Oleic Acid Protects Against Insulin Resistance by Regulating the Genes Related to the PI3K Signaling Pathway

 , , , , , ,
, , , , , ,
Abstract
1. Introduction
2. Material and Methods
2.1. Subjects
2.2. Biochemical Measurements
2.3. Adipose Tissue Sample Collection
2.4. Fatty Acid Composition of Phospholipids from Visceral Adipose Tissue
2.5. Adipocyte Culture and Fatty Acid Treatment
2.6. RNA Isolation and Quantitative RT-PCR
2.7. Statistical Analysis
3. Results
3.1. Fatty Acid Composition of VAT Phospholipids
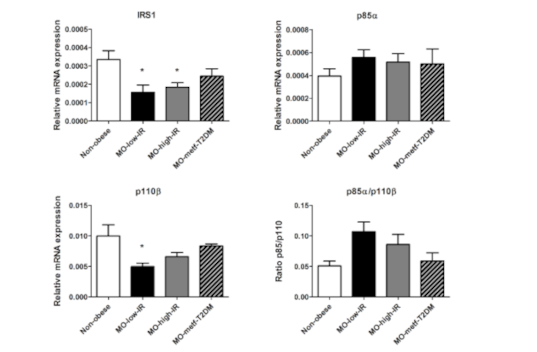
3.2. IRS1, p85α and p110β Gene Expression
3.3. Association between VAT Phospholipid Composition and mRNA Expression
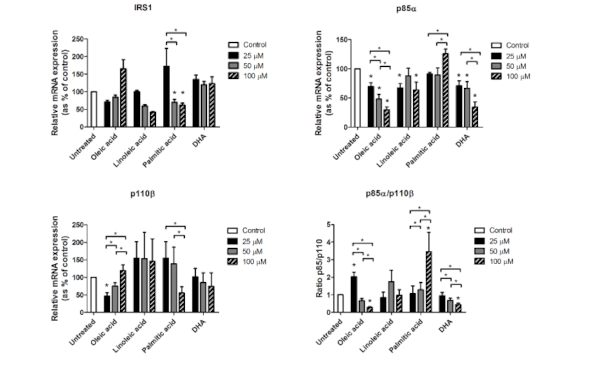
3.4. In Vitro Response to Fatty Acids
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Draznin, B. Molecular mechanisms of insulin resistance: Serine phosphorylation of insulin receptor substrate-1 and increased expression of p85alpha: The two sides of a coin. Diabetes 2006, 55, 2392–2397. [Google Scholar] [CrossRef] [PubMed]
- White, M.F. The insulin signalling system and the IRS proteins. Diabetologia 1997, 40 (Suppl. S2), S2–S17. [Google Scholar] [CrossRef] [PubMed]
- Araki, E.; Lipes, M.A.; Patti, M.E.; Bruning, J.C.; Haag, B., 3rd; Johnson, R.S.; Kahn, C.R. Alternative pathway of insulin signalling in mice with targeted disruption of the IRS-1 gene. Nature 1994, 372, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Withers, D.J.; Gutierrez, J.S.; Towery, H.; Burks, D.J.; Ren, J.M.; Previs, S.; Zhang, Y.; Bernal, D.; Pons, S.; Shulman, G.I.; et al. Disruption of IRS-2 causes type 2 diabetes in mice. Nature 1998, 391, 900–904. [Google Scholar] [CrossRef]
- Asano, T.; Fujishiro, M.; Kushiyama, A.; Nakatsu, Y.; Yoneda, M.; Kamata, H.; Sakoda, H. Role of phosphatidylinositol 3-kinase activation on insulin action and its alteration in diabetic conditions. Biol. Pharm. Bull. 2007, 30, 1610–1616. [Google Scholar] [CrossRef]
- Qiao, L.Y.; Goldberg, J.L.; Russell, J.C.; Sun, X.J. Identification of enhanced serine kinase activity in insulin resistance. J. Biol. Chem. 1999, 274, 10625–10632. [Google Scholar] [CrossRef]
- Bandyopadhyay, G.K.; Yu, J.G.; Ofrecio, J.; Olefsky, J.M. Increased p85/55/50 expression and decreased phosphotidylinositol 3-kinase activity in insulin-resistant human skeletal muscle. Diabetes 2005, 54, 2351–2359. [Google Scholar] [CrossRef]
- Ueki, K.; Algenstaedt, P.; Mauvais-Jarvis, F.; Kahn, C.R. Positive and negative regulation of phosphoinositide 3-kinase-dependent signaling pathways by three different gene products of the p85alpha regulatory subunit. Mol. Cell. Biol. 2000, 20, 8035–8046. [Google Scholar] [CrossRef]
- Soriguer, F.; Garcia-Serrano, S.; Garcia-Almeida, J.M.; Garrido-Sanchez, L.; Garcia-Arnes, J.; Tinahones, F.J.; Cardona, I.; Rivas-Marín, J.; Gallego-Perales, J.L.; García-Fuentes, E. Changes in the serum composition of free-fatty acids during an intravenous glucose tolerance test. Obesity 2009, 17, 10–15. [Google Scholar] [CrossRef]
- Boden, G.; Chen, X.; Ruiz, J.; White, J.V.; Rossetti, L. Mechanisms of fatty acid-induced inhibition of glucose uptake. J. Clin. Investig. 1994, 93, 2438–2446. [Google Scholar] [CrossRef]
- Boden, G.; Chen, X. Effects of fat on glucose uptake and utilization in patients with non-insulin-dependent diabetes. J. Clin. Investig. 1995, 96, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Yazici, D.; Sezer, H. Insulin Resistance, Obesity and Lipotoxicity. Adv. Exp. Med. Biol. 2017, 960, 277–304. [Google Scholar]
- Riccardi, G.; Giacco, R.; Rivellese, A.A. Dietary fat, insulin sensitivity and the metabolic syndrome. Clin. Nutr. 2004, 23, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Thorseng, T.; Witte, D.R.; Vistisen, D.; Borch-Johnsen, K.; Bjerregaard, P.; Jørgensen, M.E. The association between n-3 fatty acids in erythrocyte membranes and insulin resistance: The Inuit Health in Transition Study. Int. J. Circumpolar Health 2009, 68, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Gunes, O.; Tascilar, E.; Sertoglu, E.; Tas, A.; Serdar, M.A.; Kaya, G.; Kayadibi, H.; Ozcan, O. Associations between erythrocyte membrane fatty acid compositions and insulin resistance in obese adolescents. Chem. Phys. Lipids 2014, 184, 69–75. [Google Scholar] [CrossRef]
- Haag, M.; Dippenaar, N.G. Dietary fats, fatty acids and insulin resistance: Short review of a multifaceted connection. Med. Sci. Monit. 2005, 11, RA359–RA367. [Google Scholar]
- Karpe, F.; Dickmann, J.R.; Frayn, K.N. Fatty acids, obesity, and insulin resistance: Time for a reevaluation. Diabetes 2011, 60, 2441–2449. [Google Scholar] [CrossRef]
- Palomer, X.; Pizarro-Delgado, J.; Barroso, E.; Vazquez-Carrera, M. Palmitic and Oleic Acid: The Yin and Yang of Fatty Acids in Type 2 Diabetes Mellitus. Trends Endocrinol. Metab. TEM 2018, 29, 178–190. [Google Scholar] [CrossRef]
- Feng, X.T.; Wang, T.Z.; Leng, J.; Chen, Y.; Liu, J.B.; Liu, Y.; Wang, W.-J. Palmitate contributes to insulin resistance through downregulation of the Src-mediated phosphorylation of Akt in C2C12 myotubes. Biosci. Biotechnol. Biochem. 2012, 76, 1356–1361. [Google Scholar] [CrossRef]
- Ishii, M.; Maeda, A.; Tani, S.; Akagawa, M. Palmitate induces insulin resistance in human HepG2 hepatocytes by enhancing ubiquitination and proteasomal degradation of key insulin signaling molecules. Arch. Biochem. Biophys. 2015, 566, 26–35. [Google Scholar] [CrossRef]
- Bikman, B.T.; Zheng, D.; Reed, M.A.; Hickner, R.C.; Houmard, J.A.; Dohm, G.L. Lipid-induced insulin resistance is prevented in lean and obese myotubes by AICAR treatment. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R1692–R1699. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Tinahones, F.J.; Garrido-Sanchez, L.; Miranda, M.; Garcia-Almeida, J.M.; Macias-Gonzalez, M.; Ceperuelo, V.; Gluckmann, E.; Rivas-Marin, J.; Vendrell, J.; García-Fuentes, E. Obesity and insulin resistance-related changes in the expression of lipogenic and lipolytic genes in morbidly obese subjects. Obes. Surg. 2010, 20, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Serrano, S.; Moreno-Santos, I.; Garrido-Sanchez, L.; Gutierrez-Repiso, C.; Garcia-Almeida, J.M.; Garcia-Arnes, J.; Rivas-Marín, J.; Gallego-Perales, J.L.; García-Escobar, E.; Rojo-Martinez, G.; et al. Stearoyl-CoA desaturase-1 is associated with insulin resistance in morbidly obese subjects. Mol. Med. 2011, 17, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Pacheco, F.; Garcia-Serrano, S.; Garcia-Escobar, E.; Gutierrez-Repiso, C.; Garcia-Arnes, J.; Valdes, S.; Gonzalo, M.; Soriguer, F.; Moreno-Ruiz, F.J.; Rodriguez-Cañete, A.; et al. Effects of obesity/fatty acids on the expression of GPR120. Mol. Nutr. Food Res. 2014, 58, 1852–1860. [Google Scholar] [CrossRef]
- Rodriguez-Pacheco, F.; Gutierrez-Repiso, C.; Garcia-Serrano, S.; Alaminos-Castillo, M.A.; Ho-Plagaro, A.; Valdes, S.; Garcia-Arnes, J.; Gonzalo, M.; Andrade, R.J.; Moreno-Ruiz, F.J.; et al. The pro-/anti-inflammatory effects of different fatty acids on visceral adipocytes are partially mediated by GPR120. Eur. J. Nutr. 2017, 56, 1743–1752. [Google Scholar] [CrossRef]
- Gutierrez-Repiso, C.; Velasco, I.; Garcia-Escobar, E.; Garcia-Serrano, S.; Rodriguez-Pacheco, F.; Linares, F.; de Adana, M.S.R.; Rubio-Martin, E.; Garrido-Sanchez, L.; Cobos-Bravo, J.F.; et al. Does dietary iodine regulate oxidative stress and adiponectin levels in human breast milk? Antioxid. Redox Signal. 2014, 20, 847–853. [Google Scholar] [CrossRef]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int. J. Biol. Sci. 2018, 14, 1483–1496. [Google Scholar] [CrossRef]
- Barbarroja, N.; Lopez-Pedrera, R.; Mayas, M.D.; Garcia-Fuentes, E.; Garrido-Sanchez, L.; Macias-Gonzalez, M.; El Bekay, R.; Vidal-Puig, A.; Tinahones, F.J. The obese healthy paradox: Is inflammation the answer? Biochem. J. 2010, 430, 141–149. [Google Scholar] [CrossRef]
- Benito, M. Tissue specificity on insulin action and resistance: Past to recent mechanisms. Acta Physiol. 2011, 201, 297–312. [Google Scholar] [CrossRef]
- Brozinick, J.T., Jr.; Roberts, B.R.; Dohm, G.L. Defective signaling through Akt-2 and -3 but not Akt-1 in insulin-resistant human skeletal muscle: Potential role in insulin resistance. Diabetes 2003, 52, 935–941. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rondinone, C.M.; Wang, L.M.; Lonnroth, P.; Wesslau, C.; Pierce, J.H.; Smith, U. Insulin receptor substrate (IRS) 1 is reduced and IRS-2 is the main docking protein for phosphatidylinositol 3-kinase in adipocytes from subjects with non-insulin-dependent diabetes mellitus. Proc. Natl. Acad. Sci. USA 1997, 94, 4171–4175. [Google Scholar] [CrossRef] [PubMed]
- Anai, M.; Funaki, M.; Ogihara, T.; Terasaki, J.; Inukai, K.; Katagiri, H.; Fukushima, Y.; Yazaki, Y.; Kikuchi, M.; Oka, Y.; et al. Altered expression levels and impaired steps in the pathway to phosphatidylinositol 3-kinase activation via insulin receptor substrates 1 and 2 in Zucker fatty rats. Diabetes 1998, 47, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Nelson, V.L.; Jiang, Y.P.; Dickman, K.G.; Ballou, L.M.; Lin, R.Z. Adipose tissue insulin resistance due to loss of PI3K p110alpha leads to decreased energy expenditure and obesity. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1205–E1216. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Hernandez, A.; Lopez-Pastor, A.R.; Rubio-Longas, C.; Majewski, P.; Beneit, N.; Viana-Huete, V.; García-Gómez, G.; Fernandez, S.; Hribal, M.L.; Sesti, G.; et al. Specific knockout of p85alpha in brown adipose tissue induces resistance to high-fat diet-induced obesity and its metabolic complications in male mice. Mol. Metab. 2020, 31, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Cornier, M.A.; Bessesen, D.H.; Gurevich, I.; Leitner, J.W.; Draznin, B. Nutritional upregulation of p85alpha expression is an early molecular manifestation of insulin resistance. Diabetologia 2006, 49, 748–754. [Google Scholar] [CrossRef][Green Version]
- Ueki, K.; Yballe, C.M.; Brachmann, S.M.; Vicent, D.; Watt, J.M.; Kahn, C.R.; Cantley, L.C. Increased insulin sensitivity in mice lacking p85beta subunit of phosphoinositide 3-kinase. Proc. Natl. Acad. Sci. USA 2002, 99, 419–424. [Google Scholar] [CrossRef]
- Miller, M.R.; Pereira, R.I.; Langefeld, C.D.; Lorenzo, C.; Rotter, J.I.; Chen, Y.D.; Bergman, R.N.; Wagenknecht, L.E.; Norris, J.M.; Fingerlin, T.E. Levels of free fatty acids (FFA) are associated with insulin resistance but do not explain the relationship between adiposity and insulin resistance in Hispanic Americans: The IRAS Family Study. J. Clin. Endocrinol. Metab. 2012, 97, 3285–3291. [Google Scholar] [CrossRef] [PubMed]
- Piro, S.; Maniscalchi, E.T.; Monello, A.; Pandini, G.; Mascali, L.G.; Rabuazzo, A.M.; Purrello, F. Palmitate affects insulin receptor phosphorylation and intracellular insulin signal in a pancreatic alpha-cell line. Endocrinology 2010, 151, 4197–4206. [Google Scholar] [CrossRef]
- Schmitz-Peiffer, C.; Craig, D.L.; Biden, T.J. Ceramide generation is sufficient to account for the inhibition of the insulin-stimulated PKB pathway in C2C12 skeletal muscle cells pretreated with palmitate. J. Biol. Chem. 1999, 274, 24202–24210. [Google Scholar] [CrossRef]
- Rehman, K.; Haider, K.; Jabeen, K.; Akash, M.S.H. Current perspectives of oleic acid: Regulation of molecular pathways in mitochondrial and endothelial functioning against insulin resistance and diabetes. Rev. Endocr. Metab. Disord. 2020. [Google Scholar] [CrossRef] [PubMed]
- Alkhateeb, H.; Qnais, E. Preventive effect of oleate on palmitate-induced insulin resistance in skeletal muscle and its mechanism of action. J. Physiol. Biochem. 2017, 73, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Tardif, N.; Salles, J.; Landrier, J.F.; Mothe-Satney, I.; Guillet, C.; Boue-Vaysse, C.; Combaret, L.; Giraudet, C.; Patrac, V.; Bertrand-Michelh, J. Oleate-enriched diet improves insulin sensitivity and restores muscle protein synthesis in old rats. Clin. Nutr. 2011, 30, 799–806. [Google Scholar] [CrossRef]
- Salvado, L.; Coll, T.; Gomez-Foix, A.M.; Salmeron, E.; Barroso, E.; Palomer, X.; Vázquez-Carrera, M. Oleate prevents saturated-fatty-acid-induced ER stress, inflammation and insulin resistance in skeletal muscle cells through an AMPK-dependent mechanism. Diabetologia 2013, 56, 1372–1382. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, L.; Weigert, C.; Häring, H.U.; Lehmann, R. How insulin receptor substrate proteins regulate the metabolic capacity of the liver--implications for health and disease. Curr. Med. Chem. 2008, 15, 1316–1329. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, H.; Kiyomoto, H.; Nishiyama, A.; Hara, T.; Moriwaki, K.; Kaifu, K.; Ihara, G.; Fujita, Y.; Ugawa, T.; Kohno, M. Aldosterone suppresses insulin signaling via the downregulation of insulin receptor substrate-1 in vascular smooth muscle cells. Hypertension 2007, 50, 750–755. [Google Scholar] [CrossRef]
- Akalestou, E.; Genser, L.; Rutter, G.A. Glucocorticoid Metabolism in Obesity and Following Weight Loss. Front. Endocrinol. (Lausanne) 2020, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Folli, F.; Saad, M.J.; Kahn, C.R. Insulin receptor/IRS-1/PI 3-kinase signaling system in corticosteroid-induced insulin resistance. Acta Diabetol. 1996, 33, 185–192. [Google Scholar] [CrossRef]
- Weir, N.L.; Nomura, S.O.; Steffen, B.T.; Guan, W.; Karger, A.B.; Klein, R.; Klein, B.E.K.; Cotch, M.F.; Tsai, M.Y. Associations between omega-6 polyunsaturated fatty acids, hyperinsulinemia and incident diabetes by race/ethnicity: The Multi-Ethnic Study of Atherosclerosis. Clin. Nutr. 2020. [Google Scholar] [CrossRef]
- Lepretti, M.; Martucciello, S.; Burgos Aceves, M.A.; Putti, R.; Lionetti, L. Omega-3 Fatty Acids and Insulin Resistance: Focus on the Regulation of Mitochondria and Endoplasmic Reticulum Stress. Nutrients 2018, 10, 350. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F.; Ueki, K.; Fruman, D.A.; Hirshman, M.F.; Sakamoto, K.; Goodyear, L.J.; Iannacone, M.; Accili, D.; Cantley, L.C.; Kahn, C.R. Reduced expression of the murine p85alpha subunit of phosphoinositide 3-kinase improves insulin signaling and ameliorates diabetes. J. Clin. Investig. 2002, 109, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Kovalik, J.P.; Slentz, D.; Stevens, R.D.; Kraus, W.E.; Houmard, J.A.; Nicoll, J.B.; Lea-Currie, Y.R.; Everingham, K.; Kien, C.L.; Buehrer, B.M.; et al. Metabolic remodeling of human skeletal myocytes by cocultured adipocytes depends on the lipolytic state of the system. Diabetes 2011, 60, 1882–1893. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Hu, T.; Zhang, S.; Zhou, L. Associations of Different Adipose Tissue Depots with Insulin Resistance: A Systematic Review and Meta-analysis of Observational Studies. Sci. Rep. 2015, 5, 18495. [Google Scholar] [CrossRef] [PubMed]
- Cortright, R.N.; Sandhoff, K.M.; Basilio, J.L.; Berggren, J.R.; Hickner, R.C.; Hulver, M.W.; Dohm, G.L.; Houmard, J.A. Skeletal muscle fat oxidation is increased in African-American and white women after 10 days of endurance exercise training. Obesity (Silver Spring) 2006, 14, 1201–1210. [Google Scholar] [CrossRef]
- Barakat, H.; Hickner, R.C.; Privette, J.; Bower, J.; Hao, E.; Udupi, V.; Green, A.; Pories, W.; MacDonald, K. Differences in the lipolytic function of adipose tissue preparations from Black American and Caucasian women. Metabolism 2002, 51, 1514–1518. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Non-Obese | MO-Low-IR | MO-High-IR | MO-Metf-T2DM | |
|---|---|---|---|---|
| N (men/women) | 18 (6/12) | 14 (5/9) | 24 (9/15) | 9 (3/6) |
| Age (years) | 50.9 ± 16.6 | 47.8 ± 9.5 | 40.2 ± 8.4 a | 41.3 ± 12.1 |
| Weight (kg) | 74.6 ± 11.9 | 140.1 ± 29.1 a | 157.7 ± 27.2 a | 141.8 ± 22.4 a |
| BMI (kg/m2) | 27.6 ± 3.6 | 53.4 ± 6.4 a | 56.2 ± 6.6 a | 53.1 ± 6.0 a |
| Glucose (mg/dl) | 108.0 ± 24.3 | 97.0 ± 10.0 | 112.9 ± 36.5 | 169.1 ± 59.0 c |
| Cholesterol (mg/dl) | 190.9 ± 40.2 | 199.6 ± 42.9 | 196.7 ± 33.5 | 200.5 ± 28.9 |
| Triglycerides (mg/dl) | 79.4 ± 43.3 | 109.9 ± 39.7 | 175.6 ± 171.2 a | 172.0 ± 67.5 |
| Insulin (μIU/mL) | 10.8 ± 2.5 | 14.5 ± 1.8 | 29.2 ± 9.3 b | 30.7 ± 15.1 b |
| HOMA-IR | 2.9 ± 1.0 | 3.4 ± 0.5 | 8.0 ± 3.2 b | 13.7 ± 10.1 c |
| Adiponectin (μg/mL) | 20.6 ± 8.2 | 12.2 ± 4.7 a | 7.9 ± 4.1 a | 7.8 ± 4.3 a |
| Leptin (ng/mL) | 10.6 ± 3.2 | 183.2 ± 98.2 a | 151.7 ± 83.3 a | 168.2 ± 65.5 a |
| Fatty Acid | Non-Obese | MO-Low-IR | MO-High-IR | MO-Metf-TDM2 | |
|---|---|---|---|---|---|
| Lauric acid | 12:0 | 0.58 ± 0.48 | 0.11 ± 0.17 | 0.25 ± 0.38 | 0.26 ± 0.24 |
| Myristic acid | 14:0 | 1.44 ± 0.29 | 2.18 ± 1.78 | 1.28 ± 0.92 | 1.40 ± 1.10 |
| Palmitic acid | 16:0 | 22.772.8 | 22.2 ± 1.9 | 22.9 ± 3.1 | 21.7 ± 1.5 |
| Palmitoleic acid | 16:1n-7 | 2.63 ± 0.81 | 2.92 ± 1.07 | 3.01 ± 0.82 | 2.41 ± 0.49 |
| Stearic acid | 18:0 | 15.8 ± 3.9 | 12.2 ± 2.0 a | 12.8 ± 2.2 | 12.8 ± 1.2 |
| Oleic acid | 18:1n-9 | 30.4 ± 4.8 | 28.2 ± 3.5 | 27.9 ± 4.6 | 26.5 ± 5.5 |
| Vaccenic acid | 18:1n-7 | 0.94 ± 1.33 | 2.62 ± 0.55 a | 2.35 ± 1.38 a | 2.10 ± 1.04 |
| Linoleic acid | 18:2n-6 | 16.1 ± 4.5 | 17.8 ± 2.0 | 14.7 ± 3.5 | 16.4 ± 3.0 |
| Gamma-linolenic acid | 18:3n-6 | 0.03 ± 0.08 | 0.02 ± 0.06 | 0.07 ± 0.27 | 0.01 ± 0.01 |
| α-Linoleic acid | 18:3n-3 | 0.29 ± 0.32 | 0.54 ± 0.84 | 0.28 ± 0.38 | 0.55 ± 0.54 |
| Eicosanoic acid | 20:0 | 1.01 ± 0.50 | 0.54 ± 0.54 | 1.09 ± 1.06 | 1.29 ± 0.51 |
| Gondoic acid | 20:1n-9 | 0.80 ± 0.61 | 0.89 ± 0.34 | 1.12 ± 1.79 | 1.13 ± 0.58 |
| Eicosadienoic acid | 20:2n-6 | 0.47 ± 0.51 | 0.73 ± 0.59 | 0.62 ± 1.06 | 0.87 ± 0.60 |
| Dihomo-gamma-linolenic acid | 20:3n-6 | 0.73 ± 0.34 | 1.17 ± 0.57 | 1.32 ± 1.33 | 1.79 ± 0.86 |
| Arachidonic acid | 20:4n-6 | 4.18 ± 1.48 | 5.62 ± 1.39 | 6.33 ± 1.99 a | 7.24 ± 2.28 a |
| Eicosatrienoic acid | 20:3n-3 | 0.05 ± 0.09 | 0.14 ± 0.28 | 0.33 ± 0.24 | 0.18 ± 0.20 |
| Eicosapentaenoic acid | 20:5n-3 | 0.35 ± 0.29 | 0.27 ± 0.32 | 0.50 ± 0.63 | 1.04 ± 1.49 |
| Adrenic acid | 22:4n-6 | 0.37 ± 0.35 | 0.72 ± 0.55 | 1.01 ± 1.03 | 0.68 ± 0.47 |
| Docosapentaenoic acid | 22:5n-3 | 0.24 ± 0.27 | 0.23 ± 0.35 | 0.65 ± 0.86 | 0.49 ± 0.48 |
| Docosahexaenoic acid | 22:6n-3 | 0.61 ± 0.98 | 0.80 ± 0.64 | 0.77 ± 1.01 | 1.08 ± 0.74 |
| Total saturated fatty acids | 41.6 ± 5.9 | 37.2 ± 3.7 | 38.9 ± 5.0 | 37.4 ± 2.3 | |
| Total monounsaturated fatty acids | 34.7 ± 4.8 | 34.6 ± 4.0 | 34.4 ± 5.1 | 32.2 ± 5.5 | |
| n-3 polyunsaturated fatty acids | 1.54 ± 1.45 | 1.99 ± 1.28 | 2.54 ± 2.34 | 3.35 ± 3.11 | |
| n-6 polyunsaturated fatty acids | 22.0 ± 5.1 | 26.0 ± 2.1 | 24.1 ± 4.9 | 26.9 ± 3.7 | |
| Fatty Acid | IRS1 | p85α | p110β | p85/p110β | |
|---|---|---|---|---|---|
| Palmitic acid | 16:0 | r = 0.353 | r = 0.234 | r = 0.372 | r = 0.033 |
| p = 0.017 | p = 0.126 | p = 0.012 | p = 0.833 | ||
| Stearic acid | 18:0 | r = 0.103 | r = 0.484 | r = −0.179 | r = −0.152 |
| p = 0.501 | p = 0.001 | p = 0.240 | p = 0.329 | ||
| Oleic acid | 18:1n-9 | r = 0.284 | r = −0.364 | r = 0.303 | r = 0.294 |
| p = 0.059 | p = 0.015 | p = 0.043 | p = 0.056 | ||
| Linoleic acid | 18:2n-6 | r = −0.416 | r = −0.246 | r = −0.348 | r = 0.048 |
| p = 0.004 | p = 0.108 | p = 0.019 | p = 0.762 | ||
| Arachidonic acid | 20:4n-6 | r = −0.296 | r = −0.123 | r = −0.310 | r = −0.063 |
| p = 0.049 | p = 0.426 | p = 0.038 | p = 0.687 | ||
| Adrenic acid | 22:4n-6 | r = −0.305 | r = −0.139 | r = −0.171 | r = −0.083 |
| p = 0.041 | p = 0.367 | p = 0.261 | p = 0.599 | ||
| Docosahexaenoic acid | 22:6n-3 | r = 0.335 | r = −0.211 | r = 0.543 | r = 0.145 |
| p = 0.025 | p = 0.170 | p < 0.001 | p = 0.354 | ||
| Total saturated fatty acids | r = 0.221 | r = 0.446 | r = 0.044 | r = −0.047 | |
| p = 0.144 | p = 0.002 | p = 0.775 | p = 0.764 | ||
| Total unsaturated fatty acids | r = −0.221 | r = −0.446 | r = −0.044 | r = 0.047 | |
| p = 0.144 | p = 0.002 | p = 0.775 | p = 0.764 | ||
| n-6 polyunsaturated fatty acids | r = −0.587 | r = −0.266 | r = −0.497 | r = −0.062 | |
| p < 0.001 | p = 0.081 | p = 0.001 | p = 0.691 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Gómez, C.; Santiago-Fernández, C.; García-Serrano, S.; García-Escobar, E.; Gutiérrez-Repiso, C.; Rodríguez-Díaz, C.; Ho-Plágaro, A.; Martín-Reyes, F.; Garrido-Sánchez, L.; Valdés, S.; et al. Oleic Acid Protects Against Insulin Resistance by Regulating the Genes Related to the PI3K Signaling Pathway. J. Clin. Med. 2020, 9, 2615. https://doi.org/10.3390/jcm9082615
López-Gómez C, Santiago-Fernández C, García-Serrano S, García-Escobar E, Gutiérrez-Repiso C, Rodríguez-Díaz C, Ho-Plágaro A, Martín-Reyes F, Garrido-Sánchez L, Valdés S, et al. Oleic Acid Protects Against Insulin Resistance by Regulating the Genes Related to the PI3K Signaling Pathway. Journal of Clinical Medicine. 2020; 9(8):2615. https://doi.org/10.3390/jcm9082615
Chicago/Turabian StyleLópez-Gómez, Carlos, Concepción Santiago-Fernández, Sara García-Serrano, Eva García-Escobar, Carolina Gutiérrez-Repiso, Cristina Rodríguez-Díaz, Ailec Ho-Plágaro, Flores Martín-Reyes, Lourdes Garrido-Sánchez, Sergio Valdés, and et al. 2020. "Oleic Acid Protects Against Insulin Resistance by Regulating the Genes Related to the PI3K Signaling Pathway" Journal of Clinical Medicine 9, no. 8: 2615. https://doi.org/10.3390/jcm9082615
APA StyleLópez-Gómez, C., Santiago-Fernández, C., García-Serrano, S., García-Escobar, E., Gutiérrez-Repiso, C., Rodríguez-Díaz, C., Ho-Plágaro, A., Martín-Reyes, F., Garrido-Sánchez, L., Valdés, S., Rodríguez-Cañete, A., Rodríguez-Pacheco, F., & García-Fuentes, E. (2020). Oleic Acid Protects Against Insulin Resistance by Regulating the Genes Related to the PI3K Signaling Pathway. Journal of Clinical Medicine, 9(8), 2615. https://doi.org/10.3390/jcm9082615