Abstract
Metarhizium anisopliae is a broad-spectrum entomopathogenic fungus with large-scale applications in pest control; however, its slow-killing action is a limiting factor. To enhance fungal virulence, we constructed transgenic M. anisopliae strains that expressed dsRNA targeting the chitin synthase gene A in the brown planthopper Nilaparvata lugens (NlCHSA). The transcription of NlCHSA was suppressed in N. lugens infected by transgenic M. anisopliae strains. Consequently, the cumulative mortality of N. lugens caused by those transgenic strains was increased by 34.7–40.0% on day 6 after inoculation, and their median lethal times (LT50) were about 50% compared to the wild-type strain. Our results demonstrate that dsRNA expression in M. anisopliae caused RNA silencing in the host insect after infection and hence greatly enhanced the fungal virulence, highlighting a new strategy for augmentation of fungal virulence against sucking pests.
1. Introduction
The brown planthopper Nilaparvata lugens is one of the most destructive insect pests of rice through sucking and virus transmission, causing losses of billions of dollars per year [1,2]. Chemical pesticides are often used for the control of the pest but are costly and harmful to the environment and human health [3]. The entomopathogenic fungus Metarhizium anisopliae plays a crucial role in the control of insect pests in Lepidoptera, Coleoptera, Orthoptera and Hemiptera [4,5]. Recently, a strain of M. anisopliae was screened and proved highly effective to suppress the population of N. lugens [6].
RNAi has not only been widely used in the studies of gene functions but also in pest control. Currently, three major ways have been reported with potential applications: direct use of dsRNA (often mixed with nanomaterials), plant-mediated expression of dsRNA and dsRNA expressed in biocontrol microorganisms such as entomopathogenic fungi. Foliar application of dsRNA loaded onto layered double hydroxide (LDH) effectively disrupted multiple developmental stages of whitefly Bemisia tabaci in planta [7]. The SmartStax Pro® maize expressing dsRNA targeting the Snf7 of Diabrotica virgifera virgifera has been approved by the United States Department of Agriculture (USDA) and the United States Environmental Protection Agency (USEPA) and may be shortly commercialized [8]. Meanwhile, the expression of specific dsRNA in entomopathogenic fungi such as Isaria fumosorosea resulted in higher mortality of B. tabaci [9]. In order to develop RNAi-based pest control technologies, highly efficient and safe target genes are essential. In insects, there are often two chitin synthases (CHS). CHSA is responsible for chitin synthesis in the exoskeleton and trachea, while CHSB is responsible for chitin synthesis in the peritrophic membrane of the midgut [10]. Interference of CHSA with RNAi resulted in high mortality in several insects, including N. lugens [11]. Considering its safety (CHS does not exist in higher animals) and high efficiency, insect CHSA is a good RNAi target gene [12].
Theoretically, the combination of entomopathogenic fungi and RNAi technology will enhance the control effect of the entomopathogenic fungus against insect pests. In this study, we expressed the dsRNA of NlCHSA in M. anisopliae to improve its insecticidal virulence to N. lugens.
2. Materials and Methods
2.1. Fungus Strains and Insect Culture
The mycelium M. anisopliae strain CQMa 421 (strain accession number: CGMCC NO. 460) was produced on a liquid fermentation tank, and the mycelium was inoculated on rice and had a water content of 40–50% to culture conidia. Conidia and spore suspension were collected after about 15 days of culture at 28 ± 1 °C. Sporulation suspension formulation: conidium oil suspension was prepared with a mixture of water with 0.1% Tween 80. Finally, the spore suspension concentration was counted by the Petroff-hausser counter and adjusted to 8 × 1010 spore/mL.
A laboratory strain of N. lugens was collected in September 2007 from the Guangdong Academy of Agricultural Sciences in China and a colony has been reared continuously on susceptible rice varieties in the laboratory since then [13].
2.2. Vector Construction and Fungal Transformation
The construction of CHSA-RNAi was described previously [14]. A 580 bp fragment corresponding to the sense and antisense sequences of CHSA (GenBank accession no. JQ040014.1, 3829-4408 bp) was amplified by PCR using a plasmid as the template and primer pairs CHSA-F/CHSA-R and R-CHSA-F/R-CHSA-R. The antisense-TrpC terminator fragments were fused to the XbaI/BamHI (TaKaRa, Dalian, China) restriction site of the Pk2-pb vector to construct antisense-CHSA-Pk2-pb using the seamless cloning NovoRec PCR one-step directional cloning kit (Novoprotein/Sinobio, Shanghai, China). The BamHI restriction site was destroyed by primer design, and this site was added to the antisense forward primer for ligation with the gpd promoter-sense fragment. The gpd promoter-sense fragment was fused to the HindIII/BamHI restriction site and seamlessly cloned to the antisense-CHSA-Pk2-pb vector. Agrobacterium tumefaciens AGL-1 was used to transform the constructs into the M. anisopliae strain CQMa421 as previously described [15]. Transformants were selected on Czapek-Dox agar supplemented with 80 μg/mL phosphinothricin.
2.3. Verification of Transformants
Spores of RNAi transgenic strains and the wild-type strains were collected. Genomic DNA was extracted using the Fungal DNA Kit (Omega Bio-tek, Norcross, GA, USA) and confirmed by PCR using the primer pair of promoter-gpd-F/Intron-R and intron-F/TrpC-R; glyceraldehyde-3-phosphate dehydrogenase (GPD) was used as control.
For qPCR analyses, total RNA was extracted with Trizol (Thermo Fisher Scientific, Waltham, MA, USA) and cDNA was synthesized using Prime ScriptTM RT reagent Kit with gDNA Eraser (TaKaRa, Dalian, China) to remove genomic DNA. The primer pair of sense-CHSA-F/Intron-R was used to amplify the products of dsCHSA. GPD was used as control. PCR products were analyzed by electrophoresis through a 1.2% agarose gel.
2.4. Insect Bioassays
The wild-type strains and RNAi transgenic strains were assayed by topical inoculation using N. lugens 4th instar nymphs. Conidial suspensions were sprayed in 0.1% Tween 80 at concentration of 1.6 × 107 conidia/mL. Control insects were treated with 0.1% Tween 80 alone. Each treatment consisted of three replicates with 25 insects each. The entire experiment was repeated at least three times. Survival was recorded every 24 h after inoculation. LT50 (median lethal time) was used to estimate the virulence of M. anisopliae.
2.5. qPCR
Samples were collected from wildtype and RNAi strain fungal-infected nymphs of N. lunges 1–3 days post-inoculation (dpi). Additional samples were stored at –80 °C until usage. For qPCR analyses, total RNA was extracted with Trizol (Thermo Fisher Scientific) and cDNA was synthesized using a PrimeScriptTM RT reagent Kit with gDNA Eraser (TaKaRa, Dalian, China) to remove genomic DNA. A 20 μL reaction mixture including 10 μL 2 × SYBR Premix, 1 μL cDNA template, 1 μL forward primer (10 μM) and 1 μL reverse primer (10 μM) was prepared. Amplification conditions were as follows: 3 min at 95 °C, followed by 40 cycles at 95 °C for 30 s and 57 °C for 20 s. Melt curve analyses were performed to assess the specificity of amplification. The results were normalized to mRNA levels of β-actin (Table 1), and relative mRNA accumulation levels were calculated according to the delta-delta Ct method. Each experiment was repeated with three independently isolated mRNA samples (biological replicates), and each reaction was repeated three times to minimize intra-experiment variation (technical replicates).

Table 1.
Primers used in this study.
2.6. Statistical Analysis
Data were shown as means ± SE. SPSS 22.0 software for Windows (SPSS Inc., Chicago, IL, USA) was used for statistical analysis. The Student’s t-test was used for differences between groups. * means p < 0.05; ** means p < 0.01.
3. Results
3.1. Transgenic M. anisopliae Strains Expressed dsRNA of the NlCHSA
We constructed hairpin RNAs targeting the NlCHSA of N. lugens in M. anisopliae. Constructs bearing hairpin RNA structure expression were driven by the gpd promoter from M. acridum, which has been demonstrated to express hairpin RNA [14]. Due to the presence of the hairpin structure, it is difficult for PCR to amplify the complete expression element from the promoter-hairpin structure to the terminator, so we took the genome as the template and amplified the promoter of the original to the positive part of the interference fragment by use promoter-GPD-F/intron-R primer and amplified the reverse fragment to the terminator by Intron-F/TrpC-R. The results showed that transformants RNAi-strain 22 (RNAi-22) and RNAi-strain 23 (RNAi-23) of M. anisopliae could be amplified to obtain two complete elements, while the wild strain had no amplification bands (Figure 1A).
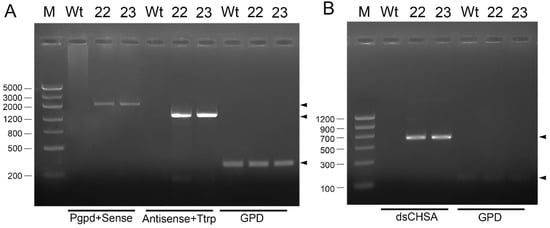
Figure 1.
Confirmation of the two transformants of Metarhizium anisopliae. (A) Amplification of gpd promoter-CHSA sense-loop and loop-antisense-Trp terminator from genomic, respectively, (B) Amplification of CHSA sense + loop from cDNA. Lane M, marker VI(A) and marker II(B); Wt, M. anisopliae wild type; 22 and 23, NlCHSA RNAi transformant lines, GPD of M. anisopliae (across intron primer) used as internal reference.
To verify the transgenic strain expressed dsRNA of the NlCHSA, we extracted its total RNA and used semi-quantitative PCR to amplify the CHSA sense chain and antisense chain, respectively, with CHSA-F/intron-R and Intron-F/R-CHSA-R. The results showed that both RNAi-22 and RNAi-23 strains expressed sense and antisense of NlCHSA, and the wild-type strain had no amplified bands. The across introns reference, GPD, has bands with cDNA amplified but not genome (Figure 1B). These results indicated that the transgenic strains 22 and 23 successfully expressed hpRNA of NlCHSA.
3.2. Transgenic M. anisopliae Strain Decreases NlCHSA Expression in N. lugens
The transgenic M. anisopliae strain 23 (RNAi-23) was used to evaluate its effect on NlCHSA expression at the end of 1 to 3 days post-inoculation (dpi). Compared with wild-type strain (WT), the RNAi-23 strain resulted in lower NlCHSA expression, decreased by 32.8% on 1 dpi (p < 0.01), 94.9% on 2 dpi (p < 0.01), and 14.8% on 3 dpi (no significant difference) (Figure 2). This result indicated that the RNAi M. anisopliae strain significantly knocked down NlCHSA expression in its host insect.
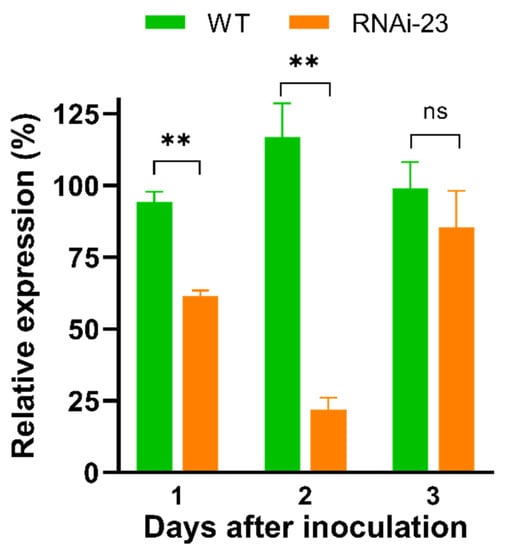
Figure 2.
Effect of transgenic M. anisopliae strain 23 on NlCHSA expression in N. lugens. Transcript levels of NlCHSA in 4th instar nymphs after infection by dsRNA expressing M. anisopliae strain 23 (RNAi-23) and wild type (WT). The data represent the mean ± SE of three independent experiments. ** p < 0.01. ns, not significant.
3.3. Transgenic M. anisopliae Strains Enhance Their Virulence to N. lugens
Among 38 transformed strains screened by bioassays, we got two highly virulent transformed strains. The mortality rate of tested insects was about 11% in the negative control (0.1% Tween 80, NC). The cumulative mortality of N. lugens (4th instars) caused by the wild-type M. anisopliae strain (WT) was 53.3% on 6 dpi, while the rate by RNAi-22 and RNAi-23 strains was 88.0% and 93.3%, respectively (Figure 3A). Infected with 1.6 × 107 conidia/mL M. anisopliae, the LT50 of N. lugens by the wild type fungus was 5.0 days, while that by the two RNAi strains was 2.5 and 2.3 days, respectively (Figure 3B).
Figure 3.
Expression of dsRNA in M. anisopliae targeting NlCHSA increases its virulence to N. lugens. (A) Survival curves of N. lugens after infection by dsRNA expressing M. anisopliae strains 22 and 23 (RNAi-22 and RNAi-23) as well as wild type (WT). N. lugens 4th nymphs were sprayed with M. anisopliae suspended in 0.1% Tween 80 at concentration of 1.6 × 107 conidia/mL. Negative control (NC) insects were sprayed with 0.1% Tween 80 alone. The data represent the mean of three independent experiments. (B) Median lethal time (LT50) of M. anisopliae WT, RNAi-22 and RNAi-23 strains against N. lugens. Probit survival analyses were used to calculate LT50. ** means p < 0.01.
4. Discussion
Our data showed that the M. anisopliae strains expressing dsRNA targeting the NlCHSA in the brown planthopper effectively knocked down NlCHSA expression in the host insect, N. lugens, and enhanced their virulence to the insect pest. Compared to the wild type M. anisopliae strain, the mortality rates of N. lugens caused by both RNAi-22 and RNAi-23 strains were increased by 34.7–40.0% (Figure 3A).
The function of CHSA was conserved in insects, and this gene can be used as a target for pest control in various pests [12]. Feeding with chitosan/dsRNA-based nanoparticles of AgCHS1 and AgCHS2 in mosquito larval repressed target gene expression [16]. Feeding bacterially expressed dsRNA of CHS1 in Mythimna separata [17], Leptinotarsa decemlineata [18], and Spodoptera exigua [19] also proved to be valid; however, it is difficult to control sap-sucking pests such as the brown planthopper by spraying dsRNA since sucking insects don’t feed plant leaves. In addition, transgenic plants expressed CHS1 dsRNA (plant-mediated RNAi) in aphids [20,21] and Helicoverpa armigera [22] were also potential for pest control. In this paper, NlCHSA in N. lugens was targeted to improve the insecticidal virulence of M. anisopliae against the brown planthopper. Since the M. anisopliae strain used here is a broad-spectrum strain capable of killing multiple pests on crops such as rice, maize and wheat [23], CHSA genes of other insect pests could be also targeted to enhance its virulence to those pests.
Microorganism-mediated delivery of host dsRNA can be used for pest control. The symbiotic bacteria Rhodococcus erythrophagus expressing the dsRNA of nitrophorin and vitellogenin from Rhodococcus rhodnii resulted in pest sterility, and the gram-negative facultative symbiotic bacterial BFo2 expressed dsα-tubulin of Frankliniella occidentalis increased pest mortality of first instar larvae and adults [24]. In a previous report, fungus M. acridum strains that expressed hairpin structure dsRNA of multiple locust genes improved the insecticidal virulence against locusts [14]. Here, we combined both entomopathogenic fungus and RNAi technology and got transgenic M. anisopliae strains with higher control effectiveness against N. lugens.
5. Conclusions
The M. anisopliae strains expressing dsRNA targeting the brown planthopper NlCHSA enhanced fungal virulence to host insects by interfering with the transcription of the target gene in the pest.
Author Contributions
J.H., H.C., Y.X. and W.Z. designed the experiments. J.H., H.C. and M.H. conducted the experiments. J.H. and W.Z. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key R&D Program (2017YFD0200900) and the National Natural Science Foundation of China (31730073).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wang, Y.; Chen, J.; Zhu, Y.C.; Ma, C.; Huang, Y.; Shen, J. Susceptibility to neonicotinoids and risk of resistance development in the brown planthopper, Nilaparvata lugens (Stal) (Homoptera: Delphacidae). Pest Manag. Sci. 2008, 64, 1278–1284. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, H.; Chen, H.; Liu, Y.; He, J.; Kang, H.; Sun, Z.; Pan, G.; Wang, Q.; Hu, J.; et al. A gene cluster encoding lectin receptor kinases confers broad-spectrum and durable insect resistance in rice. Nat. Biotechnol. 2015, 33, 301–305. [Google Scholar] [CrossRef]
- Wu, J.; Ge, L.; Liu, F.; Song, Q.; Stanley, D. Pesticide-induced planthopper population resurgence in rice cropping systems. Annu. Rev. Entomol. 2020, 65, 409–429. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.W.; St Leger, R.J. Metarhizium spp., cosmopolitan insect-pathogenic fungi: Mycological aspects. Adv. Appl. Microbiol. 2004, 54, 1–70. [Google Scholar]
- Zimmermann, G. The entomopathogenic fungus Metarhizium anisopliae and its potential as a biocontrol agent. Pestic. Sci. 1993, 37, 375–379. [Google Scholar] [CrossRef]
- Tang, J.F.; Liu, X.Y.; Ding, Y.C.; Jiang, W.J.; Xie, J.Q. Evaluation of Metarhizium anisopliae for rice planthopper control and its synergy with selected insecticides. Crop Prot. 2019, 121, 132–138. [Google Scholar] [CrossRef]
- Jain, R.G.; Fletcher, S.J.; Manzie, N.; Robinson, K.E.; Li, P.; Lu, E.; Brosnan, C.A.; Xu, Z.P.; Mitter, N. Foliar application of clay-delivered RNA interference for whitefly control. Nat. Plants 2022, 8, 535–548. [Google Scholar] [CrossRef]
- Head, G.P.; Carroll, M.W.; Evans, S.P.; Rule, D.M.; Willse, A.R.; Clark, T.L.; Storer, N.P.; Flannagan, R.D.; Samuel, L.W.; Meinke, L.J. Evaluation of SmartStax and SmartStax PRO maize against western corn rootworm and northern corn rootworm: Efficacy and resistance management. Pest Manag. Sci. 2017, 73, 1883–1899. [Google Scholar] [CrossRef]
- Chen, X.; Li, L.; Hu, Q.; Zhang, B.; Wu, W.; Jin, F.; Jiang, J. Expression of dsRNA in recombinant Isaria fumosorosea strain targets the TLR7 gene in Bemisia tabaci. BMC Biotechnol. 2015, 15, 64. [Google Scholar] [CrossRef]
- Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Specht, C.A.; Tomoyasu, Y.; Lorenzen, M.D.; Kanost, M.; Beeman, R.W. The Tribolium chitin synthase genes TcCHS1 and TcCHS2 are specialized for synthesis of epidermal cuticle and midgut peritrophic matrix. Insect Mol. Biol. 2005, 14, 453–463. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, H.W.; Huang, H.J.; Xue, J.; Wu, W.J.; Bao, Y.Y.; Xu, H.J.; Zhu, Z.R.; Cheng, J.A.; Zhang, C.X. Chitin synthase 1 gene and its two alternative splicing variants from two sap-sucking insects, Nilaparvata lugens and Laodelphax striatellus (Hemiptera: Delphacidae). Insect Biochem. Mol. Biol. 2012, 42, 637–646. [Google Scholar] [CrossRef]
- Li, T.; Chen, J.; Fan, X.; Chen, W.; Zhang, W. MicroRNA and dsRNA targeting chitin synthase A reveal a great potential for pest management of the hemipteran insect Nilaparvata lugens. Pest Manag. Sci. 2017, 73, 1529–1537. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, D.; Yao, Q.; Zhang, J.; Dong, X.; Tian, H.; Chen, J.; Zhang, W. Feeding-based RNA interference of a trehalose phosphate synthase gene in the brown planthopper, Nilaparvata lugens. Insect Mol. Biol. 2010, 19, 777–786. [Google Scholar] [CrossRef]
- Hu, J.; Xia, Y. Increased virulence in the locust-specific fungal pathogen Metarhizium acridum expressing dsRNAs targeting the host F1F0-ATPase subunit genes. Pest Manag. Sci. 2019, 75, 180–186. [Google Scholar] [CrossRef]
- Fang, W.G.; Zhang, Y.; Yang, X.; Zheng, X.; Duan, H.; Li, Y.; Pei, Y. Agrobacterium tumefaciens-mediated transformation of Beauveria bassiana using an herbicide resistance gene as a selection marker. J. Invertebr. Pathol. 2004, 85, 18–24. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, J.; Zhu, K.Y. Chitosan/double-stranded RNA nanoparticle-mediated RNA interference to silence chitin synthase genes through larval feeding in the African malaria mosquito (Anopheles gambiae). Insect Mol. Biol. 2010, 19, 683–693. [Google Scholar] [CrossRef]
- Ganbaatar, O.; Cao, B.; Zhang, Y.; Bao, D.; Bao, W.; Wuriyanghan, H. Knockdown of Mythimna separata chitinase genes via bacterial expression and oral delivery of RNAi effectors. BMC Biotechnol. 2017, 17, 9. [Google Scholar] [CrossRef]
- Shi, J.F.; Mu, L.L.; Chen, X.; Guo, W.C.; Li, G.Q. RNA interference of chitin synthase genes inhibits chitin biosynthesis and affects larval performance in Leptinotarsa decemlineata (Say). Int. J. Biol. Sci. 2016, 12, 1319–1331. [Google Scholar] [CrossRef]
- Tian, H.; Peng, H.; Yao, Q.; Chen, H.; Xie, Q.; Tang, B.; Zhang, W. Developmental control of a lepidopteran pest Spodoptera exigua by ingestion of bacteria expressing dsRNA of a non-midgut gene. PLoS ONE 2009, 4, e6225. [Google Scholar] [CrossRef]
- Zhao, Y.; Sui, X.; Xu, L.; Liu, G.; Lu, L.; You, M.; Xie, C.; Li, B.; Ni, Z.; Liang, R. Plant-mediated RNAi of grain aphid CHS1 gene confers common wheat resistance against aphids. Pest Manag. Sci. 2018, 74, 2754–2760. [Google Scholar] [CrossRef]
- Ye, C.; Jiang, Y.D.; An, X.; Yang, L.; Shang, F.; Niu, J.; Wang, J.J. Effects of RNAi-based silencing of chitin synthase gene on moulting and fecundity in pea aphids (Acyrthosiphon pisum). Sci. Rep. 2019, 9, 3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Singh, N.D.; Li, L.; Zhang, X.; Daniell, H. Engineered chloroplast dsRNA silences cytochrome p450 monooxygenase, V-ATPase and chitin synthase genes in the insect gut and disrupts Helicoverpa armigera larval development and pupation. Plant Biotechnol. J. 2015, 13, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Rajula, J.; Karthi, S.; Mumba, S.; Pittarate, S.; Thungrabeab, M.; Krutmuang, P. Chapter 4—Current status and future prospects of entomopathogenic fungi: A potential source of biopesticides. In Recent Advancement in Microbial Biotechnology; De Mandal, S., Passari, A.K., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 71–98. [Google Scholar]
- Whitten, M.M.A.; Facey, P.D.; Sol, R.D.; Fernández-Martínez, L.T.; Evans, M.C.; Mitchell, J.J.; Bodger, O.G.; Dyson, P.J. Symbiont-mediated RNA interference in insects. Proc. R. Soc. B 2016, 283, 20160042. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).