Evidences for Expression and Location of ANGPTL8 in Human Adipose Tissue

 ,
,  , , , and
, , , and 
Abstract
1. Introduction
2. Experimental Section
2.1. Study Population and Ethical Statement
2.2. Anthropometric and Biochemical Measurements. Sample collection
2.3. Enzyme Immunoassay of ANGPTL8 in Human Plasma
2.4. Immunohistochemistry
2.5. In-Situ Hybridisation
2.6. Western Blotting
2.7. Gene Expression Analysis
2.8. Statistical Analysis
3. Results
3.1. Clinical and Biochemical Characteristics of Non-Obese and Obese Subjects
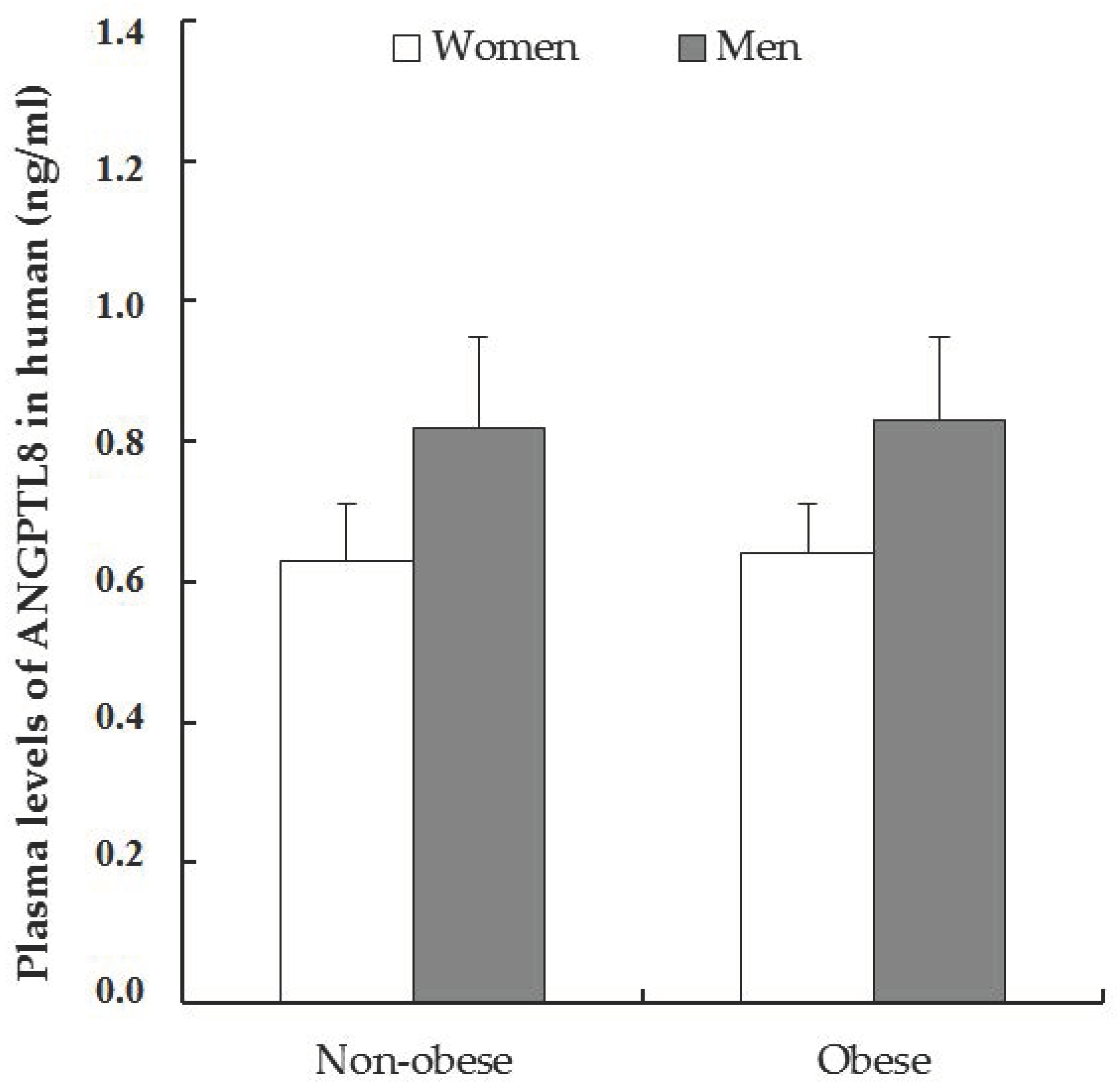
3.2. Variations in Plasma Levels of ANGPTL8 in Non-Obese and Obese Patients
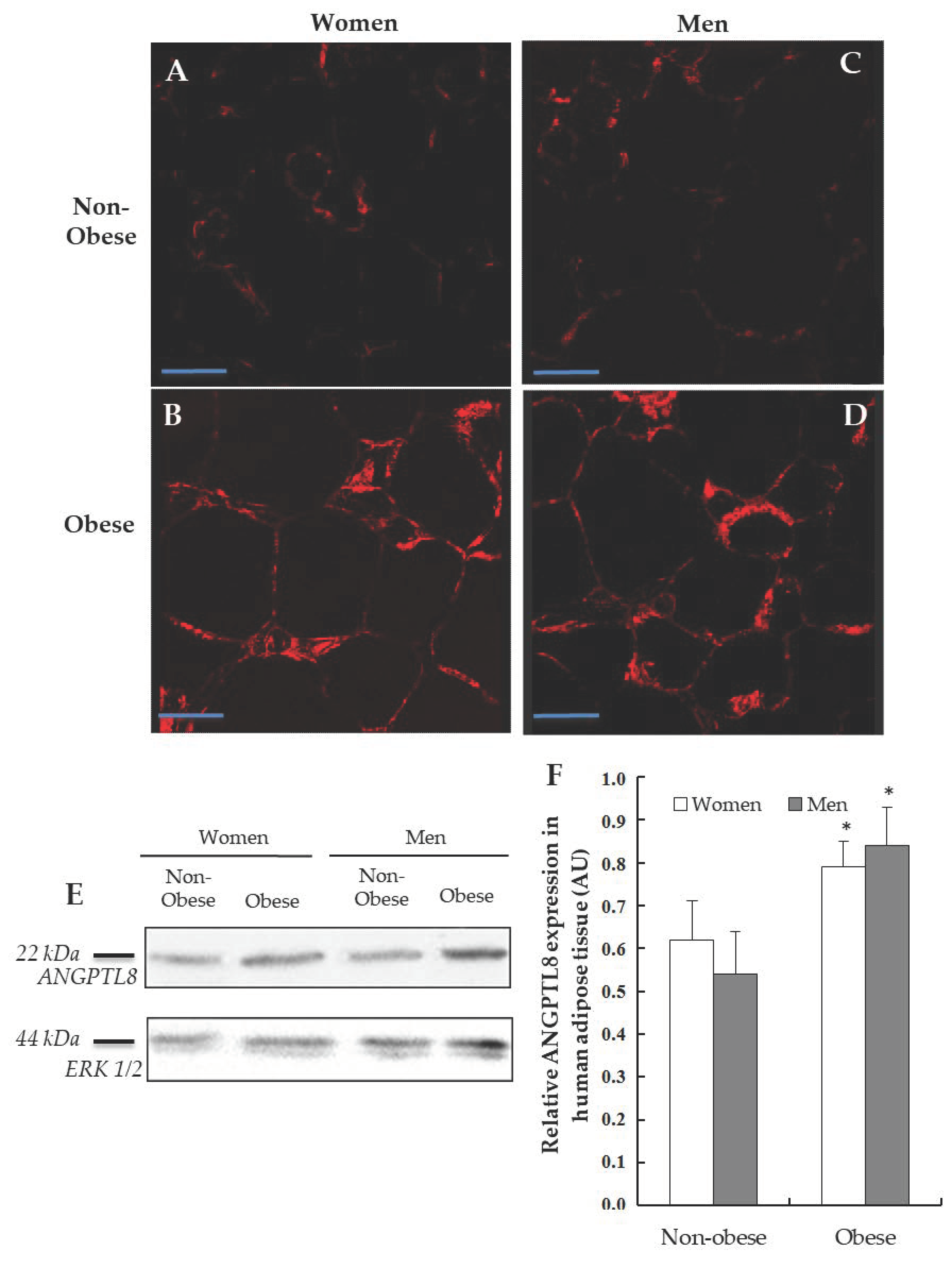
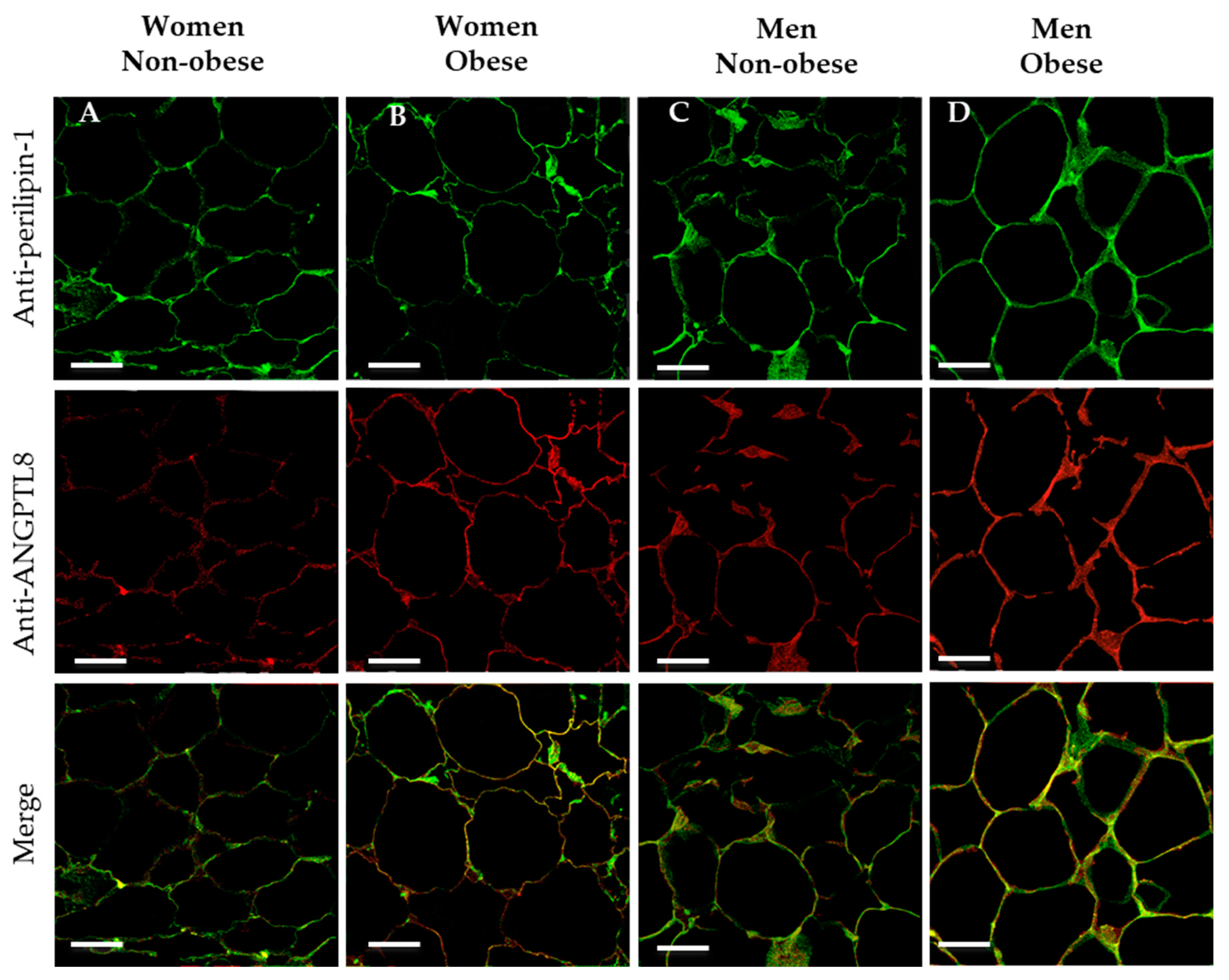
3.3. Detection of ANGPTL8 in Human Adipose Tissue
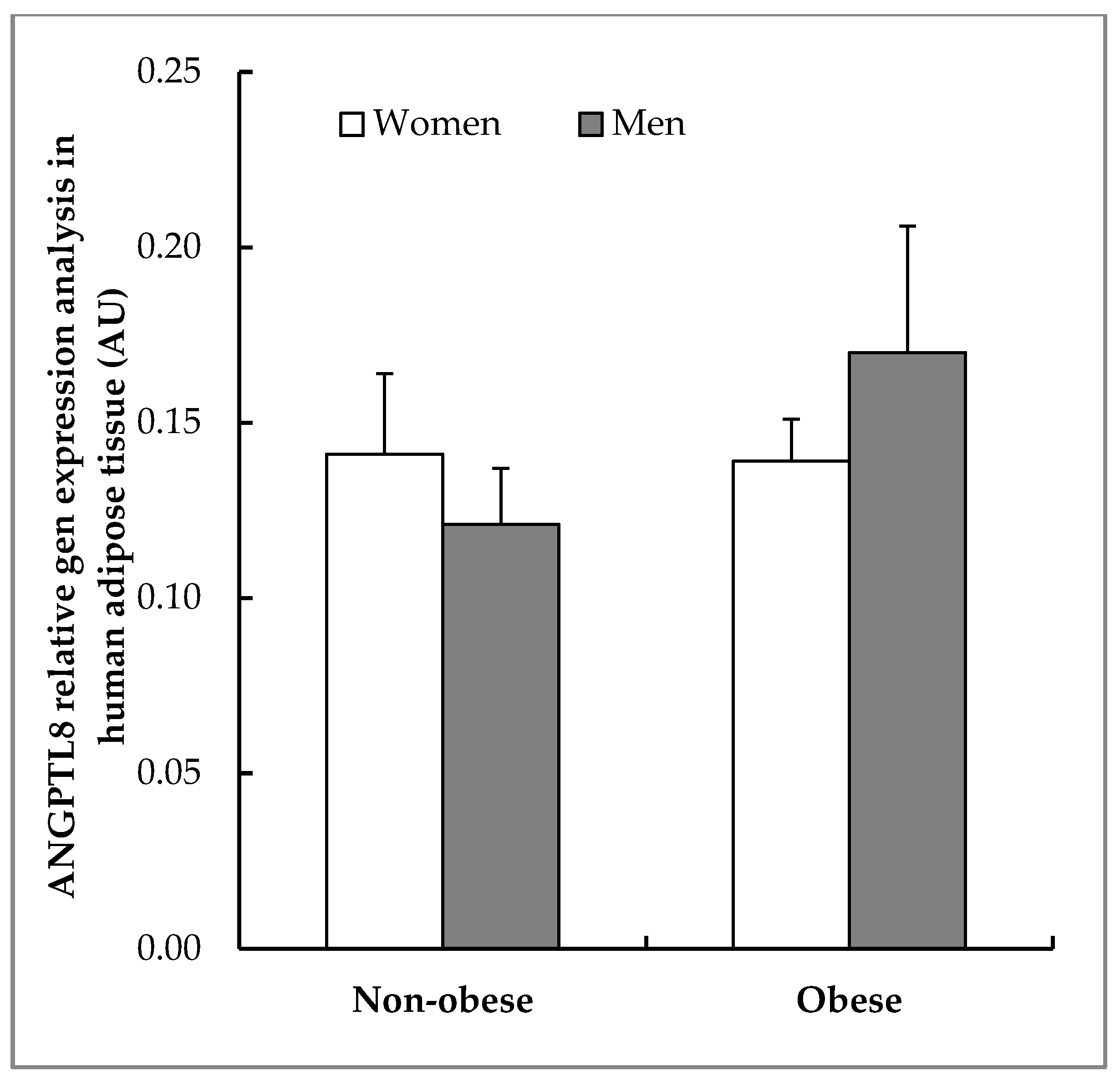
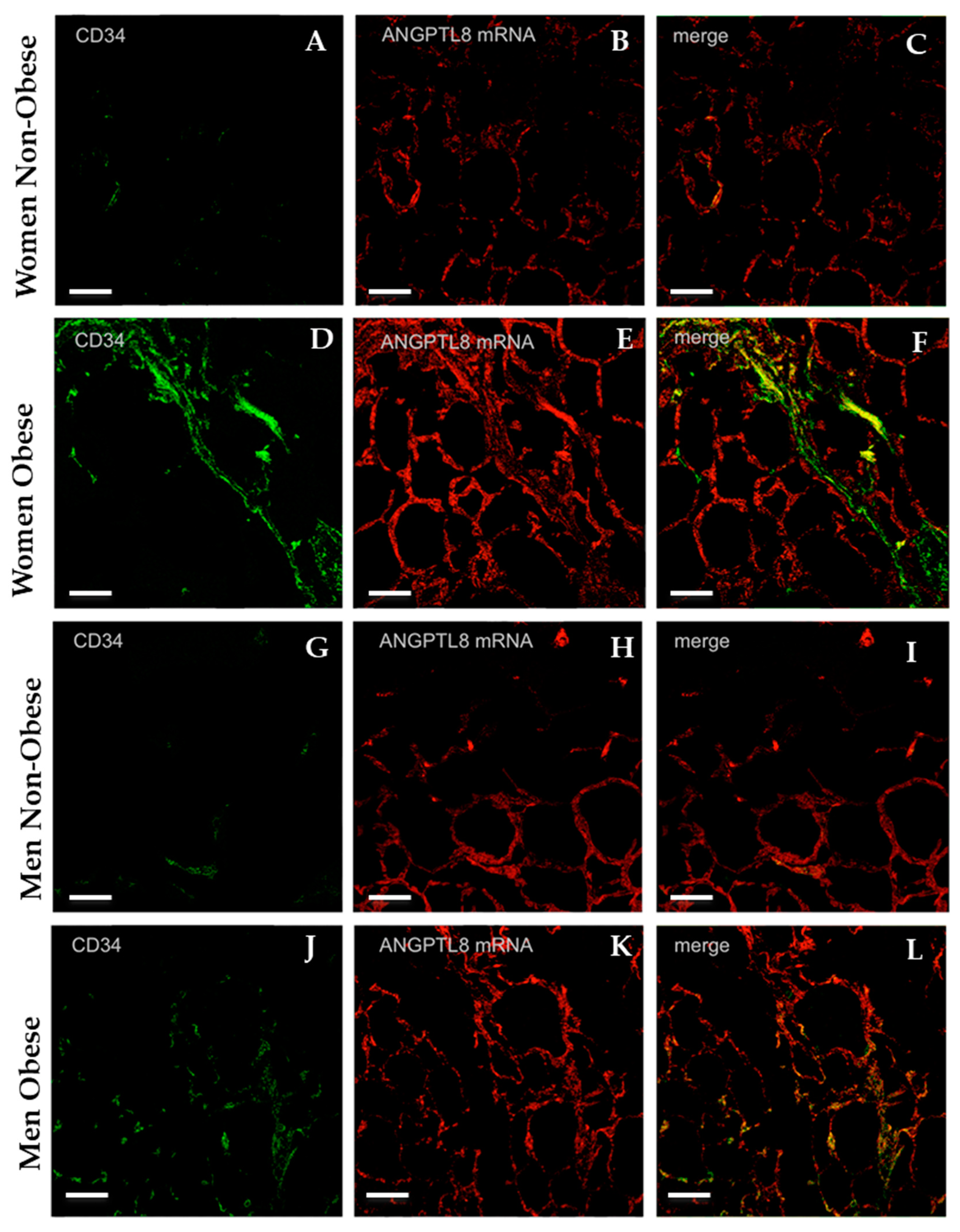
3.4. Gene Expression and Localisation of ANGPTL8 in Visceral Adipose Tissue of Non-Obese and of Obese Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hossain:, P.; Kawar, B.; El Nahas, M. Obesity and diabetes in the developing world—A growing challenge. N. Engl. J. Med. 2007, 356, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, I. Recent advances in physiological lipoprotein metabolism. Clin. Chem. Lab. Med. 2014, 52, 1695–1727. [Google Scholar] [CrossRef] [PubMed]
- Santamarina-Fojo, S.; Brewer, H.B., Jr. The familial hyperchylomicronemia syndrome. New insights into underlying genetic defects. JAMA 1991, 265, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Coleman, T.; Seip, R.L.; Gimble, J.M.; Lee, D.; Maeda, N.; Semenkovich, C.F. Cooh-terminal disruption of lipoprotein lipase in mice is lethal in homozygotes, but heterozygotes have elevated triglycerides and impaired enzyme activity. J. Biol. Chem. 1995, 270, 12518–12525. [Google Scholar] [CrossRef] [PubMed]
- Shimada, M.; Shimano, H.; Gotoda, T.; Yamamoto, K.; Kawamura, M.; Inaba, T.; Yazaki, Y.; Yamada, N. Overexpression of human lipoprotein lipase in transgenic mice. Resistance to diet-induced hypertriglyceridemia and hypercholesterolemia. J. Biol. Chem. 1993, 268, 17924–17929. [Google Scholar] [PubMed]
- Janssen, A.W.F.; Katiraei, S.; Bartosinska, B.; Eberhard, D.; Willems van Dijk, K.; Kersten, S. Loss of angiopoietin-like 4 (ANGPTL4) in mice with diet-induced obesity uncouples visceral obesity from glucose intolerance partly via the gut microbiota. Diabetologia 2018, 61, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Kersten, S. Physiological regulation of lipoprotein lipase. Biochim. Biophys. Acta 2014, 1841, 919–933. [Google Scholar] [CrossRef]
- Dijk, W.; Kersten, S. Regulation of lipoprotein lipase by ANGPTL4. Trends Endocrinol. Metab. TEM 2014, 25, 146–155. [Google Scholar] [CrossRef]
- Ren, G.; Kim, J.Y.; Smas, C.M. Identification of rifl, a novel adipocyte-enriched insulin target gene with a role in lipid metabolism. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E334–E351. [Google Scholar] [CrossRef]
- Zhang, R. Lipasin, a novel nutritionally-regulated liver-enriched factor that regulates serum triglyceride levels. Biochem. Biophys. Res. Commun. 2012, 424, 786–792. [Google Scholar] [CrossRef]
- Gray, K.A.; Yates, B.; Seal, R.L.; Wright, M.W.; Bruford, E.A. Genenames.org: The hgnc resources in 2015. Nucleic Acids Res. 2015, 43, D1079–D1085. [Google Scholar] [CrossRef] [PubMed]
- Yi, P.; Park, J.S.; Melton, D.A. Betatrophin: A hormone that controls pancreatic beta cell proliferation. Cell 2013, 153, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Yao, F.; Abou-Samra, A.B.; Zhang, R. Lipasin, thermoregulated in brown fat, is a novel but atypical member of the angiopoietin-like protein family. Biochem. Biophys. Res. Commun. 2013, 430, 1126–1131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Abou-Samra, A.B. Emerging roles of lipasin as a critical lipid regulator. Biochem. Biophys. Res. Commun. 2013, 432, 401–405. [Google Scholar] [CrossRef]
- Yi, P.; Park, J.S.; Melton, D.A. Retraction notice to: Betatrophin: A hormone that controls pancreatic beta cell proliferation. Cell 2017, 168, 326. [Google Scholar] [CrossRef]
- Cox, A.R.; Barrandon, O.; Cai, E.P.; Rios, J.S.; Chavez, J.; Bonnyman, C.W.; Lam, C.J.; Yi, P.; Melton, D.A.; Kushner, J.A. Resolving discrepant findings on ANGPTL8 in beta-cell proliferation: A collaborative approach to resolving the betatrophin controversy. PLoS ONE 2016, 11, e0159276. [Google Scholar] [CrossRef]
- Gusarova, V.; Alexa, C.A.; Na, E.; Stevis, P.E.; Xin, Y.; Bonner-Weir, S.; Cohen, J.C.; Hobbs, H.H.; Murphy, A.J.; Yancopoulos, G.D.; et al. ANGPTL8/betatrophin does not control pancreatic beta cell expansion. Cell 2014, 159, 691–696. [Google Scholar] [CrossRef]
- Abu-Farha, M.; Abubaker, J.; Al-Khairi, I.; Cherian, P.; Noronha, F.; Hu, F.B.; Behbehani, K.; Elkum, N. Higher plasma betatrophin/ANGPTL8 level in type 2 diabetes subjects does not correlate with blood glucose or insulin resistance. Sci. Rep. 2015, 5, 10949. [Google Scholar] [CrossRef]
- Espes, D.; Lau, J.; Carlsson, P.O. Increased circulating levels of betatrophin in individuals with long-standing type 1 diabetes. Diabetologia 2014, 57, 50–53. [Google Scholar] [CrossRef]
- Espes, D.; Martinell, M.; Carlsson, P.O. Increased circulating betatrophin concentrations in patients with type 2 diabetes. Int. J. Endocrinol. 2014, 2014, 323407. [Google Scholar] [CrossRef]
- Wang, L.; Song, J.; Wang, C.; Lin, P.; Liang, K.; Sun, Y.; He, T.; Li, W.; Zhao, R.; Qin, J.; et al. Circulating levels of betatrophin and irisin are not associated with pancreatic beta-cell function in previously diagnosed type 2 diabetes mellitus patients. J. Diabetes Res. 2016, 2016, 2616539. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Saito, T.; Aoki, A.; Asano, T.; Yoshida, M.; Ikoma, A.; Kusaka, I.; Toyoshima, H.; Kakei, M.; Ishikawa, S.E. Circulating betatrophin is elevated in patients with type 1 and type 2 diabetes. Endocr. J. 2015, 62, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Fenzl, A.; Itariu, B.K.; Kosi, L.; Fritzer-Szekeres, M.; Kautzky-Willer, A.; Stulnig, T.M.; Kiefer, F.W. Circulating betatrophin correlates with atherogenic lipid profiles but not with glucose and insulin levels in insulin-resistant individuals. Diabetologia 2014, 57, 1204–1208. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Ambrosi, J.; Pascual, E.; Catalan, V.; Rodriguez, A.; Ramirez, B.; Silva, C.; Gil, M.J.; Salvador, J.; Fruhbeck, G. Circulating betatrophin concentrations are decreased in human obesity and type 2 diabetes. J. Clin. Endocrinol. Metab. 2014, 99, E2004–E2009. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Yu, H.; Lu, J.; Bao, Y.; Chen, H.; Jia, W. Decreased serum betatrophin levels correlate with improved fasting plasma glucose and insulin secretion capacity after roux-en-y gastric bypass in obese chinese patients with type 2 diabetes: A 1-year follow-up. Surg. Obes. Relat. Dis. 2016, 12, 1343–1348. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, P.; He, W.; Zhang, J.; Liu, L.; Yang, Y.; Liu, Z.; Xie, J.; Shao, S.; Du, T.; et al. Circulating betatrophin levels are increased in patients with type 2 diabetes and associated with insulin resistance. J. Clin. Endocrinol. Metab. 2015, 100, E96–E100. [Google Scholar] [CrossRef]
- Guo, X.R.; Wang, X.L.; Chen, Y.; Yuan, Y.H.; Chen, Y.M.; Ding, Y.; Fang, J.; Bian, L.J.; Li, D.S. ANGPTL8/betatrophin alleviates insulin resistance via the Akt-GSK3beta or Akt-FoxO1 pathway in HepG2 cells. Exp. Cell Res. 2016, 345, 158–167. [Google Scholar]
- Ebert, T.; Kralisch, S.; Wurst, U.; Lossner, U.; Kratzsch, J.; Bluher, M.; Stumvoll, M.; Tonjes, A.; Fasshauer, M. Betatrophin levels are increased in women with gestational diabetes mellitus compared to healthy pregnant controls. Eur. J. Endocrinol. 2015, 173, 1–7. [Google Scholar] [CrossRef]
- Liu, D.; Qu, H.; Wang, H.; Duan, Y.; Liu, F.; Feng, Z.; Deng, H. Relationship between serum betatrophin levels and the first-phase of glucose-stimulated insulin secretion. Obes. Res. Clin. Pract. 2018, 12, 9–15. [Google Scholar] [CrossRef]
- Luo, M.; Peng, D. ANGPTL8: An important regulator in metabolic disorders. Front. Endocrinol. 2018, 9, 169. [Google Scholar] [CrossRef]
- Gusarova, V.; Banfi, S.; Alexa-Braun, C.A.; Shihanian, L.M.; Mintah, I.J.; Lee, J.S.; Xin, Y.; Su, Q.; Kamat, V.; Cohen, J.C.; et al. ANGPTL8 blockade with a monoclonal antibody promotes triglyceride clearance, energy expenditure, and weight loss in mice. Endocrinology 2017, 158, 1252–1259. [Google Scholar] [CrossRef] [PubMed]
- Izumi, R.; Kusakabe, T.; Noguchi, M.; Iwakura, H.; Tanaka, T.; Miyazawa, T.; Aotani, D.; Hosoda, K.; Kangawa, K.; Nakao, K. CRISPR/Cas9-mediated ANGPTL8 knockout suppresses plasma triglyceride concentrations and adiposity in rats. J. Lipid Res. 2018, 59, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Quagliarini, F.; Gusarova, V.; Gromada, J.; Valenzuela, D.M.; Cohen, J.C.; Hobbs, H.H. Mice lacking ANGPTL8 (betatrophin) manifest disrupted triglyceride metabolism without impaired glucose homeostasis. Proc. Natl. Acad. Sci. USA 2013, 110, 16109–16114. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Ambrosi, J.; Pascual-Corrales, E.; Catalan, V.; Rodriguez, A.; Ramirez, B.; Romero, S.; Vila, N.; Ibanez, P.; Margall, M.A.; Silva, C.; et al. Altered concentrations in dyslipidemia evidence a role for ANGPTL8/betatrophin in lipid metabolism in humans. J. Clin. Endocrinol. Metab. 2016, 101, 3803–3811. [Google Scholar] [CrossRef]
- Zhang, R. The ANGPTL3-4-8 model, a molecular mechanism for triglyceride trafficking. Open Biol. 2016, 6, 150272. [Google Scholar] [CrossRef]
- Zhang, R.; Abou-Samra, A.B. A dual role of lipasin (betatrophin) in lipid metabolism and glucose homeostasis: Consensus and controversy. Cardiovasc. Diabetol. 2014, 13, 133. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped blast and psi-blast: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Nidhina Haridas, P.A.; Soronen, J.; Sadevirta, S.; Mysore, R.; Quagliarini, F.; Pasternack, A.; Metso, J.; Perttila, J.; Leivonen, M.; Smas, C.M.; et al. Regulation of angiopoietin-like proteins (ANGPTLs) 3 and 8 by insulin. J. Clin. Endocrinol. Metab. 2015, 100, E1299–E1307. [Google Scholar] [CrossRef]
- Von Loeffelholz, C.; Pfeiffer, A.F.H.; Lock, J.F.; Lieske, S.; Docke, S.; Murahovschi, V.; Kriebel, J.; de Las Heras Gala, T.; Grallert, H.; Rudovich, N.; et al. ANGPTL8 (betatrophin) is expressed in visceral adipose tissue and relates to human hepatic steatosis in two independent clinical collectives. Horm. Metab. Res. 2017, 49, 343–349. [Google Scholar] [CrossRef][Green Version]
- Quagliarini, F.; Wang, Y.; Kozlitina, J.; Grishin, N.V.; Hyde, R.; Boerwinkle, E.; Valenzuela, D.M.; Murphy, A.J.; Cohen, J.C.; Hobbs, H.H. Atypical angiopoietin-like protein that regulates ANGPTL3. Proc. Natl. Acad. Sci. USA 2012, 109, 19751–19756. [Google Scholar] [CrossRef] [PubMed]
- Vatner, D.F.; Goedeke, L.; Camporez, J.G.; Lyu, K.; Nasiri, A.R.; Zhang, D.; Bhanot, S.; Murray, S.F.; Still, C.D.; Gerhard, G.S.; et al. ANGPTL8 antisense oligonucleotide improves adipose lipid metabolism and prevents diet-induced nafld and hepatic insulin resistance in rodents. Diabetologia 2018, 61, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Abou-Samra, A.B.; Zhang, R. An explanation for recent discrepancies in levels of human circulating betatrophin. Diabetologia 2014, 57, 2232–2234. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, D.; Li, L.; Li, Y.; Li, Q.; An, Z.; Sun, X.; Tian, H. Circulating betatrophin in patients with type 2 diabetes: A meta-analysis. J. Diabetes Res. 2016, 2016, 6194750. [Google Scholar] [CrossRef]
- Fu, Z.; Berhane, F.; Fite, A.; Seyoum, B.; Abou-Samra, A.B.; Zhang, R. Elevated circulating lipasin/betatrophin in human type 2 diabetes and obesity. Sci. Rep. 2014, 4, 5013. [Google Scholar] [CrossRef] [PubMed]
- Abu-Farha, M.; Sriraman, D.; Cherian, P.; AlKhairi, I.; Elkum, N.; Behbehani, K.; Abubaker, J. Circulating ANGPTL8/betatrophin is increased in obesity and reduced after exercise training. PLoS ONE 2016, 11, e0147367. [Google Scholar] [CrossRef]
- Fu, Z.; Abou-Samra, A.B.; Zhang, R. A lipasin/ANGPTL8 monoclonal antibody lowers mouse serum triglycerides involving increased postprandial activity of the cardiac lipoprotein lipase. Sci. Rep. 2015, 5, 18502. [Google Scholar] [CrossRef]
- Ejarque, M.; Borlaug, M.; Vilarrasa, N.; Martinez-Perez, B.; Llaurado, G.; Megia, A.; Helland, T.; Gutierrez, C.; Serena, C.; Folkestad, O.; et al. Angiopoietin-like protein 8/betatrophin as a new determinant of type 2 diabetes remission after bariatric surgery. Transl. Res. J. Lab. Clin. Med. 2017, 184, 35–44. [Google Scholar] [CrossRef]
- Wang, H.; Lai, Y.; Han, C.; Liu, A.; Fan, C.; Wang, H.; Zhang, H.; Ding, S.; Teng, W.; Shan, Z. The effects of serum ANGPTL8/betatrophin on the risk of developing the metabolic syndrome—A prospective study. Sci. Rep. 2016, 6, 28431. [Google Scholar] [CrossRef]
- Xie, X.; Gao, T.; Yang, M.; Chen, P.; Jin, H.; Yang, L.; Yu, X. Associations of betatrophin levels with irisin in chinese women with normal glucose tolerance. Diabetol. Metab. Syndr. 2015, 7, 26. [Google Scholar] [CrossRef]
- Wu, S.; Gao, H.; Ma, Y.; Fu, L.; Zhang, C.; Luo, X. Characterisation of betatrophin concentrations in childhood and adolescent obesity and insulin resistance. Pediatric Diabetes 2016, 17, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Christen, T.; Trompet, S.; Noordam, R.; van Klinken, J.B.; van Dijk, K.W.; Lamb, H.J.; Cobbaert, C.M.; den Heijer, M.; Jazet, I.M.; Jukema, J.W.; et al. Sex differences in body fat distribution are related to sex differences in serum leptin and adiponectin. Peptides 2018, 107, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Corrales, E.; Gomez-Ambrosi, J.; Moncada, R.; Valenti, V.; Catalan, V.; Rodriguez, A.; Ramirez, B.; Silva, C.; Gil, M.J.; Salvador, J.; et al. Circulating ANGPTL8/betatrophin concentrations are increased after surgically induced weight loss, but not after diet-induced weight loss. Obes. Surg. 2016, 26, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- Roth, C.L.; Elfers, C.; Lass, N.; Reinehr, T. Betatrophin: No relation to glucose metabolism or weight status in obese children before and after lifestyle intervention. Pediatric Diabetes 2017, 18, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Chen, R.P.; Yang, R.; Guo, X.F.; Zhang, J.C.; Chen, H. Betatrophin acts as a diagnostic biomarker in type 2 diabetes mellitus and is negatively associated with hdl-cholesterol. Int. J. Endocrinol. 2015, 2015, 479157. [Google Scholar] [CrossRef] [PubMed]
- Tuhan, H.; Abaci, A.; Anik, A.; Catli, G.; Kume, T.; Calan, O.G.; Acar, S.; Bober, E. Circulating betatrophin concentration is negatively correlated with insulin resistance in obese children and adolescents. Diabetes Res. Clin. Pract. 2016, 114, 37–42. [Google Scholar] [CrossRef]
- Davin, J.C. The glomerular permeability factors in idiopathic nephrotic syndrome. Pediatric Nephrol. 2016, 31, 207–215. [Google Scholar] [CrossRef]
- Guo, K.; Lu, J.; Yu, H.; Zhao, F.; Pan, P.; Zhang, L.; Chen, H.; Bao, Y.; Jia, W. Serum betatrophin concentrations are significantly increased in overweight but not in obese or type 2 diabetic individuals. Obesity 2015, 23, 793–797. [Google Scholar] [CrossRef]
- Hu, H.; Sun, W.; Yu, S.; Hong, X.; Qian, W.; Tang, B.; Wang, D.; Yang, L.; Wang, J.; Mao, C.; et al. Increased circulating levels of betatrophin in newly diagnosed type 2 diabetic patients. Diabetes Care 2014, 37, 2718–2722. [Google Scholar] [CrossRef]
- Lee, S.H.; Rhee, M.; Kwon, H.S.; Park, Y.M.; Yoon, K.H. Serum betatrophin concentrations and the risk of incident diabetes: A nested case-control study from chungju metabolic disease cohort. Diabetes Metab. J. 2018, 42, 53–62. [Google Scholar] [CrossRef]
- Verboven, K.; Wouters, K.; Gaens, K.; Hansen, D.; Bijnen, M.; Wetzels, S.; Stehouwer, C.D.; Goossens, G.H.; Schalkwijk, C.G.; Blaak, E.E.; et al. Abdominal subcutaneous and visceral adipocyte size, lipolysis and inflammation relate to insulin resistance in male obese humans. Sci. Rep. 2018, 8, 4677. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Britt, E.C.; Shows, H.W.; Hjelmaas, A.J.; Shetty, S.K.; Cushing, E.M.; Li, W.; Dou, A.; Zhang, R.; Davies, B.S.J. ANGPTL8 promotes the ability of ANGPTL3 to bind and inhibit lipoprotein lipase. Mol. Metab. 2017, 6, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.H.; Ke, P.Y.; Liao, C.J.; Wu, S.M.; Chi, H.C.; Tsai, C.Y.; Chen, C.Y.; Lin, Y.H.; Lin, K.H. Chromosome 19 open reading frame 80 is upregulated by thyroid hormone and modulates autophagy and lipid metabolism. Autophagy 2014, 10, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.; Moon, S.O.; Koh, K.N.; Kim, H.; Uhm, C.S.; Kwak, H.J.; Kim, N.G.; Koh, G.Y. Molecular cloning, expression, and characterization of angiopoietin-related protein. Angiopoietin-related protein induces endothelial cell sprouting. J. Biol. Chem. 1999, 274, 26523–26528. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.S.; Beigneux, A.P.; Barnes, R.H., 2nd; Tu, Y.; Gin, P.; Weinstein, M.M.; Nobumori, C.; Nyren, R.; Goldberg, I.; Olivecrona, G.; et al. Gpihbp1 is responsible for the entry of lipoprotein lipase into capillaries. Cell Metab. 2010, 12, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Haller, J.F.; Mintah, I.J.; Shihanian, L.M.; Stevis, P.; Buckler, D.; Alexa-Braun, C.A.; Kleiner, S.; Banfi, S.; Cohen, J.C.; Hobbs, H.H.; et al. ANGPTL8 requires ANGPTL3 to inhibit lipoprotein lipase and plasma triglyceride clearance. J. Lipid Res. 2017, 58, 1166–1173. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, F.; Cui, J.; Shi, Y.; Xiang, J.; Wang, X.; Zhao, N.; Yan, Q.; Greenberg, A.S.; Peng, Y.; et al. Association of betatrophin with metabolic characteristics in overweight/obese and lean women with pcos. Gynecol. Endocrinol. 2017, 33, 238–243. [Google Scholar] [CrossRef]
- Lu, X. Structure and function of angiopoietin-like protein 3 (ANGPTL3) in atherosclerosis. Curr. Med. Chem. 2019. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Women | Men | ||||||
|---|---|---|---|---|---|---|---|
| Non-obese | Obese | Non-obese | Obese | P 2-way-ANOVA | |||
| Patients | n = 22 | n = 44 | n = 20 | n = 23 | Obesity | Gender | Interaction |
| Age (years) | 58 ± 14 | 48 ± 2 | 58 ± 8 | 47 ± 3 | 0.462 | 0.957 | 0.945 |
| BMI (kg/m2) | 21.7 ± 1.9 | 45.6 ± 1.3 | 27.3 ± 1.07 | 47.2 ± 3.0 | <0.001 | 0.092 | 0.346 |
| Waist circumference (cm) | 88.8 ± 9.1 | 131.4 ± 2.7 | 77.5 ± 29.5 | 137.0 ± 6.2 | <0.001 | 0.800 | 0.453 |
| Lipid drop area (µm2) | 3958.6 ± 85.2 | 4458.1 ± 80.2 | 3984.9 ± 93 | 4376.1 ± 100.9 | <0.005 | 0.805 | 0.635 |
| Women | Men | ||||||
|---|---|---|---|---|---|---|---|
| Non-obese | Obese | Non-obese | Obese | P 2-way-ANOVA | |||
| Patients | n = 22 | n = 44 | n = 20 | n = 23 | Obesity | Gender | Inter- Action |
| Glucose (mg/dL) | 94.0 ± 8.7 | 101.6 ± 2.1 | 97.8 ± 4.7 | 98.3 ± 8.7 | 0.105 | 0.660 | 0.836 |
| Insulin (μU/mL) | 8.6 ± 1.4 | 18.2 ± 0.7 | 5.9 ± 1.7 | 24.7 ± 5.6 | <0.001 | 0.456 | 0.063 |
| HbA1C (%) | 5.2 ± 0.1 | 10.6 ± 2.9 | 5.8 ± 0.7 | 11.6 ± 4.7 | 0.053 | 0.549 | 0.418 |
| C-Peptide (ng/mL) | 2.0± 0.4 | 4.3 ± 0.2 | 2.2 ± 0.4 | 5.8 ± 0.8 | <0.001 | 0.286 | 0.459 |
| HOMA β (%) | 94.2 ± 16.1 | 134.2 ± 9.1 | 69.3 ± 10.2 | 175.4 ± 20.7 | <0.001 | 0.590 | 0.031 |
| HOMA-S (%) | 88.7 ±12.3 | 41.8 ± 5.3 | 125.5 ± 11.6 | 32.4 ± 7.7 | <0.001 | 0.133 | 0.012 |
| HOMA-IR (AU) | 1.1 ± 0.3 | 2.4 ± 0.2 | 0.8 ± 0.1 | 3.1 ± 0.6 | <0.001 | 0.578 | 0.166 |
| Triglycerides (mg/dL) | 77.6 ± 16.8 | 129.0 ± 34.4 | 108.8 ± 8.4 | 163 ± 17.5 | 0.212 | 0.449 | 0.966 |
| Cholesterol (mg/dL) | 123.0 ± 13.6 | 154.7 ± 6.7 | 162.5 ± 31.0 | 158.4 ± 11.3 | 0.601 | 0.311 | 0.442 |
| LDL-C (mg/dL) | 63.1 ± 8.3 | 86.0 ± 4.4 | 94.3 ± 19.6 | 95 ± 7.6 | 0.526 | 0.526 | 0.54 |
| HDL-C (mg/dL) | 44.4 ± 12.8 | 42.8 ± 2.2 | 55.7 ± 12.2 | 34.6 ± 3.7 | 0.092 | 0.816 | 0.147 |
| Leptin (ng/mL) | 19.9 ± 8 | 71.7 ± 7.1 | 6.8 ± 0.2 | 28.1 ± 6.8 | <0.005 | 0.062 | 0.134 |
| ANGPTL8 | ||||||||
|---|---|---|---|---|---|---|---|---|
| Women | Men | |||||||
| Non-obese n = 22 | Obese n = 44 | Non-obese n = 20 | Obese n = 23 | |||||
| Variable | ρ | p | ρ | p | ρ | p | ρ | p |
| Age (years) | −0.3714 | n.s | −0.2814 | n.s | 0.5429 | n.s | −0,4762 | n.s |
| BMI (kg/m2) | 0.2000 | n.s. | 0.0625 | n.s. | −0.8000 | n.s. | 0.5429 | n.s. |
| Waist circumference (cm) | −0.3714 | n.s. | −0.0760 | n.s. | −0.8000 | n.s. | 0.1429 | n.s. |
| Lipid drop area (µm2) | 0.2723 | n.s. | 0.1345 | n.s. | 0.1429 | n.s. | 0.0952 | n.s. |
| Glucose (mg/dL) | 0.5429 | n.s. | −0.2731 | n.s. | 0.0857 | n.s. | 0.1778 | n.s. |
| Insulin (μU/mL) | 0.5429 | n.s. | −0.1313 | n.s. | −0.1429 | n.s. | −0.7857 | p < 0.05 |
| HbA1C (%) | −0.5000 | n.s. | 0.2284 | n.s. | −0.7000 | n.s. | −0.1627 | n.s. |
| C-Peptide (ng/mL) | −0.9000 | n.s. | −0.0315 | n.s. | 0.2855 | n.s. | 0.2571 | n.s. |
| HOMA β (%) | 0.0000 | n.s. | 0.0284 | n.s. | −0.1429 | n.s. | 0.3929 | n.s. |
| HOMA-S (%) | −0.3000 | n.s. | 0.2021 | n.s. | 0.2000 | n.s. | 0.7857 | p < 0.05 |
| HOMA-IR (AU) | 0.3000 | n.s. | −0.2021 | n.s. | −0.2000 | n.s. | −0.7857 | p < 0.05 |
| Triglycerides (mg/dL) | 0.2000 | n.s. | −0.0802 | n.s. | −0.4857 | n.s. | −0.5357 | n.s. |
| Cholesterol (mg/dL) | −0.0857 | n.s. | 0.0385 | n.s. | −0.3000 | n.s. | −0.1786 | n.s. |
| LDL-C (mg/dL) | 0.05798 | n.s. | 0.0946 | n.s. | −0.8000 | n.s. | −0.5946 | n.s. |
| HDL-C (mg/dL) | 0.3143 | n.s. | −0.2888 | n.s. | 0.4000 | n.s. | 0.8895 | p < 0.05 |
| Leptin (ng/mL) | 0.7714 | n.s. | 0.0865 | n.s. | −0.1000 | n.s. | 0.5000 | n.s. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catalano-Iniesta, L.; Sánchez Robledo, V.; Iglesias-Osma, M.C.; Galán Albiñana, A.; Carrero, S.; Blanco, E.J.; Carretero-Hernández, M.; Carretero, J.; García-Barrado, M.J. Evidences for Expression and Location of ANGPTL8 in Human Adipose Tissue. J. Clin. Med. 2020, 9, 512. https://doi.org/10.3390/jcm9020512
Catalano-Iniesta L, Sánchez Robledo V, Iglesias-Osma MC, Galán Albiñana A, Carrero S, Blanco EJ, Carretero-Hernández M, Carretero J, García-Barrado MJ. Evidences for Expression and Location of ANGPTL8 in Human Adipose Tissue. Journal of Clinical Medicine. 2020; 9(2):512. https://doi.org/10.3390/jcm9020512
Chicago/Turabian StyleCatalano-Iniesta, Leonardo, Virginia Sánchez Robledo, María Carmen Iglesias-Osma, Amparo Galán Albiñana, Sixto Carrero, Enrique J. Blanco, Marta Carretero-Hernández, José Carretero, and María José García-Barrado. 2020. "Evidences for Expression and Location of ANGPTL8 in Human Adipose Tissue" Journal of Clinical Medicine 9, no. 2: 512. https://doi.org/10.3390/jcm9020512
APA StyleCatalano-Iniesta, L., Sánchez Robledo, V., Iglesias-Osma, M. C., Galán Albiñana, A., Carrero, S., Blanco, E. J., Carretero-Hernández, M., Carretero, J., & García-Barrado, M. J. (2020). Evidences for Expression and Location of ANGPTL8 in Human Adipose Tissue. Journal of Clinical Medicine, 9(2), 512. https://doi.org/10.3390/jcm9020512