catechin, epicatechin, caffeic acid, p-coumaric acid, sinapic acid, naringin, rutin, and quercetin). Similar observations were recorded by Mnafgui et al. [100] and Mahmood et al. [101]. Bearing in mind that phenolic acids and flavonoids are of great importance in the prevention or treatment of various diseases due to their antioxidant, antiangiogenic, antiproliferative, anticarcinogenic, and proapoptotic properties [102], the application of priming method on wheat seeds for the production of bioactive secondary metabolites can have potential practical importance.
5. Conclusions
It is shown that the implementation of priming agents can be an effective pre-sowing treatment for wheat plants to improve their ability to cope with adverse effects caused by low temperatures. In short, the application of priming significantly stimulated the growth of wheat seedlings. The accumulation of soluble proteins increased, and the MDA content decreased compared to seedlings obtained from unprimed seeds. The results confirmed that seed priming strengthens the antioxidant capacity of wheat under conditions of low temperature through the activity of antioxidant enzymes, the secondary metabolites’ content (phenolic acids and flavonoids), and total antioxidant capacity. Overall, this study highlights the potential implication of priming with KNO3 and GA3 for enhancing the defense mechanism of wheat seedlings under low-temperature conditions. Future research should consist of field experiments in multiple years and locations to further verify the effect of priming under limited environmental conditions.
Author Contributions
Conceptualization, B.B. and M.K.; methodology, D.J., M.S., B.B., M.K. and A.Ć.; software, M.K. and D.J.; validation, B.B. and D.J.; formal analysis, M.K., D.J. and A.Ć.; investigation, M.K.; data curation, M.K.; writing—original draft preparation, M.K. and D.J.; writing—review and editing, D.J., M.K., B.B. and M.S.; visualization, M.K. and D.J.; supervision, B.B., D.J. and M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This investigation was funded by the Ministry of Education, Science and Technological Development of the Republic of Serbia (Grant No: 451-03-68/2022-14/ 200122).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kuźniar, A.; Włodarczyk, K.; Grządziel, J.; Goraj, W.; Gałązka, A.; Wolińska, A. Culture-independent analysis of an endophytic core microbiome in two species of wheat: Triticum aestivum L. (cv.‘Hondia’) and the first report of microbiota in Triticum spelta L. (cv.‘Rokosz’). Syst. Appl. Microbiol. 2020, 43, 126025. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Ma, Z.; Boken, V.K.; Zeng, H.; Shang, J.; Igor, S.; Wang, J.; Yan, N. Regional differences in the performance of drought mitigation measures in 12 major wheat-growing regions of the world. Agric. Water Manag. 2022, 273, 107888. [Google Scholar] [CrossRef]
- Gupta, R.; Meghwal, M.; Prabhakar, P.K. Bioactive compounds of pigmented wheat (Triticum aestivum): Potential benefits in human health. Trends Food Sci. Technol. 2021, 110, 240–252. [Google Scholar] [CrossRef]
- Balli, D.; Cecchi, L.; Pieraccini, G.; Innocenti, M.; Benedettelli, S.; Mulinacci, N. What’s new on total phenols and γ-oryzanol derivatives in wheat? A comparison between modern and ancient varieties. J Food Compost Anal. 2022, 109, 104453. [Google Scholar] [CrossRef]
- Harkness, C.; Semenov, M.A.; Areal, F.; Senapati, N.; Trnka, M.; Balek, J.; Bishop, J. Adverse weather conditions for UK wheat production under climate change. Agric. For. Meteorol. 2020, 282, 107862. [Google Scholar] [CrossRef]
- Deryng, D.; Conway, D.; Ramankutty, N.; Price, J.; Warren, R. Global crop yield response to extreme heat stress under multiple climate change futures. Environ. Res. Lett. 2014, 9, 034011. [Google Scholar] [CrossRef]
- Trnka, M.; Rötter, R.P.; Ruiz-Ramos, M.; Kersebaum, K.C.; Olesen, J.E.; Žalud, Z.; Semenov, M.A. Adverse weather conditions for European wheat production will become more frequent with climate change. Nat. Clim. Change. 2014, 4, 637–643. [Google Scholar] [CrossRef]
- Powell, J.P.; Reinhard, S. Measuring the effects of extreme weather events on yields. Weather Clim. Extrem. 2016, 12, 69–79. [Google Scholar] [CrossRef]
- Barlow, K.M.; Christy, B.P.; O’leary, G.J.; Riffkin, P.A.; Nuttall, J.G. Simulating the impact of extreme heat and frost events on wheat crop production: A review. Field Crops Res. 2015, 171, 109–119. [Google Scholar] [CrossRef]
- Gu, L.; Hanson, P.J.; Post, W.M.; Kaiser, D.P.; Yang, B.; Nemani, R.; Pallardy, S.G.; Meyers, T. The 2007 eastern US spring freeze: Increased cold damage in a warming world? BioScience 2008, 58, 253–262. [Google Scholar] [CrossRef]
- Holman, J.D.; Schlegel, A.J.; Thompson, C.R.; Lingenfelser, J.E. Influence of precipitation, temperature, and 56 years on winter wheat yields in western Kansas. Crop Manag. 2011, 10, 1–10. [Google Scholar] [CrossRef]
- Zheng, B.; Chapman, S.C.; Christopher, J.T.; Frederiks, T.M.; Chenu, K. Frost trends and their estimated impact on yield in the Australian wheatbelt. J. Exp. Bot. 2015, 66, 3611–3623. [Google Scholar] [CrossRef] [PubMed]
- Crimp, S.J.; Zheng, B.; Khimashia, N.; Gobbett, D.L.; Chapman, S.; Howden, M.; Nicholls, N. Recent changes in southern Australian frost occurrence: Implications for wheat production risk. Crop Pasture Sci. 2016, 67, 801–811. [Google Scholar] [CrossRef]
- Tshewang, S.; Jessop, R.; Birchall, C. Effect of frost on triticale and wheat varieties at flowering in the north eastern Australian cereal belt. Cereal Res. Commun. 2017, 45, 655–664. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, L.; Asseng, S.; Xia, Y.; Tang, L.; Liu, B.; Cao, W.; Zhu, Y. Estimating spring frost and its impact on yield across winter wheat in China. Agric. For Meteorol. 2018, 260, 154–164. [Google Scholar] [CrossRef]
- Matysiak, K.; Kierzek, R.; Siatkowski, I.; Kowalska, J.; Krawczyk, R.; Miziniak, W. Effect of exogenous application of amino acids l-arginine and glycine on maize under temperature stress. Agronomy 2020, 10, 769. [Google Scholar] [CrossRef]
- Malek, M.; Ghaderi-Far, F.; Torabi, B.; Sadeghipour, H.R. Dynamics of seed dormancy and germination at high temperature stress is affected by priming and phytohormones in rapeseed (Brassica napus L.). J. Plant Physiol. 2022, 269, 153614. [Google Scholar] [CrossRef]
- Wang, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Wollenweber, B.; Jiang, D. Multiple heat priming enhances thermo-tolerance to a later high temperature stress via improving subcellular antioxidant activities in wheat seedlings. Plant Physiol. Biochem. 2014, 74, 185–192. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Planchon, S.; Renaut, J.; Vanková, R.; Prášil, I.T. Proteome analysis of cold response in spring and winter wheat (Triticum aestivum) crowns reveals similarities in stress adaptation and differences in regulatory processes between the growth habits. J. Proteome Res. 2013, 12, 4830–4845. [Google Scholar] [CrossRef]
- Li, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Jiang, D. Cold priming drives the sub-cellular antioxidant systems to protect photosynthetic electron transport against subsequent low temperature stress in winter wheat. Plant Physiol. Biochem. 2014, 82, 34–43. [Google Scholar] [CrossRef]
- Berova, M.; Zlatev, Z.; Stoeva, N. Effect of paclobutrazol on wheat seedlings under low temperature stress. Bulg. J. Plant Physiol. 2002, 28, 75–84. [Google Scholar]
- Xiao, L.; Asseng, S.; Wang, X.; Xia, J.; Zhang, P.; Liu, L.; Tang, L.; Cao, W.; Zhu, Y.; Liu, B. Simulating the effects of low-temperature stress on wheat biomass growth and yield. Agric. For Meteorol. 2022, 326, 109191. [Google Scholar] [CrossRef]
- Shah, T.; Latif, S.; Khan, H.; Munsif, F.; Nie, L. Ascorbic acid priming enhances seed germination and seedling growth of winter wheat under low temperature due to late sowing in Pakistan. Agronomy 2019, 9, 757. [Google Scholar] [CrossRef]
- Janowiak, F.; Maas, B.; Dörffling, K. Importance of abscisic acid for chilling tolerance of maize seedlings. J. Plant Physiol. 2002, 159, 635–643. [Google Scholar] [CrossRef]
- Ruelland, E.; Zachowski, A. How plants sense temperature. Environ. Exp. Bot. 2010, 69, 225–232. [Google Scholar] [CrossRef]
- Ji, H.; Xiao, L.; Xia, Y.; Song, H.; Liu, B.; Tang, L.; Cao, W.; Zhu, Y.; Liu, L. Effects of jointing and booting low temperature stresses on grain yield and yield components in wheat. Agric. For Meteorol. 2017, 243, 33–42. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, L.; Chen, X.; Li, J. Effects of Low-Temperature Stress during the Anther Differentiation Period on Winter Wheat Photosynthetic Performance and Spike-Setting Characteristics. Plants 2022, 11, 389. [Google Scholar] [CrossRef]
- Monroy, A.F.; Dryanova, A.; Malette, B.; Oren, D.H.; Ridha Farajalla, M.; Liu, W.; Danyluk, J.; Ubayasena, L.W.C.; Kane, K.; Scoles, G.J.; et al. Regulatory gene candidates and gene expression analysis of cold acclimation in winter and spring wheat. Plant Mol. Biol. 2007, 64, 409–423. [Google Scholar] [CrossRef]
- Theocharis, A.; Clément, C.; Barka, E.A. Physiological and molecular changes in plants grown at low temperatures. Planta 2012, 235, 1091–1105. [Google Scholar] [CrossRef]
- Takahashi, D.; Li, B.; Nakayama, T.; Kawamura, Y.; Uemura, M. Plant plasma membrane proteomics for improving cold tolerance. Front. Plant Sci. 2013, 4, 90. [Google Scholar] [CrossRef]
- Wang, Y.X.; Ya, H.U.; Chen, B.H.; Zhu, Y.F.; Dawuda, M.M.; Svetla, S. Physiological mechanisms of resistance to cold stress associated with 10 elite apple rootstocks. J. Integr. Agric. 2018, 17, 857–866. [Google Scholar] [CrossRef]
- Jan, N.; Andrabi, K.I. Cold resistance in plants: A mystery unresolved. Electron. J. Biotechnol. 2009, 12, 14–15. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; Garcıa, P.C.; Lopez-Lefebre, L.R.; Sánchez, E.; Romero, L. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed priming for abiotic stress tolerance: An overview. Acta Physiol. Plant. 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Paparella, S.; Araújo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef]
- Masondo, N.A.; Kulkarni, M.G.; Finnie, J.F.; Van Staden, J. Influence of biostimulants-seed-priming on Ceratotheca triloba germination and seedling growth under low temperatures, low osmotic potential and salinity stress. Ecotoxicol. Environ. Saf. 2018, 147, 43–48. [Google Scholar] [CrossRef]
- Ellouzi, H.; Oueslati, S.; Hessini, K.; Rabhi, M.; Abdelly, C. Seed-priming with H2O2 alleviates subsequent salt stress by preventing ROS production and amplifying antioxidant defense in cauliflower seeds and seedlings. Sci. Hortic. 2021, 288, 110360. [Google Scholar] [CrossRef]
- Li, C.; Jiang, D.; Wollenweber, B.; Li, Y.; Dai, T.; Cao, W. Waterlogging pretreatment during vegetative growth improves tolerance to waterlogging after anthesis in wheat. Plant Sci. 2011, 180, 672–678. [Google Scholar] [CrossRef]
- Chen, K.; Arora, R. Priming memory invokes seed stress-tolerance. Environ. Exp. Bot. 2013, 94, 33–45. [Google Scholar] [CrossRef]
- Sharma, S.N.; Maheshwari, A.; Sharma, C.; Shukla, N. Gene expression patterns regulating the seed metabolism in relation to deterioration/ageing of primed mung bean (Vigna radiata L.) seeds. Plant Physiol. Biochem. 2018, 124, 40–49. [Google Scholar] [CrossRef]
- Kanjevac, M.; Jakovljević, D.; Bojović, B. Improvement of physiological performance of selected cereals by modulating pregerminative metabolic activity in seeds. Cereal Res. Commun. 2021, 50, 831–839. [Google Scholar] [CrossRef]
- Hanson, J. Procedures for Handling Seeds in Genebanks; International Board for Plant Genetic Resources: Rome, Italy, 1985; Volume 1. [Google Scholar]
- Jakovljević, D.; Bojović, B.; Stanković, M.; Topuzović, M. Characteristics of in vitro seed germination of three basil genotypes under different nutrition. Kragujev. J. Sci. 2020, 42, 135–142. [Google Scholar] [CrossRef]
- Bojović, B.M.; Jakovljević, D.Z.; Ćurcić, S.S.; Stanković, M.S. Phytotoxic potential of common nettle (Urtica dioica L.) on germi-nation and early growth of cereals an vegetables. Allelopathy J. 2018, 43, 175–186. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Jakovljević, D.; Topuzović, M.; Stanković, M. Nutrient limitation as a tool for the induction of secondary metabolites with antioxidant activity in basil cultivars. Ind Crops Prod. 2019, 138, 111462. [Google Scholar] [CrossRef]
- Jakovljević, D.; Stanković, M.; Bojović, B.; Topuzović, M. Regulation of early growth and antioxidant defense mechanism of sweet basil seedlings in response to nutrition. Acta Physiol. Plant. 2017, 39, 1–13. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Goth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta. 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Jia, L.; Xu, W.; Li, W.; Ye, N.; Liu, R.; Shi, L.; Bin Rahman, R.A.N.M.; Fan, M.; Zhang, J. Class III peroxidases are activated in proanthocyanidin-deficient Arabidopsis thaliana seeds. Ann. Bot. 2013, 111, 839–847. [Google Scholar] [CrossRef]
- Kukavica, B.; Morina, F.; Janjic, N.; Boroja, M.; Jovanović, L.; Veljović-Jovanović, S. Effects of mixed saline and alkaline stress on the morphology and anatomy of Pisum sativum L.: The role of peroxidase and ascorbate oxidase in growth regulation. Arch. Biol. Sci. 2013, 65, 265–278. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and anti-oxidants by means of folin-ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Quettier-Deleu, C.; Gressier, B.; Vasseur, J.; Dine, T.; Brunet, C.; Luyckx, M.; Cazin, M.; Cazin, J.K.; Bailleul, F.; Trotin, F. Phenolic compounds and antioxidant activities of buckwheat (Fagopyrum esculentum Moench) hulls and flour. J. Ethnopharmacol. 2000, 72, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Stanković, M.S.; Radić, Z.S.; Blanco-Salas, J.; Vázquez-Pardo, F.M.; Ruiz-Téllez, T. Screening of selected species from Spanish flora as a source of bioactive substances. Ind. Crops Prod. 2017, 95, 493–501. [Google Scholar] [CrossRef]
- Ding, F.; Wang, C.; Zhang, S.; Wang, M. A jasmonate-responsive glutathione S-transferase gene SlGSTU24 mitigates cold-induced oxidative stress in tomato plants. Sci. Hortic. 2022, 303, 111231. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Szymańska, R.; Ślesak, I.; Orzechowska, A.; Kruk, J. Physiological and biochemical responses to high light and temperature stress in plants. Environ. Exp. Bot. 2017, 139, 165–177. [Google Scholar] [CrossRef]
- Bose, B.; Kumaria, S.; Tandon, P. Physiological insights into the role of temperature and light conditions on in vitro growth, embrane thermostability and antioxidative activity of Nardostachys jatamansi, an IUCN Red-listed critically endangered therapeutic plant. S. Afr. J. Bot. 2022, 146, 365–374. [Google Scholar] [CrossRef]
- Lukatkin, A.S.; Brazaitytė, A.; Bobinas, Č.; Duchovskis, P. Chilling injury in chilling-sensitive plants: A review. Agriculture 2012, 99, 111–124. [Google Scholar]
- Liu, L.; Ji, H.; An, J.; Shi, K.; Ma, J.; Liu, B.; Tang, L.; Cao, W.; Zhu, Y. Response of biomass accumulation in wheat to low-temperature stress at jointing and booting stages. Environ. Exp. Bot. 2019, 157, 46–57. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportuni-ties. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef]
- Li, Z.; Xu, J.; Gao, Y.; Wang, C.; Guo, G.; Luo, Y.; Huang, Y.; Hu, W.; Sheteiwy, M.S.; Guan, Y.; et al. The synergistic priming effect of exogenous salicylic acid and H2O2 on chilling tolerance enhancement during maize (Zea mays L.) seed germination. Front. Plant Sci. 2017, 8, 1153. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Khan, F.; Hussain, H.A.; Nie, L. Physiological and biochemical mechanisms of seed priming-induced chilling tolerance in rice cultivars. Front. Plant Sci. 2016, 7, 116. [Google Scholar] [CrossRef] [PubMed]
- Feghhenabi, F.; Hadi, H.; Khodaverdiloo, H.; Van Genuchten, M.T. Seed priming alleviated salinity stress during germination and emergence of wheat (Triticum aestivum L.). Agric. Water Manag. 2020, 231, 106022. [Google Scholar] [CrossRef]
- Jame, Y.W.; Cutforth, H.W. Simulating the effects of temperature and seeding depth on germination and emergence of spring wheat. Agric. For Meteorol. 2004, 124, 207–218. [Google Scholar] [CrossRef]
- Valluru, R.; Link, J.; Claupein, W. Consequences of early chilling stress in two Triticum species: Plastic responses and adaptive significance. Plant Biol. 2012, 14, 641–651. [Google Scholar] [CrossRef]
- Li, X.; Cai, J.; Liu, F.; Zhou, Q.; Dai, T.; Cao, W.; Jiang, D. Wheat plants exposed to winter warming are more susceptible to low temperature stress in the spring. Plant Growth Regul. 2015, 77, 11–19. [Google Scholar] [CrossRef]
- Fodorpataki, L.; Molnar, K.; Tompa, B.; Plugaru, S.R. Priming with vitamin U enhances cold tolerance of lettuce (Lactuca sativa L.). Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 592. [Google Scholar] [CrossRef]
- Iqbal, M.; Ashraf, M. Gibberellic acid mediated induction of salt tolerance in wheat plants: Growth, ionic partitioning, photosynthesis, yield and hormonal homeostasis. Environ. Exp. Bot. 2013, 86, 76–85. [Google Scholar] [CrossRef]
- Chauhan, A.; AbuAmarah, B.A.; Kumar, A.; Verma, J.S.; Ghramh, H.A.; Khan, K.A.; Ansari, M.J. Influence of gibberellic acid and different salt concentrations on germination percentage and physiological parameters of oat cultivars. Saudi J. Biol. Sci. 2019, 26, 1298–1304. [Google Scholar] [CrossRef]
- Iftikhar, A.; Ali, S.; Yasmeen, T.; Arif, M.S.; Zubair, M.; Rizwan, M.; Alhaithloul, H.A.S.; Alayafi, A.A.M.; Soliman, M.H. Effect of gibberellic acid on growth, photosynthesis and antioxidant defense system of wheat under zinc oxide nanoparticle stress. Environ. Pollut. 2019, 254, 113109. [Google Scholar] [CrossRef]
- Khan, M.N.; Khan, Z.; Luo, T.; Liu, J.; Rizwan, M.; Zhang, J.; Xu, Z.; Wu, H.; Hu, L. Seed priming with gibberellic acid and melatonin in rapeseed: Consequences for improving yield and seed quality under drought and non-stress conditions. Ind. Crops Prod. 2020, 156, 112850. [Google Scholar] [CrossRef]
- Ali, L.G.; Nulit, R.; Ibrahim, M.H.; Yien, C.Y.S. Efficacy of KNO3, SiO2 and SA priming for improving emergence, seedling growth and antioxidant enzymes of rice (Oryza sativa), under drought. Sci. Rep. 2021, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Phua, S.Y.; De Smet, B.; Remacle, C.; Chan, K.X.; Van Breusegem, F. Reactive oxygen species and organellar signaling. J. Exp. Bot. 2021, 72, 5807–5824. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Yu, X.; Shoaib, M.; Cheng, X.; Cui, Y.; Hussain, S.; Yan, J.; Zhou, J.; Chen, Q.; Gu, Y.; Zou, L.; et al. Role of rhizobia in promoting non-enzymatic antioxidants to mitigate nitrogen-deficiency and nickel stresses in Pongamia pinnata. Ecotoxicol. Environ. Saf. 2022, 241, 113789. [Google Scholar] [CrossRef]
- Khan, M.N.; Zhang, J.; Luo, T.; Liu, J.; Rizwan, M.; Fahad, S.; Xu, Z.; Hu, L. Seed priming with melatonin coping drought stress in rapeseed by regulating reactive oxygen species detoxification: Antioxidant defense system, osmotic adjustment, stomatal traits and chloroplast ultrastructure perseveration. Ind. Crops Prod. 2019, 140, 111597. [Google Scholar] [CrossRef]
- Zulfiqar, F. Effect of seed priming on horticultural crops. Sci. Hortic. 2021, 286, 110197. [Google Scholar] [CrossRef]
- Li, L.; Li, J.; Shen, M.; Zhang, C.; Dong, Y. Cold plasma treatment enhances oilseed rape seed germination under drought stress. Sci. Rep. 2015, 5, 1–10. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Rasheed, R.; Hussain, I.; Hafeez, A.; Adrees, M.; ur Rehman, M.Z.; Rizwan, M.; Ali, S. Effect of different seed priming agents on chromium accumulation, oxidative defense, glyoxalase system and mineral nutrition in canola (Brassica napus L.) cultivars. Environ. Pollut. 2022, 309, 119769. [Google Scholar] [CrossRef]
- Moori, S.; Ahmadi-Lahijani, M.J. Hormopriming instigates defense mechanisms in Thyme (Thymus vulgaris L.) seeds under cadmium stress. J. Appl. Res. Med. Aromat. Plants 2020, 19, 100268. [Google Scholar] [CrossRef]
- Nawaz, A.; Amjad, M.; Jahangir, M.M.; Khan, S.M.; Cui, H.; Hu, J. Induction of salt tolerance in tomato (’Lycopersicon esculentum’ Mill.) seeds through sand priming. Aust. J. Crop Sci. 2012, 6, 1199–1203. [Google Scholar]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, X.; Dong, Y.; Zhang, F.; He, Q.; Chen, J.; Zhu, S.; Zhao, T. Seed priming with melatonin improves salt tolerance in cotton through regulating photosynthesis, scavenging reactive oxygen species and coordinating with phytohormone signal pathways. Ind. Crops Prod. 2021, 169, 113671. [Google Scholar] [CrossRef]
- Liu, L.; Li, H.; Li, N.; Li, S.; Guo, J.; Li, X. Parental salt priming improves the low temperature tolerance in wheat offspring via modulating the seed proteome. Plant Sci. 2022, 324, 111428. [Google Scholar] [CrossRef] [PubMed]
- Ali, Q.; Daud, M.K.; Haider, M.Z.; Ali, S.; Rizwan, M.; Aslam, N.; Noman, A.; Iqbal, N.; Shahzad, F.; Deeba, F.; et al. Seed priming by sodium nitroprusside improves salt tolerance in wheat (Triticum aestivum L.) by enhancing physiological and biochemical parameters. Plant Physiol. Biochem. 2017, 119, 50–58. [Google Scholar] [CrossRef]
- Nasri, N.; Kaddour, R.; Mahmoudi, H.; Baatour, O.; Bouraoui, N.; Lachaâl, M. The effect of osmopriming on germination, seedling growth and phosphatase activities of lettuce under saline condition. Afr. J. Biotechnol. 2011, 10, 14366–14372. [Google Scholar] [CrossRef]
- Mohamed, A.A.; Castagna, A.; Ranieri, A.; di Toppi, L.S. Cadmium tolerance in Brassica juncea roots and shoots is affected by antioxidant status and phytochelatin biosynthesis. Plant Physiol. Biochem. 2012, 57, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Sun, C.; Jin, L.; Cai, Y.; Huang, Y.; Zheng, X.; Yu, T. L-Glutamate treatment enhances disease resistance of tomato fruit by inducing the expression of glutamate receptors and the accumulation of amino acids. Food Chem. 2019, 293, 263–270. [Google Scholar] [CrossRef]
- Lopez-Orenes, A.; Bueso, M.C.; Conesa, H.; Calderón, A.A.; Ferrer, M.A. Seasonal ionomic and metabolic changes in Aleppo pines growing on mine tailings under Mediterranean semi-arid climate. Sci. Total Environ. 2018, 637, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Robles, M.J.; Clemente, R.; Ferrer, M.A.; Calderón, A.; Bernal, M.P. Effects of ascorbic acid addition on the oxidative stress response of Oryza sativa L. plants to As (V) exposure. Plant Physiol. Biochem. 2022, 186, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, R.; Sultana, B.; Riaz, S.; Mushtaq, M.; Iqbal, M.; Nazir, A.; Atif, M.; Zafar, Z. Fortification of phenolics, antioxidant activities and biochemical attributes of radish root by plant leaf extract seed priming. Biocatal. Agric. Biotechnol. 2018, 16, 115–120. [Google Scholar] [CrossRef]
- Hatami, M.; Khanizadeh, P.; Bovand, F.; Aghaee, A. Silicon nanoparticle-mediated seed priming and Pseudomonas spp. inoculation augment growth, physiology and antioxidant metabolic status in Melissa officinalis L. plants. Ind. Crops Prod. 2021, 162, 113238. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X.; Hou, H.; Ma, X.; Sun, S.; Wang, H.; Kong, L. Metabolomics and gene expression analysis reveal the accumulation patterns of phenylpropanoids and flavonoids in different colored-grain wheats (Triticum aestivum L.). Food Res. Int. 2020, 138, 109711. [Google Scholar] [CrossRef]
- Kowalska, I.; Mołdoch, J.; Pawelec, S.; Podolska, G.; von Cossel, M.; Derycke, V.; Haesaert, G.; Lana, M.A.; da Silva Lopes, M.; Riche, A.B.; et al. Environmental and cultivar variability in composition, content and biological activity of phenolic acids and alkylresorcinols of winter wheat grains from a multi-site field trial across Europe. J. Cereal Sci. 2022, 107, 103527. [Google Scholar] [CrossRef]
- Ma, D.; Wang, C.; Feng, J.; Xu, B. Wheat grain phenolics: A review on composition, bioactivity, and influencing factors. J. Sci. Food Agric. 2021, 101, 6167–6185. [Google Scholar] [CrossRef]
- Shamloo, M.; Babawale, E.A.; Furtado, A.; Henry, R.J.; Eck, P.K.; Jones, P.J. Effects of genotype and temperature on accumulation of plant secondary metabolites in Canadian and Australian wheat grown under controlled environments. Sci. Rep. 2017, 7, 9133. [Google Scholar] [CrossRef]
- Mrid, R.B.; Benmrid, B.; Hafsa, J.; Boukcim, H.; Sobeh, M.; Yasri, A. Secondary metabolites as biostimulant and bioprotectant agents: A review. Sci. Total Environ. 2021, 777, 146204. [Google Scholar] [CrossRef]
- Mnafgui, W.; Hajlaoui, H.; Rizzo, V.; Muratore, G.; Elleuch, A. Priming with EDTA, IAA and Fe alleviates Pb toxicity in Trigonella Foneum graecum L. growth: Phytochemicals and secondary metabolites. J. Biotechnol. 2022, 356, 42–50. [Google Scholar] [CrossRef]
- Mahmood, S.; Wahid, A.; Azeem, M.; Zafar, S.; Bashir, R.; Sharif, O.; Ali, S. Tyrosine or lysine priming modulated phenolic metabolism and improved cadmium stress tolerance in mung bean (Vigna radiata L.). S. Afr. J. Bot. 2022, 149, 397–406. [Google Scholar] [CrossRef]
- Abotaleb, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Therapeutic potential of plant phenolic acids in the treatment of cancer. Biomolecules 2020, 10, 221. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).